
Non-Viral Delivery of Gene Therapy to the Tendon


Abstract
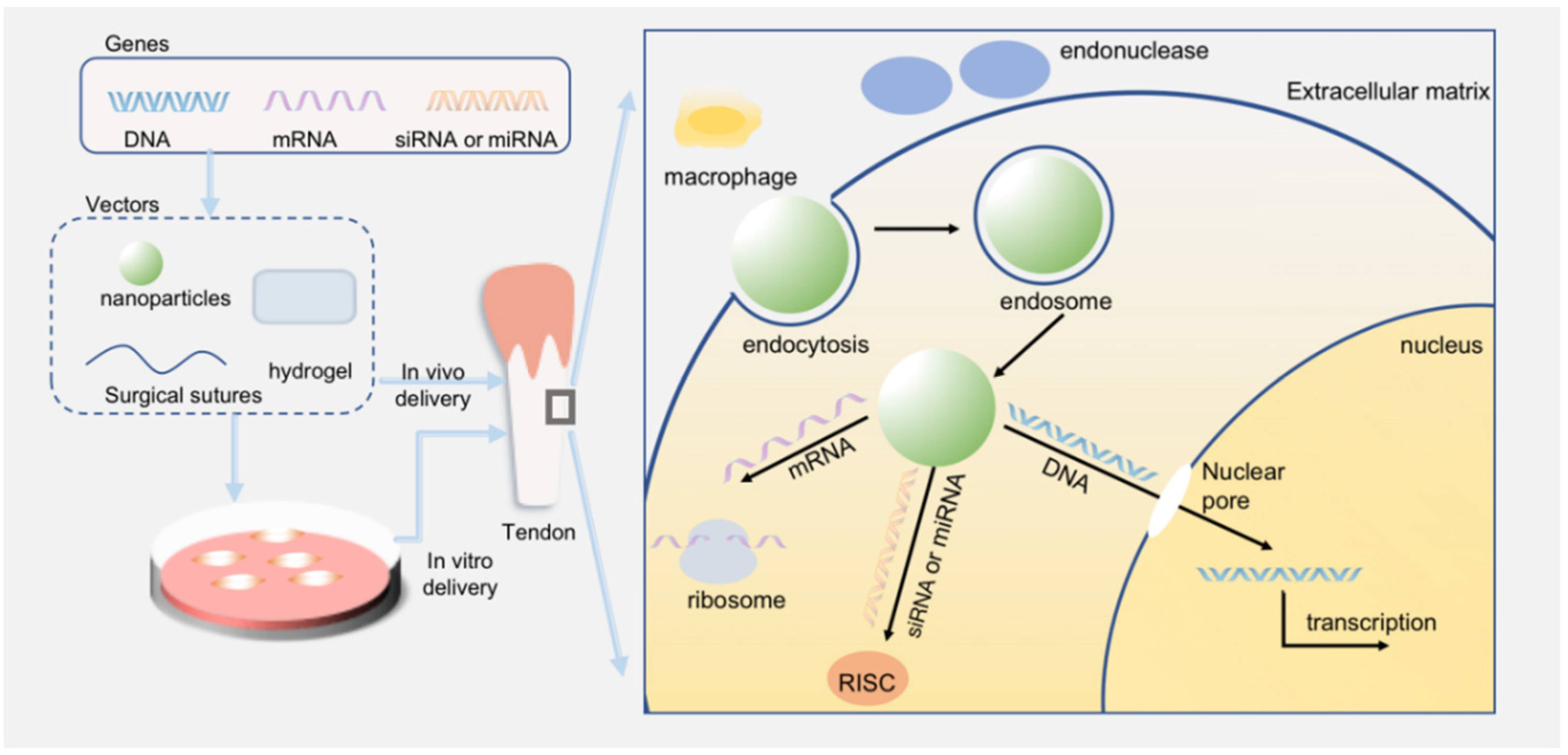
1. Introduction to the Use of Non-Viral Vectors in Tendon Injuries
2. Biological Barriers
- Avoid the degradation of endonucleases and the detection of immune systems;
- Help genes enter cells through endocytosis mediated by receptors;
- Promote lysosome escape;
- Release at a sustainable speed and be able to entrap enough drugs;
- The basic quality of biocapacity, biodegradation, and non-toxicity.
3. Different Forms of Loaded Genes
3.1. DNA
3.2. RNA
3.3. mRNA
3.4. siRNA
3.5. miRNA
4. Non-Viral Vectors for Gene Therapy and Feasibility Analysis
5. The Plasmid
6. Exosomes
7. Inorganic Nanoparticles
7.1. Gold
7.2. Ag
7.3. Silica
8. Lipids and Lipid-Based Nanoparticles
9. Polymers, Polymer-Based Nanoparticles, and Polymer-Modified Novel Tools
9.1. Polymers
9.2. Polymer-Based Nanoparticles
9.3. Polymer-Modified Novel Tools
10. Comparison between Lipid- and Polymer-Based Nanoparticles
11. Hydrogel
12. Other Worthy Delivery Systems
13. Summary and Prospect
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Voleti, P.B.; Buckley, M.R.; Soslowsky, L.J. Tendon healing: Repair and regeneration. Annu. Rev. Biomed. Eng. 2012, 14, 47–71. [Google Scholar] [CrossRef] [PubMed]
- Mienaltowski, M.J.; Birk, D.E. Structure, physiology, and biochemistry of collagens. Adv. Exp. Med. Biol. 2014, 802, 5–29. [Google Scholar] [PubMed]
- Millar, N.L.; Silbernagel, K.G.; Thorborg, K.; Kirwan, P.D.; Galatz, L.M.; Abrams, G.D.; Murrell, G.A.C.; McInnes, I.B.; Rodeo, S.A. Tendinopathy. Nat. Rev. Dis. Primers 2021, 7, 1. [Google Scholar] [CrossRef] [PubMed]
- Titan, A.L.; Foster, D.S.; Chang, J.; Longaker, M.T. Flexor Tendon: Development, Healing, Adhesion Formation, and Contributing Growth Factors. Plast. Reconstr. Surg. 2019, 144, 639e–647e. [Google Scholar] [CrossRef]
- Hafeez, M.N.; d’Avanzo, N.; Russo, V.; di Marzio, L.; Cilurzo, F.; Paolino, D.; Fresta, M.; Barboni, B.; Santos, H.A.; Celia, C. Tendon Tissue Repair in Prospective of Drug Delivery, Regenerative Medicines, and Innovative Bioscaffolds. Stem. Cells Int. 2021, 2021, 1488829. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.B.; Zhou, Y.L.; Wu, Y.F.; Liu, P.Y.; Wang, X.T. Gene therapy strategies to improve strength and quality of flexor tendon healing. Expert Opin. Biol. Ther. 2016, 16, 291–301. [Google Scholar] [CrossRef] [PubMed]
- Loiselle, A.E.; Yukata, K.; Geary, M.B.; Kondabolu, S.; Shi, S.; Jonason, J.H.; Awad, H.A.; O’Keefe, R.J. Development of antisense oligonucleotide (ASO) technology against Tgf-beta signaling to prevent scarring during flexor tendon repair. J. Orthop. Res. 2015, 33, 859–866. [Google Scholar] [CrossRef]
- Tang, J.B. Flexor Tendon Injuries. Clin. Plast. Surg. 2019, 46, 295–306. [Google Scholar] [CrossRef]
- Evans, C.H.; Huard, J. Gene therapy approaches to regenerating the musculoskeletal system. Nat. Rev. Rheumatol. 2015, 11, 234–242. [Google Scholar] [CrossRef]
- Flotte, T.R.; Carter, B.J. Adeno-associated virus vectors for gene therapy. Gene Ther. 1995, 2, 357–362. [Google Scholar]
- Clements, J.E.; Zink, M.C. Molecular biology and pathogenesis of animal lentivirus infections. Clin. Microbiol. Rev. 1996, 9, 100–117. [Google Scholar] [CrossRef] [PubMed]
- Zhan, T.; Rindtorff, N.; Betge, J.; Ebert, M.P.; Boutros, M. CRISPR/Cas9 for cancer research and therapy. Semin. Cancer Biol. 2019, 55, 106–119. [Google Scholar] [CrossRef] [PubMed]
- Aimaletdinov, A.; Mindubaeva, G.; Khalikova, S.; Kabwe, E.; Salmakova, A.; Alexandrova, N.; Rutland, C.; Rizvanov, A.; Zakirova, E. Application of gene therapy in the treatment of superficial digital flexor tendon injury in horses. Open Vet. J. 2020, 10, 261–266. [Google Scholar] [CrossRef] [PubMed]
- Bolous, N.S.; Chen, Y.; Wang, H.; Davidoff, A.M.; Devidas, M.; Jacobs, T.W.; Meagher, M.M.; Nathwani, A.C.; Neufeld, E.J.; Piras, B.A.; et al. The cost-effectiveness of gene therapy for severe hemophilia B: A microsimulation study from the United States perspective. Blood 2021, 138, 1677–1690. [Google Scholar] [CrossRef]
- Locatelli, F.; Thompson, A.A.; Kwiatkowski, J.L.; Porter, J.B.; Thrasher, A.J.; Hongeng, S.; Sauer, M.G.; Thuret, I.; Lal, A.; Algeri, M.; et al. Betibeglogene Autotemcel Gene Therapy for Non-β(0)/β(0) Genotype β-Thalassemia. N. Engl. J. Med. 2022, 386, 415–427. [Google Scholar] [CrossRef]
- Botto, C.; Rucli, M.; Tekinsoy, M.D.; Pulman, J.; Sahel, J.A.; Dalkara, D. Early and late stage gene therapy interventions for inherited retinal degenerations. Prog. Retin. Eye Res. 2022, 86, 100975. [Google Scholar] [CrossRef]
- Mendell, J.R.; Al-Zaidy, S.A.; Rodino-Klapac, L.R.; Goodspeed, K.; Gray, S.J.; Kay, C.N.; Boye, S.L.; Boye, S.E.; George, L.A.; Salabarria, S.; et al. Current Clinical Applications of In Vivo Gene Therapy with AAVs. Mol. Ther. 2021, 29, 464–488. [Google Scholar] [CrossRef]
- De Simone, G.; Quattrocchi, A.; Mancini, B.; di Masi, A.; Nervi, C.; Ascenzi, P. Thalassemias: From gene to therapy. Mol. Aspects Med. 2022, 84, 101028. [Google Scholar] [CrossRef]
- Yin, H.; Kanasty, R.L.; Eltoukhy, A.A.; Vegas, A.J.; Dorkin, J.R.; Anderson, D.G. Non-viral vectors for gene-based therapy. Nat. Rev. Genet. 2014, 15, 541–555. [Google Scholar] [CrossRef]
- Pan, X.; Veroniaina, H.; Su, N.; Sha, K.; Jiang, F.; Wu, Z.; Qi, X. Applications and developments of gene therapy drug delivery systems for genetic diseases. Asian J. Pharm. Sci. 2021, 16, 687–703. [Google Scholar] [CrossRef]
- Jeannerat, A.; Peneveyre, C.; Armand, F.; Chiappe, D.; Hamelin, R.; Scaletta, C.; Hirt-Burri, N.; Roessingh, A.d.; Raffoul, W.; Applegate, L.A.; et al. Hypoxic Incubation Conditions for Optimized Manufacture of Tenocyte-Based Active Pharmaceutical Ingredients of Homologous Standardized Transplant Products in Tendon Regenerative Medicine. Cells 2021, 10, 2872. [Google Scholar] [CrossRef] [PubMed]
- Bulcha, J.T.; Wang, Y.; Ma, H.; Tai, P.W.L.; Gao, G. Viral vector platforms within the gene therapy landscape. Signal Transduct. Target Ther. 2021, 6, 53. [Google Scholar] [CrossRef] [PubMed]
- Bessis, N.; GarciaCozar, F.J.; Boissier, M.C. Immune responses to gene therapy vectors: Influence on vector function and effector mechanisms. Gene Ther. 2004, 11 (Suppl. S1), S10–S17. [Google Scholar] [CrossRef] [PubMed]
- Baum, C.; von Kalle, C.; Staal, F.J.; Li, Z.; Fehse, B.; Schmidt, M.; Weerkamp, F.; Karlsson, S.; Wagemaker, G.; Williams, D.A. Chance or necessity? Insertional mutagenesis in gene therapy and its consequences. Mol. Ther. 2004, 9, 5–13. [Google Scholar] [CrossRef]
- Baum, C.; Kustikova, O.; Modlich, U.; Li, Z.; Fehse, B. Mutagenesis and oncogenesis by chromosomal insertion of gene transfer vectors. Hum. Gene Ther. 2006, 17, 253–263. [Google Scholar] [CrossRef]
- Thomas, C.E.; Ehrhardt, A.; Kay, M.A. Progress and problems with the use of viral vectors for gene therapy. Nat. Rev. Genet. 2003, 4, 346–358. [Google Scholar] [CrossRef]
