

The Development of a 3D PET Fibrous Scaffold Modified with an Umbilical Cord dECM for Liver Tissue Engineering


Abstract
1. Introduction
2. Materials and Methods
2.1. Preparation and Analysis of Umbilical Cord dECM
2.2. Preparation and Characterization of PET Fibrous Scaffold Modified with Umbilical Cord dECM
2.2.1. Preparation of Human UC dECM Hydrogel
2.2.2. Pretreatment and NaOH Modification of PET Fibrous Scaffolds
2.2.3. UC dECM Hydrogel Modification of Fibrous Scaffolds
2.2.4. Characterization of PET-dECM Fibrous Scaffolds
2.3. Cell Culture on the Scaffolds
2.3.1. Culture of HepaRG Cells on the Scaffolds
2.3.2. Observation of Cell Morphology
2.3.3. Cell Proliferation Assay
2.3.4. Cell Function Assay
2.4. Statistical Analysis
3. Results and Discussion
3.1. Preparation and Analysis of Umbilical Cord dECM
3.2. Characterization of PET-dECM Fibrous Scaffolds
3.3. Biological Performance Test
3.3.1. Observation of Cell Morphology
3.3.2. Cell Activity and Proliferation
3.3.3. Cell Polarity, Stemness, and Function
4. Conclusions
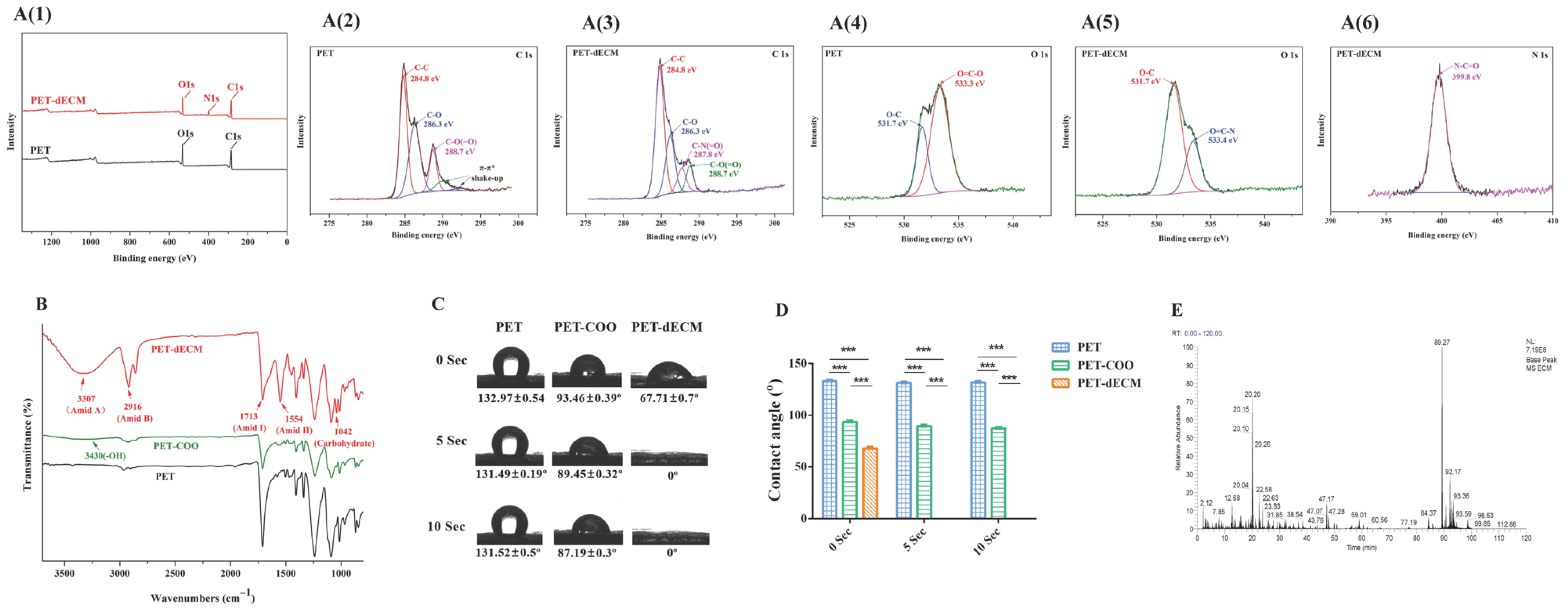
- The XPS analysis revealed that the N1s spectrum of the PET-dECM shows binding energies at 399.8 eV, corresponding to the amide groups formed after dECM modification. Concurrently, the N element content on the material surface amounted to 8.95%, suggesting that the surface element composition has changed after the modification treatment. The PET-dECM displays characteristic absorption peaks of collagen, specifically Amid II at 1554 cm−1, Amid I at 1713 cm−1, Amid B at 2916 cm−1, and Amid A at 3307 cm−1. Additionally, a characteristic absorption peak indicating the presence of carbohydrates is observed at 1042 cm−1. The dECM coating significantly enhanced the hydrophilicity of the PET material, resulting in a decrease in the water contact angle of the PET-dECM scaffold from 132.97° to 67.71° (p < 0.001). The HPLC-MS analysis of the PET-dECM revealed that the dECM coating on PET comprises collagen types I, II, III, IV, V, VI, and XI, alongside decorin and fibronectin.
