Morphological and Molecular Status of Daphne wolongensis C.D.Brickell et B.Mathew as Genetic Resource for Horticulture


Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling
2.2. Molecular Analysis
2.3. Data Analysis
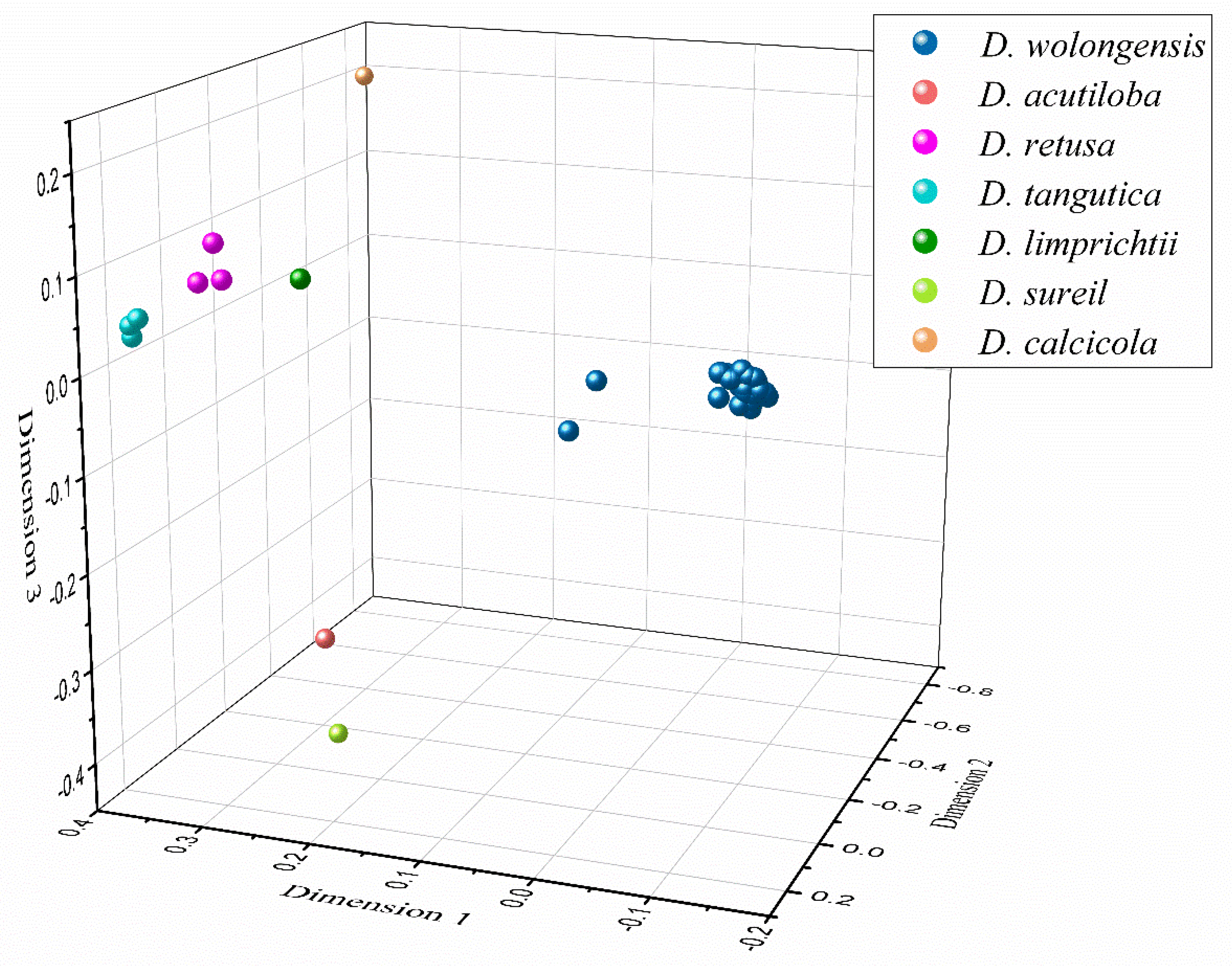
3. Results
3.1. Distribution and Habitat of D. wolongensis
