Resistance to Anthracnose (Colletotrichum lentis, Race 0) in Lens spp. Germplasm





Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material and Growing Conditions
2.2. Fungal Isolates
2.3. Fungal Inoculation and Incubation
2.4. Disease Evaluation and Statistical Analysis
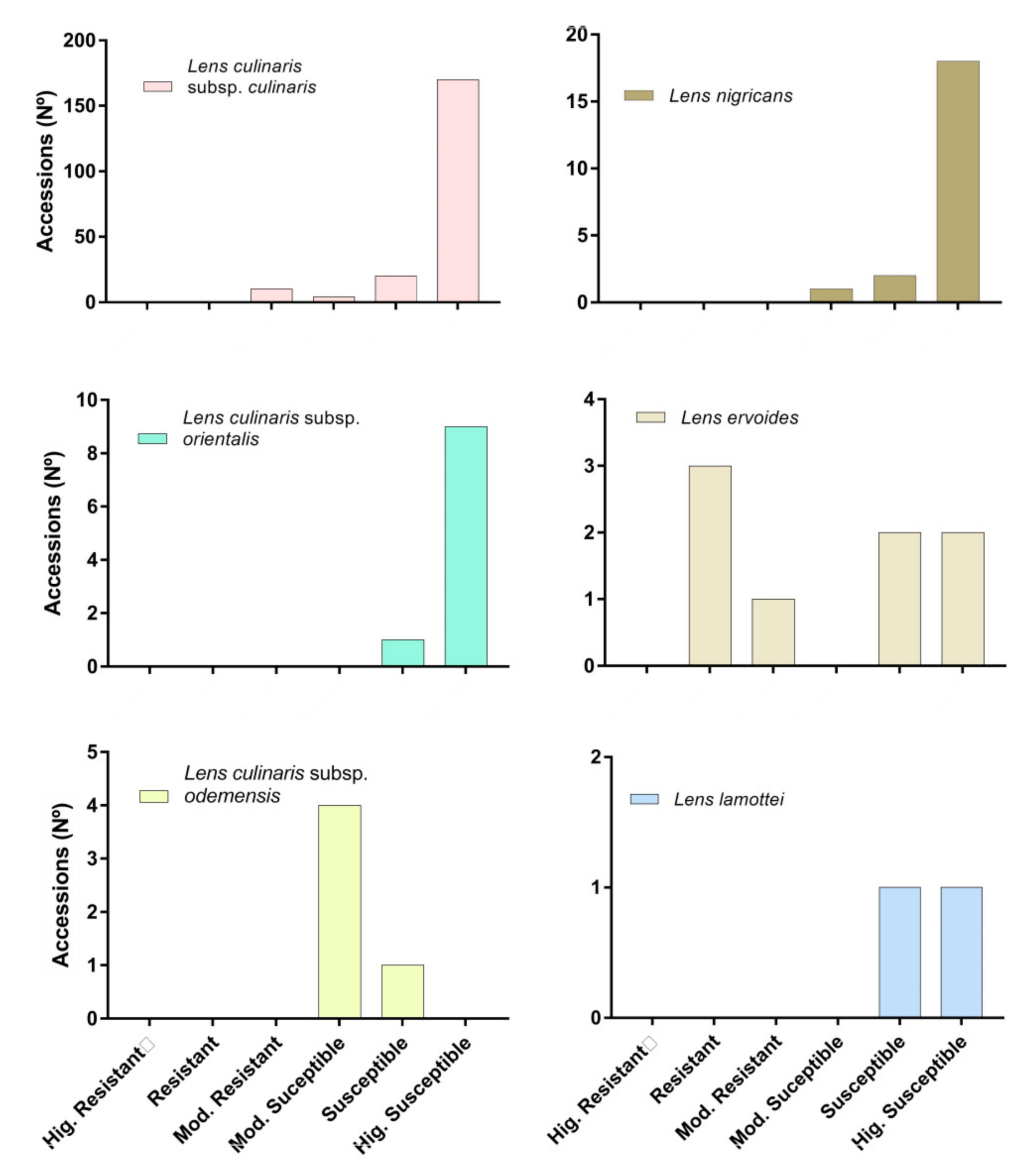
3. Results and Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- FAOSTAT-Food and Agriculture Organization. Available online: http://www.fao.org/faostat/en/#home (accessed on 20 September 2020).
