
Vermicomposting—Facts, Benefits and Knowledge Gaps


 ,
, 
{kind=link}
{kind=link}
Abstract
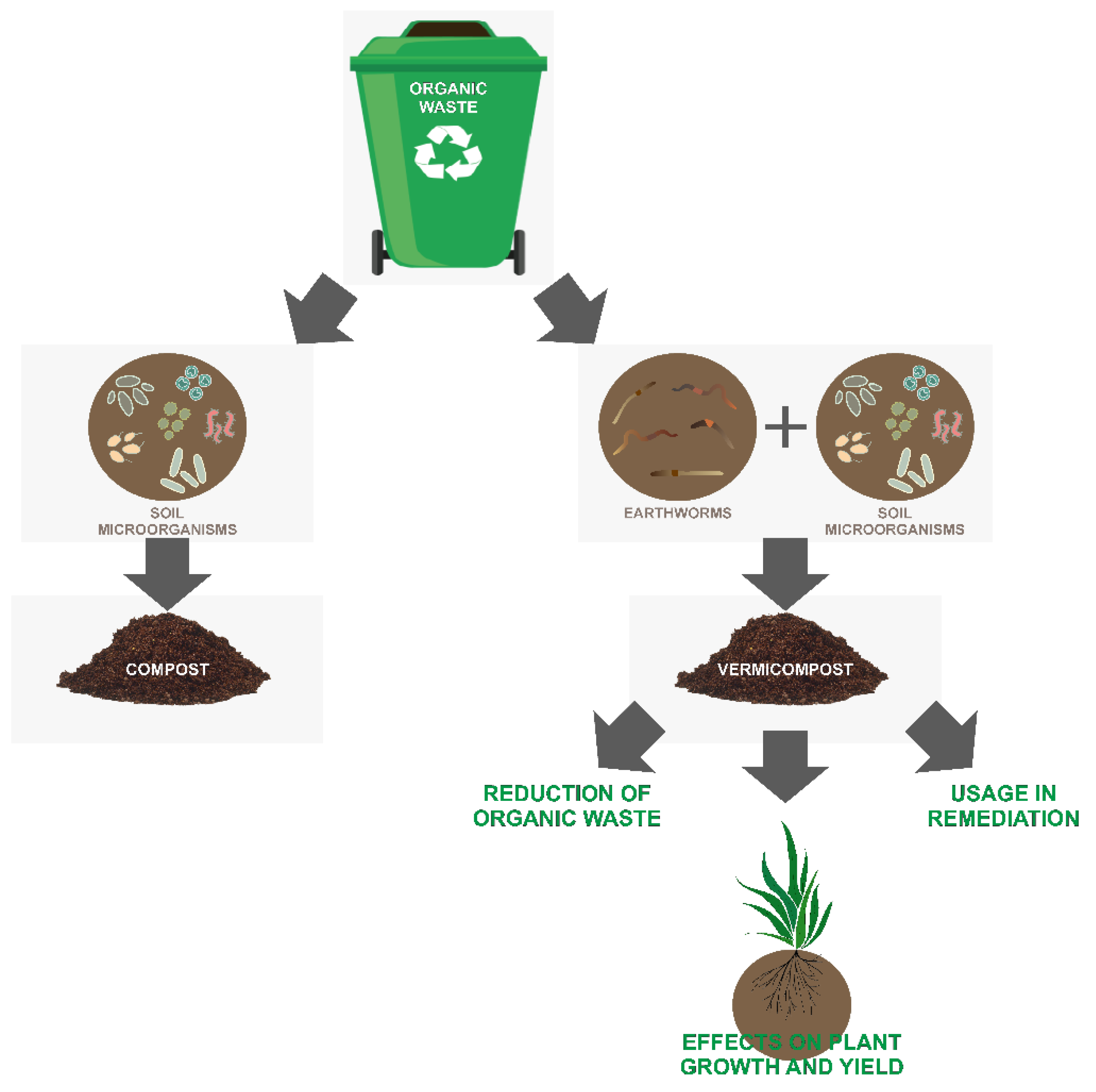
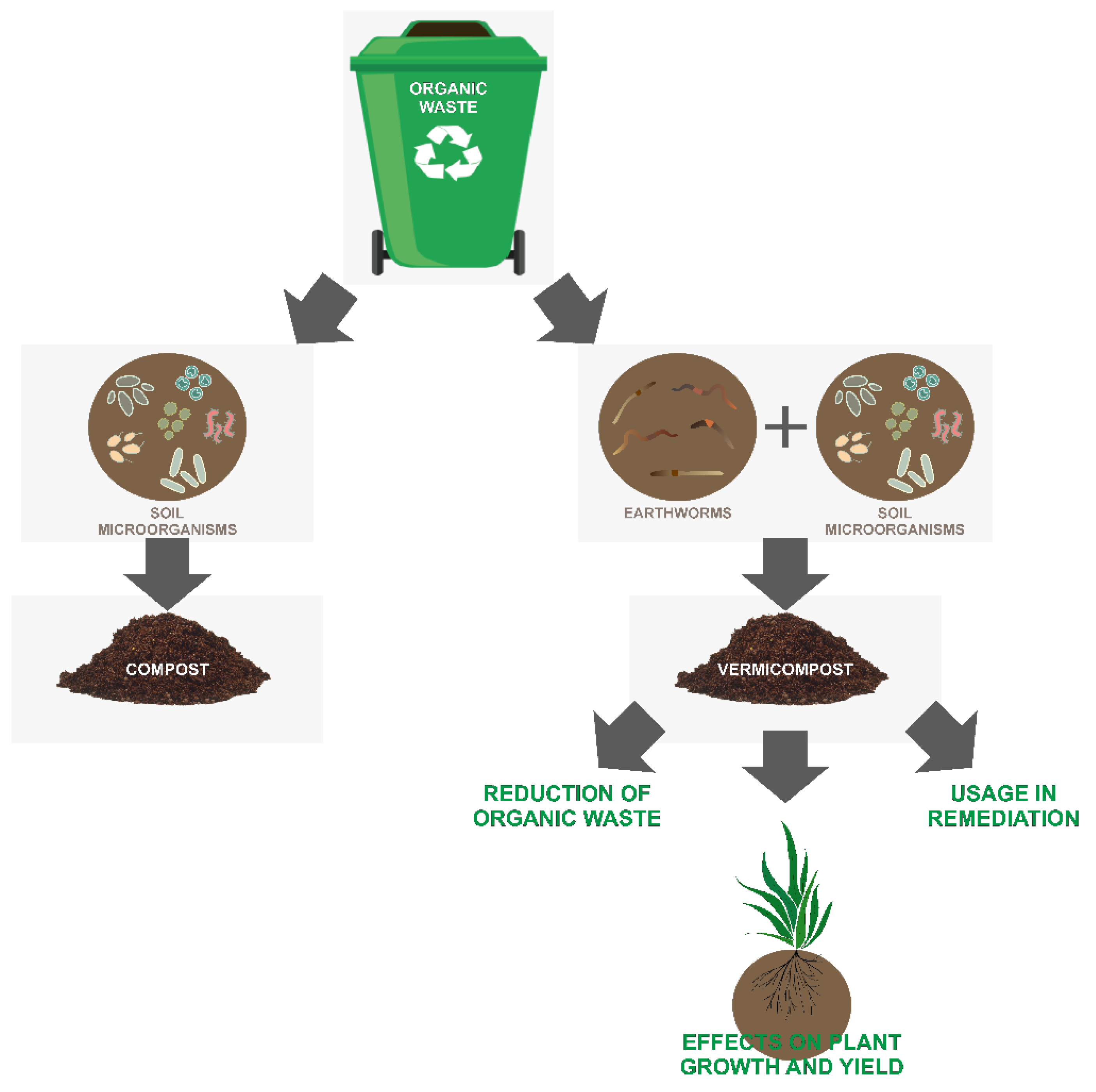
:1. Introduction
2. Bacterial Community Structure in Vermicompost
3. Effect of Vermicompost on Plant Growth and Yield
4. The Potential Use of Vermicompost in Remediation
5. Disease and Pest Control by Vermicompost
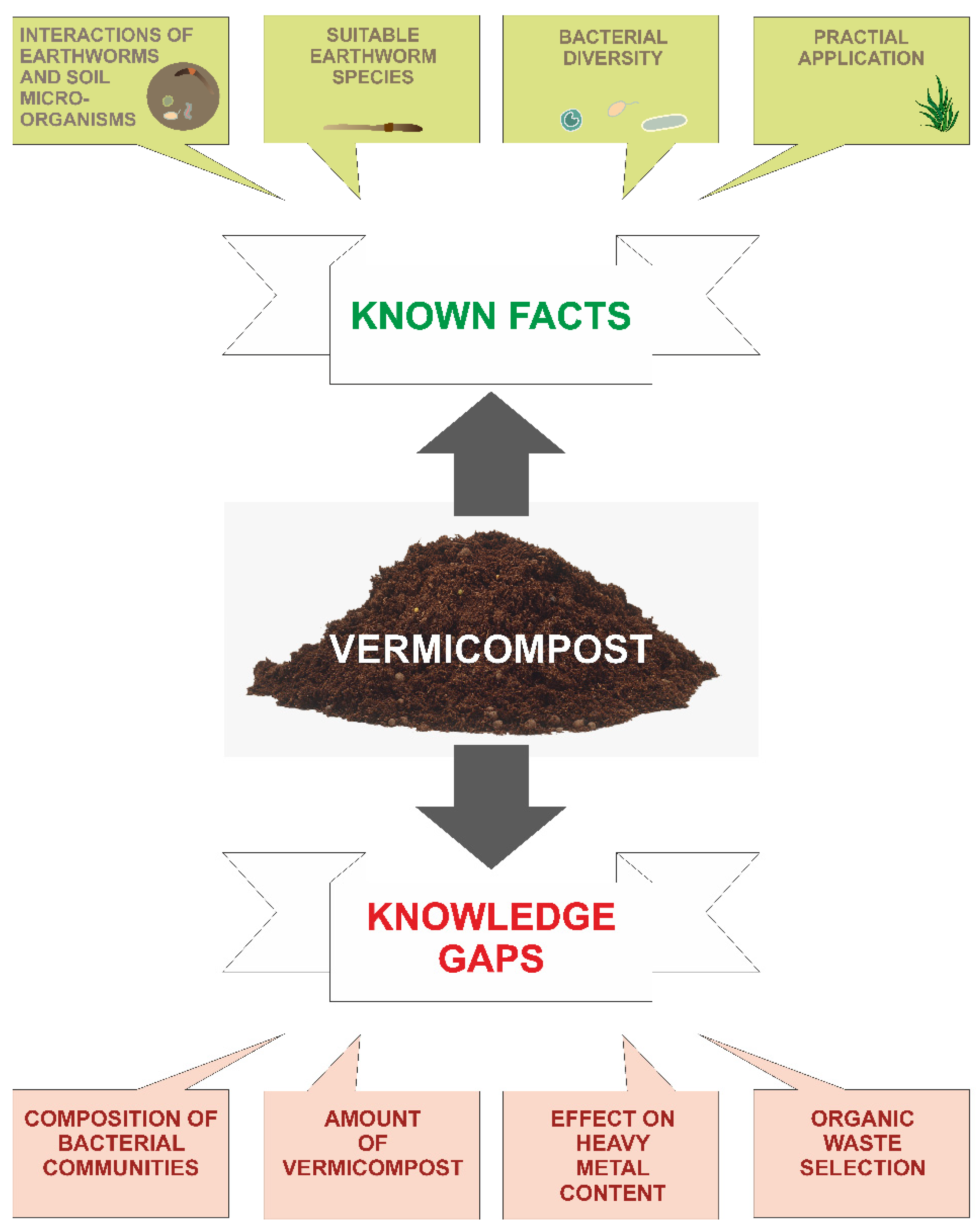
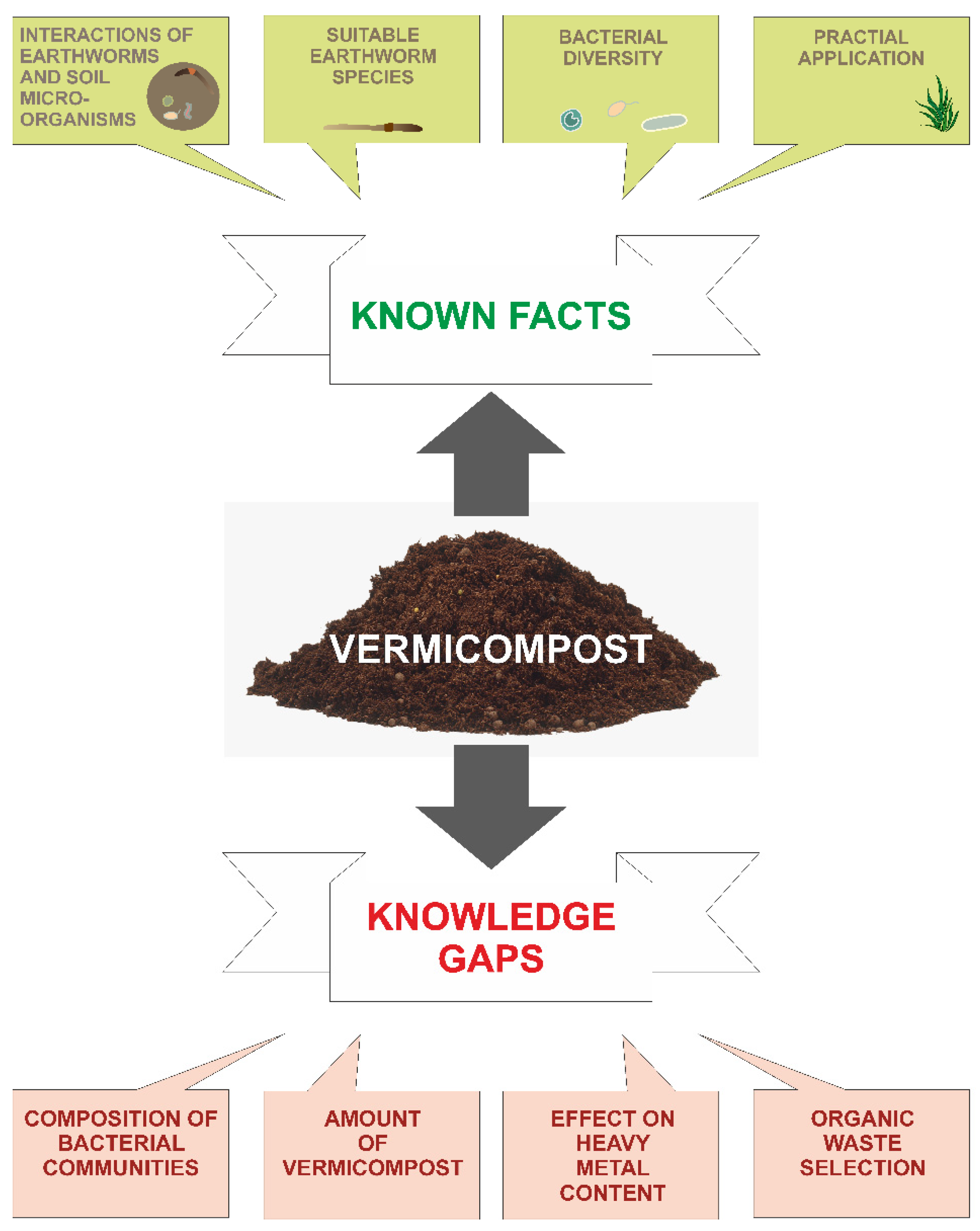
6. Knowledge Gaps
6.1. Composition of Bacterial Communities
6.2. Amount of Vermicompost
6.3. Effect on Heavy Metal Content
6.4. Organic Waste Selection
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Demirbas, A. Waste management, waste resource facilities and waste conversion processes. Energy Convers. Manag. 2011, 52, 1280–1287. [Google Scholar] [CrossRef]
- Sabbas, T.; Polettini, A.; Pomi, R.; Astrup, T.; Hjelmar, O.; Mostbauer, P.; Cappai, G.; Magel, G.; Salhofer, S.; Speiser, C.; et al. Management of municipal solid waste incineration residues. Waste Manag. 2003, 23, 61–88. [Google Scholar] [CrossRef]
- Hoornweg, D.; Bhata-Tata, P. What a Waste: A Global Review of Solid Waste Menagement; World Bank: Washington, DC, USA, 2012; ISBN 9781409406877. [Google Scholar]
- Suthar, S.; Singh, S. Vermicomposting of domestic waste by using two epigeic earthworms. J. Environ. Sci. Technol. 2008, 5, 99–106. [Google Scholar]
- Kaviraj; Sharma, S. Municipal solid waste management through vermicomposting employing exotic and local species of earthworms. Bioresour. Technol. 2003, 90, 169–173. [Google Scholar] [CrossRef]
- Dumontet, S.; Dinel, H.; Baloda, S.B. Pathogen Reduction in Sewage Sludge by Composting and Other Biological Treatments: A Review. Biol. Agric. Hortic. 1999, 16, 409–430. [Google Scholar] [CrossRef]
- Usmani, Z.; Kumar, V.; Gupta, P.; Gupta, G.; Rani, R.; Chandra, A. Enhanced soil fertility, plant growth promotion and microbial enzymatic activities of vermicomposted fly ash. Sci. Rep. 2019, 9, 10455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arancon, N.Q.; Edwards, C.A.; Babenko, A.; Cannon, J.; Galvis, P.; Metzger, J.D. Influences of vermicomposts, produced by earthworms and microorganisms from cattle manure, food waste and paper waste, on the germination, growth and flowering of petunias in the greenhouse. Appl. Soil Ecol. 2008, 39, 91–99. [Google Scholar] [CrossRef]
- Bhat, S.A.; Singh, S.; Singh, J.; Kumar, S.; Bhawana; Vig, A.P. Bioremediation and detoxification of industrial wastes by earthworms: Vermicompost as powerful crop nutrient in sustainable agriculture. Bioresour. Technol. 2018, 252, 172–179. [Google Scholar] [CrossRef]
- Samal, K.; Raj Mohan, A.; Chaudhary, N.; Moulick, S. Application of vermitechnology in waste management: A review on mechanism and performance. J. Environ. Chem. Eng. 2019, 7, 103392. [Google Scholar] [CrossRef]
- Ahmad, A.; Aslam, Z.; Bellitürk, K.; Iqbal, N.; Naeem, S.; Idrees, M.; Kaleem, Z.; Nawaz, M.Y.; Nawaz, M.; Sajjad, M.; et al. Vermicomposting Methods from Different Wastes: An Environment Friendly, Economically Viable and Socially Acceptable Approach for Crop Nutrition: A Review. Int. J. Food Sci. Agric. 2021, 5, 58–68. [Google Scholar] [CrossRef]
