
Lotus-Flower- and Lotus-Seedpod-Derived Polysaccharide: Structural Characterization and Biological Activity



Abstract
:1. Introduction
2. Material and Methods
2.1. Materials and Reagent
2.2. Extraction and Purification of Lotus-Flower- and Lotus-Seedpod-Derived Poly Nanoparticles
2.3. Structure Characterization and Molecular Face
2.3.1. Chemical Analysis of LFP and LSP
2.3.2. Monosaccharide Composition Analysis and Molecular Weight Assay
2.3.3. Infrared Spectrum Analysis
2.3.4. X-ray Diffraction (XRD) Analysis
2.3.5. SEM Morphology
2.4. Antioxidant Activity
2.4.1. 2,2′-Azino-bis (3-ethylbenzthiazoline-6-sulfonic acid) (ABTS+) Free Radical
2.4.2. DPPH Free Radical
2.4.3. Hydroxyl Radical
2.4.4. Reducing Power
2.5. Assay of Immunomodulatory Activity of LFP and LSP
2.5.1. Cell Cultivation
2.5.2. Cell Viability
2.5.3. Phagocytosis Assay
2.5.4. Scratch-Wound Assay
2.5.5. Extraction of mRNA and Real-Time Quantitative Polymerase Chain Reaction
2.5.6. The Western Blotting and Total Protein Extraction Assays
2.6. Statistical Analysis
3. Results and Analysis
3.1. Yield and Chemical Composition
3.2. Monosaccharide Composition and Molecular Weight
3.3. Fourier Transform Infrared Spectroscopy Analysis
3.4. XRD Analysis
3.5. SEM Analysis
3.6. Antioxidant Activity
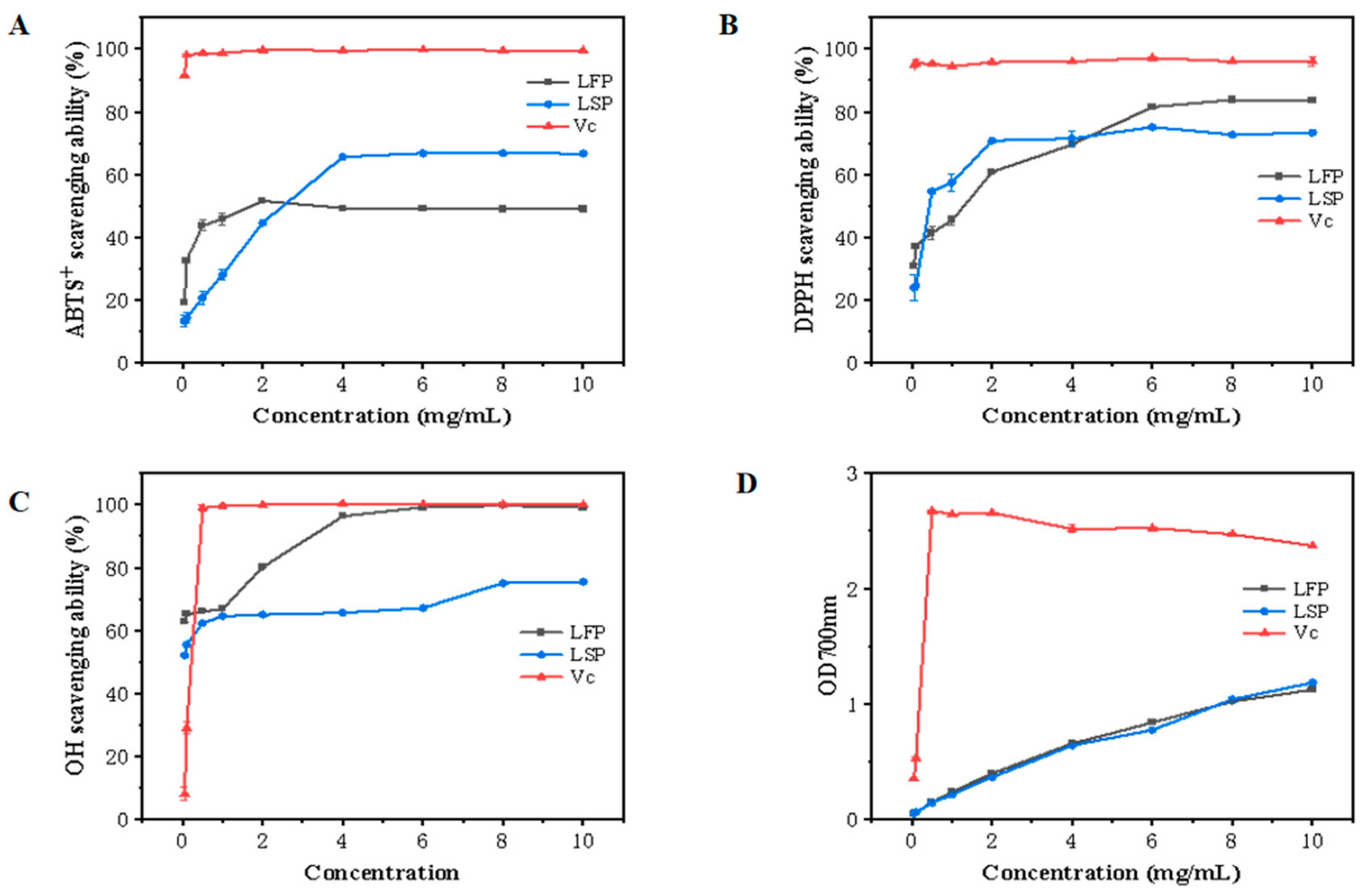
3.6.1. ABTS+ Free Radical Scavenging Activity
3.6.2. DPPH Free Radical Scavenging Assay
3.6.3. Hydroxyl Radical Scavenging Activity
3.6.4. Reducing Power
3.7. Evaluation of LFP and LSP’s Immunomodulatory Activity
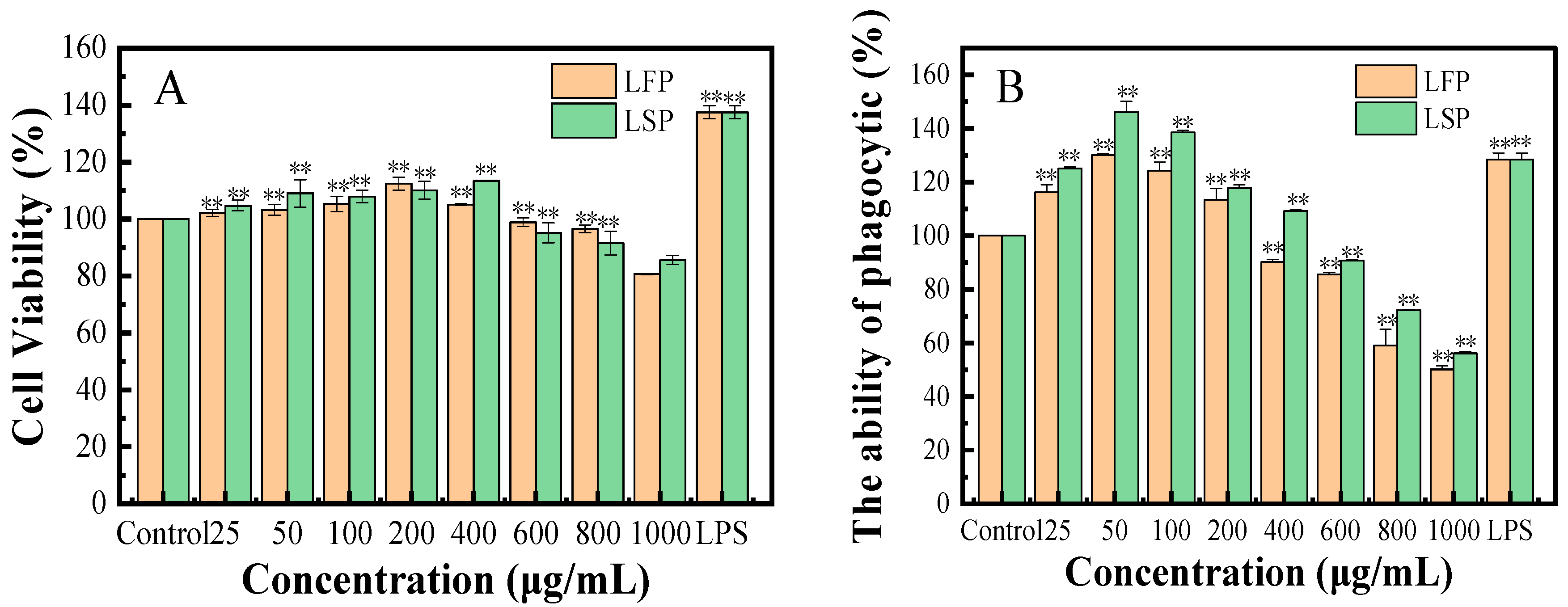
3.7.1. Cell Viability
3.7.2. Effect of LFP and LSP on the Phagocytosis of Macrophage (RAW264.7)
3.7.3. Effect of LFP and LSP on the Migration of RAW264.7
