Effects of Post-Anthesis Heat Waves on the Grain Quality of Seven European Wheat Varieties



Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material and High Temperature Treatments
2.2. Genetic Variability Analysis
2.3. Comparative Transcription Analysis
2.4. Protein Fractions Quantification
3. Results and Discussion
3.1. Genomic Analysis Revealed High Similarity on Genes Related with Grain Quality
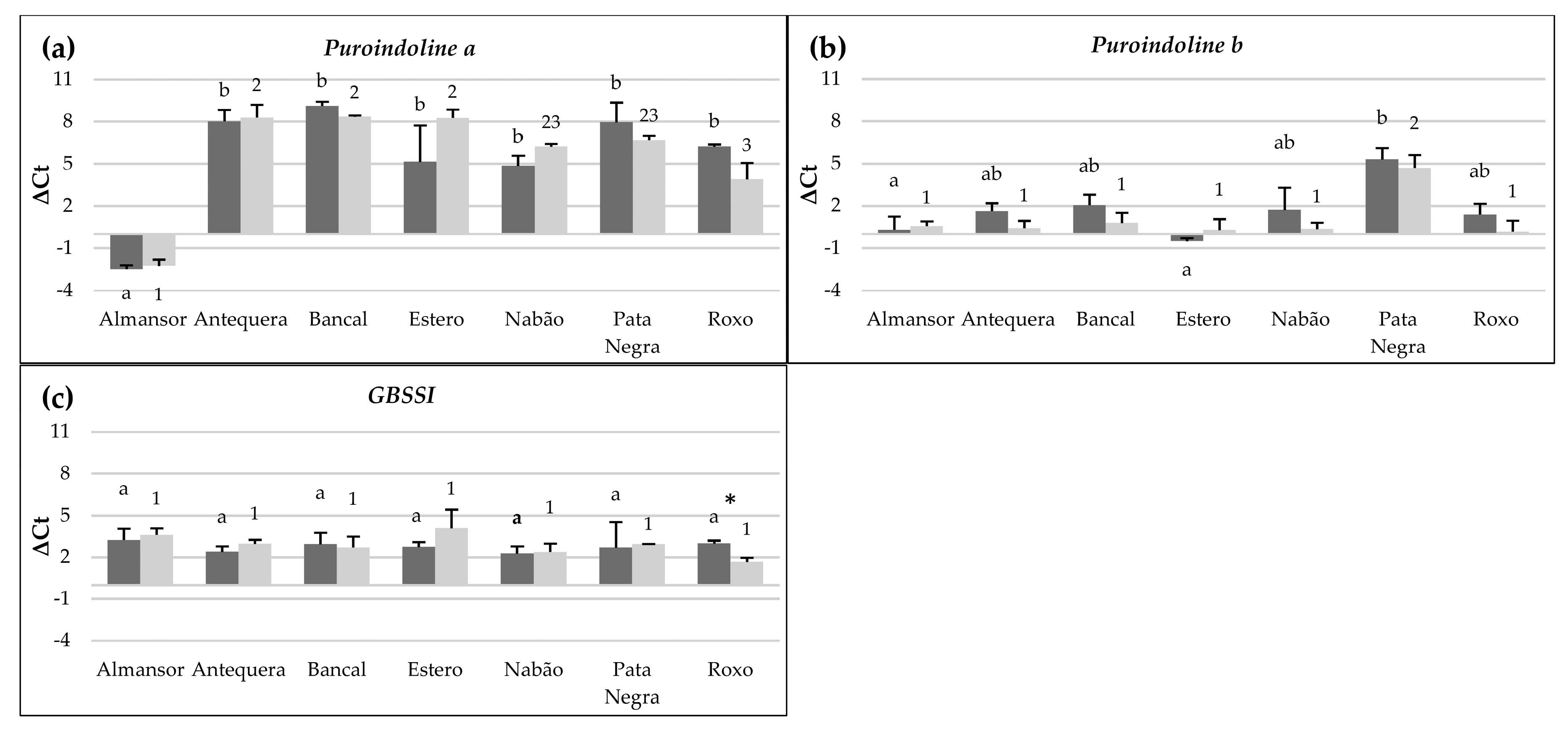
3.2. Transcription Levels of Flour Quality Related Genes in Immature Grains Vary between Wheat Varieties
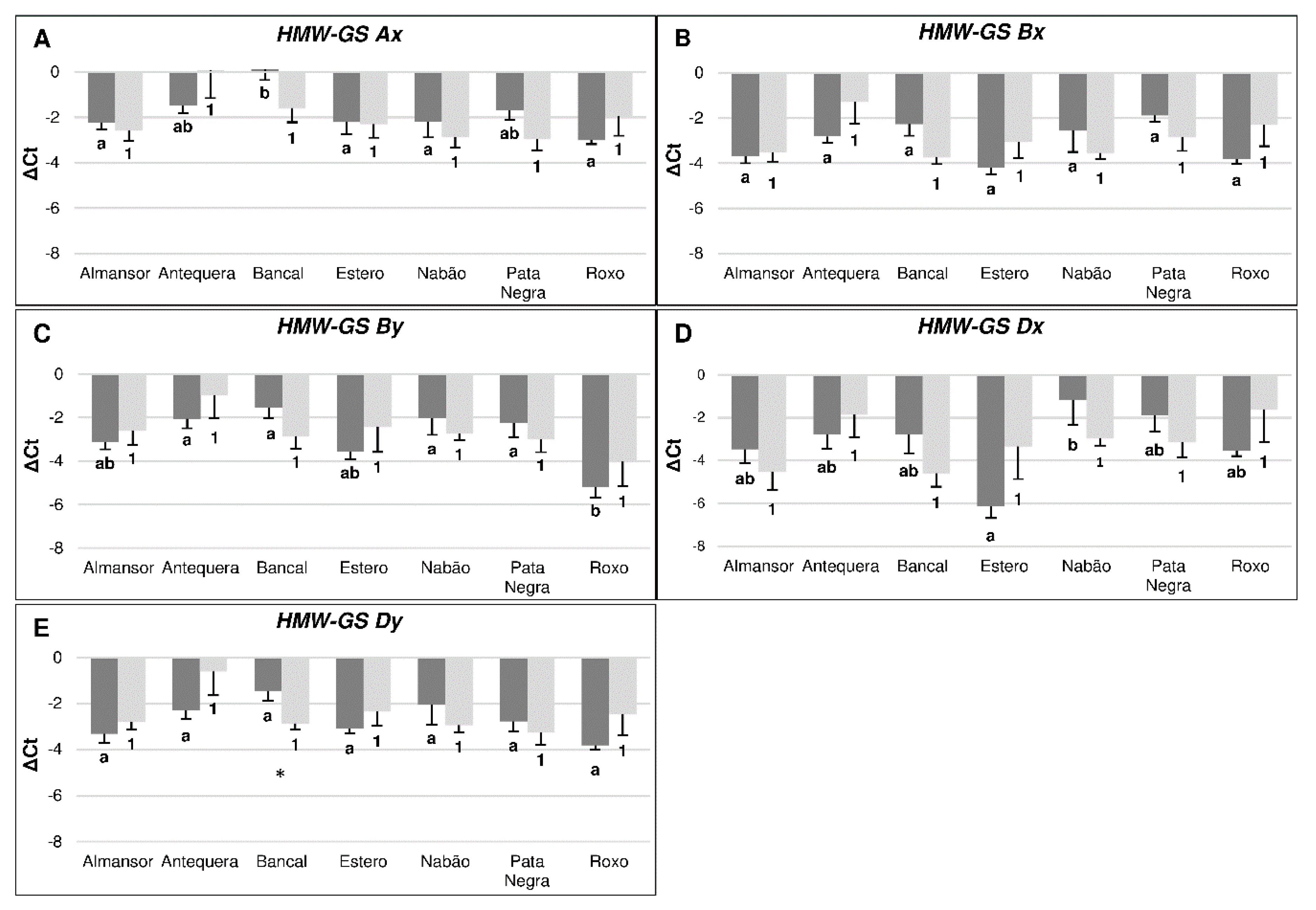
3.3. High Temperature Differentially Affects the Transcription of Flour Quality Related Genes in Immature Grains