- Elvira, C.; Goicoechea, M.; Sampedro, L.; Mato, S.; Nogales, R. Bioconversion of solid paper-pulp mill sludge by earthworms. Bioresour. Technol. 1996, 57, 173–177. [Google Scholar] [CrossRef]
- Nogales, R.; Cifuentes, C.; Benítez, E. Vermicomposting of winery wastes: A laboratory study. J. Environ. Sci. Health Part B Pestic. Food Contam. Agric. Wastes 2005, 40, 659–673. [Google Scholar] [CrossRef]
- Suthar, S. Vermicomposting potential of Perionyx sansibaricus (Perrier) in different waste materials. Bioresour. Technol. 2007, 98, 1231–1237. [Google Scholar] [CrossRef] [PubMed]
- Chattopadhyay, G.N. Use of vermicomposting biotechnology for recycling organic wastes in agriculture. Int. J. Recycl. Org. Waste Agric. 2012, 1, 8. [Google Scholar] [CrossRef] [Green Version]
- Bhat, S.A.; Singh, J.; Vig, A.P. Earthworms as Organic Waste Managers and Biofertilizer Producers. Waste Biomass Valorization 2018, 9, 1073–1086. [Google Scholar] [CrossRef]
- Lores, M.; Gómez-Brandón, M.; Pérez-Díaz, D.; Domínguez, J. Using FAME profiles for the characterization of animal wastes and vermicomposts. Soil Biol. Biochem. 2006, 38, 2993–2996. [Google Scholar] [CrossRef]
- Mahboub Khomami, A.; Padasht, M.N.; Ajili Lahiji, A.; Shirinfekr, A. The effect of sawdust vermicompost extract on Syngonium podophyllum growth and nutrition. J. Plant. Nutr. 2019, 42, 410–416. [Google Scholar] [CrossRef]
- Thakur, A.; Kumar, A.; Kumar, C.V.; Kiran, B.S.; Kumar, S.; Athokpam, V. A review on vermicomposting: By-products and its importance. Plant. Cell Biotechnol. Mol. Biol. 2021, 22, 156–164. [Google Scholar]
- Barthod, J.; Rumpel, C.; Dignac, M.F. Composting with additives to improve organic amendments. A review. Agron. Sustain. Dev. 2018, 38, 17. [Google Scholar] [CrossRef] [Green Version]
- Tognetti, C.; Laos, F.; Mazzarino, M.J.; Hernández, M.T. Composting vs. vermicomposting: A comparison of end product quality. Compos. Sci. Util. 2005, 13, 6–13. [Google Scholar] [CrossRef]
- Guo, L.; Wu, G.; Li, C.; Liu, W.; Yu, X.; Cheng, D.; Jiang, G. Vermicomposting with maize increases agricultural benefits by 304%. Agron. Sustain. Dev. 2015, 35, 1149–1155. [Google Scholar] [CrossRef] [Green Version]
- Shi, Z.; Liu, J.; Tang, Z.; Zhao, Y.; Wang, C. Vermiremediation of organically contaminated soils: Concepts, current status, and future perspectives. Appl. Soil Ecol. 2019, 147, 103377. [Google Scholar] [CrossRef]
- Bhat, S.A.; Singh, J.; Vig, A.P. Potential utilization of bagasse as feed material for earthworm Eisenia fetida and production of vermicompost. Springerplus 2015, 4, 11. [Google Scholar] [CrossRef] [Green Version]
- Bouché, M.B. PU-43-Stratégies-Lombriciennes; Lohm, U., Persson, T., Eds.; Swedish Natural Science Research Council: Stockholm, Sweden, 1977; pp. 122–132. [Google Scholar]
- Gajalakshmi, S.; Abbasi, S.A. Earthworms and vermicomposting. Indian J. Biotechnol. 2004, 3, 486–494. [Google Scholar]
- Atiyeh, R.M.; Lee, S.; Edwards, C.A.; Arancon, N.Q.; Metzger, J.D. The influence of humic acids derived from earthworm. Bioresour. Technol. 2002, 84, 7–14. [Google Scholar] [CrossRef]
