
A Molecular Integrative Study on the Inhibitory Effects of WRR and ERW on Amyloid β Peptide (1–42) Polymerization and Cell Toxicity




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Molecular Docking
2.3. Umbrella Sampling of Binding Free Energy
2.4. ThT Fluorescent Detection of Aggregation of Aβ42
2.5. Transmission Electron Microscopy Observations of Aggregated Aβ42
2.6. SH-SY5Y Cell Culture
2.7. Transfection and Expression of Secreted Aβ42
2.8. Flow Cytometry Methods to Detect Reactive Oxygen Species and Apoptosis
3. Results
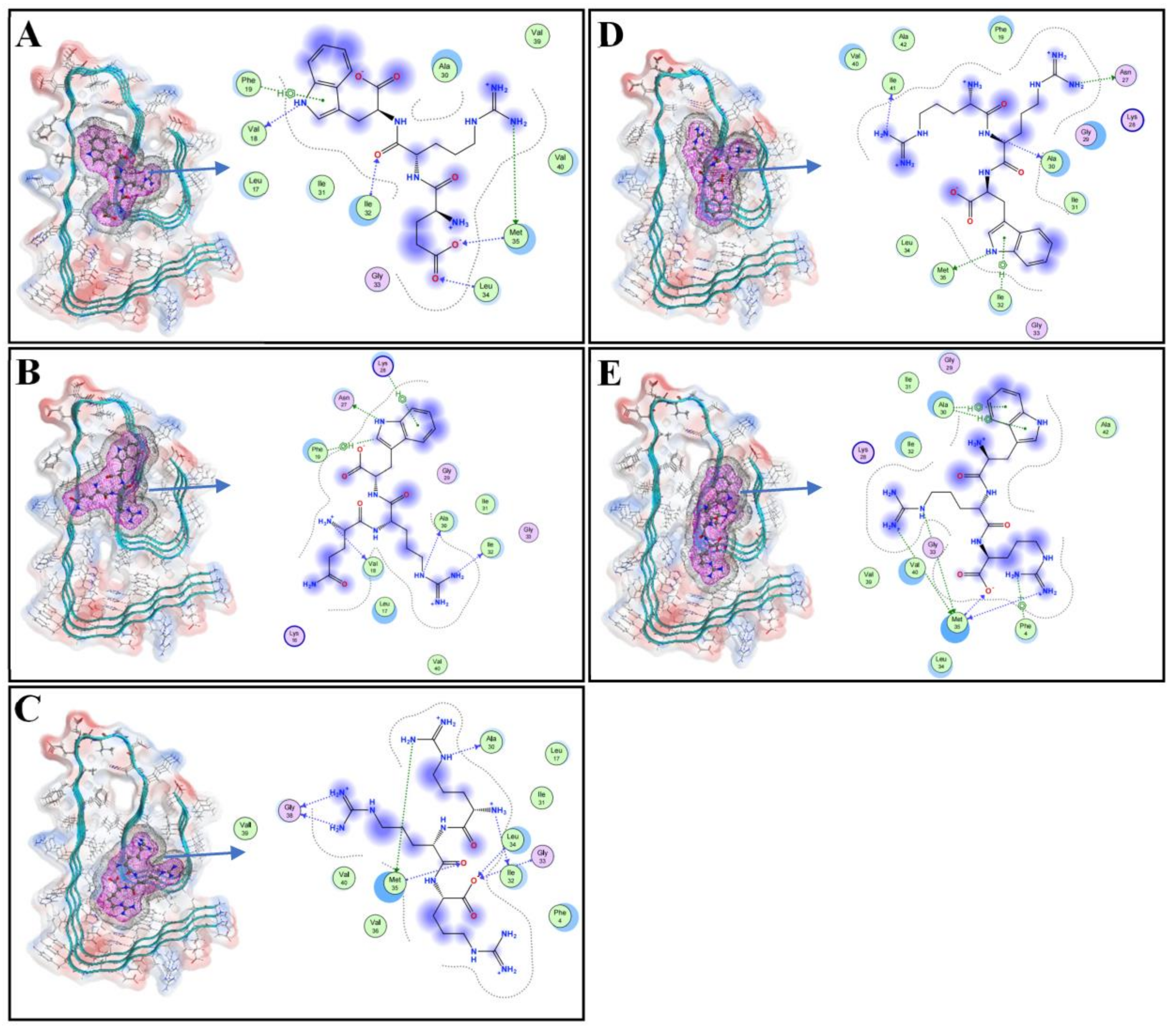
3.1. Molecular Docking of Tripeptides
3.2. Evaluation of Conformational Rationality Using a Ramachandran Plot
3.3. Umbrella Sampling Analysis of Ligand and Aβ42 Monomer
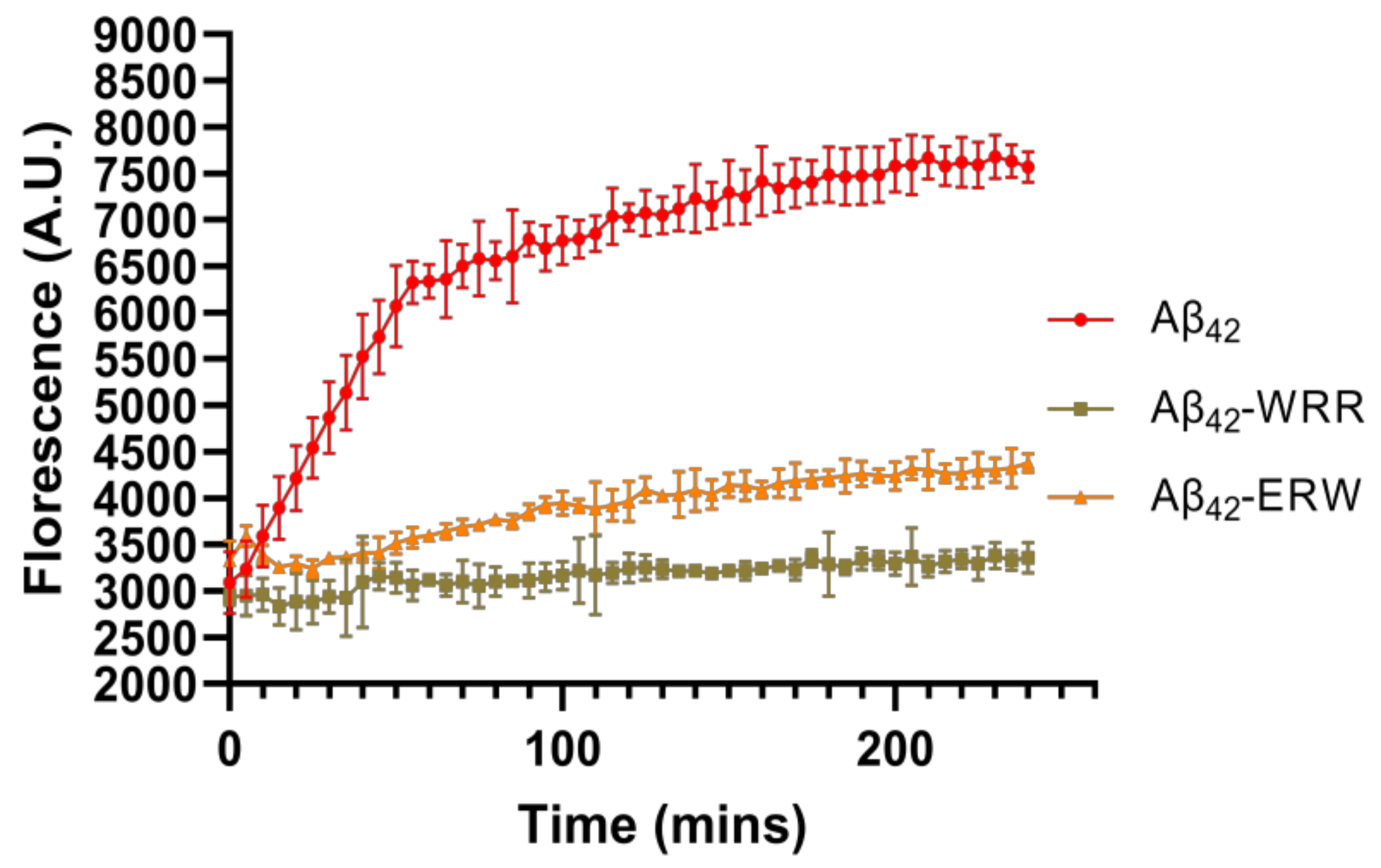
3.4. Effects of Oligopeptides on Aggregation of Aβ42 Detected by ThT Fluorescence Assay
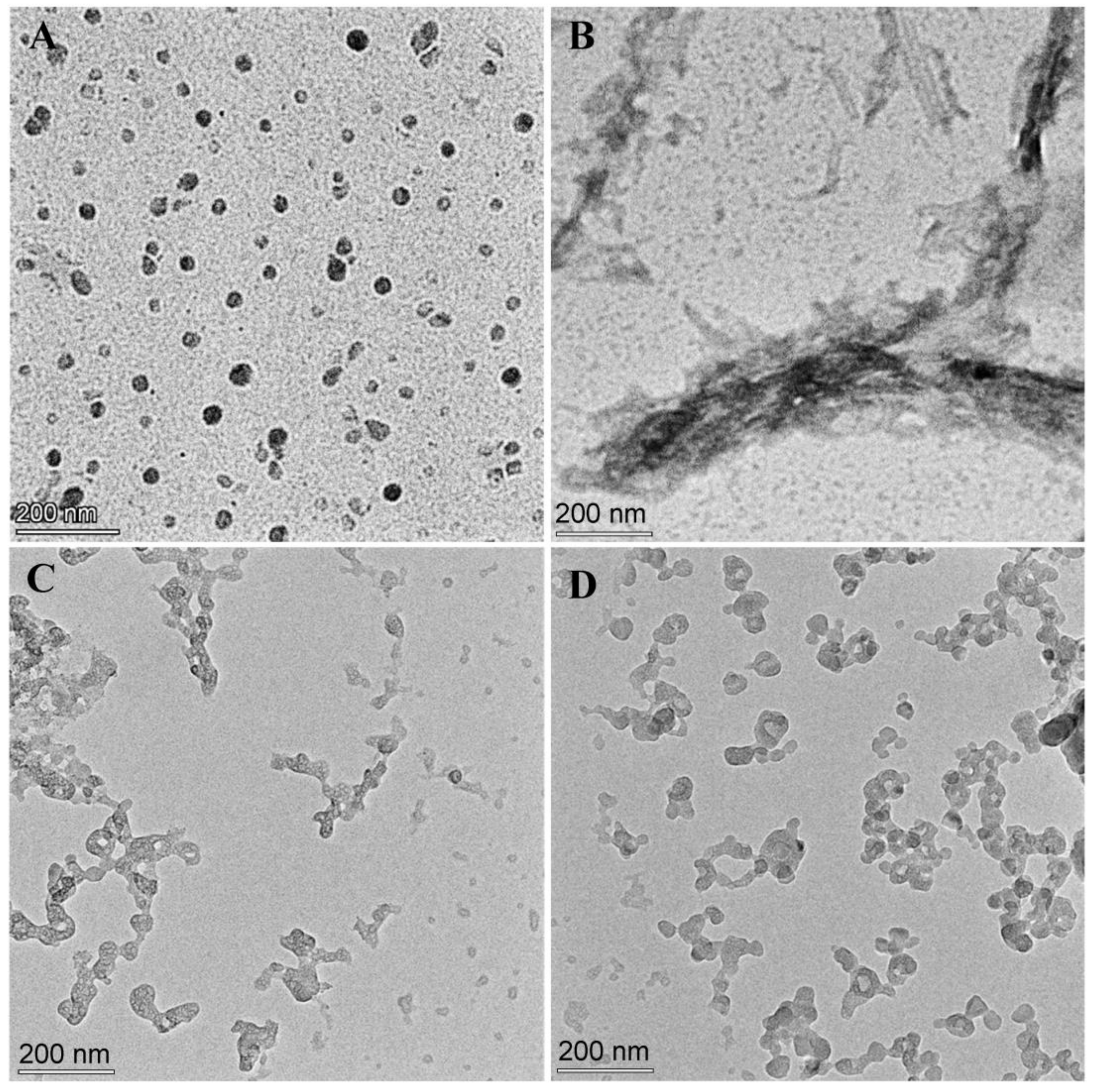
3.5. Transmission Electron Microscopy Observations of the Effect of WRR or EWR on Aβ42 Aggregation
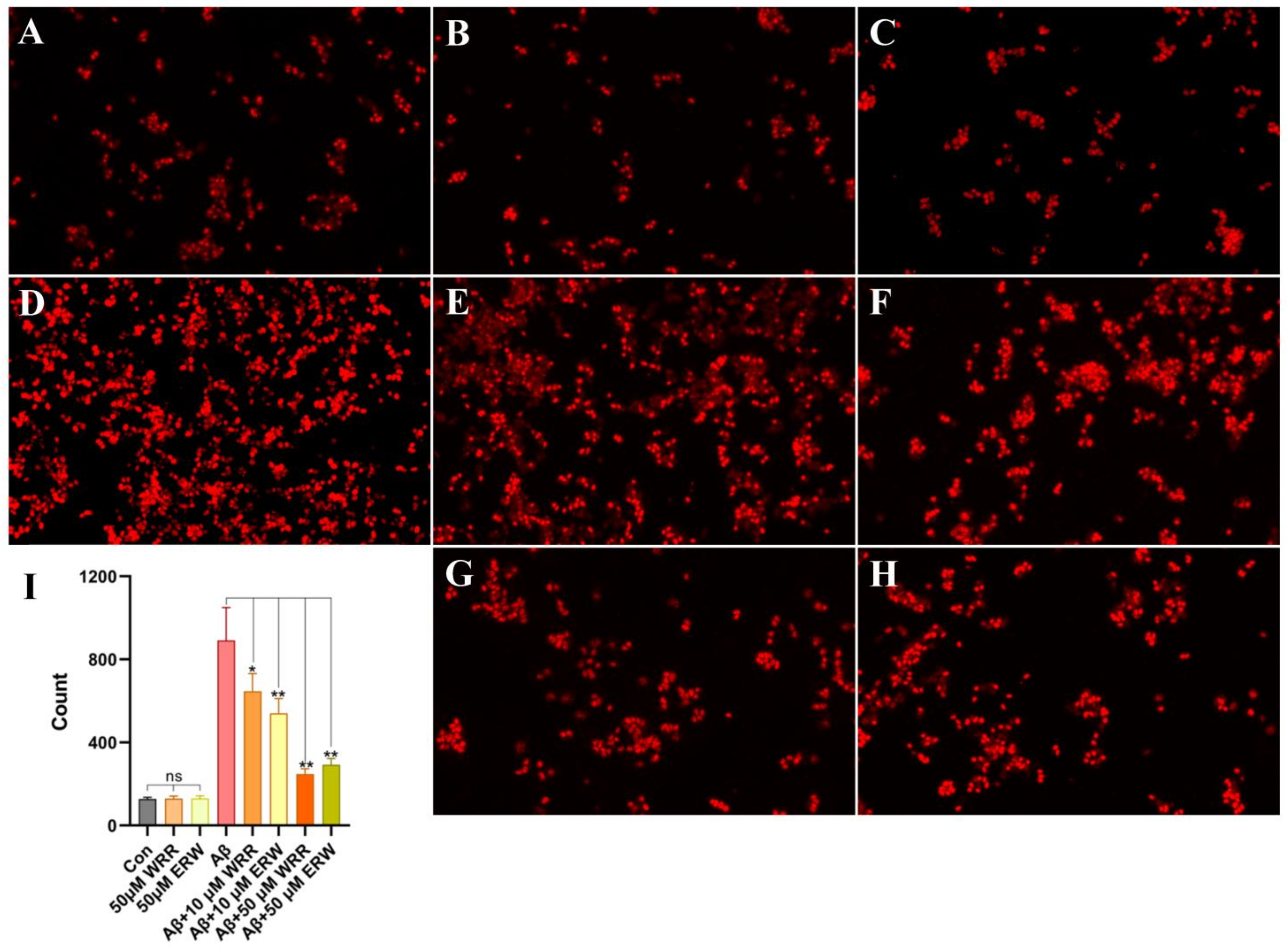
3.6. Effect of Either WRR or ERW on Cell Death Induced by Secreted Aβ42
3.7. Effect of Either WRR or ERW on Reactive Oxygen Species Produced by SH-SY5Y Cells Secreting Aβ42
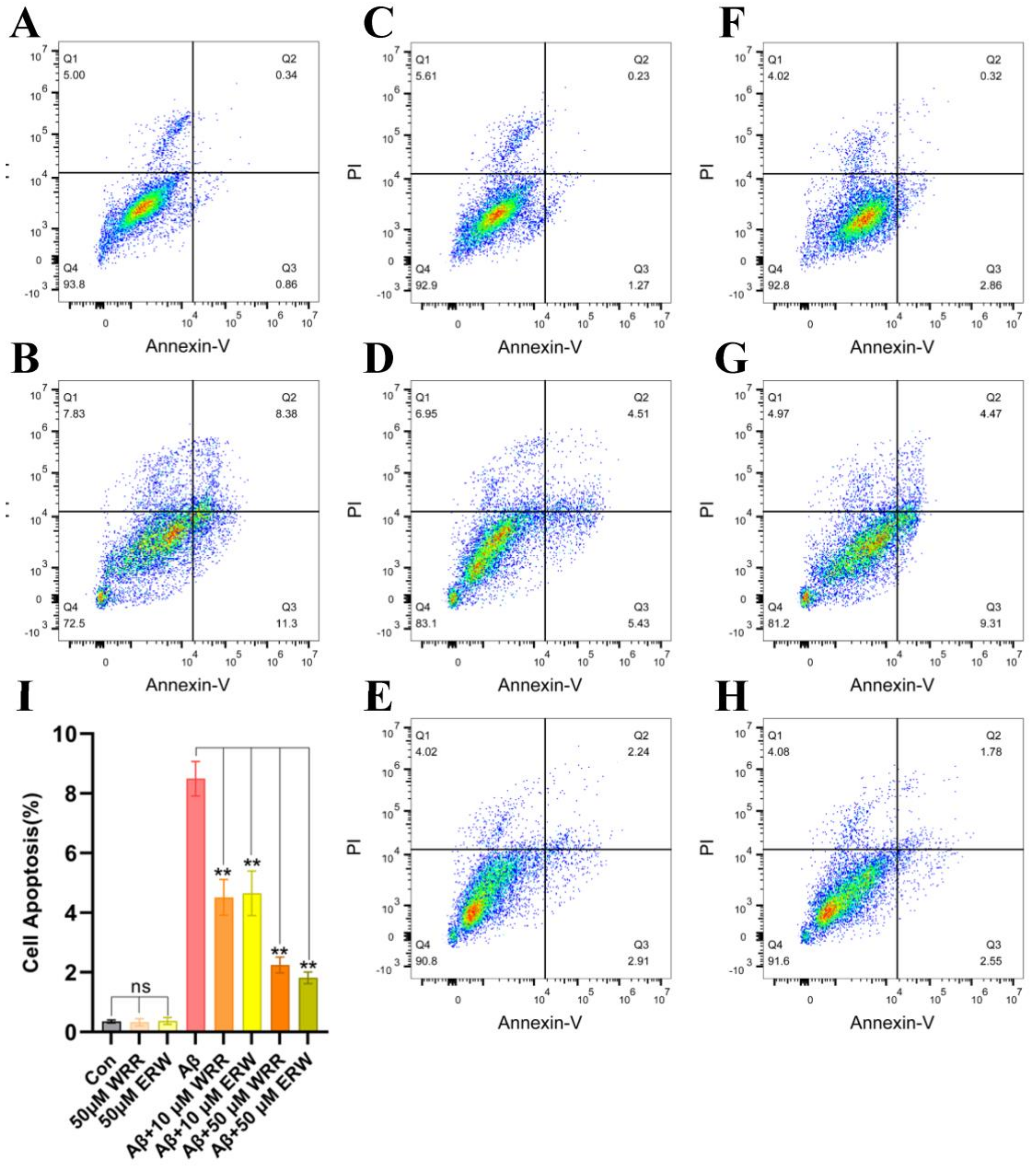