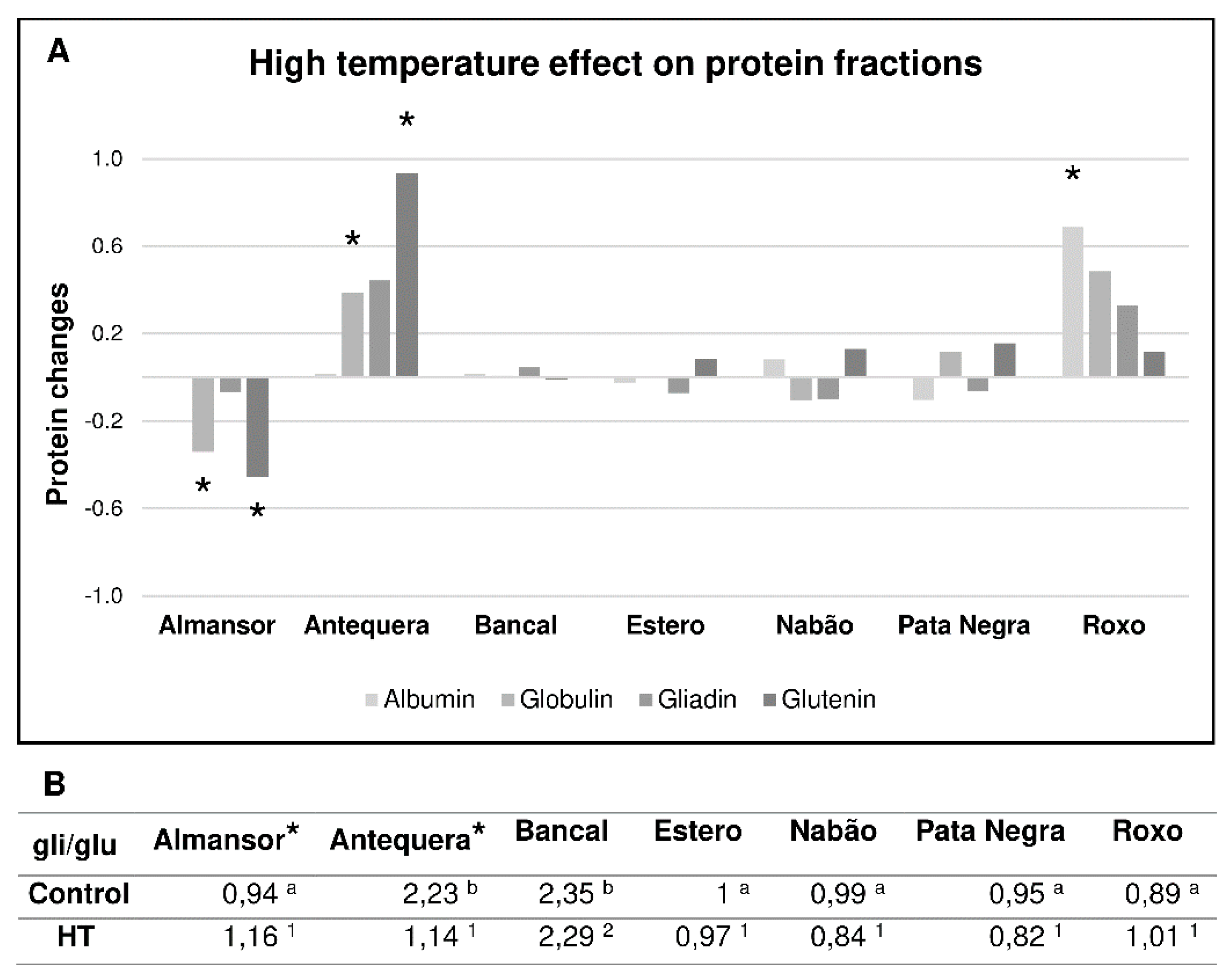
3.4. Comparative Contents of Protein Fractions in Mature Grains Vary between Varieties and with HT Treatment
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- FAO. The Cereal Supply and Demand Brief. Available online: http://www.fao.org/worldfoodsituation/csdb/en/ (accessed on 1 January 2020).