- Vivas, A.; Moreno, B.; Garcia-Rodriguez, S.; Benitez, E. Assessing the impact of composting and vermicomposting on bacterial community size and structure, and microbial functional diversity of an olive-mill waste. Bioresour. Technol. 2009, 100, 1319–1326. [Google Scholar] [CrossRef] [PubMed]
- Maji, D.; Misra, P.; Singh, S.; Kalra, A. Humic acid rich vermicompost promotes plant growth by improving microbial community structure of soil as well as root nodulation and mycorrhizal colonization in the roots of Pisum sativum. Appl. Soil Ecol. 2017, 110, 97–108. [Google Scholar] [CrossRef]
- Lv, B.; Xing, M.; Yang, J. Exploring the effects of earthworms on bacterial profiles during vermicomposting process of sewage sludge and cattle dung with high-throughput sequencing. Environ. Sci. Pollut. Res. 2018, 25, 12528–12537. [Google Scholar] [CrossRef] [PubMed]
- Arancon, N.; Van Cleave, J.; Hamasaki, R.; Nagata, K.; Felts, J. The influence of vermicompost water extracts on growth of plants propagated by cuttings. J. Plant. Nutr. 2020, 43, 176–185. [Google Scholar] [CrossRef]
- Domínguez, J.; Aira, M.; Kolbe, A.R.; Gómez-Brandón, M.; Pérez-Losada, M. Changes in the composition and function of bacterial communities during vermicomposting may explain beneficial properties of vermicompost. Sci. Rep. 2019, 9, 9657. [Google Scholar] [CrossRef] [Green Version]
- Brandón, M.G.; Aira, M.; Kolbe, A.R.; de Andrade, N.; Pérez-Losada, M.; Domínguez, J. Rapid bacterial community changes during vermicomposting of grape marc derived from red winemaking. Microorganisms 2019, 7, 473. [Google Scholar] [CrossRef] [Green Version]
- Pattnaik, S.; Reddy, M.V. Nutrient Status of Vermicompost of Urban Green Waste Processed by Three Earthworm Species—Eisenia fetida, Eudrilus eugeniae, and Perionyx excavatus. Appl. Environ. Soil Sci. 2010, 2010, 967526. [Google Scholar] [CrossRef] [Green Version]
- Chitrapriya, K.; Asokan, S.; Nagarajan, R. Estimating the Level of Phosphate Solubilising Bacteria and Azotobacter in the Vermicompost of Eudrilus Eugeniae and Perionyx Excavatus with Various Combinations of Cow- Dung and Saw-Dust. Int. J. Sci. Res. 2013, 3, 1–6. [Google Scholar]
- Sinha, R.K.; Agarwal, S.; Chauhan, K.; Valani, D. The wonders of earthworms & its vermicompost in farm production: Charles Darwin’s ‘friends of farmers’, with potential to replace destructive chemical fertilizers. Agric. Sci. 2010, 1, 76–94. [Google Scholar] [CrossRef] [Green Version]
- Gómez-Brandón, M.; Aira, M.; Lores, M.; Domínguez, J. Changes in microbial community structure and function during vermicomposting of pig slurry. Bioresour. Technol. 2011, 102, 4171–4178. [Google Scholar] [CrossRef]
- Gómez-Brandón, M.; Lores, M.; Domínguez, J. Species-specific effects of epigeic earthworms on microbial community structure during first stages of decomposition of organic matter. PLoS ONE 2012, 7, e31895. [Google Scholar] [CrossRef] [Green Version]
- Kolbe, A.R.; Aira, M.; Gómez-Brandón, M.; Pérez-Losada, M.; Domínguez, J. Bacterial succession and functional diversity during vermicomposting of the white grape marc Vitis vinifera v. Albariño. Sci. Rep. 2019, 9, 7472. [Google Scholar] [CrossRef]
- Gopal, M.; Bhute, S.S.; Gupta, A.; Prabhu, S.R.; Thomas, G.V.; Whitman, W.B.; Jangid, K. Changes in structure and function of bacterial communities during coconut leaf vermicomposting. Antonie Van Leeuwenhoek 2017, 110, 1339–1355. [Google Scholar] [CrossRef]
