Early Response of Soil Microbial Biomass and Activity to Biofertilizer Application in Degraded Brunic Arenosol and Abruptic Luvisol of Contrasting Textures


 , ,
, , 

Abstract
1. Introduction
2. Materials and Methods
2.1. Field Study Site and Soil Description
2.2. Laboratory Experiment Design
2.3. Methods
2.4. Statistical Analysis
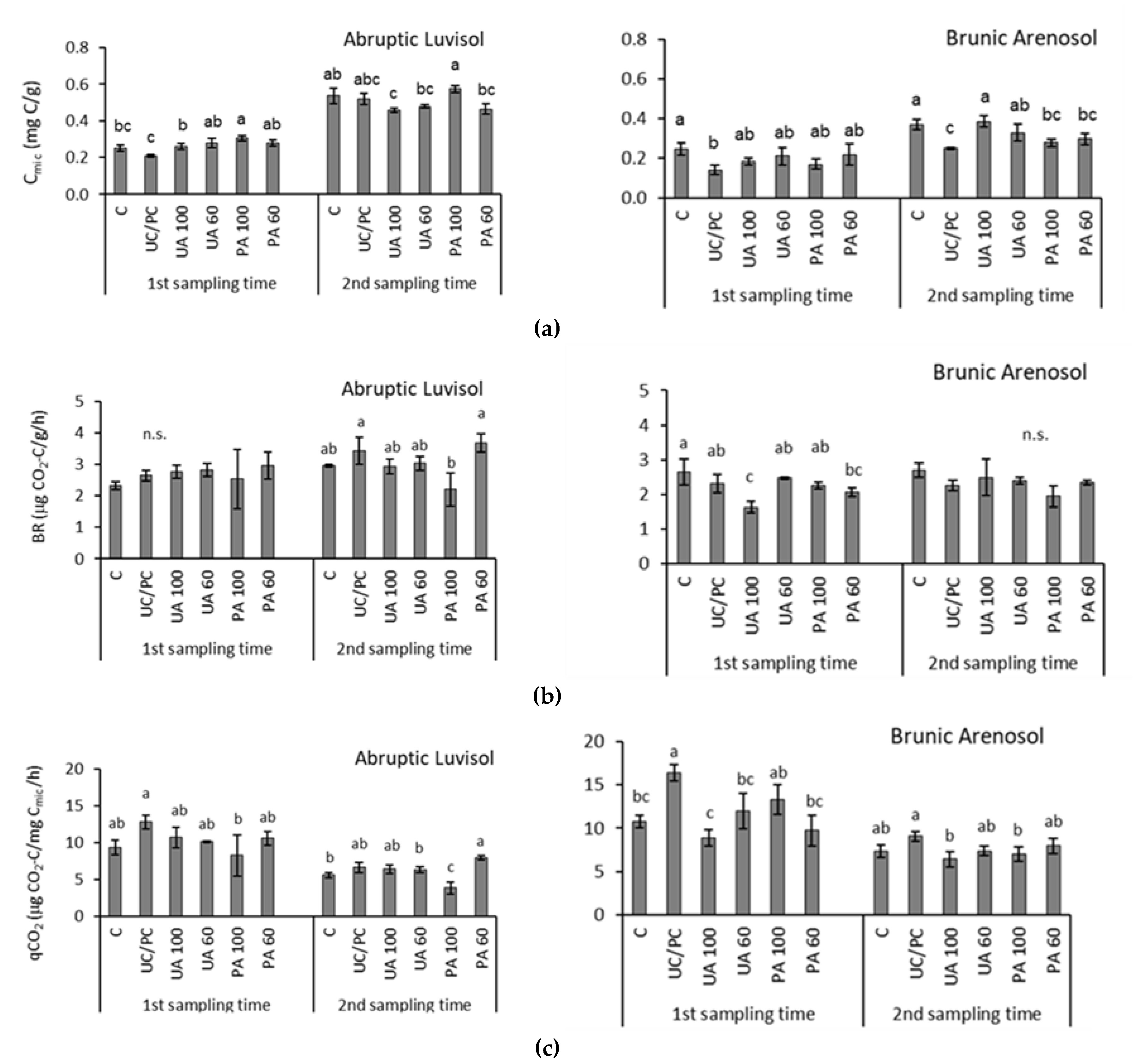
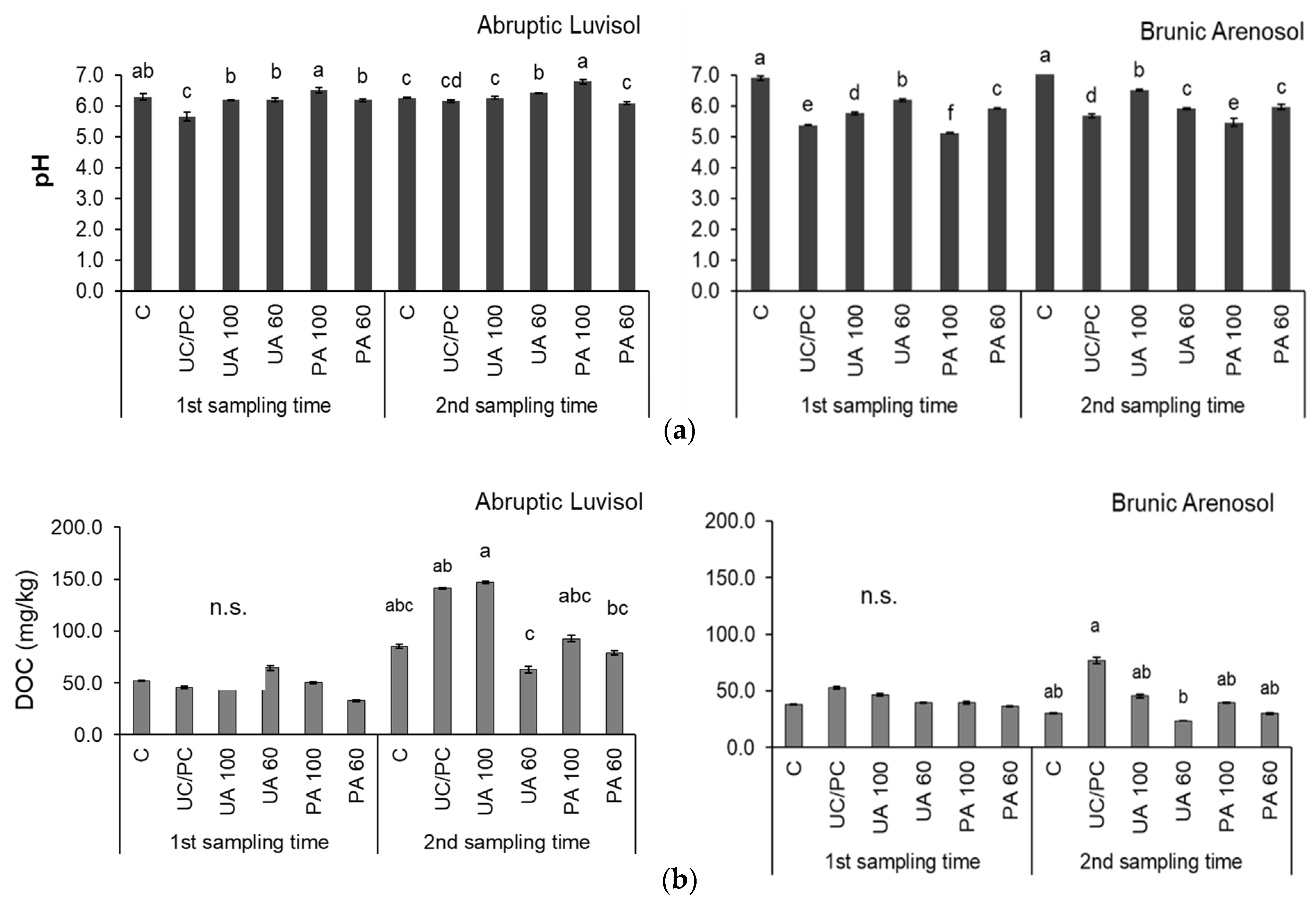
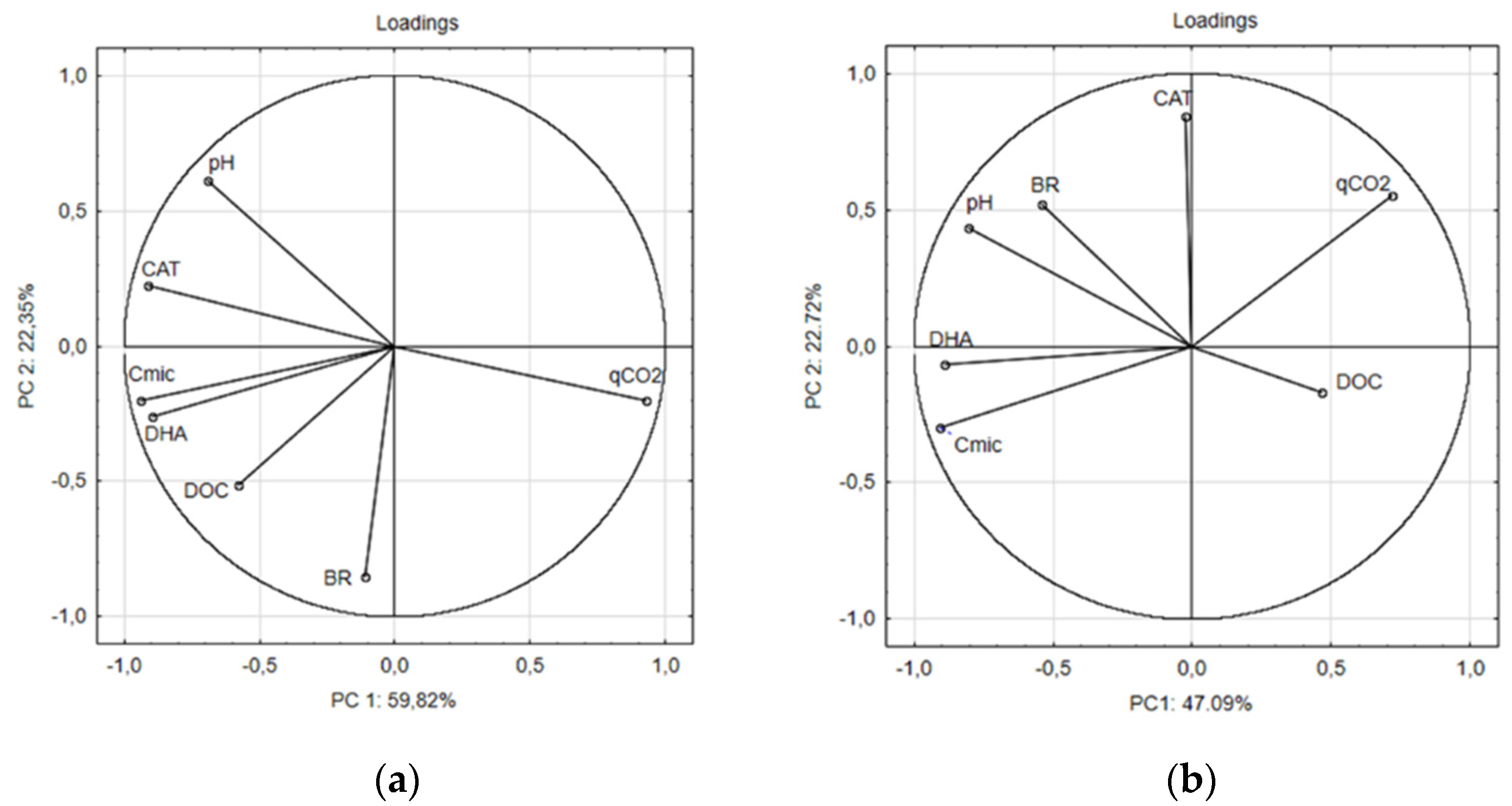
3. Results and Discussion
3.1. Physical and Chemical Properties of Soils without Fertilization
3.2. Microbial Indicators of Soils with Fertilizer and with Biofertilizers
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Prăvălie, R.; Patriche, C.V.; Bandoc, G. Quantification of land degradation sensitivity areas in Southern and Central Southeastern Europe. New results based on improving DISMED methodology with new climate data. Catena 2017, 158, 309–320. [Google Scholar] [CrossRef]
- Mącik, M.; Gryta, A.; Frąc, M. Biofertilizers in agriculture: An overview on concepts, strategies and effects on soil microorganisms. Adv. Agron. 2020, 162, 31–87. [Google Scholar] [CrossRef]
- Suhag, M. Potential of biofertilizers to replace chemical fertilizers. Int. Adv. Res. J. Sci. Eng. Technol. 2016, 3, 163–167. [Google Scholar] [CrossRef]
- Vessey, J.K. Plant growth promoting rhizobacteria as biofertilizers. Plant Soil 2003, 255, 571–586. [Google Scholar] [CrossRef]
- Itelima, J.U.; Bang, W.J.; Sila, M.D.; Onyimba, I.A.; Egbere, O.J. A review: Biofertilizer—A key player in enhancing soil fertility and crop productivity. J. Microbiol. Biotechnol. Rep. 2018, 2, 22–28. [Google Scholar]
- Ellafi, A.M.; Gadalla, A.; Galal, Y.G.M. Biofertilizers in action: Contributions of BNF in sustainable agricultural ecosystems. E-Int. Sci. Res. J. 2011, 3, 108–116. [Google Scholar]
- Faujdar, R.S.; Sharma, M. Effect of FYM, biofertilizers and Zinc on yield of maize and their residual effect on wheat. J. Soils Crops 2013, 23, 41–52. [Google Scholar]
- Mosaad, I.S.M. Effect of Mineral and Bio-Nitrogen Fertilization on Maize (Zea mays L.), some Soil Properties and Subsequent Wheat (Triticum aestivum, L.) Yield. Alex. Sci. Exch. J. 2016, 37, 550–560. [Google Scholar] [CrossRef]
- Nain, L.; Rana, A.; Joshi, M.; Jadhav, S.D.; Kumar, D.; Shivay, Y.S.; Paul, S.; Prasanna, R. Evaluation of synergistic effects of bacterial and cyanobacterial strains as biofertilizers for wheat. Plant Soil 2009, 331, 217–230. [Google Scholar] [CrossRef]
