
Static Culture Combined with Aeration in Biosynthesis of Bacterial Cellulose

 ,
, 


 ,
, 
 ,
,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Microbial Producer
2.2. Biosynthesis of Bacterial Cellulose
2.3. BC Yield
2.4. Analytical Techniques
2.4.1. Active Acidity
2.4.2. Concentration of Reducing Sugars
2.4.3. Scanning Electron Microscopy of BC
2.4.4. BC Degree of Polymerization
2.4.5. Thermomechanical Analysis
2.4.6. Thermogravimetric Analysis
2.4.7. X-ray Diffraction of BC
3. Results and Discussion
3.1. Variation of Culture Parameters during BC Biosynthesis
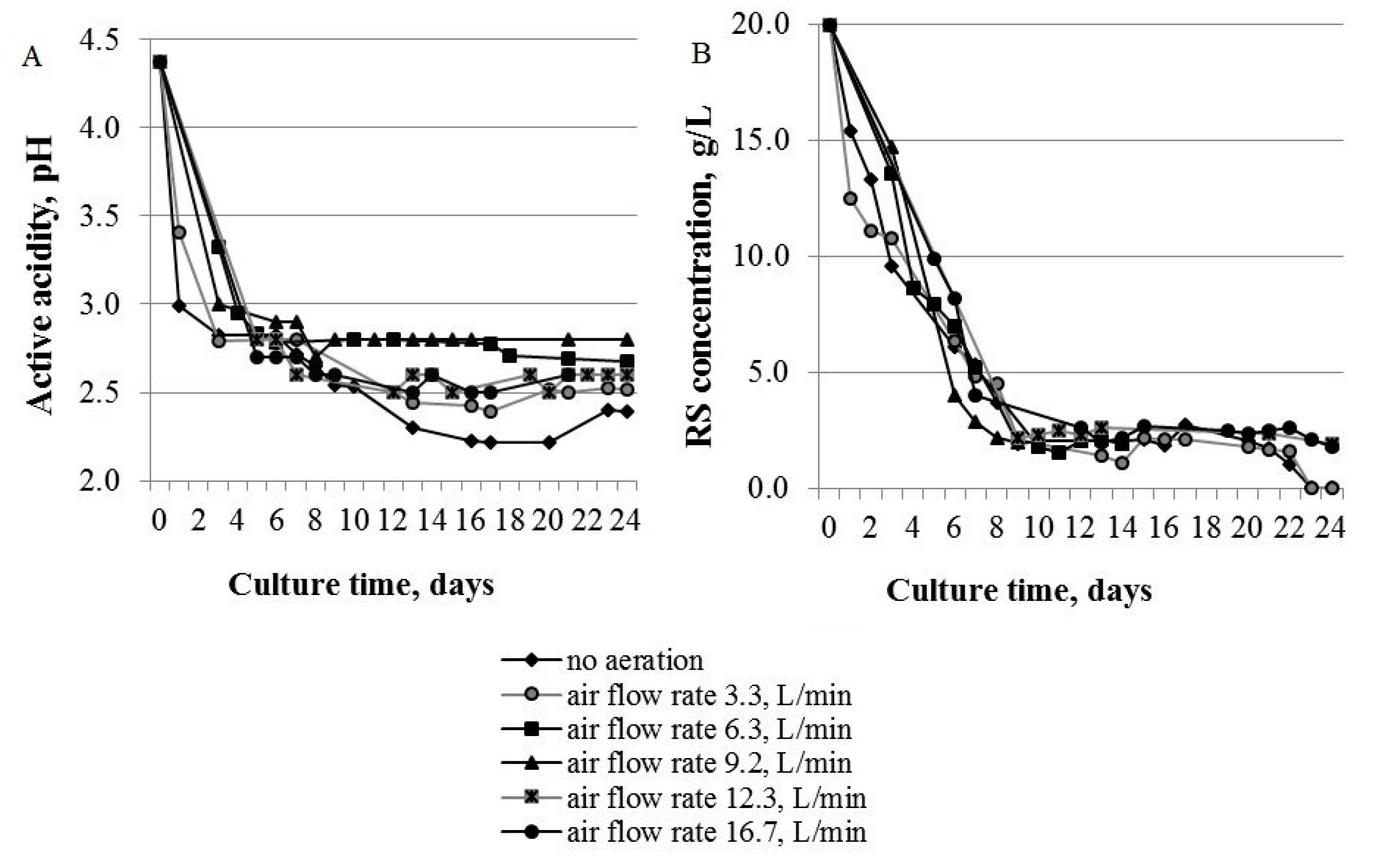
3.1.1. Active Acidity and RS Concentration
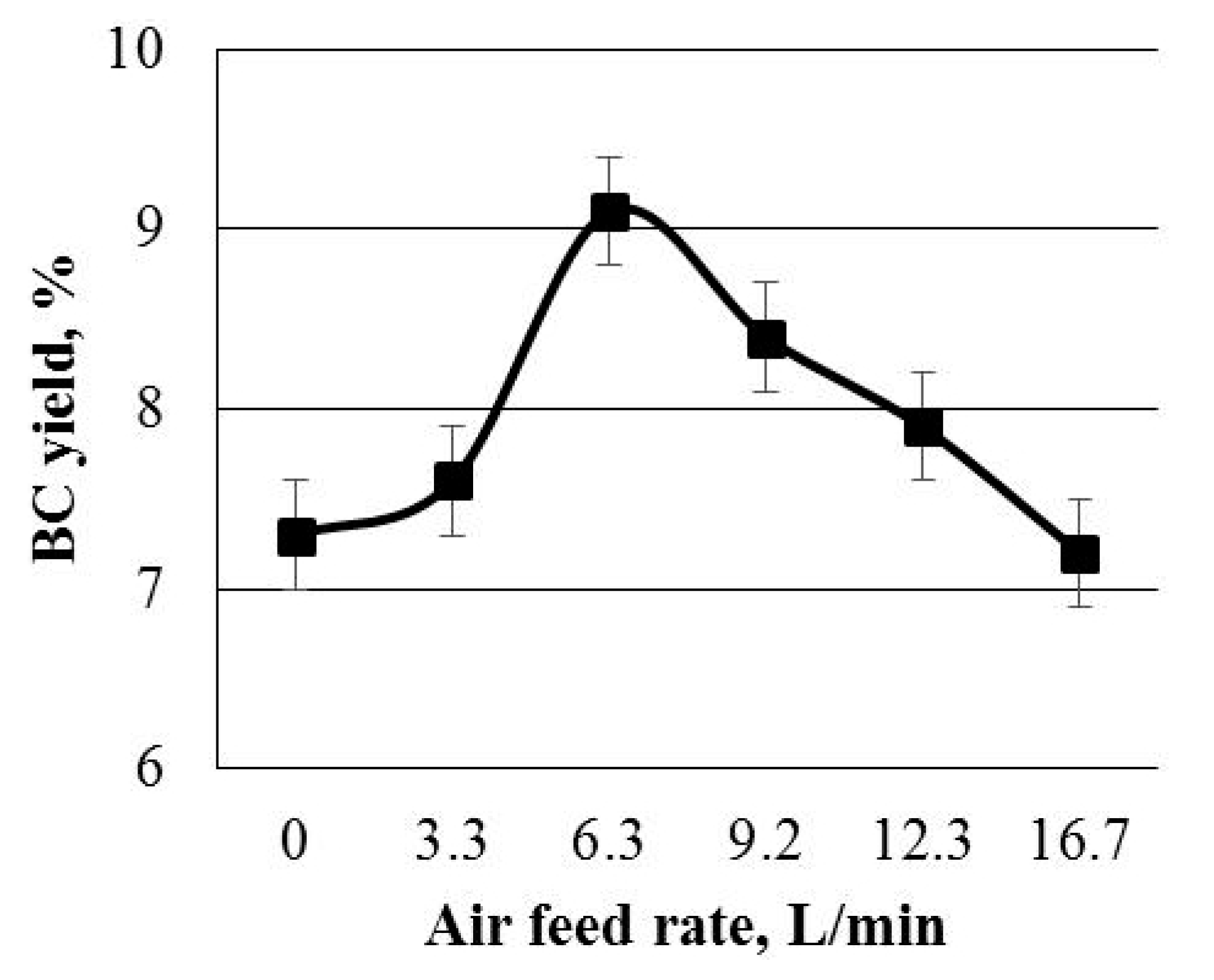
3.1.2. BC Yield
3.2. Characterization of BC Samples
3.2.1. Appearance of BC Gel-Films
3.2.2. Microstructure of BC
3.2.3. BC Degree of Polymerization
3.2.4. BC Elastic Modulus
3.2.5. Thermogravimetric Analysis (TGA)
3.2.6. X-ray Diffraction
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, J.; Tavakoli, J.; Tang, Y. Bacterial cellulose production, properties and applications with different culture methods—A review. Carbohydr. Polym. 2019, 219, 63–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klemm, D.; Petzold-Welcke, K.; Kramer, F.; Richter, T.; Raddatz, V.; Fried, W.; Nietzsche, S.; Bellmann, T.; Fischer, D. Biotech nanocellulose: A review on progress in product design and today’s state of technical and medical applications. Carbohydr. Polym. 2020, 254, 117313. [Google Scholar] [CrossRef]
- Bodea, I.M.; Beteg, F.I.; Pop, C.R.; David, A.P.; Dudescu, M.C.; Vilău, C.; Stănilă, A.; Rotar, A.M.; Catunescu, G.M. Optimi-zation of moist and oven-dried bacterial cellulose production for functional properties. Polymers 2021, 13, 2088. [Google Scholar] [CrossRef]
- Revin, V.; Dolganov, A.; Liyaskina, E.; Nazarova, N.; Balandina, A.; Devyataeva, A.; Revin, V. Characterizing Bacterial Cellulose Produced byKomagataeibacter sucrofermentans H-110 on Molasses Medium and Obtaining a Biocomposite Based on It for the Adsorption of Fluoride. Polymers 2021, 13, 1422. [Google Scholar] [CrossRef] [PubMed]
- Abol-Fotouh, D.; Hassan, M.A.; Shokry, H.; Roig, A.; Azab, M.S.; Kashyout, A.E.-H.B. Bacterial nanocellulose from agro-industrial wastes: Low-cost and enhanced production by Komagataeibacter saccharivorans MD1. Sci. Rep. 2020, 10, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Sharma, C.; Bhardwaj, N.K. Bacterial nanocellulose: Present status, biomedical applications and future perspectives. Mater. Sci. Eng. C 2019, 104, 109963. [Google Scholar] [CrossRef] [PubMed]