- Waehler, R.; Russell, S.J.; Curiel, D.T. Engineering targeted viral vectors for gene therapy. Nat. Rev. Genet. 2007, 8, 573–587. [Google Scholar] [CrossRef]
- McCarty, D.M. Self-complementary AAV vectors; Advances and applications. Mol. Ther. 2008, 16, 1648–1656. [Google Scholar] [CrossRef]
- Wu, N.; Ataai, M.M. Production of viral vectors for gene therapy applications. Curr. Opin. Biotechnol. 2000, 11, 205–208. [Google Scholar] [CrossRef]
- Kay, M.A. State-of-the-art gene-based therapies: The road ahead. Nat. Rev. Genet. 2011, 12, 316–328. [Google Scholar] [CrossRef]
- Sachdev, S.; Potočnik, T.; Rems, L.; Miklavčič, D. Revisiting the role of pulsed electric fields in overcoming the barriers to in vivo gene electrotransfer. Bioelectrochemistry 2021, 144, 107994. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.L.; Yang, Q.Q.; Yan, Y.Y.; Zhang, L.; Wang, Q.H.; Ju, F.; Tang, J.B. Gene-Loaded Nanoparticle-Coated Sutures Provide Effective Gene Delivery to Enhance Tendon Healing. Mol. Ther. 2019, 27, 1534–1546. [Google Scholar] [CrossRef] [PubMed]
- Noh, I.; Lee, K.; Rhee, Y.S. Microneedle systems for delivering nucleic acid drugs. J. Pharm. Investig. 2022, 52, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Chan, Y.K.; Wang, S.K.; Chu, C.J.; Copland, D.A.; Letizia, A.J.; Verdera, H.C.; Chiang, J.J.; Sethi, M.; Wang, M.K.; Neidermyer, W.J., Jr.; et al. Engineering adeno-associated viral vectors to evade innate immune and inflammatory responses. Sci. Transl. Med. 2021, 13, eabd3438. [Google Scholar] [CrossRef]
- Hendrikse, S.I.S.; Gras, S.L.; Ellis, A.V. Opportunities and Challenges in DNA-Hybrid Nanomaterials. ACS Nano 2019, 13, 8512–8516. [Google Scholar] [CrossRef]
- Capecchi, M.R. High efficiency transformation by direct microinjection of DNA into cultured mammalian cells. Cell 1980, 22, 479–488. [Google Scholar] [CrossRef]
- Yan, Y.; Liu, X.Y.; Lu, A.; Wang, X.Y.; Jiang, L.X.; Wang, J.C. Non-viral vectors for RNA delivery. J. Control Release 2022, 342, 241–279. [Google Scholar] [CrossRef]
- Martínez-Baz, I.; Trobajo-Sanmartín, C.; Miqueleiz, A.; Guevara, M.; Fernández-Huerta, M.; Burgui, C.; Casado, I.; Portillo, M.E.; Navascués, A.; Ezpeleta, C.; et al. Product-specific COVID-19 vaccine effectiveness against secondary infection in close contacts, Navarre, Spain, April to August 2021. Eurosurveillance 2021, 26, 2100894. [Google Scholar] [CrossRef]
- Haas, E.J.; Angulo, F.J.; McLaughlin, J.M.; Anis, E.; Singer, S.R.; Khan, F.; Brooks, N.; Smaja, M.; Mircus, G.; Pan, K.; et al. Impact and effectiveness of mRNA BNT162b2 vaccine against SARS-CoV-2 infections and COVID-19 cases, hospitalisations, and deaths following a nationwide vaccination campaign in Israel: An observational study using national surveillance data. Lancet 2021, 397, 1819–1829. [Google Scholar] [CrossRef]
- Ginn, S.L.; Alexander, I.E.; Edelstein, M.L.; Abedi, M.R.; Wixon, J. Gene therapy clinical trials worldwide to 2012—An update. J. Gene Med. 2013, 15, 65–77. [Google Scholar] [CrossRef]
- Jirikowski, G.F.; Sanna, P.P.; Maciejewski-Lenoir, D.; Bloom, F.E. Reversal of diabetes insipidus in Brattleboro rats: Intrahypothalamic injection of vasopressin mRNA. Science 1992, 255, 996–998. [Google Scholar] [CrossRef] [PubMed]
- Conry, R.M.; LoBuglio, A.F.; Wright, M.; Sumerel, L.; Pike, M.J.; Johanning, F.; Benjamin, R.; Lu, D.; Curiel, D.T. Characterization of a messenger RNA polynucleotide vaccine vector. Cancer Res. 1995, 55, 1397–1400. [Google Scholar] [PubMed]
- Boczkowski, D.; Nair, S.K.; Snyder, D.; Gilboa, E. Dendritic cells pulsed with RNA are potent antigen-presenting cells in vitro and in vivo. J. Exp. Med. 1996, 184, 465–472. [Google Scholar] [CrossRef] [PubMed]
- Hoerr, I.; Obst, R.; Rammensee, H.G.; Jung, G. In vivo application of RNA leads to induction of specific cytotoxic T lymphocytes and antibodies. Eur. J. Immunol. 2000, 30, 1–7. [Google Scholar] [CrossRef]
- Karikó, K.; Muramatsu, H.; Keller, J.M.; Weissman, D. Increased erythropoiesis in mice injected with submicrogram quantities of pseudouridine-containing mRNA encoding erythropoietin. Mol. Ther. 2012, 20, 948–953. [Google Scholar] [CrossRef] [PubMed]
- Zangi, L.; Lui, K.O.; von Gise, A.; Ma, Q.; Ebina, W.; Ptaszek, L.M.; Später, D.; Xu, H.; Tabebordbar, M.; Gorbatov, R.; et al. Modified mRNA directs the fate of heart progenitor cells and induces vascular regeneration after myocardial infarction. Nat. Biotechnol. 2013, 31, 898–907. [Google Scholar] [CrossRef] [PubMed]
- Kormann, M.S.; Hasenpusch, G.; Aneja, M.K.; Nica, G.; Flemmer, A.W.; Herber-Jonat, S.; Huppmann, M.; Mays, L.E.; Illenyi, M.; Schams, A.; et al. Expression of therapeutic proteins after delivery of chemically modified mRNA in mice. Nat. Biotechnol. 2011, 29, 154–157. [Google Scholar] [CrossRef] [PubMed]
- Breunig, M.; Lungwitz, U.; Liebl, R.; Fontanari, C.; Klar, J.; Kurtz, A.; Blunk, T.; Goepferich, A. Gene delivery with low molecular weight linear polyethylenimines. J. Gene Med. 2005, 7, 1287–1298. [Google Scholar] [CrossRef]
- Cohen, R.N.; van der Aa, M.A.; Macaraeg, N.; Lee, A.P.; Szoka, F.C., Jr. Quantification of plasmid DNA copies in the nucleus after lipoplex and polyplex transfection. J. Control Release 2009, 135, 166–174. [Google Scholar] [CrossRef]
- Ramamoorth, M.; Narvekar, A. Non viral vectors in gene therapy—An overview. J. Clin. Diagn. Res. 2015, 9, GE01–GE06. [Google Scholar] [CrossRef]
- Layzer, J.M.; McCaffrey, A.P.; Tanner, A.K.; Huang, Z.A.N.; Kay, M.A.; Sullenger, B.A. In vivo activity of nuclease-resistant siRNAs. RNA 2004, 10, 766–771. [Google Scholar] [CrossRef] [PubMed]
- Jackson, A.L.; Linsley, P.S. Recognizing and avoiding siRNA off-target effects for target identification and therapeutic application. Nat. Rev. Drug Discov. 2010, 9, 57–67. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, D.N.; Mahon, K.P.; Chikh, G.; Kim, P.; Chung, H.; Vicari, A.P.; Love, K.T.; Goldberg, M.; Chen, S.; Krieg, A.M.; et al. Lipid-derived nanoparticles for immunostimulatory RNA adjuvant delivery. Proc. Natl. Acad. Sci. USA 2012, 109, E797–E803. [Google Scholar] [CrossRef] [PubMed]
- Gargano, G.; Oliviero, A.; Oliva, F.; Maffulli, N. Small interfering RNAs in tendon homeostasis. Br. Med. Bull. 2021, 138, 58–67. [Google Scholar] [CrossRef] [PubMed]
- Deleavey, G.F.; Watts, J.K.; Damha, M.J. Chemical modification of siRNA. Curr. Protoc. Nucleic Acid Chem. 2009, 39, 16.3. [Google Scholar] [CrossRef]
- Park, J.; Ries, J.; Gelse, K.; Kloss, F.; von der Mark, K.; Wiltfang, J.; Neukam, F.W.; Schneider, H. Bone regeneration in critical size defects by cell-mediated BMP-2 gene transfer: A comparison of adenoviral vectors and liposomes. Gene Ther. 2003, 10, 1089–1098. [Google Scholar] [CrossRef]
- Lin, L.; Chen, L.; Wang, H.; Wei, X.; Fu, X.; Zhang, J.; Ma, K.; Zhou, C.; Yu, C. Adenovirus-mediated transfer of siRNA against Runx2/Cbfa1 inhibits the formation of heterotopic ossification in animal model. Biochem. Biophys. Res. Commun. 2006, 349, 564–572. [Google Scholar] [CrossRef]