- Kowalska, J.; Niewiadomska, A.; Głuchowska, K.; Kaczmarek, D. Impact of fertilizers on soil properties in the case of Solanum Tuberosum, L. during conversion to organic farming. Appl. Ecol. Environ. Res. 2017, 15, 369–383. [Google Scholar] [CrossRef]
- Khan, H.I. Appraisal of Biofertilizers in Rice: To Supplement Inorganic Chemical Fertilizer. Rice Sci. 2018, 25, 357–362. [Google Scholar] [CrossRef]
- Simarmata, T.; Hersanti; Turmuktini, T.; Fitriatin, B.N.; Setiawati, M.R. Purwanto Application of Bioameliorant and Biofertilizers to Increase the Soil Health and Rice Productivity. HAYATI J. Biosci. 2016, 23, 181–184. [Google Scholar] [CrossRef]
- Dong, L.; Li, Y.; Xu, J.; Yang, J.; Wei, G.; Shen, L.; Ding, W.; Chen, S. Biofertilizers regulate the soil microbial community and enhance Panax ginseng yields. Chin. Med. 2019, 14, 20. [Google Scholar] [CrossRef] [PubMed]
- Mosa, W.F.A.E.-G.; Paszt, L.S.; Frąc, M.; Trzciński, P. The Role of Biofertilization in Improving Apple Productivity―A Review. Adv. Microbiol. 2015, 5, 21–27. [Google Scholar] [CrossRef][Green Version]
- Babajide, P.A.; Akanbi, W.B.; Alamu, L.O.; Ewetola, E.A.; Olatunji, O.O. Growth, nodulation and biomass yield of soybean (Glycine max L.) as influenced by bio-fertilizers under simulated eroded soil condition. Res. J. Agron. 2008, 2, 96–100. [Google Scholar]
- Mukhopadhyay, S.; Maiti, S.K. Biofertilizer: VAM fungi–A future prospect for biological reclamation of mine degraded lands. Indian J. Environ. Prot. 2009, 29, 801–808. [Google Scholar]
- Aseri, G.; Jain, N.; Panwar, J.; Rao, A.; Meghwal, P. Biofertilizers improve plant growth, fruit yield, nutrition, metabolism and rhizosphere enzyme activities of Pomegranate (Punica granatum L.) in Indian Thar Desert. Sci. Hortic. 2008, 117, 130–135. [Google Scholar] [CrossRef]
- Włodarczyk, T.; Stepniewski, W.; Brzezińska, M. Dehydrogenase activity, redox potential, and emissions of carbon dioxide and nitrous oxide from Cambisols under flooding conditions. Boil. Fertil. Soils 2002, 36, 200–206. [Google Scholar] [CrossRef]
- Gianfreda, L.; Rao, M.; Piotrowska-Długosz, A.; Palumbo, G.; Colombo, C. Soil enzyme activities as affected by anthropogenic alterations: Intensive agricultural practices and organic pollution. Sci. Total. Environ. 2005, 341, 265–279. [Google Scholar] [CrossRef]