- Chen, W.; Sharma, H.C.; Muehlbauer, F.J. Compendium of Chickpea and Lentil Diseases and Pests; APS Press: St. Paul, MN, USA, 2011; pp. 6–98. [Google Scholar] [CrossRef]
- Gossen, B.D.; Anderson, K.L.; Buchwaldt, L. Host specificity to Colletotrichum truncatum from lentil. J. Plant Pathol. 2009, 31, 65–73. [Google Scholar] [CrossRef]
- Bhadauria, V.; Banniza, S.; Vandenberg, A.; Selvaraj, G.; Wei, Y. Cataloging proteins putatively secreted during the biotrophy-necrotrophy transition of the anthracnose pathogen Colletotrichum truncatum. Plant Signal. Behav. 2011, 6, 1457–1459. [Google Scholar] [CrossRef]
- Damm, U.; O’Connell, R.J.; Groenewald, J.Z.; Crous, P.W. The Colletotrichum destructivum species complex—Hemibiotrophic pathogens of forage and field crops. Stud. Mycol. 2014, 79, 49–84. [Google Scholar] [CrossRef] [PubMed]
- Buchwaldt, L.; Dzanovic, E.; Durkin, J. Lentil anthracnose: Epidemiology, fungicide decision support system, resistance and pathogen races. Can. J. Plant Pathol. 2018, 40, 189–198. [Google Scholar] [CrossRef]
- Banniza, S.; Warale, R.; Menat, J.; Cohen-Skali, A.; Armstrong-Cho, C.; Bhadauria, V. The long path to understanding the host-pathogen interactions of Colletotrichum lentis on lentil. Can. J. Plant Pathol. 2018, 40, 199–209. [Google Scholar] [CrossRef]
- Chongo, G.; Gossen, B.D.; Bernier, C.C. Infection by Colletotrichum truncatum in resistant and susceptible lentil genotypes. Can. J. Plant Pathol. 2002, 24, 81–85. [Google Scholar] [CrossRef]
- Shaikh, R.; Diederichsen, A.; Harrington, M.; Adam, J.; Conner, R.L.; Buchwaldt, L. New sources of resistance to Colletotrichum truncatum race Ct0 in Lens culinaris Medikus subsp. culinaris obtained by single plant selection in germplasm accessions. Genet. Resour. Crop Evol. 2013, 60, 193–201. [Google Scholar] [CrossRef]
- Chongo, G.; Bernier, C.C. Effects of host, inoculum concentration, wetness duration, growth stage and temperature on anthracnose of lentil. Plant Dis. 2000, 84, 544–548. [Google Scholar] [CrossRef]
- Buchwaldt, L.; Anderson, K.L.; Morrall, R.A.A.; Gossen, B.D.; Bernier, C.C. Identification of lentil germ plasm resistant to Colletotrichum truncatum and characterization of two pathogen races. Phytopathology 2004, 94, 236–243. [Google Scholar] [CrossRef]
- Armstrong-Cho, C.; Wang, J.; Wei, Y.; Banniza, S. The infection process of two pathogenic races of Colletotrichum truncatum on lentil. Can. J. Plant Pathol. 2012, 34, 58–67. [Google Scholar] [CrossRef]
- Vail, S.; Strelioff, J.V.; Tullu, A.; Vandenberg, A. Field evaluation of resistance to Colletotrichum truncatum in Lens culinaris, Lens ervoides and Lens ervoides × Lens culinaris derivatives. Field Crops Res. 2012, 126, 145–151. [Google Scholar] [CrossRef]
- Buchwaldt, L.; Shaikh, R.; Adam, J.; Tullu, A.; Slinkard, A.E. Recessive and dominant genes confer resistance to Colletotrichum truncatum in cultivated lentil. Can. J. Plant Pathol. 2013, 35, 222–231. [Google Scholar] [CrossRef]
- Tullu, A.; Buchwaldt, L.; Lulsdorf, M.; Banniza, S.; Barlow, B.; Slinkard, A.E.; Sarker, A.; Tar’an, B.; Warkentin, T.; Vandenberg, A. Sources of resistance to anthracnose (Colletotrichum truncatum) in wild lens species. Genet. Resour. Crop Evol. 2006, 53, 111–119. [Google Scholar] [CrossRef]
- Ford, R.; Banniza, S.; Photita, W.; Taylor, P.W.J. Morphological and molecular discrimination of Colletotrichum truncatum causing anthracnose on lentil in Canada. Australas. Plant Pathol. 2004, 33, 559–569. [Google Scholar] [CrossRef]
- Jeger, M.J.; Viljanen-Rollinson, S.L. The use of the area under the disease-progress curve (AUDPC) to assess quantitative disease resistance in crop cultivars. Theor. Appl. Genet. 2001, 102, 32–40. [Google Scholar] [CrossRef]
- Madden, L.V.; Hughes, G.; van den Bosch, F. The Study of Plant Disease Epidemics; APS Press: St. Paul, MN, USA, 2007; pp. 11–31. [Google Scholar] [CrossRef]