3.8. Protective Effect of Either WRR or ERW on Cell Apoptosis of SH-SY5Y Cells Secreting Aβ42
4. Discussion
5. Conclusions
6. Patents
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Bloom, G.S. Amyloid-beta and tau: The trigger and bullet in Alzheimer disease pathogenesis. JAMA Neurol. 2014, 71, 505–508. [Google Scholar] [CrossRef]
- Sun, X.; Duan, S.; Cao, A.; Villagomez, B.; Lin, R.; Chen, H.; Pi, L.; Ren, B.; Chen, R.; Chen, M.; et al. RRY Inhibits Amyloid-beta(1-42) Peptide Aggregation and Neurotoxicity. J. Alzheimer’s Dis. Rep. 2021, 5, 479–495. [Google Scholar] [CrossRef] [PubMed]
- Karran, E.; Mercken, M.; De Strooper, B. The amyloid cascade hypothesis for Alzheimer’s disease: An appraisal for the development of therapeutics. Nat. Rev. Drug Discov. 2011, 10, 698–712. [Google Scholar] [CrossRef] [PubMed]
- Reitz, C. Alzheimer’s disease and the amyloid cascade hypothesis: A critical review. Int. J. Alzheimer’s Dis. 2012, 2012, 369808. [Google Scholar] [CrossRef] [PubMed]
- Wilkins, H.M.; Swerdlow, R.H. Amyloid precursor protein processing and bioenergetics. Brain Res. Bull. 2017, 133, 71–79. [Google Scholar] [CrossRef]
- Waragai, M.; Yoshida, M.; Mizoi, M.; Saiki, R.; Kashiwagi, K.; Takagi, K.; Arai, H.; Tashiro, J.; Hashimoto, M.; Iwai, N.; et al. Increased protein-conjugated acrolein and amyloid-beta40/42 ratio in plasma of patients with mild cognitive impairment and Alzheimer’s disease. J. Alzheimer’s Dis. 2012, 32, 33–41. [Google Scholar] [CrossRef]