- Moeskops, B.; Sukristiyonubowo; Buchan, D.; Sleutel, S.; Herawaty, L.; Husen, E.; Saraswati, R.; Setyorini, D.; De Neve, S. Soil microbial communities and activities under intensive organic and conventional vegetable farming in West Java, Indonesia. Appl. Soil Ecol. 2010, 45, 112–120. [Google Scholar] [CrossRef]
- Gu, Y.; Wang, P.; Kong, C. Urease, invertase, dehydrogenase and polyphenoloxidase activities in paddy soils influenced by allelophatic rice variety. Eur. J. Soil Biol. 2009, 45, 436–444. [Google Scholar] [CrossRef]
- Salazar, S.; Sanchez, L.; Alvarez, J.; Valverde, A.; Galindo-Villardón, M.P.; Igual, J.M.; Peix, Á.; Santa-Regina, I. Correlation among soil enzyme activities under different forest system management practices. Ecol. Eng. 2011, 37, 1123–1131. [Google Scholar] [CrossRef]
- Wolińska, A.; Stepniewsk, Z. Dehydrogenase Activity in the Soil Environment. Dehydrogenases 2012, 10, 183–210. [Google Scholar] [CrossRef]
- Zhao, B.; Chen, J.; Zhang, J.; Qin, S. Soil microbial biomass and activity response to repeated drying–rewetting cycles along a soil fertility gradient modified by long-term fertilization management practices. Geoderma 2010, 160, 218–224. [Google Scholar] [CrossRef]
- Guwy, A.; Martin, S.; Hawkes, F.; Hawkes, D. Catalase activity measurements in suspended aerobic biomass and soil samples. Enzym. Microb. Technol. 1999, 25, 669–676. [Google Scholar] [CrossRef]
- Alef, K.; Nannipieri, P. Catalase activity. In Methods in applied soil microbiology and biochemistry; Alef, K., Nannipieri, P., Eds.; Academic Press: London, UK, 1995; pp. 362–363. [Google Scholar]
- Trasar-Cepeda, C.; Camiña, F.; Leirós, M.; Gil-Sotres, F. An improved method to measure catalase activity in soils. Soil Boil. Biochem. 1999, 31, 483–485. [Google Scholar] [CrossRef]
- Garcıa-Gil, J.C.; Kobza, J.; Soler-Rovira, P.; Javoreková, S.; García-Gil, J.C. Soil Microbial and Enzyme Activities Response to Pollution Near an Aluminium Smelter. CLEAN—Soil Air Water 2013, 41, 485–492. [Google Scholar] [CrossRef]
- Anderson, T.-H. Microbial eco-physiological indicators to asses soil quality. Agric. Ecosyst. Environ. 2003, 98, 285–293. [Google Scholar] [CrossRef]
- Dilly, O. Microbial energetics in soils. In Microorganisms in Soils: Roles in Genesis and Functions; Buscot, F., Varma, A., Eds.; Soil Biology Series; Springer: Berlin/Heidelberg, Germany, 2005; Volume 3, pp. 123–138. [Google Scholar]