- Monroy, F.; Aira, M.; Domínguez, J. Reduction of total coliform numbers during vermicomposting is caused by short-term direct effects of earthworms on microorganisms and depends on the dose of application of pig slurry. Sci. Total Environ. 2009, 407, 5411–5416. [Google Scholar] [CrossRef]
- Savci, S. Investigation of Effect of Chemical Fertilizers on Environment. APCBEE Procedia 2012, 1, 287–292. [Google Scholar] [CrossRef] [Green Version]
- Lončarić, Z.; Engler, M.; Karalić, K.; Bukvić, G.; Lončarić, R.; Kralik, D. Ocjena kvalitete vermikompostiranog goveđeg stajskog gnoja. Poljoprivreda 2005, 11, 57–63. [Google Scholar]
- Manivannan, S.; Balamurugan, M.; Parthasarathi, K.; Gunasekaran, G.; Ranganathan, L.S. Effect of vermicompost on soil fertility and crop productivity—Beans (Phaseolus vulgaris). J. Environ. Biol. 2009, 30, 275–281. [Google Scholar]
- Joshi, R.; Singh, J.; Vig, A.P. Vermicompost as an effective organic fertilizer and biocontrol agent: Effect on growth, yield and quality of plants. Rev. Environ. Sci. Biotechnol. 2015, 14, 137–159. [Google Scholar] [CrossRef]
- Ayastuy, M.E.; Muscolino, C.; Fernandez, J.A.; Belladonna, D.; Rodriguez, R.A.; Caro, L.; Hernandez, L.F. Effect of organic substrate and aqueous extract of vermicompost on nursery basil growth. Acta Hortic. 2020, 1268. [Google Scholar] [CrossRef]
- Arancon, N.Q.; Lee, S.; Edwards, C.A.; Atiyeh, R. Effects of humic acids derived from cattle, food and paper-waste vermicomposts on growth of greenhouse plants. Pedobiologia 2003, 47, 741–744. [Google Scholar] [CrossRef] [Green Version]
- Arancon, N.Q.; Edwards, C.A.; Lee, S.; Byrne, R. Effects of humic acids from vermicomposts on plant growth. Eur. J. Soil Biol. 2006, 42, 65–69. [Google Scholar] [CrossRef]
- Hernandez, O.L.; Calderín, A.; Huelva, R.; Martínez-Balmori, D.; Guridi, F.; Aguiar, N.O.; Olivares, F.L.; Canellas, L.P. Humic substances from vermicompost enhance urban lettuce production. Agron. Sustain. Dev. 2015, 35, 225–232. [Google Scholar] [CrossRef] [Green Version]
- Ramnarain, Y.I.; Ori, L.; Ansari, A.A. Evaluation of the use of vermicompost on the crop production of two varieties of Pak choi (Brassica rapa var. chinensis) and on the soil structure in Suriname. Asian J. Agric. 2017, 1, 73–79. [Google Scholar] [CrossRef] [Green Version]
- Morsomme, P.; Boutry, M. The plant plasma membrane H+-ATPase: Structure, function and regulation. Biochim. Biophys. Acta Biomembr. 2000, 1465, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Gholami, H.; Ghani, A.; Raouf Fard, F.; Saharkhiz, M.J.; Hazrati, H. Changes in photosynthetic pigments and uptake of some soil elements by chicory supplied with organic fertilizers. Acta Ecol. Sin. 2019, 39, 250–256. [Google Scholar] [CrossRef]
- Maruri-López, I.; Aviles-Baltazar, N.Y.; Buchala, A.; Serrano, M. Intra and extracellular journey of the phytohormone salicylic acid. Front. Plant. Sci. 2019, 10, 423. [Google Scholar] [CrossRef]