- Based on the results obtained from electron microscopy experiments, cell viability staining, cell counting, and CCK-8 cell activity assays, it was evident that the PET-dECM material facilitated the adhesion and proliferation and maintained the activity of HepaRG cells. Furthermore, the PET-dECM significantly surpassed PET and PET-COO in terms of albumin secretion, urea synthesis, and CYP3A4 expression. Additional analysis of the transcription levels of genes associated with the polarity, stemness, and function of HepaRG cells cultured on different materials showed that the RNA transcription levels of ABCC2, Occludin, and NTCP were considerably upregulated on the PET-dECM, while SOX9, LGR5, and OCT4 were significantly downregulated. ALB, CYP3A4, and CPS1 demonstrated an upward transcriptional trend. These findings imply that PET-dECM material may regulate and induce differentiation by aiding in the restoration of HepaRG cell polarity.
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kezer, C.A.; Simonetto, D.A.; Shah, V.H. Acute on Chronic Liver Failure in Patients with Alcohol-Associated Hepatitis: A Review. Clin. Liver Dis. 2023, 27, 659–670. [Google Scholar] [CrossRef] [PubMed]
- Jayalakshmi, V.T.; Bernal, W. Update on the management of acute liver failure. Curr. Opin. Crit. Care 2020, 26, 163–170. [Google Scholar] [CrossRef]
- Bernal, W.; McPhail, M.J. Acute liver failure. J. Hepatol. 2021, 74, 1489–1490. [Google Scholar] [CrossRef]
- Siddiqui, M.S.; Stravitz, R.T. Intensive care unit management of patients with liver failure. Clin. Liver Dis. 2014, 18, 957–978. [Google Scholar] [CrossRef] [PubMed]
- Bhatia, S.N.; Underhill, G.H.; Zaret, K.S.; Fox, I.J. Cell and tissue engineering for liver disease. Sci. Transl. Med. 2014, 6, 245sr2. [Google Scholar] [CrossRef] [PubMed]
- Bernau, J.; Rueff, B.; Benhamou, J.P. Fulminant and subfulminant liver failure: Definitions and causes. Semin. Liver Dis. 1986, 6, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Krisper, P.; Haditsch, B.; Stauber, R.; Jung, A.; Stadlbauer, V.; Trauner, M.; Holzer, H.; Schneditz, D. In vivo quantification of liver dialysis: Comparison of albumin dialysis and fractionated plasma separation. J. Hepatol. 2005, 43, 451–457. [Google Scholar] [CrossRef] [PubMed]
- Balcar, L.; Krawanja, J.; Scheiner, B.; Paternostro, R.; Simbrunner, B.; Semmler, G.; Jachs, M.; Hartl, L.; Stättermayer, A.F.; Schwabl, P.; et al. Impact of ammonia levels on outcome in clinically stable outpatients with advanced chronic liver disease. JHEP Rep. 2023, 5, 100682. [Google Scholar] [CrossRef] [PubMed]
- Bhatia, V.; Singh, R.; Acharya, S.K. Predictive value of arterial ammonia for complications and outcome in acute liver failure. Gut 2006, 55, 98–104. [Google Scholar] [CrossRef] [PubMed]
- Fisher, C.; Baldwin, I.; Fealy, N.; Naorungroj, T.; Bellomo, R. Ammonia Clearance with Different Continuous Renal Replacement Therapy Techniques in Patients with Liver Failure. Blood Purif. 2022, 51, 840–846. [Google Scholar] [CrossRef] [PubMed]