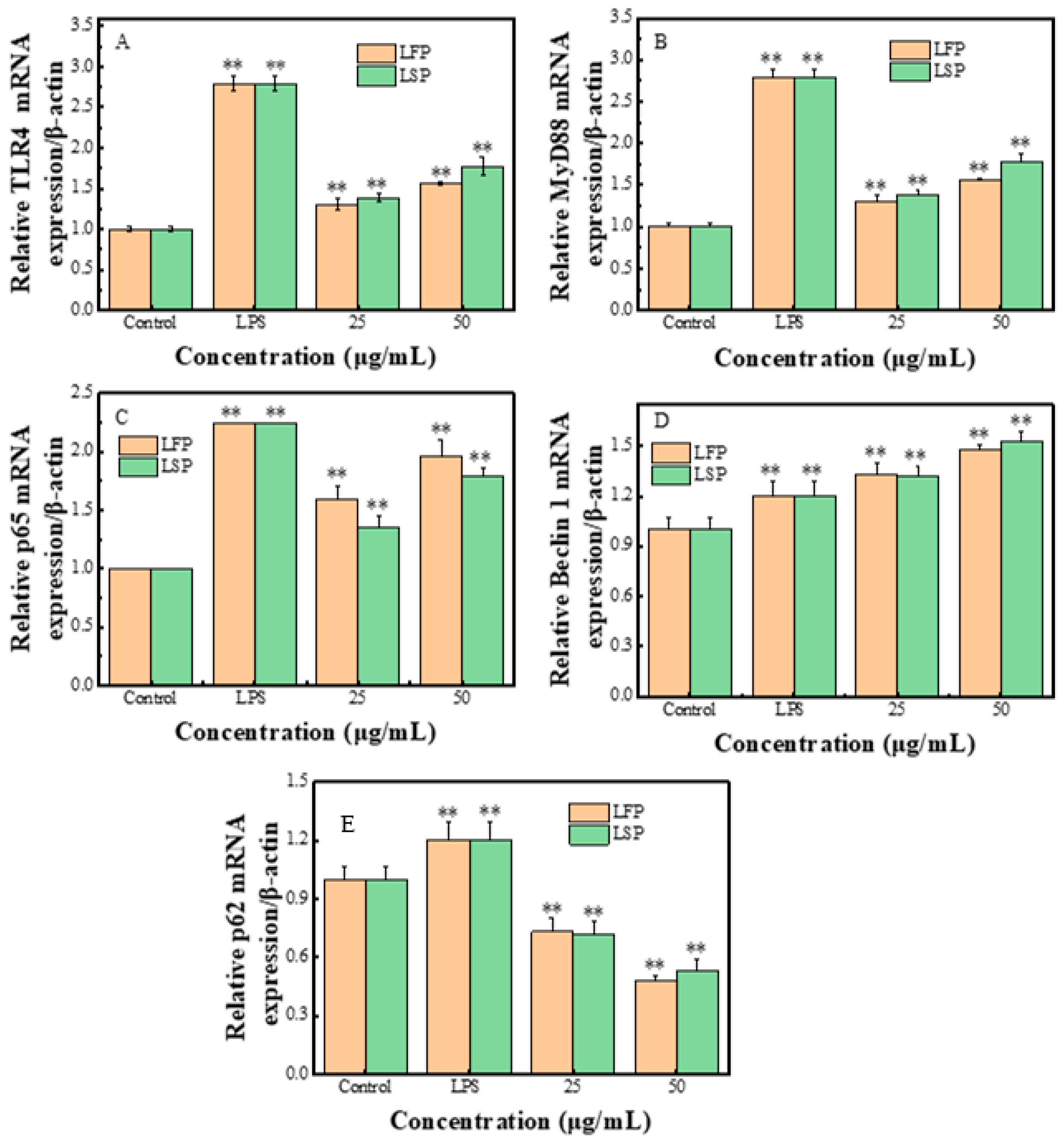
3.7.4. RT-PCR Testing Result
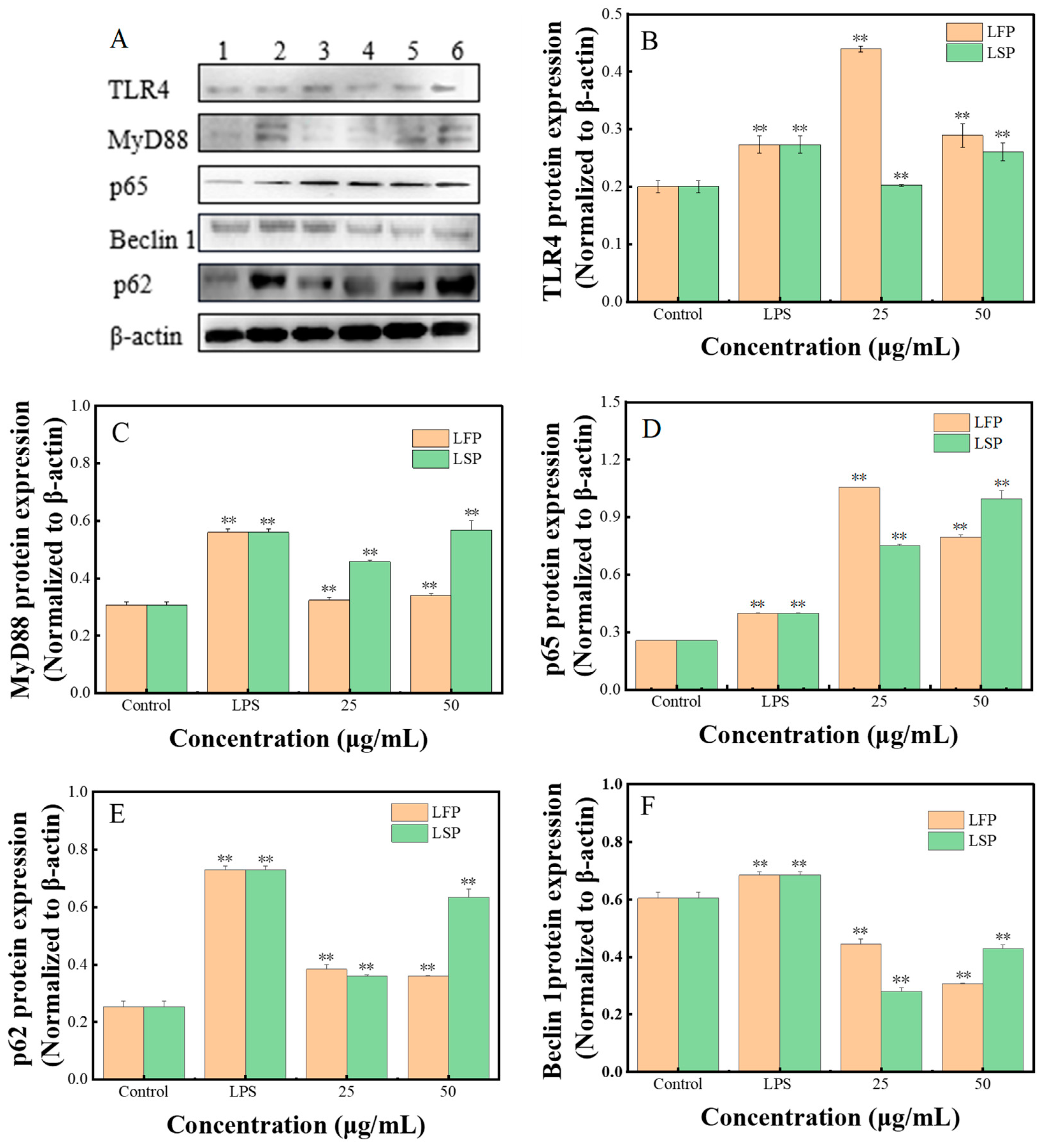
3.7.5. The Expression of the Autophagy Protein and the TLR4/NF-κB Pathway
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Mukherjee, P.; Goswami, D.; Das, B.; Banerjee, S. Nelumbo nucifera: Pharmacological Profile, Metabolite Profiling and Thera-peutic Uses. In Natural Medicines; CRC Press: Boca Raton, FL, USA, 2019; pp. 139–164. [Google Scholar]
- Chen, G.; Zhu, M.; Guo, M. Research advances in traditional and modern use of Nelumbo nucifera: Phytochemicals, health promoting activities and beyond. Crit. Rev. Food Sci. Nutr. 2019, 59 (Suppl. S1), S189–S209. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Lu, X.; Zeng, S.; Huang, X.; Guo, Z.; Zheng, Y.; Tian, Y.; Zheng, B. Nutritional composition, physiological functions and processing of lotus (Nelumbo nucifera Gaertn.) seeds: A review. Phytochem. Rev. 2015, 14, 321–334. [Google Scholar] [CrossRef]
- Fang, X.; Tian, W.; Zhang, X.; Peng, W.; Xiao, H.; Gao, L.; Wang, C.; Gao, Z. Lotus bee pollen polysaccharides significantly relieve intestinal mucosal barrier damage in mice caused by Fluorouracil. J. Food Sci. 2016, 37, 209–214. [Google Scholar]
- Wang, Z.; Sun, Q.; Zhang, H.; Wang, J.; Fu, Q.; Qiao, H.; Wang, Q. Insight into antibacterial mechanism of polysaccharides: A review. LWT 2021, 150, 111929. [Google Scholar] [CrossRef]
- Chen, F.; Huang, G. Preparation and immunological activity of polysaccharides and their derivatives. Int. J. Biol. Macromol. 2018, 112, 211–216. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Chen, J.; Guo, Z.; Zheng, Y.; Rea, M.C.; Su, H.; Zheng, X.; Zheng, B.; Miao, S. Using polysaccharides for the enhancement of functionality of foods: A review. Trends Food Sci. Technol. 2019, 86, 311–327. [Google Scholar] [CrossRef]