- Murray, P.S.; Kirkwood, C.M.; Gray, M.C.; Ikonomovic, M.D.; Paljug, W.R.; Abrahamson, E.E.; Henteleff, R.A.; Hamilton, R.L.; Kofler, J.K.; Klunk, W.E.; et al. Beta-Amyloid 42/40 ratio and kalirin expression in Alzheimer disease with psychosis. Neurobiol. Aging 2012, 33, 2807–2816. [Google Scholar] [CrossRef]
- Peng, X.L.; Hou, L.; Xu, S.H.; Hua, Y.; Zhou, S.J.; Zhang, Y.; Zheng, Y.P.; Fu, Y.H.; Xu, Q.; Zhang, L.S.; et al. Novel APP K724M mutation causes Chinese early-onset familial Alzheimer’s disease and increases amyloid-beta42 to amyloid-beta40 ratio. Neurobiol. Aging 2014, 35, 2657e1–2657e6. [Google Scholar] [CrossRef]
- Selkoe, D.J.; Hardy, J. The amyloid hypothesis of Alzheimer’s disease at 25 years. EMBO Mol. Med. 2016, 8, 595–608. [Google Scholar] [CrossRef]
- Edwards, F.A. A Unifying Hypothesis for Alzheimer’s Disease: From Plaques to Neurodegeneration. Trends Neurosci. 2019, 42, 310–322. [Google Scholar] [CrossRef]
- Lane, C.A.; Hardy, J.; Schott, J.M. Alzheimer’s disease. Eur. J. Neurol. 2018, 25, 59–70. [Google Scholar] [CrossRef] [PubMed]
- Kawasaki, T.; Onodera, K.; Kamijo, S. Identification of novel short peptide inhibitors of soluble 37/48 kDa oligomers of amyloid beta42. Biosci. Biotechnol. Biochem. 2011, 75, 1496–1501. [Google Scholar] [CrossRef] [PubMed]