- Dutkowski, P.; Oberkofler, C.E.; Béchir, M.; Müllhaupt, B.; Geier, A.; Raptis, D.A.; Clavien, P.A. The model for end-stage liver disease allocation system for liver transplantation saves lives, but increases morbidity and cost: A prospective outcome analysis. Liver Transpl. 2011, 17, 674–684. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, L.M.; Moreira, L.F.; Pinto, M.A.; Henriques-Pons, A.; Alves, L.A. Domino Hepatocyte Transplantation: A Therapeutic Alternative for the Treatment of Acute Liver Failure. Can. J. Gastroenterol. Hepatol. 2018, 2018, 2593745. [Google Scholar] [CrossRef] [PubMed]
- Kimmann, M.; Trebicka, J. Acute-on-chronic liver failure: Current interventional treatment options and future challenges. J. Pers. Med. 2023, 13, 1052. [Google Scholar] [CrossRef] [PubMed]
- Mazza, G.; Al-Akkad, W.; Rombouts, K.; Pinzani, M. Liver tissue engineering: From implantable tissue to whole organ engineering. Hepatol. Commun. 2018, 2, 131–141. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Chan, H.F.; Wang, H.; Shao, D.; Tao, Y.; Li, M. Stem cell therapy and tissue engineering strategies using cell aggregates and decellularized scaffolds for the rescue of liver failure. J. Tissue Eng. 2021, 12, 2041731420986711. [Google Scholar] [CrossRef] [PubMed]
- Langer, R.; Vacanti, J. Tissue engineering. Science 1993, 260, 920–926. [Google Scholar] [CrossRef] [PubMed]
- Kazemnejad, S.; Khanmohammadi, M.; Mobini, S.; Taghizadeh-Jahed, M.; Khanjani, S.; Arasteh, S.; Golshahi, H.; Torkaman, G.; Ravanbod, R.; Heidari-Vala, H.; et al. Comparative repair capacity of knee osteochondral defects using regenerated silk fiber scaffolds and fibrin glue with/without autologous chondrocytes during 36 weeks in rabbit model. Cell Tiss. Res. 2016, 364, 559–572. [Google Scholar] [CrossRef] [PubMed]
- Sedghi, R.; Shaabani, A.; Sayyari, N. Electrospun triazole-based chitosan nanofibers as a novel scaffold for bone tissue repair and regeneration. Carbohydr. Polym. 2020, 230, 115707. [Google Scholar] [CrossRef] [PubMed]
- Sivashankari, P.R.; Prabaharan, M. Three-dimensional porous scaffolds based on agarose/chitosan/graphene oxide composite for tissue engineering. Int. J. Biol. Macromol. 2019, 146, 222–231. [Google Scholar] [CrossRef] [PubMed]
- Glicklis, R.; Shapiro, L.; Agbaria, R.; Merchuk, J.C.; Cohen, S. Hepatocyte behavior within three-dimensional porous alginate scaffolds. Biotechnol. Bioeng. 2000, 67, 344–353. [Google Scholar] [CrossRef]
- Karamanos, N.K. Extracellular matrix: Key structural and functional meshwork in health and disease. FEBS J. 2019, 286, 2826–2829. [Google Scholar] [CrossRef] [PubMed]
- Franchi, M.; Piperigkou, Z.; Mastronikolis, N.S.; Karamanos, N. Extracellular matrix biomechanical roles and adaptation in health and disease. FEBS J. 2024, 291, 430–440. [Google Scholar] [CrossRef] [PubMed]
- Dong, C.; Lv, Y. Application of collagen scaffold in tissue engineering: Recent advances and new perspectives. Polymers 2016, 8, 42. [Google Scholar] [CrossRef] [PubMed]
- Peng, G.; Li, S.; Peng, Q.; Li, Y.; Weng, J.; Jia, Z.; Kang, J.; Lei, X.; Zhang, G.; Gao, Y. Immobilization of native type I collagen on polypropylene fabrics as a substrate for HepG2 cell culture. J. Biomater. Appl. 2017, 32, 93–103. [Google Scholar] [CrossRef] [PubMed]