- Wang, L.; Liu, H.-M.; Qin, G.-Y. Structure characterization and antioxidant activity of polysaccharides from Chinese quince seed meal. Food Chem. 2017, 234, 314–322. [Google Scholar] [CrossRef]
- Wang, L.; Zhao, Z.; Zhao, H.; Liu, M.; Lin, C.; Li, L.; Ma, B. Pectin polysaccharide from Flos Magnoliae (Xin Yi, Magnolia biondii Pamp. flower buds): Hot-compressed water extraction, purification and partial structural characterization. Food Hydrocoll. 2021, 122, 107061. [Google Scholar] [CrossRef]
- Qiu, J.; Zhang, H.; Wang, Z. Ultrasonic degradation of Polysaccharides from Auricularia auricula and the antioxidant activity of their degradation products. LWT 2019, 113, 108266. [Google Scholar] [CrossRef]
- Wang, L.; Mao, Y.-G.; Zeng, X.; Liu, N.; Niu, C.-F.; Li, X.-X.; Ma, B.-J.; Guo, L.-P.; Yang, X.-L. Structure and Bioactivities of a Novel Polysaccharide Extracted from Dendrobium huoshanense by Subcritical Water. Front. Nutr. 2022, 9, 877871. [Google Scholar] [CrossRef]
- Gao, C.; Wang, Y.; Wang, C.; Wang, Z. Antioxidant and immunological activity in vitro of polysaccharides from Gomphidius rutilus mycelium. Carbohydr. Polym. 2013, 92, 2187–2192. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Guo, Z.; Zhang, Z.; Li, X.; Wang, X.; Liu, Y.; Wang, X. Ferulic acid isolated from propolis inhibits porcine parvovirus rep-lication potentially through Bid-mediate apoptosis. Int. Immunopharmacol. 2020, 83, 106379. [Google Scholar] [CrossRef]