- Olaetxea, M.; De Hita, D.; Garcia, C.A.; Fuentes, M.; Baigorri, R.; Mora, V.; Garnica, M.; Urrutia, O.; Erro, J.; Zamarreño, A.M.; et al. Hypothetical framework integrating the main mechanisms involved in the promoting action of rhizospheric humic substances on plant root- and shoot- growth. Appl. Soil Ecol. 2018, 123, 521–537. [Google Scholar] [CrossRef]
- Zandonadi, D.B.; Santos, M.P.; Dobbss, L.B.; Olivares, F.L.; Canellas, L.P.; Binzel, M.L.; Okorokova-Façanha, A.L.; Façanha, A.R. Nitric oxide mediates humic acids-induced root development and plasma membrane H+-ATPase activation. Planta 2010, 231, 1025–1036. [Google Scholar] [CrossRef]
- Mora, V.; Bacaicoa, E.; Baigorri, R.; Zamarreño, A.M.; García-Mina, J.M. NO and IAA Key Regulators in the Shoot Growth Promoting Action of Humic Acid in Cucumis sativus L. J. Plant. Growth Regul. 2014, 33, 430–439. [Google Scholar] [CrossRef]
- Plavšin, I.; Velki, M.; Ečimović, S.; Vrandečić, K.; Ćosić, J. Inhibitory effect of earthworm coelomic fluid on growth of the plant parasitic fungus Fusarium oxysporum. Eur. J. Soil Biol. 2017, 78, 1–6. [Google Scholar] [CrossRef]
- McManus, P.S.; Stockwell, V.O.; Sundin, G.W.; Jones, A.L. Antibiotic use in plant agriculture. Annu. Rev. Phytopathol. 2002, 40, 443–465. [Google Scholar] [CrossRef]
- Szczech, M.M. Suppressiveness of vermicompost against fusarium wilt of tomato. J. Phytopathol. 1999, 147, 155–161. [Google Scholar] [CrossRef]
- Amooaghaie, R.; Golmohammadi, S. Effect of Vermicompost on Growth, Essential Oil, and Health of Thymus Vulgaris. Compos. Sci. Util. 2017, 25, 166–177. [Google Scholar] [CrossRef]
- Sallaku, G.; Babaj, I.; Kaciu, S.; Balliu, A. The influence of vermicompost on plant growth characteristics of cucumber (Cucumis sativus L.) seedlings under saline conditions. J. Food Agric. Environ. 2009, 7, 869–872. [Google Scholar]
- Lazcano, C.; Sampedro, L.; Zas, R.; Domínguez, J. Vermicompost enhances germination of the maritime pine (Pinus pinaster Ait.). New For. 2010, 39, 387–400. [Google Scholar] [CrossRef] [Green Version]
- Ievinsh, G. Vermicompost treatment differentially affects seed germination, seedling growth and physiological status of vegetable crop species. Plant. Growth Regul. 2011, 65, 169–181. [Google Scholar] [CrossRef]
- Goswami, L.; Sarkar, S.; Mukherjee, S.; Das, S.; Barman, S.; Raul, P.; Bhattacharyya, P.; Mandal, N.C.; Bhattacharya, S.; Bhattacharya, S.S. Vermicomposting of Tea Factory Coal Ash: Metal accumulation and metallothionein response in Eisenia fetida (Savigny) and Lampito mauritii (Kinberg). Bioresour. Technol. 2014, 166, 96–102. [Google Scholar] [CrossRef]
- He, X.; Zhang, Y.; Shen, M.; Zeng, G.; Zhou, M.; Li, M. Effect of vermicomposting on concentration and speciation of heavy metals in sewage sludge with additive materials. Bioresour. Technol. 2016, 218, 867–873. [Google Scholar] [CrossRef]
- Wang, Y.; Han, W.; Wang, X.; Chen, H.; Zhu, F.; Wang, X.; Lei, C. Speciation of heavy metals and bacteria in cow dung after vermicomposting by the earthworm, Eisenia fetida. Bioresour. Technol. 2017, 245, 411–418. [Google Scholar] [CrossRef] [PubMed]
- Maity, S.; Bhattacharya, S.; Chaudhury, S. Metallothionein response in earthworms Lampito mauritii (Kinberg) exposed to fly ash. Chemosphere 2009, 77, 319–324. [Google Scholar] [CrossRef]