- D’Amora, U.; D’Este, M.; Eglin, D.; Safari, F.; Sprecher, C.; Gloria, A.; De Santis, R.; Alini, M.; Ambrosio, L. Collagen density gradient on three-dimensional printed poly(“-caprolactone) scaffolds for interface tissue engineering. J. Tissue Eng. Regen. Med. 2018, 12, 321–329. [Google Scholar] [CrossRef] [PubMed]
- Kuo, Y.J.; Chen, C.H.; Dash, P.; Lin, Y.C.; Hsu, C.W.; Shih, S.J.; Chung, R.J. Angiogenesis, osseointegration, and antibacterial applications of polyelectrolyte multilayer coatings incorporated with silver/strontium containing mesoporous bioactive glass on 316L stainless steel. Front. Bioeng. Biotechnol. 2022, 10, 818137. [Google Scholar] [CrossRef] [PubMed]
- Vorotnikova, E.; McIntosh, D.; Dewilde, A.; Zhang, J.; Reing, J.E.; Zhang, L.; Cordero, K.; Bedelbaeva, K.; Gourevitch, D.; Heber-Katz, E.; et al. Extracellular matrix-derived products modulate endothelial and progenitor cell migration and proliferation in vitro and stimulate regenerative healing in vivo. Matrix Biol. 2010, 29, 690–700. [Google Scholar] [CrossRef] [PubMed]
- Barkan, D.; Green, J.E.; Chambers, A.F. Extracellular matrix: A gatekeeper in the transition from dormancy to metastatic growth. Eur. J. Cancer 2010, 46, 1181–1188. [Google Scholar] [CrossRef] [PubMed]
- De Santis, M.M.; Alsafadi, H.N.; Tas, S.; Bölükbas, D.A.; Prithiviraj, S.; Da Silva, I.A.N.; Mittendorfer, M.; Ota, C.; Stegmayr, J.; Daoud, F.; et al. Extracellular-Matrix-Reinforced Bioinks for 3D Bioprinting Human Tissue. Adv. Mater. 2021, 33, 2005476. [Google Scholar] [CrossRef] [PubMed]
- Saheli, M.; Sepantafar, M.; Pournasr, B.; Farzaneh, Z.; Vosough, M.; Piryaei, A.; Baharvand, H. Three-dimensional liver-derived extracellular matrix hydrogel promotes liver organoids function. J. Cell. Biochem. 2018, 119, 4320–4333. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Han, W.; Kim, H.; Ha, D.; Jang, J.; Kim, B.S.; Cho, D. Development of Liver Decellularized Extracellular Matrix Bioink for Three-Dimensional Cell Printing-Based Liver Tissue Engineering. Biomacromolecules 2017, 18, 1229–1237. [Google Scholar] [CrossRef]
- Damania, A.; Kumar, A.; Teotia, A.K.; Kimura, H.; Kamihira, M.; Ijima, H.; Sarin, S.K.; Kumar, A. Decellularized Liver Matrix-Modified Cryogel Scaffolds as Potential Hepatocyte Carriers in Bioartificial Liver Support Systems and Implantable Liver Constructs. ACS Appl. Mater. Interfaces 2018, 10, 114–126. [Google Scholar] [CrossRef] [PubMed]
- Nie, X.; Chuah, Y.J.; Zhu, W.; He, P.; Peck, Y.; Wang, D.-A. Decellularized tissue engineered hyaline cartilage graft for articular cartilage repair. Biomaterials 2020, 235, 119821. [Google Scholar] [CrossRef] [PubMed]
- Safari, F.; Fani, N.; Eglin, D.; Alini, M.; Stoddart, M.J.; Eslaminejad, M.B. Human umbilical cord-derived scaffolds for cartilage tissue engineering. J. Biomed. Mater. Res. Part A 2019, 107, 1793–1802. [Google Scholar] [CrossRef] [PubMed]