- Araújo, A.S.F.; Santos, V.; Monteiro, R. Responses of soil microbial biomass and activity for practices of organic and conventional farming systems in Piauí state, Brazil. Eur. J. Soil Boil. 2008, 44, 225–230. [Google Scholar] [CrossRef]
- Zhao, F.; Wang, J.; Zhang, L.; Ren, C.; Han, X.; Yang, G.; Doughty, R.; Deng, J. Understory Plants Regulate Soil Respiration through Changes in Soil Enzyme Activity and Microbial C, N, and P Stoichiometry Following Afforestation. Forests 2018, 9, 436. [Google Scholar] [CrossRef]
- Nannipieri, P.; Ascher-Jenull, J.; Ceccherini, M.T.; Landi, L.; Pietramellara, G.; Renella, G. Microbial diversity and soil functions. Eur. J. Soil Sci. 2003, 54, 655–670. [Google Scholar] [CrossRef]
- Anderson, T.-H.; Domsch, K.H. Soil microbial biomass: The eco-physiological approach. Soil Boil. Biochem. 2010, 42, 2039–2043. [Google Scholar] [CrossRef]
- Wardle, D.A.; Ghani, A. A critique of the microbial metabolic quotient (qCO2) as a bioindicator of disturbance and ecosystem development. Soil Biol. Biochem. 1995, 27, 1601–1610. [Google Scholar] [CrossRef]
- Araújo, A.S.F.; Oliveira, J.; Araújo, R.; Gomes, R. Biofertilizers on soil microbial biomass and activity. Rev. Bras. Ciênc. Agrár. —Braz. J. Agric. Sci. 2014, 9, 545–549. [Google Scholar] [CrossRef]
- Zu Schweinsberg-Mickan, M.S.; Müller, T. Impact of effective microorganisms and other biofertilizers on soil microbial characteristics, organic-matter decomposition, and plant growth. J. Plant Nutr. Soil Sci. 2009, 172, 704–712. [Google Scholar] [CrossRef]
- Singh, G.; Kumar, D.; Sharma, P.; Krakauer, N. Effect of organics, biofertilizers and crop residue application on soil microbial activity in rice – wheat and rice-wheat mungbean cropping systems in the Indo-Gangetic plains. Cogent Geosci. 2015, 1, 1085296. [Google Scholar] [CrossRef]
- Piotrowska, A.; Długosz, J.; Zamorski, R.; Bogdanowicz, P. Changes in some biological and chemical properties of an arable soil treated with the microbial biofertilizer UGmax. Pol. J. Environ. Stud. 2012, 21, 455–463. [Google Scholar]
- Borowik, K.; Rusek, P.; Schab, S.; Rusek, Ł. Biofertilizer and Method of Its Production (Bionawóz i sposób jego wytwarzania). Patent Application No. 431350, 10 March 2019. [Google Scholar]
- Bieganowski, A.; Chojecki, T.; Ryżak, M.; Sochan, A.; Lamorski, K. Methodological Aspects of Fractal Dimension Estimation on the Basis of Particle Size Distribution. Vadose Zone J. 2013, 12. [Google Scholar] [CrossRef]