- Lv, B.; Xing, M.; Yang, J. Speciation and transformation of heavy metals during vermicomposting of animal manure. Bioresour. Technol. 2016, 209, 397–401. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Zhu, P.; Xue, J. Comparative Study on Physical and Chemical Characteristics of Sludge Vermicomposted by Eisenia fetida. Procedia Environ. Sci. 2012, 16, 418–423. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Zhang, Y.; Lian, J.; Chao, J.; Gao, Y.; Yang, F.; Zhang, L. Impact of fly ash and phosphatic rock on metal stabilization and bioavailability during sewage sludge vermicomposting. Bioresour. Technol. 2013, 136, 281–287. [Google Scholar] [CrossRef]
- Merdy, P.; Gharbi, L.T.; Lucas, Y. Pb, Cu and Cr interactions with soil: Sorption experiments and modelling. Colloids Surf. A Physicochem. Eng. Asp. 2009, 347, 192–199. [Google Scholar] [CrossRef]
- Sedlak, D.L.; Chan, P.G. Reduction of hexavalent chromium by ferrous iron. Geochim. Cosmochim. Acta 1997, 61, 2185–2192. [Google Scholar] [CrossRef]
- Qu, M.; Chen, J.; Huang, Q.; Chen, J.; Xu, Y.; Luo, J.; Wang, K.; Gao, W.; Zheng, Y. Bioremediation of hexavalent chromium contaminated soil by a bioleaching system with weak magnetic fields. Int. Biodeterior. Biodegrad. 2018, 128, 41–47. [Google Scholar] [CrossRef]
- García, A.C.; Izquierdo, F.G.; Sobrinho, N.M.B.D.A.; Castro, R.N.; Santos, L.A.; De Souza, L.G.A.; Berbara, R.L.L. Humified insoluble solid for efficient decontamination of nickel and lead in industrial effluents. J. Environ. Chem. Eng. 2013, 1, 916–924. [Google Scholar] [CrossRef]
- Chen, W.; Habibul, N.; Liu, X.Y.; Sheng, G.P.; Yu, H.Q. FTIR and synchronous fluorescence heterospectral two-dimensional correlation analyses on the binding characteristics of copper onto dissolved organic matter. Environ. Sci. Technol. 2015, 49, 2052–2058. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Tian, Y.; Hu, D.; Fan, J.; Shen, M.; Zeng, G. Is vermicompost the possible in situ sorbent? Immobilization of Pb, Cd and Cr in sediment with sludge derived vermicompost, a column study. J. Hazard. Mater. 2019, 367, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Bakar, A.A.; Mahmood, N.Z.; Da Silva, J.A.T.; Abdullah, N.; Jamaludin, A.A. Vermicomposting of sewage sludge by Lumbricus rubellus using spent mushroom compost as feed material: Effect on concentration of heavy metals. Biotechnol. Bioprocess. Eng. 2011, 16, 1036–1043. [Google Scholar] [CrossRef]
- Rorat, A.; Suleiman, H.; Grobelak, A.; Grosser, A.; Kacprzak, M.; Płytycz, B.; Vandenbulcke, F. Interactions between sewage sludge-amended soil and earthworms—Comparison between Eisenia fetida and Eisenia andrei composting species. Environ. Sci. Pollut. Res. 2016, 23, 3026–3035. [Google Scholar] [CrossRef]
- Fried, G.; Chauvel, B.; Reynaud, P.; Sache, I. Decreases in Crop Production by Non-native Weeds, Pests, and Pathogens. In Impact of Biological Invasions on Ecosystem Services; Springer: Cham, Switzerland, 2017. [Google Scholar] [CrossRef]
- Fernandes, M.E.S.; Alves, F.M.; Pereira, R.C.; Aquino, L.A.; Fernandes, F.L.; Zanuncio, J.C. Lethal and sublethal effects of seven insecticides on three beneficial insects in laboratory assays and field trials. Chemosphere 2016, 156, 45–55. [Google Scholar] [CrossRef]