- Žilić, S.; Barać, M.; Pešić, M.; Dodig, D.; Ignjatović-Micić, D. Characterization of proteins from grain of different bread and durum wheat genotypes. Int. J. Mol. Sci. 2011, 12, 5878–5894. [Google Scholar] [CrossRef] [PubMed]
- Branlard, G.; Dardevet, M.; Saccomano, R.; Lagoutte, F.; Gourdon, J. Genetic diversity of wheat storage proteins and bread wheat quality. Euphytica 2001, 119, 59–67. [Google Scholar] [CrossRef]
- Guzmán, C.; Alvarez, J.B. Wheat waxy proteins: polymorphism, molecular characterization and effects on starch properties. Theor. Appl. Genet. 2016, 129, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Morris, C.F. Puroindolines: The molecular genetic basis of wheat grain hardness. Plant Mol. Biol. 2002, 48, 633–647. [Google Scholar] [CrossRef] [PubMed]
- Semenov, M.A.; Shewry, P.R. Modelling predicts that heat stress, not drought, will increase vulnerability of wheat in Europe. Sci. Rep. 2011, 1, 66. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, M. Stress-induced changes in wheat grain composition and quality. Crit. Rev. Food Sci. Nutr. 2014, 54, 1576–1583. [Google Scholar] [CrossRef] [PubMed]
- Blumenthal, C.S.; Barlow, E.W.R.; Wrigley, C.W. Growth Environment and Wheat Quality: the Effect of Heat Stress on Dough Properties and Gluten Proteins. J. Cereal Sci. 1993, 18, 3–21. [Google Scholar] [CrossRef]
- Altenbach, S.B.; Kothari, K.M.; Lieu, D. Environmental Conditions During Wheat Grain Development Alter Temporal Regulation of Major Gluten Protein Genes. Cereal Chem. 2002, 79, 279–285. [Google Scholar] [CrossRef]
- Hurkman, W.J.; McCue, K.F.; Altenbach, S.B.; Korn, A.; Tanaka, C.K.; Kothari, K.M.; Johnson, E.L.; Bechtel, D.B.; Wilson, J.D.; Anderson, O.D.; et al. Effect of temperature on expression of genes encoding enzymes for starch biosynthesis in developing wheat endosperm. Plant Sci. 2003, 164, 873–881. [Google Scholar] [CrossRef]
- DuPont, F.M.; Hurkman, W.J.; Vensel, W.H.; Chan, R.; Lopez, R.; Tanaka, C.; Altenbach, S.B. Differential accumulation of sulfur-rich and sulfur-poor wheat flour proteins is affected by temperature and mineral nutrition during grain development. J. Cereal Sci. 2006, 44, 101–112. [Google Scholar] [CrossRef]
- DuPont, F.M.; Hurkman, W.J.; Vensel, W.H.; Tanaka, C.; Kothari, K.M.; Chung, O.K.; Altenbach, S.B. Protein accumulation and composition in wheat grains: Effects of mineral nutrients and high temperature. Eur. J. Agron. 2006, 25, 96–107. [Google Scholar] [CrossRef]
- Zhang, Y.; Pan, J.; Huang, X.; Guo, D.; Lou, H.; Hou, Z.; Su, M.; Liang, R.; Xie, C.; You, M.; et al. Differential effects of a post-anthesis heat stress on wheat (Triticum aestivum L.) grain proteome determined by iTRAQ. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Jørgensen, A.D.; Li, H.; Søndergaard, I.; Finnie, C.; Svensson, B.; Jiang, D.; Wollenweber, B.; Jacobsen, S. Implications of high-temperature events and water deficits on protein profiles in wheat (Triticum aestivum L. cv. Vinjett) grain. Proteomics 2011, 11, 1684–1695. [Google Scholar] [CrossRef] [PubMed]
- World Meteorological Organization. Guidelines on the definition and monitoring of extreme weather and climate events-Draft Version-first review by TT-DEWCE; Geneva, Switzerland, 2015. [Google Scholar]
- Cardoso, R.M.; Soares, P.M.M.; Lima, D.C.A.; Miranda, P.M.A. Mean and extreme temperatures in a warming climate: EURO CORDEX and WRF regional climate high-resolution projections for Portugal. Clim. Dyn. 2019, 52, 129–157. [Google Scholar] [CrossRef]
- ANPOC.; INIAV.; IpBeja; Ceres; Germen; Cerealis. Lista de Variedade Recomendadas de Trigo Mole; Lisboa, Portugal, 2014. [Google Scholar]
- Paolacci, A.R.; Tanzarella, O.A.; Porceddu, E.; Ciaffi, M. Identification and validation of reference genes for quantitative RT-PCR normalization in wheat. BMC Mol. Biol. 2009, 10, 11. [Google Scholar] [CrossRef] [PubMed]