- Podgorbunskikh, E.M.; Bychkov, A.L.; Lomovsky, O.I. Determination of Surface Accessibility of the Cellulose Substrate According to Enzyme Sorption. Polymers 2019, 11, 1201. [Google Scholar] [CrossRef] [Green Version]
- Daskalova, A.; Angelova, L.; Filipov, E.; Aceti, D.; Mincheva, R.; Carrete, X.; Kerdjoudj, H.; Dubus, M.; Chevrier, J.; Trifonov, A.; et al. Biomimetic Hierarchical Structuring of PLA by Ultra-Short Laser Pulses for Processing of Tissue Engineered Matrices: Study of Cellular and Antibacterial Behavior. Polymers 2021, 13, 2577. [Google Scholar] [CrossRef] [PubMed]
- Gregory, D.A.; Tripathi, L.; Fricker, A.T.R.; Asare, E.; Orlando, I.; Raghavendran, V.; Roy, I. Bacterial cellulose: A smart biomaterial with diverse applications. Mater. Sci. Eng. R Rep. 2021, 145, 100623. [Google Scholar] [CrossRef]
- Mbituyimana, B.; Liu, L.; Ye, W.; Boni, B.O.O.; Zhang, K.; Chen, J.; Thomas, S.; Vasilievich, R.V.; Shi, Z.; Yang, G. Bacterial cellulose-based composites for biomedical and cosmetic applications: Research progress and existing products. Carbohydr. Polym. 2021, 273, 118565. [Google Scholar] [CrossRef]
- Shirai, A.; Takahashi, M.; Kaneko, H.; Nishimura, S.-I.; Ogawa, M.; Nishi, N.; Tokura, S. Biosynthesis of a novel polysaccharide by Acetobacter xylinum. Int. J. Biol. Macromol. 1994, 16, 297–300. [Google Scholar] [CrossRef]
- Aloni, Y.; Delmer, D.P.; Benziman, M. Achievement of high rates of in vitro synthesis of 1,4-beta-D-glucan: Activation by cooperative interaction of the Acetobacter xylinum enzyme system with GTP, polyethylene glycol, and a protein factor. Proc. Natl. Acad. Sci. USA 1982, 79, 6448–6452. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Catchmark, J.; Vogler, E. Factors impacting the formation of sphere-like bacterial cellulose particles and their bio-compatibility for human osteoblast growth. Biomacromolecules 2013, 14, 3444–3452. [Google Scholar] [CrossRef] [PubMed]
- Cai, Q.; Hu, C.; Yang, N.; Wang, Q.; Wang, J.; Pan, H.; Hu, Y.; Ruan, C. Enhanced activity and stability of industrial lipases immobilized onto spherelike bacterial cellulose. Int. J. Biol. Macromol. 2017, 109, 1174–1181. [Google Scholar] [CrossRef]