3.2. Morphological Diversity of D. wolongensis
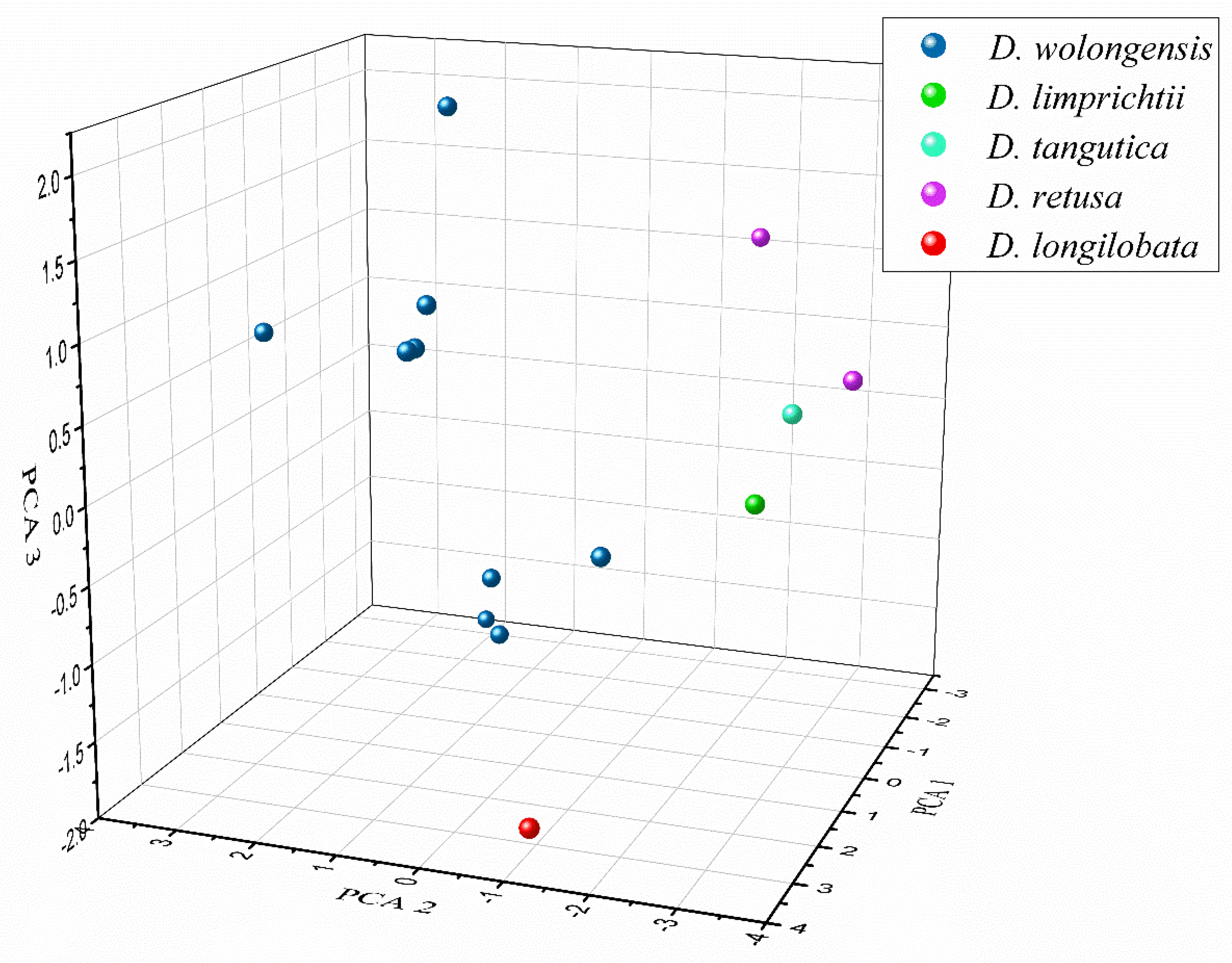
3.3. Differences among Related Species
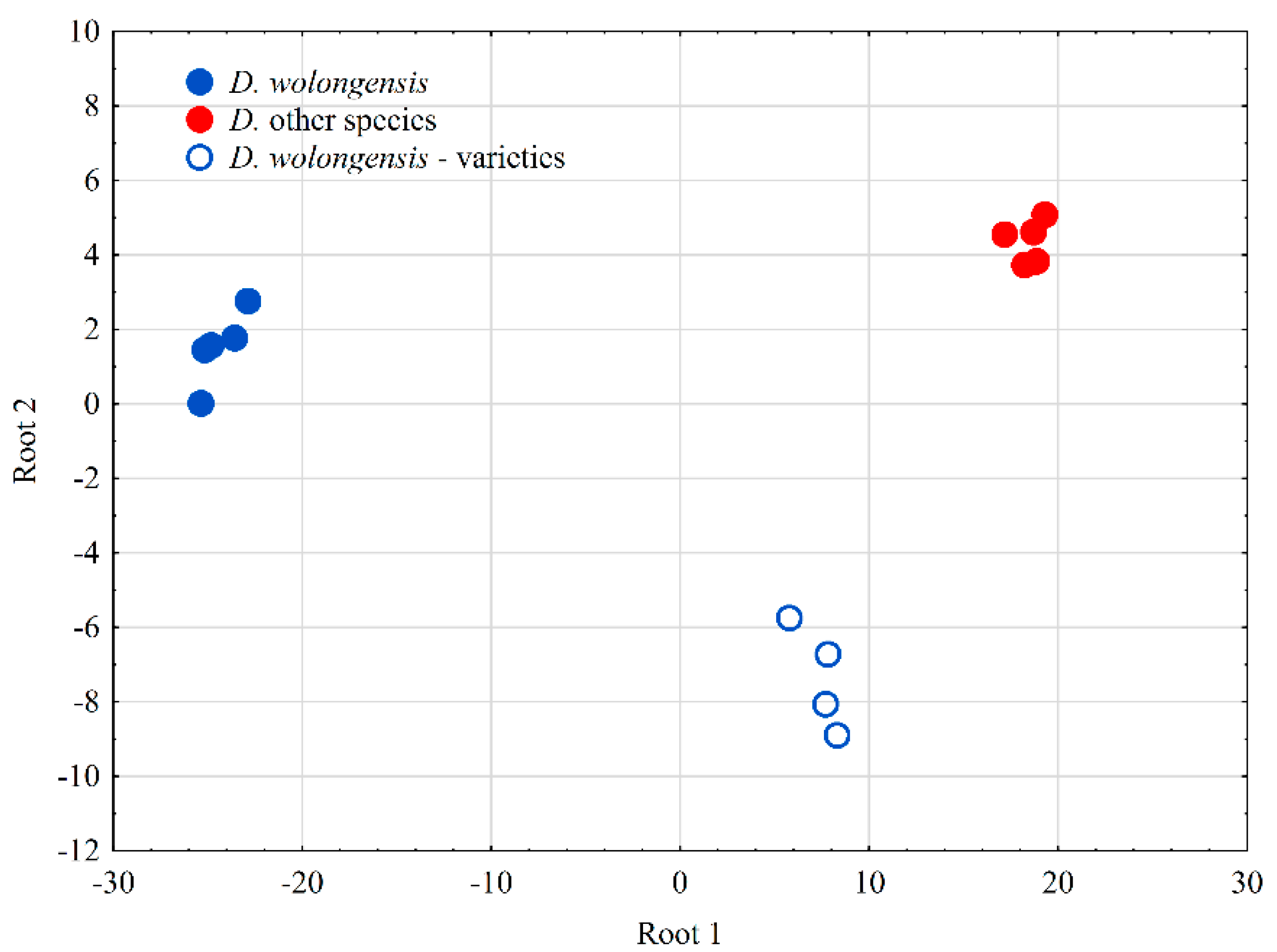
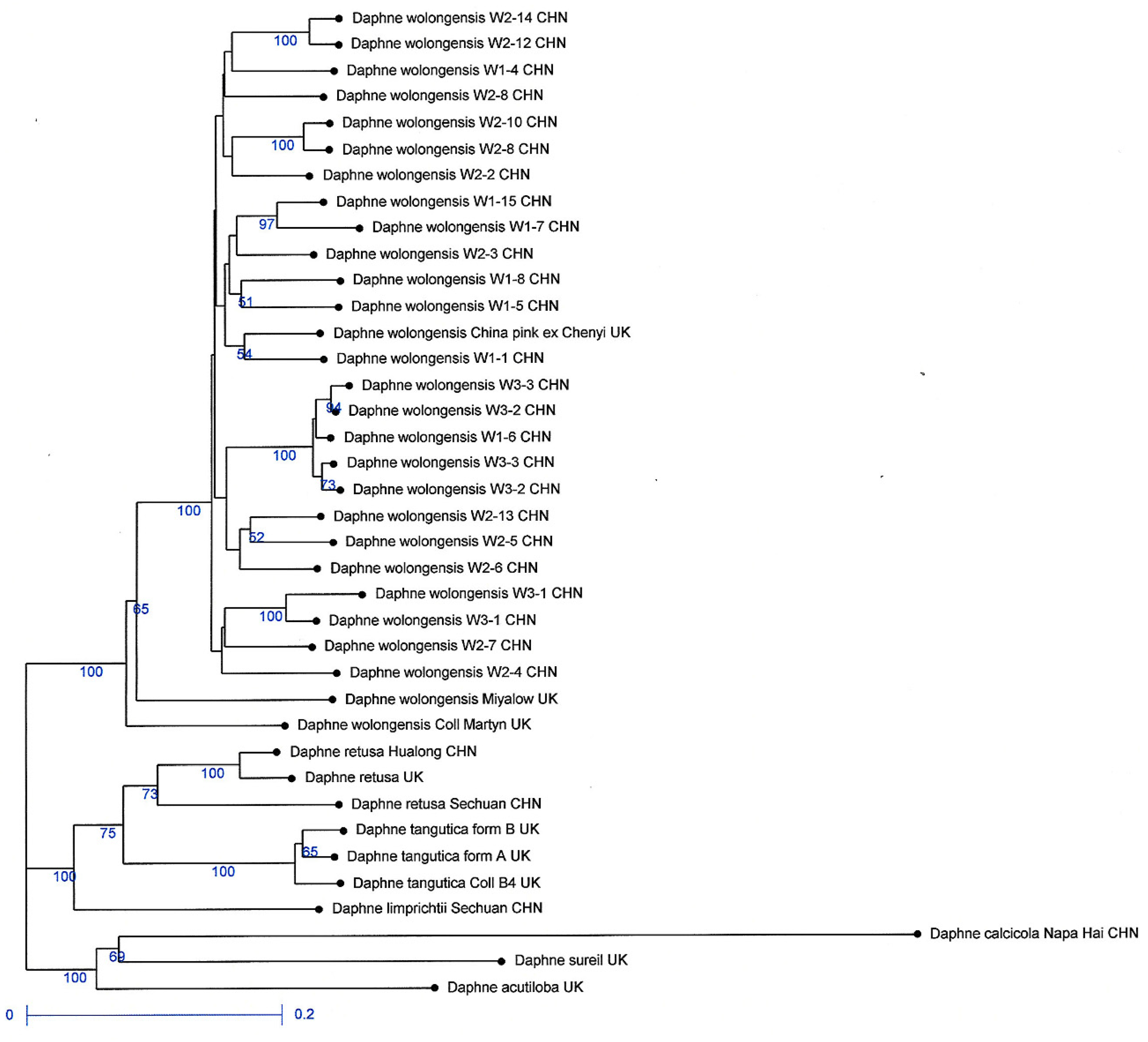
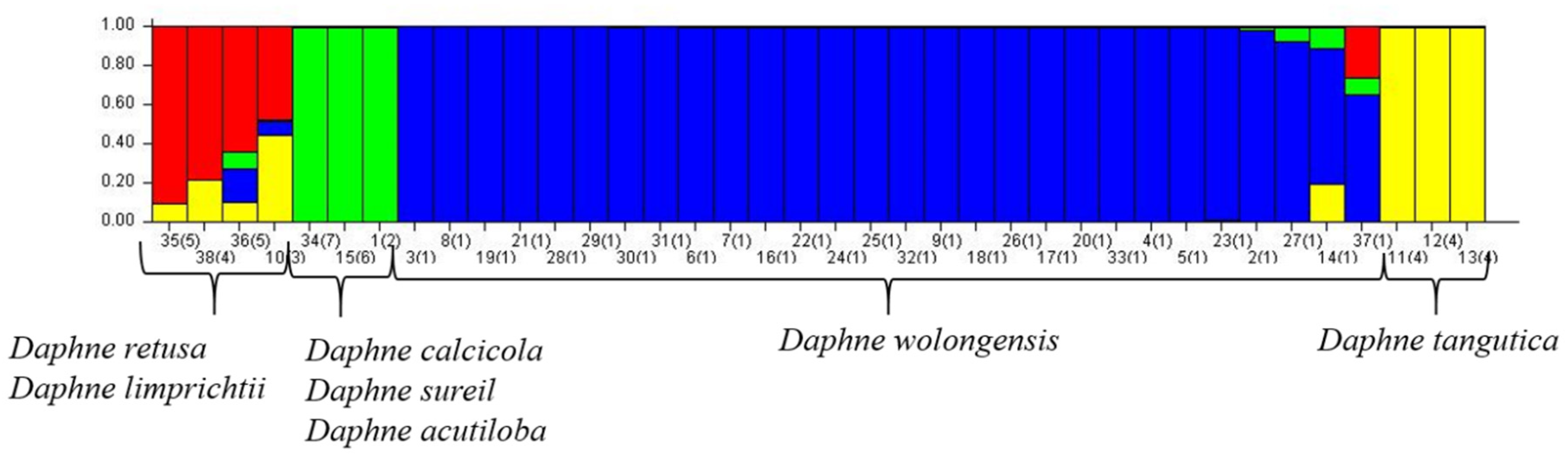
3.4. Genetic Diversity of D. wolongensis and Related Species
4. Discussion
4.1. Status and Diversity of D. wolongensis
4.2. Conservation Status
4.3. Impact for Horticulture
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hong, S.P. Daphne L. In The Genera of Vascular Plants of Korea; Park, C.W., Ed.; Academy Publishing Co.: Seoul, Korea, 2007; pp. 633–634. [Google Scholar]
- Wang, Y.; Gilbert, M.G.; Mathew, B.F.; Eckell, B. Daphne L. In Flora of China; Wu, Z.Y., Raven, P.H., Hong, D.Y., Eds.; Science Press: Beijing, China; Missouri Botanical Garden Press: St. Louis, MI, USA, 2007; Volume 13, pp. 230–245. [Google Scholar]
- Flora of China Home Page. Available online: http://flora.huh.harvard.edu/china/ (accessed on 30 July 2020).