- Lookhart, G.; Bean, S.R. Separation and Characterization of Wheat Protein Fractions. Cereal Chem. 1995, 72, 527–532. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Payne, P.I.; Nightingale, M.A.; Krattiger, A.F.; Holt, L.M. The relationship between HMW glutenin subunit composition and the bread making quality of British grown wheat varieties. J. Sci. Food Agric. 1987, 40, 51–65. [Google Scholar] [CrossRef]
- Rasheed, A.; Xia, X.; Yan, Y.; Appels, R.; Mahmood, T.; He, Z. Wheat seed storage proteins: Advances in molecular genetics, diversity and breeding applications. J. Cereal Sci. 2014, 60, 11–24. [Google Scholar] [CrossRef]
- Nakamura, T.; Vrinten, P.; Saito, M.; Konda, M. Rapid classification of partial waxy wheats using PCR-based markers. Genome 2002, 45, 1150–1156. [Google Scholar] [CrossRef]
- Bowden, P.; Edwards, J.; Fergson, N.; McNee, T.; Manning, B.; Raoberts, K.; Schipp, A.; Schulze, K.; Wilkins, J. Wheat Growth & Development; White, J., Edwards, J., Eds.; NSW Department of Primary Industries: Orange, Australia, 2007. [Google Scholar]
- Igrejas, G.; Gaborit, T.; Oury, F.-X.; Chiron, H.; Marion, D.; Branlard, G. Genetic and Environmental Effects on Puroindoline-a and Puroindoline-b Content and their Relationship to Technological Properties in French Bread Wheats. J. Cereal Sci. 2001, 34, 37–47. [Google Scholar] [CrossRef]
- Farooq, M.; Bramley, H.; Palta, J.a.; Siddique, K.H.M. Heat Stress in Wheat during Reproductive and Grain-Filling Phases. CRC. Crit. Rev. Plant Sci. 2011, 30, 491–507. [Google Scholar] [CrossRef]
- Dhaka, V.; Khatkar, B.S. Effects of gliadin/glutenin and HMW-GS/LMW-GS ratio on dough rheological properties and breadmaking potential of wheat varieties. J. Food Qual. 2015, 38, 71–82. [Google Scholar] [CrossRef]
- Barak, S.; Mudgil, D.; Khatkar, B.S. Relationship of gliadin and glutenin proteins with dough rheology, flour pasting and bread making performance of wheat varieties. LWT-Food Sci. Technol. 2013, 51, 211–217. [Google Scholar] [CrossRef]
- Balla, K.; Rakszegi, M.; Li, Z.; Békés, F.; Bencze, S.; Veisz, O. Quality of winter wheat in relation to heat and drought shock after anthesis. Czech. J. Food Sci. 2011, 29, 117–128. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Almansor | Antequera | Bancal | Estero | Nabão | Pata Negra | Roxo | Chinese Spring | ||
|---|---|---|---|---|---|---|---|---|---|
| Glu-1 loci | Ax | 2* | 1 | 2* | 1 | 2* | 1 | 2* | Null |
| Bx + By | 17 + 18 | 7 + 8 | 7 + 9 | 17 + 18 | 7 + 8 | 17 + 18 | 7 + 8 | 7 + 8 | |
| Dx + Dy | 5 + 10 | 5 + 10 | 5 + 10 | 5 + 10 | 5 + 10 | 5 + 10 | 5 + 10 | 2 + 12 | |
| Glu-1 Score | 10 | 10 | 9 | 10 | 10 | 10 | 10 | 8 | |
| Waxy loci | Wx-A1 | WT | WT | WT | WT | WT | WT | Null | WT |
| Wx-B1 | Null | WT | WT | WT | WT | WT | WT | WT | |
| Wx-D1 | WT | WT | Null | WT | WT | Null | WT | WT | |
| Waxy type | Partial waxy | Wild Type | Partial waxy | Wild Type | Wild Type | Partial waxy | Partial waxy | Wild Type | |
| Hardness locus | Pina-D1a | + | - | - | - | - | - | - | + |
| Pinb-D1a | - | + | - | + | - | + | - | + | |
| Hardness | Hard | Hard | Hard | Hard | Hard | Hard | Hard | Soft | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tomás, D.; Viegas, W.; Silva, M. Effects of Post-Anthesis Heat Waves on the Grain Quality of Seven European Wheat Varieties. Agronomy 2020, 10, 268. https://doi.org/10.3390/agronomy10020268
Tomás D, Viegas W, Silva M. Effects of Post-Anthesis Heat Waves on the Grain Quality of Seven European Wheat Varieties. Agronomy. 2020; 10(2):268. https://doi.org/10.3390/agronomy10020268
Chicago/Turabian StyleTomás, Diana, Wanda Viegas, and Manuela Silva. 2020. "Effects of Post-Anthesis Heat Waves on the Grain Quality of Seven European Wheat Varieties" Agronomy 10, no. 2: 268. https://doi.org/10.3390/agronomy10020268
APA StyleTomás, D., Viegas, W., & Silva, M. (2020). Effects of Post-Anthesis Heat Waves on the Grain Quality of Seven European Wheat Varieties. Agronomy, 10(2), 268. https://doi.org/10.3390/agronomy10020268