- Kawasaki, T.; Kamijo, S. Inhibition of aggregation of amyloid beta42 by arginine-containing small compounds. Biosci. Biotechnol. Biochem. 2012, 76, 762–766. [Google Scholar] [CrossRef] [PubMed]
- Novick, P.A.; Lopes, D.H.; Branson, K.M.; Esteras-Chopo, A.; Graef, I.A.; Bitan, G.; Pande, V.S. Design of beta-amyloid aggregation inhibitors from a predicted structural motif. J. Med. Chem. 2012, 55, 3002–3010. [Google Scholar] [CrossRef] [PubMed]
- Yuan, N.; Ye, L.; Sun, Y.; Wu, H.; Xiao, Z.; Fu, W.; Chen, Z.; Pei, Y.; Min, Y.; Wang, D. Molecular Integrative Analysis of the Inhibitory Effects of Dipeptides on Amyloid beta Peptide 1-42 Polymerization. Int. J. Mol. Sci. 2023, 24, 7673. [Google Scholar] [CrossRef]
- Liu, F.F.; Dong, X.Y.; He, L.; Middelberg, A.P.; Sun, Y. Molecular insight into conformational transition of amyloid beta-peptide 42 inhibited by (-)-epigallocatechin-3-gallate probed by molecular simulations. J. Phys. Chem. B 2011, 115, 11879–11887. [Google Scholar] [CrossRef]
- Reddy, G.; Straub, J.E.; Thirumalai, D. Dynamics of locking of peptides onto growing amyloid fibrils. Proc. Natl. Acad. Sci. USA 2009, 106, 11948–11953. [Google Scholar] [CrossRef]