- Sanluis-Verdes, A.; Sanluis-Verdes, N.; Manso-Revilla, M.J.; Castro-Castro, A.M.; Pombo-Otero, J.; Fraga-Mariño, M.; Sanchez-Ibañez, J.; Doménech, N.; Rendal-Vázquez, M.E. Tissue engineering for neurodegenerative diseases using human amniotic membrane and umbilical cord. Cell Tissue Bank. 2017, 18, 1–15. [Google Scholar] [CrossRef]
- Ramzan, F.; Ekram, S.; Frazier, T.; Salim, A.; Mohiuddin, O.A.; Khan, I. Decellularized Human Umbilical Tissue-Derived Hydrogels Promote Proliferation and Chondrogenic Differentiation of Mesenchymal Stem Cells. Bioengineering 2022, 9, 239. [Google Scholar] [CrossRef]
- Shupe, T.; Williams, M.; Brown, A.; Willenberg, B.; Petersen, B.E. Method for the decellularization of intact rat liver. Organogenesis 2010, 6, 134–136. [Google Scholar] [CrossRef] [PubMed]
- Prest, T.A.; Yeager, E.; LoPresti, S.T.; Zygelyte, E.; Martin, M.J.; Dong, L.; Gibson, A.; Olutoye, O.O.; Brown, B.N.; Cheetham, J. Nerve-specific, xenogeneic extracellular matrix hydrogel promotes recovery following peripheral nerve injury. J. Biomed. Mater. Res. Part A 2018, 106, 450–459. [Google Scholar] [CrossRef] [PubMed]
- Pla-Palacín, I.; Sainz-Arnal, P.; Morini, S.; Almeida, M.; Baptista, P.M. Liver Bioengineering Using Decellularized Whole-Liver Scaffolds. Methods Mol. Biol. 2018, 1577, 293–305. [Google Scholar] [PubMed]
- Crapo, P.M.; Gilbert, T.W.; Badylak, S.F. An overview of tissue and whole organ decellularization processes. Biomaterials 2011, 32, 3233–3243. [Google Scholar] [CrossRef] [PubMed]
- Bui, V.T.; Liu, X.; Ko, S.H.; Choi, H.S. Super-amphiphilic surface of nano silica/polyurethane hybrid coated PET film via a plasma treatment. J. Colloid Interface Sci. 2015, 453, 209–215. [Google Scholar] [CrossRef] [PubMed]
- Rabchinskii, M.K.; Ryzhkov, S.A.; Kirilenko, D.A.; Ulin, N.V.; Baidakova, M.V.; Shnitov, V.V.; Pavlov, S.I.; Chumakov, R.G.; Stolyarova, D.Y.; Besedina, N.A.; et al. From graphene oxide towards aminated graphene: Facile synthesis, its structure and electronic properties. Sci. Rep. 2020, 10, 6902. [Google Scholar] [CrossRef] [PubMed]
- Szafran, K.; Jurak, M.; Mroczka, R.; Wiącek, A.E. Preparation and Surface Characterization of Chitosan-Based Coatings for PET Materials. Molecules 2023, 28, 2375. [Google Scholar] [CrossRef]
- Zhou, Y.; Chen, C.; Guo, Z.; Xie, S.; Hu, J.; Lu, H. SR-FTIR as a tool for quantitative mapping of the content and distribution of extracellular matrix in decellularized book-shape bioscaffolds. BMC Musculoskelet. Disord. 2018, 19, 220. [Google Scholar] [CrossRef]
- Kim, H.; Panda, P.K.; Sadeghi, K.; Seo, J. Poly (vinyl alcohol)/hydrothermally treated tannic acid composite films as sustainable antioxidant and barrier packaging materials. Prog. Org. Coat. 2023, 174, 107305. [Google Scholar] [CrossRef]
- Jha, S.; Malviya, R.; Fuloria, S.; Sundram, S.; Subramaniyan, V.; Sekar, M.; Sharma, P.K.; Chakravarthi, S.; Wu, Y.S.; Mishra, N.; et al. Characterization of Microwave-Controlled Polyacrylamide Graft Copolymer of Tamarind Seed Polysaccharide. Polymers 2022, 14, 1037. [Google Scholar] [CrossRef] [PubMed]
- Woźniak, A.; Smok, W.; Szewczenko, J.; Staszuk, M.; Chladek, G. Influence of Hybrid Surface Modification on Biocompatibility and Physicochemical Properties of Ti-6Al-4V ELI Titanium. J. Funct. Biomater. 2024, 20, 52. [Google Scholar] [CrossRef] [PubMed]