- Webb, D.A. Daphne, L. In Flora Europaea; Tutin, T.G., Heywood, V.H., Burges, N.A., Moore, D.M., Valentine, D.H., Walters, S.M., Webb, D.A., Eds.; Cambridge University Press: Cambridge, UK, 1978; Volume 2, pp. 256–258. [Google Scholar]
- Brickell, C.D.; Mathew, B. Daphne, the Genus in the Wild & in Cultivation; Alpine Garden Society: Woking, UK, 1976; p. 194. [Google Scholar]
- Keissler, K. Die Arten der Gattung Daphne aus der Section Daphnanthes. Bot. Jahrbucher 1898, 25, 29–124. [Google Scholar]
- Halda, J.J. Lýkovce Daphne Skalky-Skalničky; Klub Skalničkářů: Brno, Czech Republic, 1976; pp. 11–68. [Google Scholar]
- White, R. Daphnes: A Practical Guide for Gardeners; Timber Press: Portland, OR, USA, 2006; p. 224. [Google Scholar]
- Halda, J.J. The Genus Daphne; Sen: Dobré, Czech Republic, 2001; p. 231. [Google Scholar]
- Rix, M. Plants from Panda country. Country Life J. 1992, 42–43. [Google Scholar]
- White, R. (Blackthorn Nursery, Leighton Buzzard, UK). Personal communication, 2011.
- Brickell, C.D.; Mathew, B. 575. Daphne wolongensis. Bot. Mag. 2007, 24, 18–21, (plate 575, photo). [Google Scholar]
- Rehder, A. Daphne wilsonii. In Plantae Wilsonianae: An Enumeration of the Woody Plants Collected in Western China for the Arnold Arboretum of Harvard University during the Years 1907, 1908, and 1910; Sargent, C.S., Ed.; The University Press: Cambridge, UK, 1916; Volume 2, pp. 540–541. [Google Scholar]
- IUCN Standards and Petitions Subcommittee 2014. Guidelines for Using the IUCN Red List Categories and Criteria, Version 11. Available online: www.iucnredlist.org/documents/RedListGuidelines.pdf (accessed on 30 July 2020).
- Holubec, V.; Smekalova, T.; Leišová-Svobodová, L. Morphological and molecular evaluation of the Far East fruit genetic resources of Lonicera caerulea L.—Vegetation, ethnobotany, use and conservation. Genet. Resour. Crop. Evol. 2018, 66, 121–141. [Google Scholar] [CrossRef]
- Vos, P.; Hogers, R.; Bleeker, M.; Reijans, M.; Van De Lee, T.; Hornes, M.; Friters, A.; Pot, J.; Paleman, J.; Kuiper, M.; et al. AFLP: A new technique for DNA fingerprinting. Nucleic Acids Res. 1995, 23, 4407–4414. [Google Scholar] [CrossRef]
- Perrier, X.; Jacquemoud-Collet, J.P. DARwin Software Version 6.0. Available online: http://darwin.cirad.fr/darwin (accessed on 30 July 2020).
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar]
- Falush, D.; Stephens, M.; Pritchard, J.K. Inference of population structure using multilocus genotype data: Dominant markers and null alleles. Mol. Ecol. Notes 2007, 7, 574–578. [Google Scholar] [CrossRef]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of cluster of individuals using the software Structure: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef]
- Nei, M. Analysis of gene diversity in subdivided populations. Proc. Natl. Acad. Sci. USA 1973, 70, 3321–3323. [Google Scholar] [CrossRef]
- Lewontin, R.C. The Apportionment of Human Diversity. Evol. Biol. 1972, 6, 381–398. [Google Scholar] [CrossRef]
- Shannon, C.E. and Weaver, W. The Mathematical Theory of Communication; The University of Illinois Press: Champaign, IL, USA, 1949. [Google Scholar]
- Yeh, F.C.; Boyle, T.; Yang, R.-C.; Boyle, T. Popgene Version 1.31. Microsoft Window-Based Freeware for Population Genetic Analysis; University of Alberta: Edmonton, AB, Canada. Available online: https://sites.ualberta.ca/~fyeh/popgene.pdf (accessed on 2 October 2017).