- Zubair, M.; Ekholm, A.; Nybom, H.; Renvert, S.; Widen, C.; Rumpunen, K. Effects of Plantago major L. leaf extracts on oral ep-ithelial cells in a scratch assay. J. Ethnopharmacol. 2012, 141, 825–830. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Chen, F.; Cheng, H.; Huang, G. Modification and application of polysaccharide from traditional Chinese medicine such as Dendrobium officinale. Int. J. Biol. Macromol. 2020, 157, 385–393. [Google Scholar] [CrossRef] [PubMed]
- Fang, C.; Chen, G.; Kan, J. Comparison on characterization and biological activities of Mentha haplocalyx polysaccharides at different solvent extractions. Int. J. Biol. Macromol. 2020, 154, 916–928. [Google Scholar] [CrossRef] [PubMed]
- Hong, T.; Yin, J.-Y.; Nie, S.-P.; Xie, M.-Y. Applications of infrared spectroscopy in polysaccharide structural analysis: Progress, challenge and perspective. Food Chem. X 2021, 12, 100168. [Google Scholar] [CrossRef]
- Sofronov, O.O.; Bakker, H.J. Vibrational Relaxation Dynamics of the Core and Outer Part of Proton-Hydration Clusters. J. Phys. Chem. B 2019, 123, 6222–6228. [Google Scholar] [CrossRef]
- Wang, R.; Chen, P.; Jia, F.; Tang, J.; Ma, F. Optimization of polysaccharides from Panax japonicus C.A. Meyer by RSM and its anti-oxidant activity. Int. J. Biol. Macromol. 2012, 50, 331–336. [Google Scholar] [CrossRef]
- Lee, Y.X.; Ahmad, F.; Kakooei, S.; Azmi, Y.; Kabir, S. Fire-Resistant Properties of Modified Epoxy Based Intumescent Fire-Retardant Coating: Hybridizing of Oyster Shell Powder with Glass Frit. IOP Conf. Ser. Mater. Sci. Eng. 2021, 1176, 012025. [Google Scholar] [CrossRef]
- Liu, X.; Renard, C.M.; Bureau, S.; Le Bourvellec, C. Revisiting the contribution of ATR-FTIR spectroscopy to characterize plant cell wall polysaccharides. Carbohydr. Polym. 2021, 262, 117935. [Google Scholar] [CrossRef]