- Bieganowski, A.; Ryżak, M.; Sochan, A.; Barna, G.; Hernádi, H.; Beczek, M.; Polakowski, C.; Makó, A. Laser Diffractometry in the Measurements of Soil and Sediment Particle Size Distribution. Adv. Agron. 2018, 151, 215–279. [Google Scholar] [CrossRef]
- Yu, Z.; Chen, L.; Pan, S.; Li, Y.; Kuzyakov, Y.; Xu, J.; Brookes, P.; Luo, Y. Feedstock determines biochar-induced soil priming effects by stimulating the activity of specific microorganisms. Eur. J. Soil Sci. 2018, 69, 521–534. [Google Scholar] [CrossRef]
- Trevors, J. Rapid gas chromatographic method to measure H2O2 oxidoreductase (catalase) activity in soil. Soil Boil. Biochem. 1984, 16, 525–526. [Google Scholar] [CrossRef]
- Walkiewicz, A.; Brzezińska, M. Interactive effects of nitrate and oxygen on methane oxidation in three different soils. Soil Boil. Biochem. 2019, 133, 116–118. [Google Scholar] [CrossRef]
- Casida, L.E.; Klein, D.A.; Santoro, T. Soil Dehydrogenase Activity. Soil Sci. 1964, 98, 371–376. [Google Scholar] [CrossRef]
- Anderson, J.; Domsch, K. A physiological method for the quantitative measurement of microbial biomass in soils. Soil Boil. Biochem. 1978, 10, 215–221. [Google Scholar] [CrossRef]
- Šimek, M.; Kalčík, J. Carbon and nitrate utilization in soils: The effect of long-term fertilization on potential denitrification. Geoderma 1998, 83, 269–280. [Google Scholar] [CrossRef]
- Gliński, J.; Stępniewski, W. Soil Aeration and Its Role for Plants; CRC Press: Boca Raton, FL, USA, 1985. [Google Scholar]
- Ullah, S.; Frasier, R.; King, L.; Picotte-Anderson, N.; Moore, T.R. Potential fluxes of N2O and CH4 from soils of three forest types in Eastern Canada. Soil Boil. Biochem. 2008, 40, 986–994. [Google Scholar] [CrossRef]
- Wang, W.; Dalal, R.C.; Moody, P.; Smith, C.J. Relationships of soil respiration to microbial biomass, substrate availability and clay content. Soil Boil. Biochem. 2003, 35, 273–284. [Google Scholar] [CrossRef]
- Hemkemeyer, M.; Christensen, B.T.; Martens, R.; Tebbe, C.C. Soil particle size fractions harbour distinct microbial communities and differ in potential for microbial mineralisation of organic pollutants. Soil Boil. Biochem. 2015, 90, 255–265. [Google Scholar] [CrossRef]
- Bollmann, A.; Conrad, R. Influence of O2availability on NO and N2O release by nitrification and denitrification in soils. Glob. Chang. Boil. 1998, 4, 387–396. [Google Scholar] [CrossRef]