- Czaja, W.; Romanovicz, D.; Brown, R.M. Structural investigations of microbial cellulose produced in stationary and agitated culture. Cellulose 2004, 11, 403–411. [Google Scholar] [CrossRef]
- Tantratian, S.; Tammarate, P.; Krusong, W.; Bhattarakosol, P.; Phunsri, A. Effect of dissolved oxygen on cellulose production by Acetobacter sp. J. Sci. Res. Chula. Univ. 2005, 30, 179–186. [Google Scholar]
- Krystynowicz, A.; Czaja, W.; Wiktorowska-Jezierska, A.; Goncalves-Miskiewicz, M.; Turkiewicz, M.; Bielecki, S. Factors af-fecting the yield and properties of bacterial cellulose. J. Ind. Microbiol. Biotechnol. 2002, 29, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Zhu, C.; Yang, J.; Nie, Y.; Chen, C.; Sun, D. Recent advances in bacterial cellulose. Cellulose 2013, 21, 1–30. [Google Scholar] [CrossRef]
- Kouda, T.; Yano, H.; Yoshinaga, F.; Kaminoyama, M.; Kamiwano, M. Characterization of non-newtonian behavior during mixing of bacterial cellulose in a bioreactor. J. Ferment. Bioeng. 1996, 82, 382–386. [Google Scholar] [CrossRef]
- Kouda, T.; Naritomi, T.; Yano, H.; Yoshinaga, F. Effects of oxygen and carbon dioxide pressures on bacterial cellulose pro-duction by Acetobacter in aerated and agitated culture. J. Ferment. Bioeng. 1997, 84, 124–127. [Google Scholar] [CrossRef]
- Chao, Y.; Sugano, Y.; Shoda, M. Bacterial cellulose production under oxygen-enriched air at different fructose concentrations in a 50-liter, internal-loop airlift reactor. Appl. Microbiol. Biotechnol. 2001, 55, 673–679. [Google Scholar] [CrossRef]
- Wu, S.-C.; Li, M.-H. Production of bacterial cellulose membranes in a modified airlift bioreactor by Gluconacetobacter xylinus. J. Biosci. Bioeng. 2015, 120, 444–449. [Google Scholar] [CrossRef] [PubMed]
- Song, H.-J.; Li, H.; Seo, J.-H.; Kim, M.-J.; Kim, S.-J. Pilot-scale production of bacterial cellulose by a spherical type bubble column bioreactor using saccharified food wastes. Korean J. Chem. Eng. 2009, 26, 141–146. [Google Scholar] [CrossRef]
- Zahan, K.A.; Pa’e, N.; Muhamad, I.I. An evaluation of fermentation period and discs rotation speed of rotary discs reactor for bacterial cellulose production. Sains Malays. 2016, 45, 393–400. [Google Scholar]
- Zhong, C. Industrial-Scale Production and Applications of Bacterial Cellulose. Front. Bioeng. Biotechnol. 2020, 8. [Google Scholar] [CrossRef]
- Chakravorty, S.; Bhattacharya, S.; Chatzinotas, A.; Chakraborty, W.; Bhattacharya, D.; Gachhui, R. Kombucha tea fermen-tation: Microbial and biochemical dynamics. Int. J. Food Microbiol. 2016, 220, 63–72. [Google Scholar] [CrossRef]