- Zhao, Z.; Wang, L.; Ruan, Y.; Wen, C.; Ge, M.; Qian, Y.; Ma, B. Physicochemical properties and biological activities of polysac-charides from the peel of Dioscorea opposita Thunb. extracted by four different methods. Food Sci. Hum. Wellness 2013, 12, 130–139. [Google Scholar] [CrossRef]
- Zhao, C.; Li, X.; Miao, J.; Jing, S.; Li, X.; Huang, L.; Gao, W. The effect of different extraction techniques on property and bioactivity of polysaccharides from Dioscorea hemsleyi. Int. J. Biol. Macromol. 2017, 102, 847–856. [Google Scholar] [CrossRef] [PubMed]
- Santos-Sánchez, N.; Salas-Coronado, R.; Villanueva-Cañongo, C.; Hernández-Carlos, B. Antioxidant compounds and their antioxidant mechanism. Antioxidants 2019, 10, 1–29. [Google Scholar]
- Safdar, B.; ZhiHua, P.; Xinqi, L.; Jatoi, M.A.; Rashid, M.T. Influence of different extraction techniques on recovery, purity, antioxidant activities, and microstructure of flaxseed gum. J. Food Sci. 2020, 85, 3168–3182. [Google Scholar] [CrossRef]
- Wang, X.-Q.; Wang, W.; Peng, M.; Zhang, X.-Z. Free radicals for cancer theranostics. Biomaterials 2020, 266, 120474. [Google Scholar] [CrossRef]
- Wang, J.; Hu, S.; Nie, S.; Yu, Q.; Xie, M. Reviews on Mechanisms of In Vitro Antioxidant Activity of Polysaccharides. Oxidative Med. Cell. Longev. 2015, 2016, 5692852. [Google Scholar] [CrossRef]
- Wang, H.; Ma, C.; Sun-Waterhouse, D.; Wang, J.; Waterhouse, G.; Kang, W. Immunoregulatory polysaccharides from Apocynum venetum L. flowers stimulate phagocytosis and cytokine expression via activating the NF-κB/MAPK signaling pathways in RAW264. 7 cells. Food Sci. Hum. Wellness 2022, 11, 806–814. [Google Scholar] [CrossRef]