- Richardson, S.H.; Starborg, T.; Lu, Y.; Humphries, S.M.; Meadows, R.S.; Kadler, K.E. Tendon development requires regulation of cell condensation and cell shape via cadherin-11-mediated cell-cell junctions. Mol. Cell Biol. 2007, 27, 6218–6228. [Google Scholar] [CrossRef]
- Gagarina, V.; Carlberg, A.L.; Pereira-Mouries, L.; Hall, D.J. Cartilage oligomeric matrix protein protects cells against death by elevating members of the IAP family of survival proteins. J. Biol. Chem. 2008, 283, 648–659. [Google Scholar] [CrossRef]
- Sun, H.B.; Li, Y.; Fung, D.T.; Majeska, R.J.; Schaffler, M.B.; Flatow, E.L. Coordinate regulation of IL-1beta and MMP-13 in rat tendons following subrupture fatigue damage. Clin. Orthop. Relat. Res. 2008, 466, 1555–1561. [Google Scholar] [CrossRef]
- Sakamoto, K.; Owada, Y.; Shikama, Y.; Wada, I.; Waguri, S.; Iwamoto, T.; Kimura, J. Involvement of Na+/Ca2+ exchanger in migration and contraction of rat cultured tendon fibroblasts. J. Physiol. 2009, 587, 5345–5359. [Google Scholar] [CrossRef] [PubMed]
- Xue, M.; Smith, M.M.; Little, C.B.; Sambrook, P.; March, L.; Jackson, C.J. Activated protein C mediates a healing phenotype in cultured tenocytes. J. Cell Mol. Med. 2009, 13, 749–757. [Google Scholar] [CrossRef] [PubMed]
- Xue, T.; Mao, Z.; Lin, L.; Hou, Y.; Wei, X.; Fu, X.; Zhang, J.; Yu, C. Non-virus-mediated transfer of siRNAs against Runx2 and Smad4 inhibit heterotopic ossification in rats. Gene Ther. 2010, 17, 370–379. [Google Scholar] [CrossRef][Green Version]
- Lu, P.; Zhang, G.R.; Song, X.H.; Zou, X.H.; Wang, L.L.; Ouyang, H.W. Col V siRNA engineered tenocytes for tendon tissue engineering. PLoS ONE 2011, 6, e21154. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Fu, Y.; Tong, W.; Geng, Y.; Lui, P.P.; Tang, T.; Zhang, X.; Dai, K. Uniaxial mechanical tension promoted osteogenic differentiation of rat tendon-derived stem cells (rTDSCs) via the Wnt5a-RhoA pathway. J. Cell Biochem. 2012, 113, 3133–3142. [Google Scholar] [CrossRef] [PubMed]
- Cui, Q.; Fu, S.; Li, Z. Hepatocyte growth factor inhibits TGF-β1-induced myofibroblast differentiation in tendon fibroblasts: Role of AMPK signaling pathway. J. Physiol. Sci. 2013, 63, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Nakahara, H.; Hasegawa, A.; Otabe, K.; Ayabe, F.; Matsukawa, T.; Onizuka, N.; Ito, Y.; Ozaki, T.; Lotz, M.K.; Asahara, H. Transcription factor Mohawk and the pathogenesis of human anterior cruciate ligament degradation. Arthritis Rheum. 2013, 65, 2081–2089. [Google Scholar] [CrossRef]
- Ruan, H.; Liu, S.; Li, F.; Li, X.; Fan, C. Prevention of Tendon Adhesions by ERK2 Small Interfering RNAs. Int. J. Mol. Sci. 2013, 14, 4361–4371. [Google Scholar] [CrossRef]
- Chen, L.; Jiang, C.; Tiwari, S.R.; Shrestha, A.; Xu, P.; Liang, W.; Sun, Y.; He, S.; Cheng, B. TGIF1 Gene Silencing in Tendon-Derived Stem Cells Improves the Tendon-to-Bone Insertion Site Regeneration. Cell Physiol. Biochem. 2015, 37, 2101–2114. [Google Scholar] [CrossRef]
- Chen, L.; Liu, J.; Tao, X.; Wang, G.; Wang, Q.; Liu, X. The role of Pin1 protein in aging of human tendon stem/progenitor cells. Biochem. Biophys. Res. Commun. 2015, 464, 487–492. [Google Scholar] [CrossRef]
- Chen, W.; Tang, H.; Zhou, M.; Hu, C.; Zhang, J.; Tang, K. Dexamethasone inhibits the differentiation of rat tendon stem cells into tenocytes by targeting the scleraxis gene. J. Steroid. Biochem. Mol. Biol. 2015, 152, 16–24. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Chen, L.; Sun, L.; Chen, H.; Sun, Y.; Jiang, C.; Cheng, B. Silencing of TGIF1 in bone mesenchymal stem cells applied to the post-operative rotator cuff improves both functional and histologic outcomes. J. Mol. Histol. 2015, 46, 241–249. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, Q.; Wei, X.; Xu, J.; Chen, Q.; Song, S.; Lu, Z.; Wang, Z. Targeted knockout of TNF-α by injection of lentivirus-mediated siRNA into the subacromial bursa for the treatment of subacromial bursitis in rats. Mol. Med. Rep. 2015, 12, 4389–4395. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Chen, S.; Jiang, S.; Zheng, W.; Tu, B.; Liu, S.; Ruan, H.; Fan, C. RelA/p65 inhibition prevents tendon adhesion by modulating inflammation, cell proliferation, and apoptosis. Cell Death Dis. 2017, 8, e2710. [Google Scholar] [CrossRef]
- Jamil, S.; Mousavizadeh, R.; Roshan-Moniri, M.; Tebbutt, S.J.; McCormack, R.G.; Duronio, V.; Scott, A. Angiopoietin-like 4 Enhances the Proliferation and Migration of Tendon Fibroblasts. Med. Sci. Sports Exerc. 2017, 49, 1769–1777. [Google Scholar] [CrossRef]
- Freeberg, M.A.T.; Farhat, Y.M.; Easa, A.; Kallenbach, J.G.; Malcolm, D.W.; Buckley, M.R.; Benoit, D.S.W.; Awad, H.A. Serpine1 Knockdown Enhances MMP Activity after Flexor Tendon Injury in Mice: Implications for Adhesions Therapy. Sci. Rep. 2018, 8, 5810. [Google Scholar] [CrossRef]
- Nichols, A.E.C.; Settlage, R.E.; Werre, S.R.; Dahlgren, L.A. Novel roles for scleraxis in regulating adult tenocyte function. BMC Cell Biol. 2018, 19, 14. [Google Scholar] [CrossRef]
- Shukunami, C.; Takimoto, A.; Nishizaki, Y.; Yoshimoto, Y.; Tanaka, S.; Miura, S.; Watanabe, H.; Sakuma, T.; Yamamoto, T.; Kondoh, G.; et al. Scleraxis is a transcriptional activator that regulates the expression of Tenomodulin, a marker of mature tenocytes and ligamentocytes. Sci. Rep. 2018, 8, 3155. [Google Scholar] [CrossRef]
- Xu, H.; Liu, F. Downregulation of FOXP1 correlates with tendon stem/progenitor cells aging. Biochem. Biophys. Res. Commun. 2018, 504, 96–102. [Google Scholar] [CrossRef]
- Jackson, J.E.; Kopecki, Z.; Anderson, P.J.; Cowin, A.J. Increasing the level of cytoskeletal protein Flightless I reduces adhesion formation in a murine digital flexor tendon model. J. Orthop. Surg. Res. 2020, 15, 362. [Google Scholar] [CrossRef]
- Liao, X.; Falcon, N.D.; Mohammed, A.A.; Paterson, Y.Z.; Mayes, A.G.; Guest, D.J.; Saeed, A. Synthesis and Formulation of Four-Arm PolyDMAEA-siRNA Polyplex for Transient Downregulation of Collagen Type III Gene Expression in TGF-β1 Stimulated Tenocyte Culture. ACS Omega 2020, 5, 1496–1505. [Google Scholar] [CrossRef] [PubMed]
- Deshmukh, V.; Seo, T.; O’Green, A.L.; Ibanez, M.; Hofilena, B.; Kc, S.; Stewart, J.; Dellamary, L.; Chiu, K.; Ghias, A.; et al. SM04755, a small-molecule inhibitor of the Wnt pathway, as a potential topical treatment for tendinopathy. J. Orthop. Res. 2021, 39, 2048–2061. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhou, Z.; Liu, Y.; Wang, Z.; Kang, Y. Inhibition of Smad3 promotes the healing of rotator cuff injury in a rat model. J. Orthop. Res. 2021, 39, 204–218. [Google Scholar] [CrossRef] [PubMed]
- Xu, K.; Shao, Y.; Xia, Y.; Qian, Y.; Jiang, N.; Liu, X.; Yang, L.; Wang, C. Tenascin-C regulates migration of SOX10 tendon stem cells via integrin-α9 for promoting patellar tendon remodeling. Biofactors 2021, 47, 768–777. [Google Scholar] [CrossRef]