- Marsh, A.J.; O'Sullivan, O.; Hill, C.; Ross, R.; Cotter, P.D. Sequence-based analysis of the bacterial and fungal compositions of multiple kombucha (tea fungus) samples. Food Microbiol. 2013, 38, 171–178. [Google Scholar] [CrossRef]
- Sharma, C.; Bhardwaj, N.K. Biotransformation of fermented black tea into bacterial nanocellulose via symbiotic interplay of microorganisms. Int. J. Biol. Macromol. 2019, 132, 166–177. [Google Scholar] [CrossRef]
- Gladysheva, E.K.; Skiba, E.A.; Zolotukhin, V.; Sakovich, G.V. Study of the Conditions for the Biosynthesis of Bacterial Cellulose by the Producer Medusomyces gisevii Sa-12. Appl. Biochem. Microbiol. 2018, 54, 179–187. [Google Scholar] [CrossRef]
- Shavyrkina, N.; Budaeva, V.; Skiba, E.; Mironova, G.; Bychin, N.; Gismatulina, Y.; Kashcheyeva, E.; Sitnikova, A.; Shilov, A.; Kuznetsov, P.; et al. Scale-Up of Biosynthesis Process of Bacterial Nanocellulose. Polymers 2021, 13, 1920. [Google Scholar] [CrossRef]
- Hallac, B.B.; Ragauskas, A.J. Analyzing cellulose degree of polymerization and its relevancy to cellulosic ethanol. Biofuels Bioprod. Biorefin. 2011, 5, 215–225. [Google Scholar] [CrossRef]
- French, A.D. Idealized powder diffraction patterns for cellulose polymorphs. Cellulose 2013, 21, 885–896. [Google Scholar] [CrossRef]
- French, A.D. Increment in evolution of cellulose crystallinity analysis. Cellulose 2020, 27, 5445–5448. [Google Scholar] [CrossRef]
- Yurkevich, D.I.; Kutyshenko, V.P. Medusomyces (Tea fungus): A scientific history, composition, features of physiology and metabolism. Biophysics 2002, 47, 1035–1048. [Google Scholar]
- Krusong, W.; Pothimon, R.; La China, S.; Thompson, A.K. Consecutive bacterial cellulose production by luffa sponge enmeshed with cellulose microfibrils of Acetobacter xylinum under continuous aeration. 3 Biotech 2021, 11, 1–9. [Google Scholar] [CrossRef]
- Gama, M.; Dourado, F.; Bielecki, S. Bacterial nanocellulose. From Biotechnology to Bio-Economy, 1st ed.; Elsevier: Amsterdam, Netherlands, 2016. [Google Scholar]
- Singhsa, P.; Narain, R.; Manuspiya, H. Physical structure variations of bacterial cellulose produced by different Komaga-taeibacter xylinus strains and carbon sources in static and agitated conditions. Cellulose 2018, 25, 1571–1581. [Google Scholar] [CrossRef]