- Bobadilla, A.; Arévalo, J.; Sarró, E.; Byrne, H.; Maini, P.; Carraro, T.; Balocco, S.; Meseguer, A.; Alarcón, T. In vitro cell migration quantification method for scratch assays. J. R. Soc. Interface 2019, 16, 20180709. [Google Scholar] [CrossRef]
- Song, Y.-R.; Han, A.-R.; Park, S.-G.; Cho, C.-W.; Rhee, Y.-K.; Hong, H.-D. Effect of enzyme-assisted extraction on the physico-chemical properties and bioactive potential of lotus leaf polysaccharides. Int. J. Biol. Macromol. 2020, 153, 169–179. [Google Scholar] [CrossRef]
- Wu, D.-T.; Feng, K.-L.; Huang, L.; Gan, R.-Y.; Hu, Y.-C.; Zou, L. Deep Eutectic Solvent-Assisted Extraction, Partially Structural Characterization, and Bioactivities of Acidic Polysaccharides from Lotus Leaves. Foods 2021, 10, 2330. [Google Scholar] [CrossRef]
- Hwang, Y.-H.; Jang, S.-A.; Lee, A.; Cho, C.-W.; Song, Y.-R.; Hong, H.-D.; Ha, H.; Kim, T. Polysaccharides isolated from lotus leaves (LLEP) exert anti-osteoporotic effects by inhibiting osteoclastogenesis. Int. J. Biol. Macromol. 2020, 161, 449–456. [Google Scholar] [CrossRef] [PubMed]
- Shtein, I.; Bar-On, B.; Popper, Z. Plant and algal structure: From cell walls to biomechanical function. Physiol. Plant. 2018, 164, 56–66. [Google Scholar] [CrossRef] [PubMed]
- Feng, K.-L.; Huang, L.; Wu, D.-T.; Li, F.; Gan, R.-Y.; Qin, W.; Zou, L. Physicochemical properties and in vitro bioactivities of pol-ysaccharides from lotus leaves extracted by different techniques and solvents. J. Food Meas. Characteri-Zation 2022, 16, 1583–1594. [Google Scholar] [CrossRef]
- Ma, G.; Li, G.; Wu, J.; Xiong, W.; Wu, L. Purification, antioxidant and immunomodulatory activities of polysaccharides from a white lotus seedpod. Food Mach. 2021, 37, 156–162. [Google Scholar]