- Cai, C.; Zhang, X.; Li, Y.; Liu, X.; Wang, S.; Lu, M.; Yan, X.; Deng, L.; Liu, S.; Wang, F.; et al. Self-Healing Hydrogel Embodied with Macrophage-Regulation and Responsive-Gene-Silencing Properties for Synergistic Prevention of Peritendinous Adhesion. Adv. Mater. 2022, 34, e2106564. [Google Scholar] [CrossRef]
- Wang, C.; Zhou, Z.; Song, W.; Cai, Z.; Ding, Z.; Chen, D.; Xia, F.; He, Y. Inhibition of IKKβ/NF-κB signaling facilitates tendinopathy healing by rejuvenating inflamm-aging induced tendon-derived stem/progenitor cell senescence. Mol. Ther. Nucleic Acids 2022, 27, 562–576. [Google Scholar] [CrossRef]
- Yang, Q.Q.; Zhang, L.; Zhou, Y.L.; Tang, J.B. Morphological changes of macrophages and their potential contribution to tendon healing. Colloids Surf B Biointerfaces 2022, 209, 112145. [Google Scholar] [CrossRef]
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef]
- Berezikov, E.; Guryev, V.; van de Belt, J.; Wienholds, E.; Plasterk, R.H.; Cuppen, E. Phylogenetic shadowing and computational identification of human microRNA genes. Cell 2005, 120, 21–24. [Google Scholar] [CrossRef]
- Jakob, P.; Landmesser, U. Role of microRNAs in stem/progenitor cells and cardiovascular repair. Cardiovasc. Res. 2012, 93, 614–622. [Google Scholar] [CrossRef]
- Ambros, V. The functions of animal microRNAs. Nature 2004, 431, 350–355. [Google Scholar] [CrossRef] [PubMed]
- Esau, C.C.; Monia, B.P. Therapeutic potential for microRNAs. Adv. Drug Deliv. Rev. 2007, 59, 101–114. [Google Scholar] [CrossRef] [PubMed]
- Friedman, R.C.; Farh, K.K.; Burge, C.B.; Bartel, D.P. Most mammalian mRNAs are conserved targets of microRNAs. Genome Res. 2009, 19, 92–105. [Google Scholar] [CrossRef] [PubMed]
- Cai, X.; Cai, M.; Lou, L. Identification of differentially expressed genes and small molecule drugs for the treatment of tendinopathy using microarray analysis. Mol. Med. Rep. 2015, 11, 3047–3054. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Xu, Q.; Sun, W.X.; Zhang, Z.F. High expression of VEGFA in MSCs promotes tendon-bone healing of rotator cuff tear via microRNA-205–5p. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 4081–4088. [Google Scholar] [PubMed]
- Thankam, F.G.; Boosani, C.S.; Dilisio, M.F.; Agrawal, D.K. MicroRNAs associated with inflammation in shoulder tendinopathy and glenohumeral arthritis. Mol. Cell Biochem. 2018, 437, 81–97. [Google Scholar] [CrossRef]
- Thankam, F.G.; Boosani, C.S.; Dilisio, M.F.; Gross, R.M.; Agrawal, D.K. Genes interconnecting AMPK and TREM-1 and associated microRNAs in rotator cuff tendon injury. Mol. Cell Biochem. 2019, 454, 97–109. [Google Scholar] [CrossRef]
- Chen, Q.; Lu, H.; Yang, H. Chitosan inhibits fibroblasts growth in Achilles tendon via TGF-β1/Smad3 pathway by miR-29b. Int. J. Clin. Exp. Pathol. 2014, 7, 8462–8470. [Google Scholar]
- Marie, C.; Vandermeulen, G.; Quiviger, M.; Richard, M.; Préat, V.; Scherman, D. pFARs, plasmids free of antibiotic resistance markers, display high-level transgene expression in muscle, skin and tumour cells. J. Gene Med. 2010, 12, 323–332. [Google Scholar] [CrossRef]
- Gantenbein, B.; Tang, S.; Guerrero, J.; Higuita-Castro, N.; Salazar-Puerta, A.I.; Croft, A.S.; Gazdhar, A.; Purmessur, D. Non-viral Gene Delivery Methods for Bone and Joints. Front. Bioeng. Biotechnol. 2020, 8, 598466. [Google Scholar] [CrossRef]
- Hardee, C.L.; Arevalo-Soliz, L.M.; Hornstein, B.D.; Zechiedrich, L. Advances in Non-Viral DNA Vectors for Gene Therapy. Genes 2017, 8, 65. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, E.G. Nonviral DNA vectors for immunization and therapy: Design and methods for their obtention. J. Mol. Med. 2004, 82, 500–509. [Google Scholar] [CrossRef] [PubMed]
- Mairhofer, J.; Grabherr, R. Rational vector design for efficient non-viral gene delivery: Challenges facing the use of plasmid DNA. Mol. Biotechnol. 2008, 39, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Cranenburgh, R.M.; Hanak, J.A.; Williams, S.G.; Sherratt, D.J. Escherichia coli strains that allow antibiotic-free plasmid selection and maintenance by repressor titration. Nucleic Acids Res. 2001, 29, E26. [Google Scholar] [CrossRef]
- Wang, C.; Song, W.; Chen, B.; Liu, X.; He, Y. Exosomes Isolated from Adipose-Derived Stem Cells: A New Cell-Free Approach to Prevent the Muscle Degeneration Associated with Torn Rotator Cuffs. Am. J. Sports Med. 2019, 47, 3247–3255. [Google Scholar] [CrossRef]
- El-Andaloussi, S.; Lee, Y.; Lakhal-Littleton, S.; Li, J.; Seow, Y.; Gardiner, C.; Alvarez-Erviti, L.; Sargent, I.L.; Wood, M.J. Exosome-mediated delivery of siRNA in vitro and in vivo. Nat. Protoc. 2012, 7, 2112–2126. [Google Scholar] [CrossRef]
- Wassmer, S.J.; Carvalho, L.S.; Gyorgy, B.; Vandenberghe, L.H.; Maguire, C.A. Exosome-associated AAV2 vector mediates robust gene delivery into the murine retina upon intravitreal injection. Sci. Rep. 2017, 7, 45329. [Google Scholar] [CrossRef]
- Caoduro, C.; Hervouet, E.; Girard-Thernier, C.; Gharbi, T.; Boulahdour, H.; Delage-Mourroux, R.; Pudlo, M. Carbon nanotubes as gene carriers: Focus on internalization pathways related to functionalization and properties. Acta Biomater. 2017, 49, 36–44. [Google Scholar] [CrossRef]
- Liu, J.; Cui, L.; Losic, D. Graphene and graphene oxide as new nanocarriers for drug delivery applications. Acta Biomater. 2013, 9, 9243–9257. [Google Scholar] [CrossRef]
- Li, L.L.; Wu, P.; Hwang, K.; Lu, Y. An exceptionally simple strategy for DNA-functionalized up-conversion nanoparticles as biocompatible agents for nanoassembly, DNA delivery, and imaging. J. Am. Chem. Soc. 2013, 135, 2411–2414. [Google Scholar] [CrossRef]
- Wang, Y.; Zhao, J.; Liu, Y.; Liu, G.; Ding, S.; Li, Y.; Xia, J.; Li, H. Synergy between plasmonic and sites on gold nanoparticle-modified bismuth-rich bismuth oxybromide nanotubes for the efficient photocatalytic CC coupling synthesis of ethane. J. Colloid Interface Sci. 2022, 616, 649–658. [Google Scholar] [CrossRef] [PubMed]
- Figueroa, E.R.; Lin, A.Y.; Yan, J.; Luo, L.; Foster, A.E.; Drezek, R.A. Optimization of PAMAM-gold nanoparticle conjugation for gene therapy. Biomaterials 2014, 35, 1725–1734. [Google Scholar] [CrossRef] [PubMed]
- Jeon, H.J.; Yi, S.C.; Oh, S.G. Preparation and antibacterial effects of Ag-SiO2 thin films by sol-gel method. Biomaterials 2003, 24, 4921–4928. [Google Scholar] [CrossRef]
- Samberg, M.E.; Oldenburg, S.J.; Monteiro-Riviere, N.A. Evaluation of silver nanoparticle toxicity in skin in vivo and keratinocytes in vitro. Environ. Health Perspect. 2010, 118, 407–413. [Google Scholar] [CrossRef]
- Asharani, P.V.; Hande, M.P.; Valiyaveettil, S. Anti-proliferative activity of silver nanoparticles. BMC Cell Biol. 2009, 10, 65. [Google Scholar] [CrossRef]
- Foldbjerg, R.; Olesen, P.; Hougaard, M.; Dang, D.A.; Hoffmann, H.J.; Autrup, H. PVP-coated silver nanoparticles and silver ions induce reactive oxygen species, apoptosis and necrosis in THP-1 monocytes. Toxicol. Lett. 2009, 190, 156–162. [Google Scholar] [CrossRef]
- Cheung, T.S.; Lau, P.M.; Lu, H.; Ho, H.P.; Lui, P.P.Y.; Kong, S.K. Cytotoxic and sublethal effects of silver nanoparticles on tendon-derived stem cells—Implications for tendon engineering. Toxicol. Res. 2016, 5, 318–330. [Google Scholar] [CrossRef]