- Tascher, G.; Burban, A.; Camus, S.; Plumel, M.; Chanon, S.; Le Guevel, R.; Shevchenko, V.; Van Dorsselaer, A.; Lefai, E.; Guguen-Guillouzo, C.; et al. In-Depth Proteome Analysis Highlights HepaRG Cells as a Versatile Cell System Surrogate for Primary Human Hepatocytes. Cells 2019, 8, 192. [Google Scholar] [CrossRef]
- Jackson, J.P.; Li, L.; Chamberlain, E.D.; Wang, H.; Ferguso, S.S. Contextualizing Hepatocyte Functionality of Cryopreserved HepaRG Cell Cultures. Drug Metab. Dispos. 2016, 44, 1463–1479. [Google Scholar] [CrossRef]
- Nibourg, G.A.A.; Chamuleau, R.A.F.M.; van der Hoeven, T.V.; Maas, M.A.W.; Ruiter, A.F.C.; Lamers, W.H.; Oude Elferink, R.P.J.; van Gulik, T.M.; Hoekstra, R. Liver progenitor cell line HepaRG differentiated in a bioartificial liver effectively supplies liver support to rats with acute liver failure. PLoS ONE 2012, 7, e38778. [Google Scholar] [CrossRef]
- Hoekstra, R.; Nibourg, G.A.A.; van der Hoeven, T.V.; Ackermans, M.T.; Hakvoort, T.B.M.; van Gulik, T.M.; Lamers, W.H.; Oude Elferink, R.P.; Chamuleau, R.A.F.M. The HepaRG cell line is suitable for bioartificial liver application. Int. J. Biochem. Cell Biol. 2011, 43, 1483–1489. [Google Scholar] [CrossRef] [PubMed]
- Rose, C.F.; Amodio, P.; Bajaj, J.S.; Dhiman, R.K.; Montagnese, S.; Taylor-Robinson, S.D.; Vilstrup, H.; Jalan, R. Hepatic encephalopathy: Novel insights into classification, pathophysiology and therapy. J. Hepatol. 2020, 73, 1526–1547. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Boyer, I.L. The maintenance and generation of membrane polarity in hepatocytes. Hepatology 2004, 39, 892–899. [Google Scholar] [CrossRef] [PubMed]
- Cheung, I.D.; Bagnat, M.; Ma, T.P.; Datta, A.; Evason, K.; Moore, J.C.; Lawson, N.D.; Mostov, K.E.; Moens, C.B.; Stainier, D.Y.R. Regulation of intrahepatic biliary duct morphogenesis by Claudin 15-like b. Dev. Biol. 2012, 361, 68–78. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.C.; Pan, D.G.; Liao, W.; Jia, Z.D.; Pan, M.X.; Weng, J.; Han, X.; Li, S.; Li, Y.; Liang, K.Y.; et al. Application of 3D Hepatic Plate-Like Liver Model for Statin-Induced Hepatotoxicity Evaluation. Front. Bioeng. Biotechnol. 2022, 10, 826093. [Google Scholar] [CrossRef] [PubMed]
 : cell structure. (D) Immunofluorescence staining. Scale bar: 100 μm. (E) DNA content. Biochemical assays for (F) collagen, (G) glycosaminoglycans (GAGs), and (H) elastin in native UC and UC dECM. Data are given as mean ± SD, n = 3; *: p < 0.05, and ***: p < 0.001.
: cell structure. (D) Immunofluorescence staining. Scale bar: 100 μm. (E) DNA content. Biochemical assays for (F) collagen, (G) glycosaminoglycans (GAGs), and (H) elastin in native UC and UC dECM. Data are given as mean ± SD, n = 3; *: p < 0.05, and ***: p < 0.001.
: cell structure. (D) Immunofluorescence staining. Scale bar: 100 μm. (E) DNA content. Biochemical assays for (F) collagen, (G) glycosaminoglycans (GAGs), and (H) elastin in native UC and UC dECM. Data are given as mean ± SD, n = 3; *: p < 0.05, and ***: p < 0.001.