- Jiang, M.-H.; Zhu, L.; Jiang, J.-G. Immunoregulatory actions of polysaccharides from Chinese herbal medicine. Expert Opin. Ther. Targets 2010, 14, 1367–1402. [Google Scholar] [CrossRef]
- Ren, D.; Lin, D.; Alim, A.; Zheng, Q.; Yang, X. Chemical characterization of a novel polysaccharide ASKP-1 from Artemisia sphaerocephala Krasch seed and its macrophage activation via MAPK, PI3k/Akt and NF-κB signaling pathways in RAW264.7 cells. Food Funct. 2017, 8, 1299–1312. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.P.; Yan, F.; Wei, W.; Wang, F.F.; Wang, Z.R.; Nie, J.H.; Jin, M.; Pang, Y.L.; Qin, M.Q.; Wang, L.F. CD38 deficiency protects the retina from ischaemia/reperfusion injury partly via suppression of TLR4/MyD88/NF-κB signalling. Exp. Eye Re-Search 2022, 219, 109058. [Google Scholar] [CrossRef]
- Gerada, C.; Ryan, K. Autophagy, the innate immune response and cancer. Mol. Oncol. 2020, 14, 1913–1929. [Google Scholar] [CrossRef]
- Maiuri, M.C.; Criollo, A.; Kroemer, G. Crosstalk between apoptosis and autophagy within the Beclin 1 interactome. EMBO J. 2010, 29, 515–516. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Forward (5′-3′) | Reverse (5′-3′) |
|---|---|---|
| TLR4 | ATTTCCGCTTCCTGGTCT | GTCATCCCACTTCCTTCCT |
| MyD88 | CCGCCTGTCTCTGTTCTT | GTCCGCTTGTGTCTCCA |
| P65 | ATGCGCTTCCGCTACAA | GTGACCAGGGAGATGCG |
| Beclin 1 | GAGCGATGGTAGTTCTGGA | CCCGATGCTCTTCACCT |
| p62 | CTTTGACTGAGCGACAGCA | GCCACAACCCCAAACTACA |
| β-actin | ACCCCAGCAAGGACACTGAGCAAG | GGCCCCTCCTGTTATTATGGGGGT |
| Sample | LFP | LSP |
|---|---|---|
| Yield (%) | 5.57 ± 0.04 | 2.51 ± 0.03 |