- Gremer, L.; Scholzel, D.; Schenk, C.; Reinartz, E.; Labahn, J.; Ravelli, R.B.G.; Tusche, M.; Lopez-Iglesias, C.; Hoyer, W.; Heise, H.; et al. Fibril structure of amyloid-beta(1-42) by cryo-electron microscopy. Science 2017, 358, 116–119. [Google Scholar] [CrossRef]
- Hodsdon, M.E.; Ponder, J.W.; Cistola, D.P. The NMR solution structure of intestinal fatty acid-binding protein complexed with palmitate: Application of a novel distance geometry algorithm. J. Mol. Biol. 1996, 264, 585–602. [Google Scholar] [CrossRef]
- Tam, B.; Sinha, S.; Wang, S.M. Combining Ramachandran plot and molecular dynamics simulation for structural-based variant classification: Using TP53 variants as model. Comput. Struct. Biotechnol. J. 2020, 18, 4033–4039. [Google Scholar] [CrossRef]
- Mannige, R.V. An exhaustive survey of regular peptide conformations using a new metric for backbone handedness (h). PeerJ 2017, 5, e3327. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Wang, B.; Pei, J.; Luo, Y.; Yuan, N.; Xiao, Z.; Wu, H.; Luo, C.; Wang, J.; Wei, S.; et al. Molecular dynamic and pharmacological studies on protein-engineered hirudin variants of Hirudinaria manillensis and Hirudo medicinalis. Br. J. Pharmacol. 2022, 179, 3740–3753. [Google Scholar] [CrossRef] [PubMed]