: cell structure. (D) Immunofluorescence staining. Scale bar: 100 μm. (E) DNA content. Biochemical assays for (F) collagen, (G) glycosaminoglycans (GAGs), and (H) elastin in native UC and UC dECM. Data are given as mean ± SD, n = 3; *: p < 0.05, and ***: p < 0.001.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer (5′-3′) | Reverse Primer (5′-3′) |
|---|---|---|
| ABCC2 | TCTCTCGATACTCTGTGGCAC | CTGGAATCCGTAGGAGATGAAGA |
| Occludin | GACTTCAGGCAGCCTCGTTAC | GCCAGTTGTGTAGTCTGTCTCA |
| NTCP | GGCCGTCACAGTTCTCTCTG | GGTGGCAATCAAGAGTGGTGT |
| SOX9 | AGCGAACGCACATCAAGAC | CTGTAGGCGATCTGTTGGGG |
| LGR5 | CACCTCCTACCTAGACCTCAGT | CGCAAGACGTAACTCCTCCAG |
| OCT4 | GGGAGATTGATAACTGGTGTGTT | GTGTATATCCCAGGGTGATCCTC |
| ALB | TGCAACTCTTCGTGAAACCTATG | ACATCAACCTCTGGTCTCACC |
| CPS1 | AATGAGGTGGGCTTAAAGCAAG | AGTTCCACTCCACAGTTCAGA |
| CYP3A4 | AAGTCGCCTCGAAGATACACA | AAGGAGAGAACACTGCTCGTG |
| β-Actin | CATGTACGTTGCTATCCAGGC | CTCCTTAATGTCACGCACGAT |
| Sample | Elementary Composition | ||
|---|---|---|---|
| C (%) | N (%) | O (%) | |
| PET | 71.34 | 0.47 | 27.91 |
| PET-dECM | 67.03 | 8.95 | 23.53 |
| Protein Sequence Number | Protein Type | Coverage Rate (%) | Peptides | Unique Peptides | Molecular Weight (kDa) |
|---|---|---|---|---|---|
| P02452 | Collagen alpha-1(I) chain | 26 | 27 | 5 | 138.9 |
| A0A384MDU2 | Collagen, type I, alpha 2 | 30 | 29 | 17 | 129.2 |
| P28481 | Collagen alpha-1(II) chain | 3 | 3 | 1 | 141.9 |
| P02461 | Collagen alpha-1(III) chain | 24 | 26 | 25 | 138.5 |
| A0A024RDW8 | Collagen, type IV, alpha 2 | 4 | 5 | 5 | 167.4 |
| B2ZZ86 | Collagen type V alpha 1 | 6 | 8 | 7 | 183.5 |
| P05997 | Collagen alpha-2(V) chain | 10 | 10 | 10 | 144.8 |
| D9ZGF2 | Collagen, type VI, alpha 3 | 4 | 12 | 12 | 343.5 |
| D3DT71 | Collagen, type XI, alpha 1 | 2 | 3 | 2 | 176.5 |
| P07585 | Decorin | 3 | 1 | 1 | 39.7 |
| A0A024R462 | Fibronectin | 3 | 4 | 4 | 259 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Zhang, Y.; Zhong, K.; Liao, S.; Zhang, G. The Development of a 3D PET Fibrous Scaffold Modified with an Umbilical Cord dECM for Liver Tissue Engineering. Polymers 2024, 16, 1794. https://doi.org/10.3390/polym16131794
Li Y, Zhang Y, Zhong K, Liao S, Zhang G. The Development of a 3D PET Fibrous Scaffold Modified with an Umbilical Cord dECM for Liver Tissue Engineering. Polymers. 2024; 16(13):1794. https://doi.org/10.3390/polym16131794
Chicago/Turabian StyleLi, Yang, Yang Zhang, Kebo Zhong, Shuguang Liao, and Guifeng Zhang. 2024. "The Development of a 3D PET Fibrous Scaffold Modified with an Umbilical Cord dECM for Liver Tissue Engineering" Polymers 16, no. 13: 1794. https://doi.org/10.3390/polym16131794
APA StyleLi, Y., Zhang, Y., Zhong, K., Liao, S., & Zhang, G. (2024). The Development of a 3D PET Fibrous Scaffold Modified with an Umbilical Cord dECM for Liver Tissue Engineering. Polymers, 16(13), 1794. https://doi.org/10.3390/polym16131794