- Radu, D.R.; Lai, C.Y.; Jeftinija, K.; Rowe, E.W.; Jeftinija, S.; Lin, V.S. A polyamidoamine dendrimer-capped mesoporous silica nanosphere-based gene transfection reagent. J. Am. Chem. Soc. 2004, 126, 13216–13217. [Google Scholar] [CrossRef]
- Chang, F.P.; Kuang, L.Y.; Huang, C.A.; Jane, W.N.; Hung, Y.; Hsing, Y.C.; Mou, C.Y. A simple plant gene delivery system using mesoporous silica nanoparticles as carriers. J. Mater. Chem. B 2013, 1, 5279–5287. [Google Scholar] [CrossRef]
- Kar, M.; Vijayakumar, P.S.; Prasad, B.L.; Gupta, S.S. Synthesis and characterization of poly-L-lysine-grafted silica nanoparticles synthesized via NCA polymerization and click chemistry. Langmuir 2010, 26, 5772–5781. [Google Scholar] [CrossRef]
- Hartono, S.B.; Gu, W.; Kleitz, F.; Liu, J.; He, L.; Middelberg, A.P.; Yu, C.; Lu, G.Q.; Qiao, S.Z. Poly-L-lysine functionalized large pore cubic mesostructured silica nanoparticles as biocompatible carriers for gene delivery. ACS Nano 2012, 6, 2104–2117. [Google Scholar] [CrossRef] [PubMed]
- Xia, T.; Kovochich, M.; Liong, M.; Meng, H.; Kabehie, S.; George, S.; Zink, J.I.; Nel, A.E. Polyethyleneimine coating enhances the cellular uptake of mesoporous silica nanoparticles and allows safe delivery of siRNA and DNA constructs. ACS Nano 2009, 3, 3273–3286. [Google Scholar] [CrossRef] [PubMed]
- Suwalski, A.; Dabboue, H.; Delalande, A.; Bensamoun, S.F.; Canon, F.; Midoux, P.; Saillant, G.; Klatzmann, D.; Salvetat, J.P.; Pichon, C. Accelerated Achilles tendon healing by PDGF gene delivery with mesoporous silica nanoparticles. Biomaterials 2010, 31, 5237–5245. [Google Scholar] [CrossRef] [PubMed]
- Brevet, D.; Hocine, O.; Delalande, A.; Raehm, L.; Charnay, C.; Midoux, P.; Durand, J.O.; Pichon, C. Improved gene transfer with histidine-functionalized mesoporous silica nanoparticles. Int. J. Pharm. 2014, 471, 197–205. [Google Scholar] [CrossRef]
- Manni, L.S.; Fong, W.K.; Mezzenga, R. Lipid-based mesophases as matrices for nanoscale reactions. Nanoscale Horiz. 2020, 5, 914–927. [Google Scholar] [CrossRef]
- Paunovska, K.; Loughrey, D.; Dahlman, J.E. Drug delivery systems for RNA therapeutics. Nat. Rev. Genet 2022, 23, 265–280. [Google Scholar] [CrossRef]
- Fraley, R.; Subramani, S.; Berg, P.; Papahadjopoulos, D. Introduction of liposome-encapsulated SV40 DNA into cells. J. Biol. Chem. 1980, 255, 10431–10435. [Google Scholar] [CrossRef]
- Qin, Y.; Tian, Y.; Liu, Y.; Li, D.; Zhang, H.; Yang, Y.; Qi, J.; Wang, H.; Gan, L. Hyaluronic acid-modified cationic niosomes for ocular gene delivery: Improving transfection efficiency in retinal pigment epithelium. J. Pharm. Pharmacol. 2018, 70, 1139–1151. [Google Scholar] [CrossRef]
- Zylberberg, C.; Gaskill, K.; Pasley, S.; Matosevic, S. Engineering liposomal nanoparticles for targeted gene therapy. Gene Ther. 2017, 24, 441–452. [Google Scholar] [CrossRef]
- Khurana, A.; Allawadhi, P.; Khurana, I.; Allwadhi, S.; Weiskirchen, R.; Banothu, A.K.; Chhabra, D.; Joshi, K.; Bharani, K.K. Role of nanotechnology behind the success of mRNA vaccines for COVID-19. Nano Today 2021, 38, 101142. [Google Scholar] [CrossRef]
- Huang, H.; Zhang, C.; Yang, S.; Xiao, W.; Zheng, Q.; Song, X. The investigation of mRNA vaccines formulated in liposomes administrated in multiple routes against SARS-CoV-2. J. Control Release 2021, 335, 449–456. [Google Scholar] [CrossRef] [PubMed]
- Ermilova, I.; Swenson, J. DOPC versus DOPE as a helper lipid for gene-therapies: Molecular dynamics simulations with DLin-MC3-DMA. Phys. Chem. Chem. Phys. 2020, 22, 28256–28268. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, N.; Shino, K.; Natsuume, T.; Horibe, S.; Matsumoto, N.; Kaneda, Y.; Ochi, T. Early biological effect of in vivo gene transfer of platelet-derived growth factor (PDGF)-B into healing patellar ligament. Gene Ther. 1998, 5, 1165–1170. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.T.; Liu, P.Y.; Tang, J.B. Tendon healing in vitro: Genetic modification of tenocytes with exogenous PDGF gene and promotion of collagen gene expression. J. Hand Surg. Am. 2004, 29, 884–890. [Google Scholar] [CrossRef]
- Liu, Y.; Feng, L.; Xu, J.; Yang, Z.; Wu, T.; Zhang, J.; Shi, L.; Zhu, D.; Zhang, J.; Li, G. MiR-378a suppresses tenogenic differentiation and tendon repair by targeting at TGF-beta2. Stem Cell Res. Ther. 2019, 10, 108. [Google Scholar] [CrossRef]
- Delalande, A.; Gosselin, M.P.; Suwalski, A.; Guilmain, W.; Leduc, C.; Berchel, M.; Jaffres, P.A.; Baril, P.; Midoux, P.; Pichon, C. Enhanced Achilles tendon healing by fibromodulin gene transfer. Nanomedicine 2015, 11, 1735–1744. [Google Scholar] [CrossRef]
- Olins, D.E.; Olins, A.L.; von Hippel, P.H. Model nucleoprotein complexes: Studies on the interaction of cationic homopolypeptides with DNA. J. Mol. Biol. 1967, 24, 157–176. [Google Scholar] [CrossRef]
- Laemmli, U.K. Characterization of DNA condensates induced by poly(ethylene oxide) and polylysine. Proc. Natl. Acad. Sci. USA 1975, 72, 4288–4292. [Google Scholar] [CrossRef]
- Alexis, F.; Pridgen, E.; Molnar, L.K.; Farokhzad, O.C. Factors affecting the clearance and biodistribution of polymeric nanoparticles. Mol. Pharm. 2008, 5, 505–515. [Google Scholar] [CrossRef]
- Bazile, D.; Prud’homme, C.; Bassoullet, M.T.; Marlard, M.; Spenlehauer, G.; Veillard, M. Stealth Me.PEG-PLA nanoparticles avoid uptake by the mononuclear phagocytes system. J. Pharm. Sci. 1995, 84, 493–498. [Google Scholar] [CrossRef]
- Lungwitz, U.; Breunig, M.; Blunk, T.; Göpferich, A. Polyethylenimine-based non-viral gene delivery systems. Eur. J. Pharm. Biopharm. 2005, 60, 247–266. [Google Scholar] [CrossRef] [PubMed]
- Godbey, W.T.; Wu, K.K.; Mikos, A.G. Size matters: Molecular weight affects the efficiency of poly(ethylenimine) as a gene delivery vehicle. J. Biomed. Mater. Res. 1999, 45, 268–275. [Google Scholar] [CrossRef]
- Wightman, L.; Kircheis, R.; Rössler, V.; Carotta, S.; Ruzicka, R.; Kursa, M.; Wagner, E. Different behavior of branched and linear polyethylenimine for gene delivery in vitro and in vivo. J. Gene Med. 2001, 3, 362–372. [Google Scholar] [CrossRef] [PubMed]
- Duan, S.; Cao, D.; Li, X.; Zhu, H.; Lan, M.; Tan, Z.; Song, Z.; Zhu, R.; Yin, L.; Chen, Y. Topology-assisted, photo-strengthened DNA/siRNA delivery mediated by branched poly(β-amino ester)s via synchronized intracellular kinetics. Biomater. Sci. 2020, 8, 290–301. [Google Scholar] [CrossRef] [PubMed]
- Lv, H.; Zhang, S.; Wang, B.; Cui, S.; Yan, J. Toxicity of cationic lipids and cationic polymers in gene delivery. J. Control Release 2006, 114, 100–109. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H.K.; Lemieux, P.; Vinogradov, S.V.; Gebhart, C.L.; Guérin, N.; Paradis, G.; Bronich, T.K.; Alakhov, V.Y.; Kabanov, A.V. Evaluation of polyether-polyethyleneimine graft copolymers as gene transfer agents. Gene Ther. 2000, 7, 126–138. [Google Scholar] [CrossRef] [PubMed]