- Moral, J.; Xaviér, C.J.; Viruega, J.R.; Roca, L.F.; Caballero, J.; Trapero, A. Variability in susceptibility to anthracnose in the world collection of olive cultivars of Cordoba (Spain). Front. Plant Sci. 2017, 8, 1892. [Google Scholar] [CrossRef] [PubMed]
- Barulina, H. Lentils of the USSR and other countries. Bull. Appl. Genet. Plant Breed. 1930, 40, 1–319. [Google Scholar]
- Coyne, C.; McGee, R. Lentil. Genetic and Genomic Resources of Grain Legume Improvement; Elsevier: Amsterdam, The Netherlands, 2013; pp. 157–180. [Google Scholar] [CrossRef]
- Buchwaldt, L.; Diederichsen, A. New disease resistant lentil germplasm identified at Plant Gene Resources of Canada (PGRC). In Proceedings of the 5th Canadian Pulse Research Workshop, London, ON, Canada, 28–30 November 2004; p. 204. [Google Scholar]
- Vail, S.; Vandenberg, A. Evaluation of a clonal propagation protocol to obtain replicated disease data on infection by Colletotrichum truncatum in Lens culinaris. Crop Sci. 2010, 50, 926–932. [Google Scholar] [CrossRef]
- Gela, T.S.; Banniza, S.; Vandenberg, A. Lack of effective resistance to the virulent race of Colletotrichum lentis in Lens culinaris Medikus subsp. culinaris. Plant Gen. Res. 2020, 18, 81–87. [Google Scholar] [CrossRef]
- Fiala, J.V.; Tullu, A.; Banniza, S.; Séguin-Swartz, G.; Vandenberg, A. Interspecies transfer of resistance to anthracnose in lentil (Lens culinaris Medic.). Crop Sci. 2009, 49, 825–830. [Google Scholar] [CrossRef]
- Tullu, A.; Bett, K.E.; Banniza, S.; Vail, S.; Vandenberg, A. Widening the genetic base of cultivated lentil through hybridization of Lens culinaris ‘Eston’ and L. ervoides accession IG 72815. Can. J. Plant Sci. 2013, 93, 1037–1047. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Country of Origin | Nº of Accessions | Accessions Resistant (R) | Accessions Moderately Resistant (MR) |
|---|---|---|---|
| Afghanistan | 1 | - | - |
| Algeria | 2 | - | PI297287 |
| Bosnia | 1 | - | - |
| Brazil | 1 | - | - |
| Bulgaria | 1 | - | - |
| Canada | 1 | - | - |
| Chile | 7 | - | - |
| Czech Republic | 2 | - | - |
| Egypt | 3 | - | - |
| Ethiopia | 2 | - | - |
| France | 4 | - | - |
| Germany | 2 | - | - |
| Guatemala | 1 | - | - |
| India | 10 | - | - |
| Iran | 43 | - | PI431809, PI432005, PI432033, PI432071 |
| Israel | 4 | PI572330 | - |
| Italy | 3 | - | - |
| Jordan | 3 | - | PI477921 |
| Lebanon | 1 | - | - |
| Mexico | 4 | - | - |
| Montenegro | 1 | - | - |
| Morocco | 3 | - | - |
| North Macedonia | 2 | - | - |
| Pakistan | 4 | - | - |
| Peru | 1 | - | - |
| Portugal | 1 | - | PI283604 |
| Republic of Cyprus | 3 | - | - |
| Russia | 2 | - | - |
| Serbia | 5 | - | PI572327 |
| Syria | 5 | - | - |
| Spain | 88 | - | BGE001814, PI298644 |
| State of Palestine | 1 | - | - |
| Tajikistan | 1 | - | - |
| Turkey | 21 | PI572334, PI572338 | - |
| Ukraine | 1 | - | - |
| United States | 1 | - | - |
| Unknown | 13 | - | W627756 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barilli, E.; Moral, J.; Aznar-Fernández, T.; Rubiales, D. Resistance to Anthracnose (Colletotrichum lentis, Race 0) in Lens spp. Germplasm. Agronomy 2020, 10, 1799. https://doi.org/10.3390/agronomy10111799
Barilli E, Moral J, Aznar-Fernández T, Rubiales D. Resistance to Anthracnose (Colletotrichum lentis, Race 0) in Lens spp. Germplasm. Agronomy. 2020; 10(11):1799. https://doi.org/10.3390/agronomy10111799
Chicago/Turabian StyleBarilli, Eleonora, Juan Moral, Thaïs Aznar-Fernández, and Diego Rubiales. 2020. "Resistance to Anthracnose (Colletotrichum lentis, Race 0) in Lens spp. Germplasm" Agronomy 10, no. 11: 1799. https://doi.org/10.3390/agronomy10111799
APA StyleBarilli, E., Moral, J., Aznar-Fernández, T., & Rubiales, D. (2020). Resistance to Anthracnose (Colletotrichum lentis, Race 0) in Lens spp. Germplasm. Agronomy, 10(11), 1799. https://doi.org/10.3390/agronomy10111799