- Hamarashid, N.H.; Othman, M.A.; Hussain, M.A.H. Effect of soil texture on chemical compositions, microbial populations and carbon mineralization in soil. Egypt. J. Exp. Biol. 2010, 6, 59–64. [Google Scholar]
- Walkiewicz, A.; Brzezińska, M.; Wnuk, E.; Jabłoński, B. Soil properties and not high CO2 affect CH4 production and uptake in periodically waterlogged arable soils. J. Soils Sediments 2019, 20, 1231–1240. [Google Scholar] [CrossRef]
- Helmi, H.; Hechmi, S.; Khelil, M.N.; Zoghlami, I.R.; Benzarti, S.; Mokni-Tlili, S.; Hassen, A.; Jedidi, N. Repetitive land application of urban sewage sludge: Effect of amendment rates and soil texture on fertility and degradation parameters. Catena 2019, 172, 11–20. [Google Scholar] [CrossRef]
- Tahir, S.; Marschner, B. Clay addition to sandy soil: Effect of clay concentration and ped size on microbial biomass and nutrient dynamics after addition of low C/N ratio residue. J. soil Sci. Plant Nutr. 2016, 16, 864–875. [Google Scholar] [CrossRef][Green Version]
- Wolf, J.; Broeke, M.H.-T.; Rötter, R.P. Simulation of nitrogen leaching in sandy soils in The Netherlands with the ANIMO model and the integrated modelling system STONE. Agric. Ecosyst. Environ. 2005, 105, 523–540. [Google Scholar] [CrossRef]
- Khadem, A.; Raiesi, F. Influence of biochar on potential enzyme activities in two calcareous soils of contrasting texture. Geoderma 2017, 308, 149–158. [Google Scholar] [CrossRef]
- Yang, L.; Li, T.; Li, F.; Lemcoff, J.H.; Cohen, S. Fertilization regulates soil enzymatic activity and fertility dynamics in a cucumber field. Sci. Hortic. 2008, 116, 21–26. [Google Scholar] [CrossRef]
- Monreal, C.; Bergstrom, D.W. Soil enzymatic factors expressing the influence of land use, tillage system and texture on soil biochemical quality. Can. J. Soil Sci. 2000, 80, 419–428. [Google Scholar] [CrossRef]
- Dinesh, D.; Chithra, L.; Baskar, M.; Rajan, K.; Senthilraja, K.; Sankar, M.; Kumar, R.; Sivakumar, K. Variation of Soil Microbial Growth and Enzyme Activities by Application of Treated Distillery Effluent in Maize Crop Grown Under Sandy Loam Soils. Int. J. Curr. Microbiol. Appl. Sci. 2017, 6, 1334–1348. [Google Scholar] [CrossRef]
- Dinesh, R.; Srinivasan, V.; Hamza, S.; Manjusha, A. Short-term incorporation of organic manures and biofertilizers influences biochemical and microbial characteristics of soils under an annual crop [Turmeric (Curcuma longa L.)]. Bioresour. Technol. 2010, 101, 4697–4702. [Google Scholar] [CrossRef]