- Ghanta, J.; Shen, C.L.; Kiessling, L.L.; Murphy, R.M. A strategy for designing inhibitors of beta-amyloid toxicity. J. Biol. Chem. 1996, 271, 29525–29528. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Biancalana, M.; Koide, S.; Shea, J.E. Binding modes of thioflavin-T to the single-layer beta-sheet of the peptide self-assembly mimics. J. Mol. Biol. 2009, 394, 627–633. [Google Scholar] [CrossRef] [PubMed]
- Qiu, T.; Liu, Q.; Chen, Y.X.; Zhao, Y.F.; Li, Y.M. Abeta42 and Abeta40: Similarities and differences. J. Pept. Sci. 2015, 21, 522–529. [Google Scholar] [CrossRef]
- Rajasekhar, K.; Madhu, C.; Govindaraju, T. Natural Tripeptide-Based Inhibitor of Multifaceted Amyloid β Toxicity. ACS Chem. Neurosci. 2016, 7, 1300–1310. [Google Scholar] [CrossRef]
- Hu, X.; Zhang, Q.; Wang, W.; Yuan, Z.; Zhu, X.; Chen, B.; Chen, X. Tripeptide GGH as the Inhibitor of Copper-Amyloid-β-Mediated Redox Reaction and Toxicity. ACS Chem. Neurosci. 2016, 7, 1255–1263. [Google Scholar] [CrossRef]
- Lobello, K.; Ryan, J.M.; Liu, E.; Rippon, G.; Black, R. Targeting Beta amyloid: A clinical review of immunotherapeutic approaches in Alzheimer’s disease. Int. J. Alzheimer’s Dis. 2012, 2012, 628070. [Google Scholar] [CrossRef]
- Soto, C.; Sigurdsson, E.M.; Morelli, L.; Kumar, R.A.; Castano, E.M.; Frangione, B. Beta-sheet breaker peptides inhibit fibrillogenesis in a rat brain model of amyloidosis: Implications for Alzheimer’s therapy. Nat. Med. 1998, 4, 822–826. [Google Scholar] [CrossRef]