- Aktar, W.; Sengupta, D.; Chowdhury, A. Impact of pesticides use in agriculture: Their benefits and hazards. Interdiscip. Toxicol. 2009, 2, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Yatoo, A.M.; Ali, M.N.; Baba, Z.A.; Hassan, B. Sustainable management of diseases and pests in crops by vermicompost and vermicompost tea. A review. Agron. Sustain. Dev. 2021, 41, 1–26. [Google Scholar] [CrossRef]
- Roubalová, R.; Procházková, P.; Hanč, A.; Dvořák, J.; Bilej, M. Mutual interactions of E. andrei earthworm and pathogens during the process of vermicomposting. Environ. Sci. Pollut. Res. 2020, 27, 33429–33437. [Google Scholar] [CrossRef]
- Rodríguez-Canché, L.G.; Cardoso Vigueros, L.; Maldonado-Montiel, T.; Martínez-Sanmiguel, M. Pathogen reduction in septic tank sludge through vermicomposting using Eisenia fetida. Bioresour. Technol. 2010, 101, 3548–3553. [Google Scholar] [CrossRef] [PubMed]
- Pachepsky, Y.A.; Sadeghi, A.M.; Bradford, S.A.; Shelton, D.R.; Guber, A.K.; Dao, T. Transport and fate of manure-borne pathogens: Modeling perspective. Agric. Water Manag. 2006, 86, 81–92. [Google Scholar] [CrossRef]
- Monroy, F.; Aira, M.; Domínguez, J. Changes in density of nematodes, protozoa and total coliforms after transit through the gut of four epigeic earthworms (Oligochaeta). Appl. Soil Ecol. 2008, 39, 127–132. [Google Scholar] [CrossRef]
- Dales, R.P.; Kalaç, Y. Phagocytic defence by the earthworm Eisenia foetida against certain pathogenic bacteria. Comp. Biochem. Physiol. Part. A Physiol. 1992, 101, 487–490. [Google Scholar] [CrossRef]
- Sarma, B.K.; Singh, P.; Susheel, P.; Harikesh, S. Vermicompost as Modulator of Plant Growth and Disease Suppression. Glob. Sci. Books 2010, 4, 58–66. [Google Scholar]
- Jangra, M.; Sindhu, S.; Gulati, R.; Batra, V.K. Studies on efficacy of vermicompost for the management of Polyphagotarsonemus latus (Banks) (Acari: Tarsonemidae) infesting chilli (Capsicum annuum L.) in Haryana. Pharm. Innovat. J. 2019, 8, 86–89. [Google Scholar]
- You, X.; Kimura, N.; Okura, T.; Murakami, S.; Okano, R.; Shimogami, Y.; Matsumura, A.; Tokumoto, H.; Ogata, Y.; Tojo, M. Suppressive effects of vermicomposted-bamboo powder on cucumber damping-off. Jpn. Agric. Res. Q. 2019, 53, 13–19. [Google Scholar] [CrossRef] [Green Version]
- Basco, M.J.; Bisen, K.; Keswani, C.; Singh, H.B. Biological management of Fusarium wilt of tomato using biofortified vermicompost. Mycosphere 2017, 8, 467–483. [Google Scholar] [CrossRef]
- Liu, D.; Liu, D.; Liu, L.; Wang, Y.; Zhang, Y. Screening and identification of antagonistic bacteria from vermicompost against Fusarium oxysporum f. sp. cucumerinum. Acta Agric. Scand. Sect. B Soil Plant. Sci. 2021, 71, 266–272. [Google Scholar] [CrossRef]
- Simsek Ersahin, Y.; Haktanir, K.; Yanar, Y. Vermicompost suppresses Rhizoctonia solani Kühn in cucumber seedlings. J. Plant Dis. Prot. 2009, 116, 182–188. [Google Scholar] [CrossRef]
- Szczech, M.; Smolinska, U. Comparison of suppressiveness of vermicomposts produced from animal manures and sewage sludge against phytophthora nicotianae breda de haan var. nicotianae. J. Phytopathol. 2001, 149, 77–82. [Google Scholar] [CrossRef]