- Petersen, H.; Fechner, P.M.; Martin, A.L.; Kunath, K.; Stolnik, S.; Roberts, C.J.; Fischer, D.; Davies, M.C.; Kissel, T. Polyethylenimine-graft-poly(ethylene glycol) copolymers: Influence of copolymer block structure on DNA complexation and biological activities as gene delivery system. Bioconjug. Chem. 2002, 13, 845–854. [Google Scholar] [CrossRef]
- Muxika, A.; Etxabide, A.; Uranga, J.; Guerrero, P.; de la Caba, K. Chitosan as a bioactive polymer: Processing, properties and applications. Int. J. Biol. Macromol. 2017, 105, 1358–1368. [Google Scholar] [CrossRef]
- Pack, D.W.; Hoffman, A.S.; Pun, S.; Stayton, P.S. Design and development of polymers for gene delivery. Nat. Rev. Drug Discov. 2005, 4, 581–593. [Google Scholar] [CrossRef]
- Gu, Y.; Guo, Y.; Wang, C.; Xu, J.; Wu, J.; Kirk, T.B.; Ma, D.; Xue, W. A polyamidoamne dendrimer functionalized graphene oxide for DOX and MMP-9 shRNA plasmid co-delivery. Mater. Sci. Eng. C Mater. Biol. Appl. 2017, 70, 572–585. [Google Scholar] [CrossRef]
- Xu, W.; Wang, Y.; Li, S.; Ke, Z.; Yan, Y.; Li, S.; Xing, Z.; Wang, C.; Zeng, F.; Liu, R.; et al. Efficient gene and siRNA delivery with cationic polyphosphoramide with amino moieties in the main chain. RSC Adv. 2015, 5, 50425–50432. [Google Scholar] [CrossRef]
- Park, H.J.; Lee, J.; Kim, M.J.; Kang, T.J.; Jeong, Y.; Um, S.H.; Cho, S.W. Sonic hedgehog intradermal gene therapy using a biodegradable poly(beta-amino esters) nanoparticle to enhance wound healing. Biomaterials 2012, 33, 9148–9156. [Google Scholar] [CrossRef] [PubMed]
- Chan, K.M.; Fu, S.C.; Wong, Y.P.; Hui, W.C.; Cheuk, Y.C.; Wong, M.W. Expression of transforming growth factor beta isoforms and their roles in tendon healing. Wound Repair Regen. 2008, 16, 399–407. [Google Scholar] [CrossRef] [PubMed]
- Malcolm, D.W.; Freeberg, M.A.T.; Wang, Y.; Sims, K.R., Jr.; Awad, H.A.; Benoit, D.S.W. Diblock Copolymer Hydrophobicity Facilitates Efficient Gene Silencing and Cytocompatible Nanoparticle-Mediated siRNA Delivery to Musculoskeletal Cell Types. Biomacromolecules 2017, 18, 3753–3765. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhang, L.; Zhao, W.; Wu, Y.; Zhu, C.; Yang, Y. Nanoparticle-mediated delivery of TGF-beta1 miRNA plasmid for preventing flexor tendon adhesion formation. Biomaterials 2013, 34, 8269–8278. [Google Scholar] [CrossRef]
- Yan, Z.; Meng, X.; Su, Y.; Chen, Y.; Zhang, L.; Xiao, J. Double layer composite membrane for preventing tendon adhesion and promoting tendon healing. Mater. Sci. Eng. C Mater. Biol. Appl. 2021, 123, 111941. [Google Scholar] [CrossRef]
- Yang, Z.; Tu, Q.; Zhu, Y.; Luo, R.; Li, X.; Xie, Y.; Maitz, M.F.; Wang, J.; Huang, N. Mussel-inspired coating of polydopamine directs endothelial and smooth muscle cell fate for re-endothelialization of vascular devices. Adv. Healthc. Mater. 2012, 1, 548–559. [Google Scholar] [CrossRef]
- Park, J.; Brust, T.F.; Lee, H.J.; Lee, S.C.; Watts, V.J.; Yeo, Y. Polydopamine-based simple and versatile surface modification of polymeric nano drug carriers. ACS Nano 2014, 8, 3347–3356. [Google Scholar] [CrossRef]
- Zhou, Y.L.; Yang, Q.Q.; Zhang, L.; Tang, J.B. Nanoparticle-coated sutures providing sustained growth factor delivery to improve the healing strength of injured tendons. Acta Biomater. 2021, 124, 301–314. [Google Scholar] [CrossRef]
- Parchi, P.D.; Vittorio, O.; Andreani, L.; Battistini, P.; Piolanti, N.; Marchetti, S.; Poggetti, A.; Lisanti, M. Nanoparticles for Tendon Healing and Regeneration: Literature Review. Front. Aging Neurosci. 2016, 8, 202. [Google Scholar] [CrossRef]
- Fröhlich, E. The role of surface charge in cellular uptake and cytotoxicity of medical nanoparticles. Int. J. Nanomed. 2012, 7, 5577–5591. [Google Scholar] [CrossRef] [PubMed]
- Leroueil, P.R.; Hong, S.; Mecke, A.; Baker, J.R., Jr.; Orr, B.G.; Holl, M.M.B. Nanoparticle interaction with biological membranes: Does nanotechnology present a Janus face? Acc. Chem. Res. 2007, 40, 335–342. [Google Scholar] [CrossRef]
- Leroueil, P.R.; Berry, S.A.; Duthie, K.; Han, G.; Rotello, V.M.; McNerny, D.Q.; Baker, J.R., Jr.; Orr, B.G.; Holl, M.M. Wide varieties of cationic nanoparticles induce defects in supported lipid bilayers. Nano Lett. 2008, 8, 420–424. [Google Scholar] [CrossRef] [PubMed]
- Raffa, V.; Gherardini, L.; Vittorio, O.; Bardi, G.; Ziaei, A.; Pizzorusso, T.; Riggio, C.; Nitodas, S.; Karachalios, T.; Al-Jamal, K.T.; et al. Carbon nanotube-mediated wireless cell permeabilization: Drug and gene uptake. Nanomedicine 2011, 6, 1709–1718. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.H.; Kim, S.H.; Lee, M.; Kim, W.J.; Park, T.G.; Ko, K.S.; Kim, S.W. Non-viral systemic delivery of Fas siRNA suppresses cyclophosphamide-induced diabetes in NOD mice. J. Control Release 2010, 143, 88–94. [Google Scholar] [CrossRef]
- Wang, C.; Du, L.; Zhou, J.; Meng, L.; Cheng, Q.; Wang, C.; Wang, X.; Zhao, D.; Huang, Y.; Zheng, S.; et al. Elaboration on the Distribution of Hydrophobic Segments in the Chains of Amphiphilic Cationic Polymers for Small Interfering RNA Delivery. ACS Appl. Mater. Interfaces 2017, 9, 32463–32474. [Google Scholar] [CrossRef]
- Deng, Z.J.; Morton, S.W.; Ben-Akiva, E.; Dreaden, E.C.; Shopsowitz, K.E.; Hammond, P.T. Layer-by-layer nanoparticles for systemic codelivery of an anticancer drug and siRNA for potential triple-negative breast cancer treatment. ACS Nano 2013, 7, 9571–9584. [Google Scholar] [CrossRef]
- Shaabani, E.; Sharifiaghdam, M.; de Keersmaecker, H.; de Rycke, R.; de Smedt, S.; Faridi-Majidi, R.; Braeckmans, K.; Fraire, J.C. Layer by Layer Assembled Chitosan-Coated Gold Nanoparticles for Enhanced siRNA Delivery and Silencing. Int. J. Mol. Sci. 2021, 22, 831. [Google Scholar] [CrossRef]
- Chono, S.; Li, S.D.; Conwell, C.C.; Huang, L. An efficient and low immunostimulatory nanoparticle formulation for systemic siRNA delivery to the tumor. J. Control Release 2008, 131, 64–69. [Google Scholar] [CrossRef]
- Yang, X.Z.; Dou, S.; Sun, T.M.; Mao, C.Q.; Wang, H.X.; Wang, J. Systemic delivery of siRNA with cationic lipid assisted PEG-PLA nanoparticles for cancer therapy. J. Control Release 2011, 156, 203–211. [Google Scholar] [CrossRef]
- Zhang, Y.; Teng, Z.; Ni, Q.; Tao, J.; Cao, X.; Wen, Y.; Wu, L.; Fang, C.; Wan, B.; Zhang, X.; et al. Orderly Curled Silica Nanosheets with a Small Size and Macromolecular Loading Pores: Synthesis and Delivery of Macromolecules to Eradicate Drug-Resistant Cancer. ACS Appl. Mater. Interfaces 2020, 12, 57810–57820. [Google Scholar] [CrossRef]
- Jiang, X.; Qu, W.; Pan, D.; Ren, Y.; Williford, J.M.; Cui, H.; Luijten, E.; Mao, H.Q. Plasmid-templated shape control of condensed DNA-block copolymer nanoparticles. Adv. Mater. 2013, 25, 227–232. [Google Scholar] [CrossRef] [PubMed]
- Anselmo, A.C.; Zhang, M.; Kumar, S.; Vogus, D.R.; Menegatti, S.; Helgeson, M.E.; Mitragotri, S. Elasticity of nanoparticles influences their blood circulation, phagocytosis, endocytosis, and targeting. ACS Nano 2015, 9, 3169–3177. [Google Scholar] [CrossRef] [PubMed]