- Tjernberg, L.O.; Naslund, J.; Lindqvist, F.; Johansson, J.; Karlstrom, A.R.; Thyberg, J.; Terenius, L.; Nordstedt, C. Arrest of beta-amyloid fibril formation by a pentapeptide ligand. J. Biol. Chem. 1996, 271, 8545–8548. [Google Scholar] [CrossRef]
- Singh, K.; Kaur, A.; Goyal, D.; Goyal, B. Mechanistic insights into the mitigation of Aβ aggregation and protofibril destabilization by a D-enantiomeric decapeptide rk10. Phys. Chem. Chem. Phys. 2022, 24, 21975–21994. [Google Scholar] [CrossRef] [PubMed]
- Narang, S.S.; Goyal, D.; Goyal, B. Inhibition of Alzheimer’s amyloid-β42 peptide aggregation by a bi-functional bis-tryptoline triazole: Key insights from molecular dynamics simulations. J. Biomol. Struct. Dyn. 2020, 38, 1598–1611. [Google Scholar] [CrossRef] [PubMed]
- Asadbegi, M.; Shamloo, A. Identification of a Novel Multifunctional Ligand for Simultaneous Inhibition of Amyloid-Beta (Aβ42) and Chelation of Zinc Metal Ion. ACS Chem. Neurosci. 2019, 10, 4619–4632. [Google Scholar] [CrossRef] [PubMed]
- Kaur, A.; Shuaib, S.; Goyal, D.; Goyal, B. Interactions of a multifunctional di-triazole derivative with Alzheimer’s Aβ42 monomer and Aβ42 protofibril: A systematic molecular dynamics study. Phys. Chem. Chem. Phys. 2020, 22, 1543–1556. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, Z.; Ye, L.; Yuan, N.; Che Ajuyo, N.M.; Xiao, Z.; Liu, L.; Chen, Z.; Pei, Y.; Min, Y.; Wang, D. A Molecular Integrative Study on the Inhibitory Effects of WRR and ERW on Amyloid β Peptide (1–42) Polymerization and Cell Toxicity. Polymers 2023, 15, 4356. https://doi.org/10.3390/polym15224356
Wu Z, Ye L, Yuan N, Che Ajuyo NM, Xiao Z, Liu L, Chen Z, Pei Y, Min Y, Wang D. A Molecular Integrative Study on the Inhibitory Effects of WRR and ERW on Amyloid β Peptide (1–42) Polymerization and Cell Toxicity. Polymers. 2023; 15(22):4356. https://doi.org/10.3390/polym15224356
Chicago/Turabian StyleWu, Zhongyun, Lianmeng Ye, Nan Yuan, Nuela Manka’a Che Ajuyo, Zhengpan Xiao, Liangwang Liu, Zuqian Chen, Yechun Pei, Yi Min, and Dayong Wang. 2023. "A Molecular Integrative Study on the Inhibitory Effects of WRR and ERW on Amyloid β Peptide (1–42) Polymerization and Cell Toxicity" Polymers 15, no. 22: 4356. https://doi.org/10.3390/polym15224356
APA StyleWu, Z., Ye, L., Yuan, N., Che Ajuyo, N. M., Xiao, Z., Liu, L., Chen, Z., Pei, Y., Min, Y., & Wang, D. (2023). A Molecular Integrative Study on the Inhibitory Effects of WRR and ERW on Amyloid β Peptide (1–42) Polymerization and Cell Toxicity. Polymers, 15(22), 4356. https://doi.org/10.3390/polym15224356