- Arancon, N.Q.; Edwards, C.A.; Yardim, E.N.; Oliver, T.J.; Byrne, R.J.; Keeney, G. Suppression of two-spotted spider mite (Tetranychus urticae), mealy bug (Pseudococcus sp.) and aphid (Myzus persicae) populations and damage by vermicomposts. Crop. Prot. 2007, 26, 29–39. [Google Scholar] [CrossRef]
- Cardoza, Y.J.; Buhler, W.G. Soil organic amendment impacts on corn resistance to Helicoverpa zea: Constitutive or induced? Pedobiologia 2012, 55, 343–347. [Google Scholar] [CrossRef]
- Xiao, Z.; Liu, M.; Jiang, L.; Chen, X.; Griffiths, B.S.; Li, H.; Hu, F. Vermicompost increases defense against root-knot nematode (Meloidogyne incognita) in tomato plants. Appl. Soil Ecol. 2016, 105, 177–186. [Google Scholar] [CrossRef]
- Vogt, T. Phenylpropanoid biosynthesis. Mol. Plant. 2010, 3, 2–20. [Google Scholar] [CrossRef] [Green Version]
- Bavaresco, L.; Pezzutto, S.; Gatti, M.; Mattivi, F. Role of the variety and some environmental factors on grape stilbenes. Vitis J. Grapevine Res. 2007, 46, 57–61. [Google Scholar]
- Mupambwa, H.A.; Mnkeni, P.N.S. Optimizing the vermicomposting of organic wastes amended with inorganic materials for production of nutrient-rich organic fertilizers: A review. Environ. Sci. Pollut. Res. 2018, 25, 10577–10595. [Google Scholar] [CrossRef]
- Blouin, M.; Barrere, J.; Meyer, N.; Lartigue, S.; Barot, S.; Mathieu, J. Vermicompost significantly affects plant growth. A meta-analysis. Agron. Sustain. Dev. 2019, 39, 34. [Google Scholar] [CrossRef]
- Swati, A.; Hait, S. Fate and bioavailability of heavy metals during vermicomposting of various organic wastes—A review. Process. Saf. Environ. Prot. 2017, 109, 30–45. [Google Scholar] [CrossRef]
- Chand, S.; Pandey, A.; Patra, D.D. Influence of nickel and lead applied in combination with vermicompost on growth and accumulation of heavy metals by Mentha arvensis Linn. cv. “Kosi.”. Indian J. Nat. Prod. Resour. 2012, 3, 256–261. [Google Scholar]
- Chand, S.; Kumari, R.; Patra, D.D. Effect of Nickel and Vermicompost on Growth, Yield, Accumulation of Heavy Metals and Essential Oil Quality of Tagetes minuta. J. Essent. Oil Bear. Plants 2015, 18, 767–774. [Google Scholar] [CrossRef]
- Sengupta, S.; Mukherjee, S.; Halder, S. Enrichment of vermicompost for improving soil quality and ensuring Zn and Fe bioavailability through rice grain. J. Pharmacogn. Phytochem. 2020, 9, 246–254. [Google Scholar]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vuković, A.; Velki, M.; Ečimović, S.; Vuković, R.; Štolfa Čamagajevac, I.; Lončarić, Z. Vermicomposting—Facts, Benefits and Knowledge Gaps. Agronomy 2021, 11, 1952. https://doi.org/10.3390/agronomy11101952
Vuković A, Velki M, Ečimović S, Vuković R, Štolfa Čamagajevac I, Lončarić Z. Vermicomposting—Facts, Benefits and Knowledge Gaps. Agronomy. 2021; 11(10):1952. https://doi.org/10.3390/agronomy11101952
Chicago/Turabian StyleVuković, Ana, Mirna Velki, Sandra Ečimović, Rosemary Vuković, Ivna Štolfa Čamagajevac, and Zdenko Lončarić. 2021. "Vermicomposting—Facts, Benefits and Knowledge Gaps" Agronomy 11, no. 10: 1952. https://doi.org/10.3390/agronomy11101952
APA StyleVuković, A., Velki, M., Ečimović, S., Vuković, R., Štolfa Čamagajevac, I., & Lončarić, Z. (2021). Vermicomposting—Facts, Benefits and Knowledge Gaps. Agronomy, 11(10), 1952. https://doi.org/10.3390/agronomy11101952