- Pacheco, G.; Nogueira, C.; Meneguin, A.; Trovatti, E.; Silva, M.C.; Machado, R.T.; Ribeiro, S.; Filho, E.S.; Barud, H.D.S. Development and characterization of bacterial cellulose produced by cashew tree residues as alternative carbon source. Ind. Crop. Prod. 2017, 107, 13–19. [Google Scholar] [CrossRef] [Green Version]
- Um, I.C.; Ki, C.S.; Kweon, H.Y.; Lee, K.G.; Ihm, D.W.; Park, Y.H. Wet spinning of silk polymer II. Effect of drawing on the structure characteristics and properties of filament. Int. J. Biol. Macromol. 2004, 34, 107–119. [Google Scholar] [CrossRef]
- Cheng, K.-C.; Catchmark, J.M.; Demirci, A. Enhanced Production of Bacterial Cellulose Production by Using Biofilm Reactor and its Material Property Analysis. J. Biol. Eng. 2009. [Google Scholar] [CrossRef] [Green Version]
- Skiba, E.; Gladysheva, E.K.; Golubev, D.S.; Budaeva, V.V.; Aleshina, L.; Sakovich, G.V. Self-standardization of quality of bacterial cellulose produced by Medusomyces gisevii in nutrient media derived from Miscanthus biomass. Carbohydr. Polym. 2020, 252, 117178. [Google Scholar] [CrossRef]
- Orlovska, I.; Podolich, O.; Kukharenko, O.; Zaets, I.; Reva, O.; Khirunenko, L.; Zmejkoski, D.; Rogalsky, S.; Barh, D.; Tiwari, S.; et al. Bacterial Cellulose Retains Ro-bustness but Its Synthesis Declines After Exposure to a Mars-Like Environment Simulated Outside the International Space Station. Astrobiology 2021, 21, 706–717. [Google Scholar] [CrossRef] [PubMed]
- Forte, A.; Dourado, F.; Mota, A.; Neto, B.; Gama, M.; Ferreira, E.C. Life cycle assessment of bacterial cellulose production. Int. J. Life Cycle Assess. 2021, 26, 864–878. [Google Scholar] [CrossRef]
- Pillai, M.M.; Tran, H.N.; Sathishkumar, G.; Manimekalai, K.; Yoon, J.; Lim, D.; Noh, I.; Bhattacharyya, A. Symbiotic culture of nanocellulose pellicle: A potential matrix for 3D bioprinting. Mater. Sci. Eng. C 2020, 119, 111552. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H.T.; Saha, N.; Ngwabebhoh, F.A.; Zandraa, O.; Saha, T.; Saha, P. Kombucha-derived bacterial cellulose from diverse wastes: A prudent leather alternative. Cellulose 2021, 28, 9335–9353. [Google Scholar] [CrossRef]
- Laavanya, D.; Shirkole, S.; Balasubramanian, P. Current challenges, applications and future perspectives of SCOBY cellulose of Kombucha fermentation. J. Clean. Prod. 2021, 295, 126454. [Google Scholar] [CrossRef]