- Miller, M.P. Tools for Population Genetic Analysis (TFPGA), Version 1.3; Northern Arizona University: Tempe, AZ, USA, 1997; p. 30. [Google Scholar]
- Nei, M. Estimation of average heterozygosity and genetic distance from a small number of individuals. Genetics 1978, 89, 583–590. [Google Scholar] [PubMed]
- Rogers, Z.S. A World Checklist of Thymelaeaceae; Missouri Botanical Garden: St. Louis, MO, USA, 2009; Last Modified 2012. Tropicos Home Page; Available online: http://www.tropicos.org/Project/Thymelaeaceae (accessed on 30 September 2020).
- Ghosh, J.; Midday, M.; Dey, S.K.; Maity, D. Daphne thanguensis sp. nov. (Thymelaeaceae) from Sikkim Himalaya. Nord. J. Bot. 2018, 36, 1–5. [Google Scholar] [CrossRef]
- Birks, H.; Willis, K.J. Alpines, trees, and refugia in Europe. Plant Ecol. Divers. 2008, 1, 147–160. [Google Scholar] [CrossRef]
- Hengeveld, R.; Haeck, J. The distribution of the abundance I. Measurements. J. Biogeogr. 1982, 9, 303–316. [Google Scholar] [CrossRef]
- Vucetich, J.A.; Waite, T.A. Spatial patterns of demography and genetic processes across the species’ range: Null hypotheses for landscape conservation genetics. Conserv. Genet. 2003, 4, 639–645. [Google Scholar] [CrossRef]
- Alleaume-Benharira, M.; Pen, I.R.; Ronce, O. Geographical patterns of adaptation within a species’ range: Interactions between drift and gene flow. J. Evol. Biol. 2006, 19, 203–215. [Google Scholar] [CrossRef] [PubMed]
- Gaudeul, M.; Taberlet, P.; Till-Bottraud, I. Genetic diversity in an endangered alpine plant, Eryngium alpinum L. (Apiaceae), inferred from amplified fragment length polymorphism markers. Mol. Ecol. 2000, 9, 1625–1637. [Google Scholar] [CrossRef] [PubMed]
- Eckstein, R.L.; O’Neill, R.A.; Danihelka, J.; Otte, A.; Köhler, W. Genetic structure among and within peripheral and central populations of three endangered floodplain violets. Mol. Ecol. 2006, 15, 2367–2379. [Google Scholar] [CrossRef] [PubMed]
- Medrano, M.; Herrera, C.M. Geographical Structuring of Genetic Diversity Across the Whole Distribution Whitege of Narcissus longispathus, a Habitat-specialist, Mediterranean Narrow Endemic. Ann. Bot. 2008, 102, 183–194. [Google Scholar] [CrossRef]
- Castilla, A.R.; Alonso, C.; Herrera, C.M. Genetic structure of the shrub Daphne laureola across the Baetic Ranges, a Mediterranean glacial refugium and biodiversity hotspot. Plant Biol. 2011, 14, 515–524. [Google Scholar] [CrossRef]
- Thompson, J.D. Plant evolution in the Mediterranean, 1st ed.; Oxford University Press: Oxford, UK, 2005; pp. 77–80. [Google Scholar]
- Médail, F.; Diadema, K. Glacial refugia influence plant diversity patterns in the Mediterranean. Basin. J. Biogeogr. 2009, 36, 1333–1345. [Google Scholar] [CrossRef]
- Rankin, D. (University of Edinburgh, Edinburgh, UK). Personal communication, 2007.
- Heywood, V. Monitoring of areas and species /populations to assess effectiveness of conservation /managementactions. In Crop Wild Relatives. A Manual of In Situ Conservation; Hunter, D., Heywood, V., Eds.; Earthscan: London, UK; Washington, DC, USA, 2011; pp. 295–313. [Google Scholar]
- Iriondo, J.M.; Maxted, N.; Duloo, M.E. Conserving Plant Genetic Diversity in Protected Areas; CAB International: Wallingford, UK, 2008; p. 212. [Google Scholar]
- Joshi, S.P.; Ranjekar, P.K.; Gupta, V.S. Molecular markers in plant genome analysis. Curr. Sci. 1999, 77, 230–240. [Google Scholar]