- Santos, V.B.; Araújo, A.S.F.; Leite, L.F.; Nunes, L.A.P.L.; De Melo, W.J. Soil microbial biomass and organic matter fractions during transition from conventional to organic farming systems. Geoderma 2012, 170, 227–231. [Google Scholar] [CrossRef]
- Tejada, M.; Garcia, C.; González, J.; Hernandez, T. Use of organic amendment as a strategy for saline soil remediation: Influence on the physical, chemical and biological properties of soil. Soil Boil. Biochem. 2006, 38, 1413–1421. [Google Scholar] [CrossRef]
- Hungria, M.; Franchini, J.C.; Brandão-Junior, O.; Kaschuk, G.; Souza, R.A. Soil microbial activity and crop sustainability in a long-term experiment with three soil-tillage and two crop-rotation systems. Appl. Soil Ecol. 2009, 42, 288–296. [Google Scholar] [CrossRef]
- Pietri, J.A.; Brookes, P. Relationships between soil pH and microbial properties in a UK arable soil. Soil Boil. Biochem. 2008, 40, 1856–1861. [Google Scholar] [CrossRef]
- Tan, W.; Wang, J.; Bai, W.; Qi, J.; Chen, W. Soil bacterial diversity correlates with precipitation and soil pH in long-term maize cropping systems. Sci. Rep. 2020, 10, 6012. [Google Scholar] [CrossRef]
- Peiffer, J.A.; Spor, A.; Koren, O.; Jin, Z.; Tringe, S.G.; Dangl, J.L.; Buckler, E.; Ley, R. Diversity and heritability of the maize rhizosphere microbiome under field conditions. Proc. Natl. Acad. Sci. USA 2013, 110, 6548–6553. [Google Scholar] [CrossRef]
- Havlin, J.L.; Beaton, J.D.; Tisdale, S.L.; Nelson, W.L. Soil Fertility and Fertilizers, 6th ed.; Prentice-Hall, Inc.: Upper Saddle River, NJ, USA, 1999; p. 499. [Google Scholar]
- Adak, T.; Singha, A.; Kumar, K.; Shukla, S.K.; Singh, A.; Singh, V.K. Soil organic carbon, dehydrogenase activity, nutrient availability and leaf nutrient content as affected by organic and inorganic source of nutrient in mango orchard soil. J. soil Sci. Plant Nutr. 2014, 14, 394–406. [Google Scholar] [CrossRef]
- Albiach, R.; Canet, R.; Pomares, F.; Ingelmo, F. Organic matter components and aggregate stability after the application of different amendments to a horticultural soil. Bioresour. Technol. 2001, 76, 125–129. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Type | Clay | Silt | Sand | Texture | P2O5 | K2O | Mg | N-NO3 | N-NH4 | Nmin | DOC | Corg | pHH2O |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| (%) | (mg/kg) | (kg/ha) | (mg/kg) | (%) | |||||||||