- D’Souza, A.A.; Shegokar, R. Polyethylene glycol (PEG): A versatile polymer for pharmaceutical applications. Expert Opin. Drug Deliv. 2016, 13, 1257–1275. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Tian, J.; Chen, X. Effect of surface properties on liposomal siRNA delivery. Biomaterials 2016, 79, 56–68. [Google Scholar] [CrossRef]
- Coué, G.; Engbersen, J.F. Functionalized linear poly(amidoamine)s are efficient vectors for intracellular protein delivery. J. Control Release 2011, 152, 90–98. [Google Scholar] [CrossRef]
- Cohen, S.; Coué, G.; Beno, D.; Korenstein, R.; Engbersen, J.F. Bioreducible poly(amidoamine)s as carriers for intracellular protein delivery to intestinal cells. Biomaterials 2012, 33, 614–623. [Google Scholar] [CrossRef]
- No, Y.J.; Castilho, M.; Ramaswamy, Y.; Zreiqat, H. Role of Biomaterials and Controlled Architecture on Tendon/Ligament Repair and Regeneration. Adv. Mater. 2020, 32, e1904511. [Google Scholar] [CrossRef]
- Shadwick, R.E. Elastic energy storage in tendons: Mechanical differences related to function and age. J. Appl. Physiol. 1990, 68, 1033–1040. [Google Scholar] [CrossRef]
- Race, A.; Amis, A.A. The mechanical properties of the two bundles of the human posterior cruciate ligament. J. Biomech. 1994, 27, 13–24. [Google Scholar] [CrossRef]
- Bosteels, J.; Weyers, S.; Kasius, J.; Broekmans, F.J.; Mol, B.W.; D’Hooghe, T.M. Anti-adhesion therapy following operative hysteroscopy for treatment of female subfertility. Cochrane Database Syst. Rev. 2017, 11, CD011110. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Jha, A.K.; Harrington, D.A.; Farach-Carson, M.C.; Jia, X. Hyaluronic Acid-Based Hydrogels: From a Natural Polysaccharide to Complex Networks. Soft Matter 2012, 8, 3280–3294. [Google Scholar] [CrossRef] [PubMed]
- Burdick, J.A.; Prestwich, G.D. Hyaluronic acid hydrogels for biomedical applications. Adv. Mater. 2011, 23, H41–H56. [Google Scholar] [CrossRef] [PubMed]
- Sorushanova, A.; Delgado, L.M.; Wu, Z.; Shologu, N.; Kshirsagar, A.; Raghunath, R.; Mullen, A.M.; Bayon, Y.; Pandit, A.; Raghunath, M.; et al. The Collagen Suprafamily: From Biosynthesis to Advanced Biomaterial Development. Adv. Mater. 2019, 31, e1801651. [Google Scholar] [CrossRef] [PubMed]
- Marinho-Soriano, E. Agar polysaccharides from Gracilaria species (Rhodophyta, Gracilariaceae). J. Biotechnol. 2001, 89, 81–84. [Google Scholar] [CrossRef]
- Bhattarai, N.; Gunn, J.; Zhang, M. Chitosan-based hydrogels for controlled, localized drug delivery. Adv. Drug Deliv. Rev. 2010, 62, 83–99. [Google Scholar] [CrossRef]
- Sabra, W.; Zeng, A.P.; Deckwer, W.D. Bacterial alginate: Physiology, product quality and process aspects. Appl. Microbiol. Biotechnol. 2001, 56, 315–325. [Google Scholar] [CrossRef]
- Ghosh, S.K.; Park, J.; Na, S.; Kim, M.P.; Ko, H. A Fully Biodegradable Ferroelectric Skin Sensor from Edible Porcine Skin Gelatine. Adv. Sci. 2021, 8, 2005010. [Google Scholar] [CrossRef]
- Klemm, D.; Heublein, B.; Fink, H.P.; Bohn, A. Cellulose: Fascinating biopolymer and sustainable raw material. Angew Chem. Int. Ed. Engl. 2005, 44, 3358–3393. [Google Scholar] [CrossRef]
- Habibi, Y.; Lucia, L.A.; Rojas, O.J. Cellulose nanocrystals: Chemistry, self-assembly, and applications. Chem. Rev. 2010, 110, 3479–3500. [Google Scholar] [CrossRef]
- Moon, R.J.; Martini, A.; Nairn, J.; Simonsen, J.; Youngblood, J. Cellulose nanomaterials review: Structure, properties and nanocomposites. Chem. Soc. Rev. 2011, 40, 3941–3994. [Google Scholar] [CrossRef] [PubMed]
- Schulze, J.; Hendrikx, S.; Schulz-Siegmund, M.; Aigner, A. Microparticulate poly(vinyl alcohol) hydrogel formulations for embedding and controlled release of polyethylenimine (PEI)-based nanoparticles. Acta Biomater. 2016, 45, 210–222. [Google Scholar] [CrossRef] [PubMed]
- Sefton, M.V.; May, M.H.; Lahooti, S.; Babensee, J.E. Making microencapsulation work: Conformal coating, immobilization gels and in vivo performance. J. Control Release 2000, 65, 173–186. [Google Scholar] [CrossRef]
- Tanaka, F.; Koga, T.; Winnik, F.M. Temperature-responsive polymers in mixed solvents: Competitive hydrogen bonds cause cononsolvency. Phys. Rev. Lett. 2008, 101, 028302. [Google Scholar] [CrossRef] [PubMed]
- Tighe, B.J. A decade of silicone hydrogel development: Surface properties, mechanical properties, and ocular compatibility. Eye Contact Lens 2013, 39, 4–12. [Google Scholar] [CrossRef]
- Nagase, K. Thermoresponsive interfaces obtained using poly(N-isopropylacrylamide)-based copolymer for bioseparation and tissue engineering applications. Adv. Colloid Interface Sci. 2021, 295, 102487. [Google Scholar] [CrossRef]
- Grosser, T.; Ricciotti, E.; FitzGerald, G.A. The Cardiovascular Pharmacology of Nonsteroidal Anti-Inflammatory Drugs. Trends Pharmacol. Sci. 2017, 38, 733–748. [Google Scholar] [CrossRef]
- Bindu, S.; Mazumder, S.; Bandyopadhyay, U. Non-steroidal anti-inflammatory drugs (NSAIDs) and organ damage: A current perspective. Biochem. Pharmacol. 2020, 180, 114147. [Google Scholar] [CrossRef]
- Zhou, Y.L.; Yang, Q.Q.; Yan, Y.Y.; Zhu, C.; Zhang, L.; Tang, J.B. Localized delivery of miRNAs targets cyclooxygenases and reduces flexor tendon adhesions. Acta Biomater. 2018, 70, 237–248. [Google Scholar] [CrossRef]
- Sgambato, A.; Cipolla, L.; Russo, L. Bioresponsive Hydrogels: Chemical Strategies and Perspectives in Tissue Engineering. Gels 2016, 2, 28. [Google Scholar] [CrossRef]
- Liu, X.; Steiger, C.; Lin, S.; Parada, G.A.; Liu, J.; Chan, H.F.; Yuk, H.; Phan, N.V.; Collins, J.; Tamang, S.; et al. Ingestible hydrogel device. Nat. Commun. 2019, 10, 493. [Google Scholar] [CrossRef] [PubMed]
- Mo, F.; Jiang, K.; Zhao, D.; Wang, Y.; Song, J.; Tan, W. DNA hydrogel-based gene editing and drug delivery systems. Adv. Drug Deliv. Rev. 2021, 168, 79–98. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Sun, B.; Zhao, C.; Wang, Z.; Teng, S.; Yang, M.; Cui, Z.; Zhu, G.; Yu, Y. Three-Dimensional Tendon Scaffold Loaded with TGF-beta1 Gene Silencing Plasmid Prevents Tendon Adhesion and Promotes Tendon Repair. ACS Biomater. Sci. Eng. 2021, 7, 5739–5748. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reference | Year | Target Gene | Function on Tendon | Type of Study | Title |
|---|---|---|---|---|---|
| [57] | 2006 | Runx2 Cbfa1 | The silencing of Runx2/Cbfa1 inhibits the formation of heterotopic ossification. | in vitro | Adenovirus-mediated transfer of siRNA against Runx2/Cbfa1 inhibits the formation of heterotopic ossification in animal model |
| [58] | 2007 | cadherin-11 | Cell–cell junctions and alignment of collagen fibrils are mediated by cadherin-11, which promotes tendon formation. | in vitro | Tendon development requires regulation of cell condensation and cell shape via cadherin-11-mediated cell-cell junctions |
| [59] | 2008 | COMP | COMP protects chondrocytes against apoptosis via elevating the proteins of the IAP family. | in vitro | Cartilage oligomeric matrix protein protects cells against death by elevating members of the IAP family of survival proteins |