- Zeven, A.C.; de Wet, J.M.J. Dictionary of Cultivated Plants and Their Regions of Diversity; Pudoc: Wageningen, The Netherlands, 1982; 259p. [Google Scholar]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; Da Fonseca, G.A.B.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Locality/* Source of Living Material | Altitude | Habitat Description | No. of Samples |
|---|---|---|---|---|
| D. wolongensis | W1, Wolong Valley, Sechuan | 2060 | open forest along rivulet | 23 |
| D. wolongensis | W2, Wolong Valley, Sechuan | 2084 | shady gorge along fast flowing stream | 22 |
| D. wolongensis | W3, Wolong Valley, Sechuan | 2100 | forest along small river | 22 |
| D. wolongensis | Coll Martyn Rix, ex United Kingdom * | 1 | ||
| D. wolongensis | Miya—Low, ex United Kingdom * | 1 | ||
| D. wolongensis | China Pink ex ChenYi, ex United Kingdom * | 1 | ||
| D. wolongensis | Kevock Star, ex United Kingdom * | |||
| D. acutiloba | CDC 626, ex United Kingdom * | 1 | ||
| D. retusa | Zheduo Shan, Kanding, Sechuan | 4100 | subalpine shrubland | 1 |
| D. retusa | Huanglong, Min Shan, Sechuan | 4300 | alpine shrubland | 1 |
| D. retusa | ex United Kingdom * | 1 | ||
| D. tangutica | Zhongdian, Yunnan | 3600 | open forest and shrubland | 1 |
| D. tangutica | form A, ex United Kingdom * | 1 | ||
| D. tangutica | form B, ex United Kingdom * | 1 | ||
| D. tangutica | Coll B4, Keith Rushforth, ex United Kingdom * | 1 | ||
| D. limprichtii | Jiuzhai, Min Shan, Sechuan | 4100 | alpine shrubland | 1 |
| D. longilobata | Baimucun, Lunang, Tibet | 3600 | deciduous forest | 10 |
| D. sureil | ex United Kingdom * | 1 | ||
| D. calcicola | Zhongdian, NapaHai, Yunnan | 3644 | evergreen oak forest (Q. aquifolioides) | 1 |
| Species/Populations | n a | P (%) b | ĥ c | I d | GST e | Nm f |
|---|---|---|---|---|---|---|
| D. wolongensis—W1 | 7 | 24.7 | 0.088 | 0.133 | ||
| D. wolongensis—W2 | 12 | 25.1 | 0.087 | 0.131 | ||
| D. wolongensis—W3 | 6 | 15.5 | 0.061 | 0.091 | ||
| D. wolongensis—varieties | 3 | 28.4 | 0.126 | 0.181 | ||
| D. wolongensis—total | 28 | 43.87 | 0.108 | 0.171 | 0.237 | 1.614 |
| D. retusa | 3 | 20.3 | 0.09 | 0.129 | ||
| D. tangutica | 3 | 5.4 | 0.024 | 0.034 | ||
| D. sureil, calcicola, acutiloba | 4 | 66.1 | 0.294 | 0.421 | ||
| D. spp. | 10 | 67.05 | 0.284 | 0.434 | 0.521 | 0.460 |
| Total | 38 | 99.43 | 0.215 | 0.351 | 0.301 | 1.164 |
| D. wolongensis—W1 | D. wolongensis—W2 | D. wolongensis—W3 | D. wolongensis—Varieties | D. acutiloba | D. retusa | D. tangutica | D. limprichtii | D. sureil | D. calcicola | |
|---|---|---|---|---|---|---|---|---|---|---|
| D. wolongensis—W1 | 0.000 | |||||||||
| D. wolongensis—W2 | 0.012 | 0.000 | ||||||||
| D. wolongensis—W3 | 0.028 | 0.032 | 0.000 | |||||||
| D. wolongensis—varieties | 0.030 | 0.028 | 0.053 | 0.000 | ||||||
| D. acutiloba | 0.444 | 0.441 | 0.450 | 0.361 | 0.000 | |||||
| D. retusa | 0.274 | 0.265 | 0.287 | 0.206 | 0.370 | 0.000 | ||||
| D. tangutica | 0.361 | 0.352 | 0.376 | 0.285 | 0.424 | 0.146 | 0.000 | |||
| D. limprichtii | 0.292 | 0.284 | 0.317 | 0.234 | 0.459 | 0.134 | 0.265 | 0.000 | ||
| D. sureil | 0.463 | 0.461 | 0.484 | 0.398 | 0.408 | 0.424 | 0.470 | 0.450 | 0.000 | |
| D. calcicola | 0.884 | 0.881 | 0.880 | 0.794 | 0.752 | 0.741 | 0.955 | 0.697 | 0.627 | 0.000 |
| Species | Population | Origin | K1 | K2 | K3 | K4 | K |
|---|---|---|---|---|---|---|---|
| Daphne wolongensis | W1 | China | 0.004 | 0.001 | 0.994 | 0.001 | 3 |