| Brunic Arenosol | 0.93 | 12.4 | 86.6 | sandy | 174 | 29 | 12 | <1.39 | 2.14 | 22.0 | 37.93 | 1.54 | 6.29 |
| Abruptic Luvisol | 5.4 | 60.3 | 34.3 | silt loam | 48 | 53 | 36 | 2.91 | 6.57 | 75.6 | 52.30 | 3.76 | 6.90 |
| CAT | DHA | Cmic | BR | DOC | pH | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Sampling Time | Ist | IInd | Ist | IInd | Ist | IInd | Ist | IInd | Ist | IInd | Ist | IInd |
| Abruptic Luvisol | ||||||||||||
| UA 100 | + | n.s. | n.s. | n.s. | + | n.s. | n.s. | n.s. | n.s. | n.s. | + | n.s. |
| UA 60 | + | n.s. | n.s. | n.s. | + | n.s. | n.s. | n.s. | n.s. | - | + | + |
| PA 100 | + | + | n.s. | n.s. | + | + | n.s. | - | n.s. | n.s. | + | + |
| PA 60 | + | n.s. | n.s. | n.s. | + | n.s. | n.s. | n.s. | n.s. | n.s. | + | n.s. |
| Brunic Arenosol | ||||||||||||
| UA 100 | n.s. | n.s. | n.s. | + | n.s. | + | - | n.s. | n.s. | n.s. | + | + |
| UA 60 | + | n.s. | n.s. | n.s. | n.s. | + | n.s. | n.s. | n.s. | n.s. | + | + |
| PA 100 | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | - | - |
| PA 60 | n.s. | n.s. | + | + | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | + | + |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Walkiewicz, A.; Brzezińska, M.; Bieganowski, A.; Sas-Paszt, L.; Frąc, M. Early Response of Soil Microbial Biomass and Activity to Biofertilizer Application in Degraded Brunic Arenosol and Abruptic Luvisol of Contrasting Textures. Agronomy 2020, 10, 1347. https://doi.org/10.3390/agronomy10091347
Walkiewicz A, Brzezińska M, Bieganowski A, Sas-Paszt L, Frąc M. Early Response of Soil Microbial Biomass and Activity to Biofertilizer Application in Degraded Brunic Arenosol and Abruptic Luvisol of Contrasting Textures. Agronomy. 2020; 10(9):1347. https://doi.org/10.3390/agronomy10091347
Chicago/Turabian StyleWalkiewicz, Anna, Małgorzata Brzezińska, Andrzej Bieganowski, Lidia Sas-Paszt, and Magdalena Frąc. 2020. "Early Response of Soil Microbial Biomass and Activity to Biofertilizer Application in Degraded Brunic Arenosol and Abruptic Luvisol of Contrasting Textures" Agronomy 10, no. 9: 1347. https://doi.org/10.3390/agronomy10091347
APA StyleWalkiewicz, A., Brzezińska, M., Bieganowski, A., Sas-Paszt, L., & Frąc, M. (2020). Early Response of Soil Microbial Biomass and Activity to Biofertilizer Application in Degraded Brunic Arenosol and Abruptic Luvisol of Contrasting Textures. Agronomy, 10(9), 1347. https://doi.org/10.3390/agronomy10091347