| [60] | 2008 | IL-1beta | The silencing of IL-1beta regulates MMP-13, which is also affected by fatigue loading. | in vitro | Coordinate regulation of IL-1beta and MMP-13 in rat tendons following sub-rupture fatigue damage |
| [61] | 2009 | NCX | NCX involves in the role of fibroblasts during tendon healing | in vitro | Involvement of Na+/Ca2+ exchanger in migration and contraction of rat cultured tendon fibroblasts |
| [62] | 2009 | APC | APC promotes the proliferation of tenocytes and the synthesis of collagen1. | in vitro | Activated protein C mediates a healing phenotype in cultured tenocytes |
| [63] | 2010 | Rnux2 Smad4 | The inhibition of Runx2 and Smad4 could prevent heterotopic ossification. | in vitro | Non-virus-mediated transfer of siRNAs against Runx2 and Smad4 inhibit heterotopic ossification in rats |
| [64] | 2011 | collagen V | Collagen V α1 plays an important role in tendon regeneration. | in vitro | Col V siRNA engineered tenocytes for tendon tissue engineering |
| [65] | 2012 | Wnt5a | The Wnt5a-RhoA pathway plays an important role in uniaxial mechanical tendon-induced osteogenic differentiation. | in vitro | Uniaxial mechanical tension promoted osteogenic differentiation of rat tendon-derived stem cells (rTDSCs) via the Wnt5a-RhoA pathway |
| [66] | 2013 | AMPKα1 | HGF inhibits GF-β1-induced myofibroblastic differentiation via AMPK. | in vitro | Hepatocyte growth factor inhibits TGF-β1-induced myofibroblast differentiation in tendon fibroblasts: role of AMPK signaling pathway |
| [67] | 2013 | Mohawk | The inhibition of MKX would downregulate COL1A1 and TNXB and upregulate SOX9. | in vitro | Transcription factor Mohawk and the pathogenesis of human anterior cruciate ligament degradation |
| [68] | 2013 | ERK2 | Tendon adhesion will be regulated by the inhibition of ERK2. | in vivo | Prevention of Tendon Adhesions by ERK2 Small Interfering RNAs |
| [69] | 2015 | TGIF1 | TGIF1 could prevent tendon-to-bone from chondrogenic differentiation. | in vitro | TGIF1 Gene Silencing in Tendon-Derived Stem Cells Improves the Tendon-to-Bone Insertion Site Regeneration |
| [70] | 2015 | Pin1 | Pin1 plays an important role in the progression of TSPCs aging. | in vitro | The role of Pin1 protein in aging of human tendon stem/progenitor cells |
| [71] | 2015 | scleraxis | Scleraxis is vital to the differentiation of TSCs to tenocytes. | in vitro | Dexamethasone inhibits the differentiation of rat tendon stem cells into tenocytes by targeting the scleraxis gene |
| [72] | 2015 | TGIF1 | Rats perform better functions after being treated with TGIF1-siRNA BMSCs. | in vitro | Silencing of TGIF1 in bone mesenchymal stem cells applied to the post-operative rotator cuff improves both functional and histologic outcomes |
| [73] | 2015 | TNF-α | NF-κB, MMP1, MMP9, COX-1, and COX-2 which involve in inflammation may be downregulated. | in vivo | Targeted knockout of TNF-α by injection of lentivirus-mediated siRNA into the subacromial bursa for the treatment of subacromial bursitis in rats |
| [74] | 2017 | RelA/p65 | p65 plays a core role in fibrosis by inhibiting cell proliferation and the expression of ECM. | in vitro | RelA/p65 inhibition prevents tendon adhesion by modulating inflammation, cell proliferation, and apoptosis |
| [75] | 2017 | ANGPTL4 | ANGPTL4 serves as a multifunctional protein to regulate cell migration and proliferation. | in vitro | Angiopoietin-like 4 Enhances the Proliferation and Migration of Tendon Fibroblasts |
| [76] | 2018 | serpine1 | The inhibition of serpine1 promotes the activity of MMP, which could protect tendons against adhesion. | in vitro | Serpine1 Knockdown Enhances MMP Activity after Flexor Tendon Injury in Mice: Implications for Adhesions Therapy |
| [77] | 2018 | scleraxis | Scx regulates several mechanosensitive proteins involved in adhesion. | in vitro | Novel roles for scleraxis in regulating adult tenocyte function |
| [78] | 2018 | scleraxis | Scx enhances the level of tenomodulin. | in vitro | Scleraxis is a transcriptional activator that regulates the expression of Tenomodulin, a marker of mature tenocytes and ligamentocytes |
| [79] | 2018 | FOXP1 | FOXP1 promotes self-renewal of TSPCs by decreasing E2F1, pRb and cylin D1. | in vitro | Downregulation of FOXP1 correlates with tendon stem/progenitor cells aging |
| [80] | 2020 | Flightless I | Flii could reduce the proliferation and migration of human tenocyte. | in vitro | Increasing the level of cytoskeletal protein Flightless I reduces adhesion formation in a murine digital flexor tendon model |
| [81] | 2020 | Collagen III | polyDMAEA-siRNA polyexes show more promising efficiency compared to PEI-siRNA. | in vitro | Synthesis and Formulation of Four-Arm PolyDMAEA-siRNA Polyplex for Transient Downregulation of Collagen Type III Gene Expression in TGF-β1 Stimulated Tenocyte Culture |
| [82] | 2021 | CLK2 DYRK1A | SM04755 reduces inflammation and enhances tenocytes differentiation by inhibiting CLK2 and DYRK1A | in vitro | SM04755, a small-molecule inhibitor of the Wnt pathway, as a potential topical treatment for tendinopathy |
| [83] | 2021 | Smad3 | the inhibition of transforming. Growth factor-β (TGF-β1)/Smad2/3 signal pathway could enhance tendon healing. | in vivo | Inhibition of Smad3 promotes the healing of rotator cuff injury in a rat model |
| [84] | 2021 | ITGA9 | Tenascin-C promotes the regeneration of tendons via ITGA9-mediated migration of STSCs. | in vitro | Tenascin-C regulates migration of SOX10 tendon stem cells via integrin-α9 for promoting patellar tendon remodeling |
| [85] | 2022 | Smad3 | The delivery system serves as an effective antiadhesion barrier, which could also decrease inflammation. | in vivo | Self-Healing Hydrogel Embodied with Macrophage-Regulation and Responsive-Gene-Silencing Properties for Synergistic Prevention of Peritendinous Adhesion |
| [86] | 2022 | IKKβ | Blocking KKβ/NF-κB pathway in vivo could treat RCT well. | in vivo | Inhibition of IKKβ/NF-κB signaling facilitates tendinopathy healing by rejuvenating inflamm-aging induced tendon-derived stem/progenitor cell senescence |
| [87] | 2022 | COX | The inhibition of COX could transform M1 to M2. | in vivo | Morphological changes of macrophages and their potential contribution to tendon healing |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jin, J.; Yang, Q.Q.; Zhou, Y.L. Non-Viral Delivery of Gene Therapy to the Tendon. Polymers 2022, 14, 3338. https://doi.org/10.3390/polym14163338
Jin J, Yang QQ, Zhou YL. Non-Viral Delivery of Gene Therapy to the Tendon. Polymers. 2022; 14(16):3338. https://doi.org/10.3390/polym14163338
Chicago/Turabian StyleJin, Jing, Qian Qian Yang, and You Lang Zhou. 2022. "Non-Viral Delivery of Gene Therapy to the Tendon" Polymers 14, no. 16: 3338. https://doi.org/10.3390/polym14163338
APA StyleJin, J., Yang, Q. Q., & Zhou, Y. L. (2022). Non-Viral Delivery of Gene Therapy to the Tendon. Polymers, 14(16), 3338. https://doi.org/10.3390/polym14163338