| Carbohydrate (mg/g) | 22.86 ± 0.52 | 41.24 ± 0.94 |
| Protein (mg/g) | 4.63 ± 0.07 | 6.62 ± 0.26 |
| Polyphenol (mg/g) | 2.89 ± 0.02 | 6.66 ± 0.07 |
| Uronic acid (mg/g) | 28.5 ± 0.16 | 34.03 ± 0.35 |
| Sample | LFP | LSP |
|---|---|---|
| Monosaccharide (mol%) | ||
| Fucose | 0.18 | 2.70 |
| Rhamnose | 0.43 | 1.02 |
| Arabinose | 2.26 | 8.15 |
| Glucose | 45.22 | 45.63 |
| Galactose | 32.14 | 20.63 |
| Mannose | 4.28 | 1.44 |
| Fructose | 8.20 | 2.59 |
| Galacturonic acid | 6.28 | 16.45 |
| Glucuronic acid | 1.01 | 1.38 |
| Molecular weight (Da) | ||
| Polymer dispersity index (PDI) | 1.50 | 2.95 |
| Number—average molecular weight (Mn) | 8.23 × 105 | 3.18 × 104 |
| Weight—average molecular weight (Mw) | 1.24 × 106 | 9.37 × 104 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Z.; Wang, L.; Zeng, D.; Ma, X.; Wang, H. Lotus-Flower- and Lotus-Seedpod-Derived Polysaccharide: Structural Characterization and Biological Activity. Polymers 2023, 15, 3828. https://doi.org/10.3390/polym15183828
Zhang Z, Wang L, Zeng D, Ma X, Wang H. Lotus-Flower- and Lotus-Seedpod-Derived Polysaccharide: Structural Characterization and Biological Activity. Polymers. 2023; 15(18):3828. https://doi.org/10.3390/polym15183828
Chicago/Turabian StyleZhang, Zhiqiang, Li Wang, Dai Zeng, Xia Ma, and Hui Wang. 2023. "Lotus-Flower- and Lotus-Seedpod-Derived Polysaccharide: Structural Characterization and Biological Activity" Polymers 15, no. 18: 3828. https://doi.org/10.3390/polym15183828
APA StyleZhang, Z., Wang, L., Zeng, D., Ma, X., & Wang, H. (2023). Lotus-Flower- and Lotus-Seedpod-Derived Polysaccharide: Structural Characterization and Biological Activity. Polymers, 15(18), 3828. https://doi.org/10.3390/polym15183828