| Daphne wolongensis | W1 | China | 0.002 | 0.001 | 0.996 | 0.001 | 3 |
| Daphne wolongensis | W1 | China | 0.001 | 0.001 | 0.997 | 0.002 | 3 |
| Daphne wolongensis | W1 | China | 0.007 | 0.014 | 0.977 | 0.003 | 3 |
| Daphne wolongensis | W1 | China | 0 | 0 | 0.999 | 0 | 3 |
| Daphne wolongensis | W1 | China | 0.003 | 0.002 | 0.995 | 0.001 | 3 |
| Daphne wolongensis | W1 | China | 0 | 0 | 0.999 | 0 | 3 |
| Daphne wolongensis | W2 | China | 0 | 0 | 0.999 | 0 | 3 |
| Daphne wolongensis | W2 | China | 0.001 | 0 | 0.998 | 0.001 | 3 |
| Daphne wolongensis | W2 | China | 0.002 | 0.002 | 0.995 | 0.001 | 3 |
| Daphne wolongensis | W2 | China | 0 | 0 | 0.999 | 0 | 3 |
| Daphne wolongensis | W2 | China | 0.001 | 0.001 | 0.998 | 0 | 3 |
| Daphne wolongensis | W2 | China | 0.001 | 0.001 | 0.986 | 0.012 | 3 |
| Daphne wolongensis | W2 | China | 0.001 | 0.001 | 0.998 | 0 | 3 |
| Daphne wolongensis | W2 | China | 0.003 | 0.001 | 0.994 | 0.002 | 3 |
| Daphne wolongensis | W2 | China | 0.001 | 0.001 | 0.998 | 0 | 3 |
| Daphne wolongensis | W2 | China | 0.001 | 0 | 0.997 | 0.001 | 3 |
| Daphne wolongensis | W2 | China | 0.001 | 0 | 0.999 | 0 | 3 |
| Daphne wolongensis | W3 | China | 0.001 | 0.001 | 0.998 | 0 | 3 |
| Daphne wolongensis | W3 | China | 0 | 0 | 0.999 | 0 | 3 |
| Daphne wolongensis | W3 | China | 0.001 | 0.001 | 0.997 | 0 | 3 |
| Daphne wolongensis | W3 | China | 0.006 | 0.071 | 0.923 | 0 | 3 |
| Daphne wolongensis | W3 | China | 0 | 0 | 0.999 | 0 | 3 |
| Daphne wolongensis | W3 | China | 0 | 0 | 0.999 | 0 | 3 |
| Daphne wolongensis | Coll Martyn | UK | 0.001 | 0.111 | 0.691 | 0.196 | 3 |
| Daphne wolongensis | China pink | UK | 0.001 | 0.001 | 0.998 | 0 | 3 |
| Daphne wolongensis | Miyalow | UK | 0.26 | 0.089 | 0.65 | 0.001 | 3 |
| Daphne tangutica | form A | UK | 0.002 | 0 | 0.001 | 0.997 | 4 |
| Daphne tangutica | form B | UK | 0.002 | 0.001 | 0 | 0.997 | 4 |
| Daphne tangutica | Coll B4 | UK | 0.001 | 0.001 | 0.001 | 0.997 | 4 |
| Daphne sureil | White fls | UK | 0.001 | 0.996 | 0.001 | 0.001 | 2 |
| Daphne calcicola | Napa Hai | China | 0.001 | 0.999 | 0 | 0 | 2 |
| Daphne acutiloba | UK | 0.003 | 0.99 | 0.002 | 0.005 | 2 | |
| Daphne retusa | Sechuan | China | 0.903 | 0.002 | 0.002 | 0.093 | 1 |
| Daphne retusa | Hualong | China | 0.782 | 0.002 | 0.001 | 0.215 | 1 + 4 |
| Daphne retusa | UK | 0.476 | 0.008 | 0.067 | 0.449 | 1 + 4 | |
| Daphne limprichtii | Sechuan | China | 0.64 | 0.083 | 0.175 | 0.102 | 1 + 3 + 4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Holubec, V.; Leišová-Svobodová, L. Morphological and Molecular Status of Daphne wolongensis C.D.Brickell et B.Mathew as Genetic Resource for Horticulture. Agronomy 2020, 10, 1628. https://doi.org/10.3390/agronomy10111628
Holubec V, Leišová-Svobodová L. Morphological and Molecular Status of Daphne wolongensis C.D.Brickell et B.Mathew as Genetic Resource for Horticulture. Agronomy. 2020; 10(11):1628. https://doi.org/10.3390/agronomy10111628
Chicago/Turabian StyleHolubec, Vojtěch, and Leona Leišová-Svobodová. 2020. "Morphological and Molecular Status of Daphne wolongensis C.D.Brickell et B.Mathew as Genetic Resource for Horticulture" Agronomy 10, no. 11: 1628. https://doi.org/10.3390/agronomy10111628
APA StyleHolubec, V., & Leišová-Svobodová, L. (2020). Morphological and Molecular Status of Daphne wolongensis C.D.Brickell et B.Mathew as Genetic Resource for Horticulture. Agronomy, 10(11), 1628. https://doi.org/10.3390/agronomy10111628