- Gismatulina, Y.A.; Budaeva, V.V.; Sitnikova, A.E.; Bychin, N.V.; Gladysheva, E.K.; Shavyrkina, N.A.; Mironova, G.F.; Sevastyanova, Y.V. Bacterial nanocellulose and softwood pulp for composite paper. Proc. Univ. Appl. Chem. Biotechnol. 2021, 11, 460–471. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Aeration Rate, L/min | Proportion of Uniform Gel-Films, % | BC Yield, % |
|---|---|---|
| No aeration (control) | 100 | 7.3 ± 0.3 |
| 3.3 | 89.9 ± 1.5 | 7.6 ± 0.3 |
| 6.3 | 72.4 ± 2.2 | 9.1 ± 0.3 |
| 9.2 | 64.9 ± 1.9 | 8.4 ± 0.3 |
| 12.3 | 60.7 ± 2.5 | 7.9 ± 0.3 |
| 16.7 | 53.2 ± 2.6 | 7.2 ± 0.3 |
| Parameter | Aeration Rate, L/min | |||||
|---|---|---|---|---|---|---|
| No Aeration | 3.3 | 6.3 | 9.2 | 12.3 | 16.7 | |
| Average microfibril width, ±40 nm | 107 | 93 | 81 | 101 | 85 | 97 |
| Degree of polymerization, ±100 | 4300 | 4200 | 3800 | 2550 | 2100 | 950 |
| Young’s modulus, ±10 MPa | 910 | 860 | 795 | 490 | 360 | 315 |
| TGA summary data | ||||||
| Sample weight loss at the first stage, ±0.5% | 4.1 | 5.0 | 3.9 | 4.5 | 4.1 | 2.8 |
| Sample weight loss at the second stage (within sample decomposition range), ±0.1% | 7.8 | 17.9 | 12.4 | 19.5 | 37.4 | 19.1 |
| Onset temperature of decomposition, ±5 °C | 353 | 372 | 374 | 330 | 308 | 297 |
| Sample weight loss at the third stage, ±1.0% | 62.5 | 49.2 | 63.6 | 52.4 | 30.4 | 45.7 |
| Sample weight loss at the fourth stage, ±0.5% | 8.4 | 9.1 | 5.9 | 6.0 | 9.0 | 18.3 |
| Unburnable residue, ±1.0% | 17.2 | 18.9 | 14.3 | 17.6 | 19.1 | 14.2 |
| Concentrations of cellulose allomorphs and crystallinity index as measured by X-ray diffraction | ||||||
| Iα-allomorph, ±5% | 98 | 94 | 94 | 96 | 94 | 97 |
| Iβ-allomorph, ±5% | 2 | 6 | 6 | 4 | 6 | 3 |
| Index of crystallinity (reflection geometry), ±5% | 93 | 93 | 89 | 87-90 | 93 | 91 |
| Index of crystallinity (transmission geometry), ±5% | 89 | 95 | 92 | 92 | 93 | 92 |
| Crystallite size of <110>, ±0.5 nm | 7.7 | 7.8 | 7.8 | 7.8 | 7.8 | 7.7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shavyrkina, N.A.; Skiba, E.A.; Kazantseva, A.E.; Gladysheva, E.K.; Budaeva, V.V.; Bychin, N.V.; Gismatulina, Y.A.; Kashcheyeva, E.I.; Mironova, G.F.; Korchagina, A.A.; et al. Static Culture Combined with Aeration in Biosynthesis of Bacterial Cellulose. Polymers 2021, 13, 4241. https://doi.org/10.3390/polym13234241
Shavyrkina NA, Skiba EA, Kazantseva AE, Gladysheva EK, Budaeva VV, Bychin NV, Gismatulina YA, Kashcheyeva EI, Mironova GF, Korchagina AA, et al. Static Culture Combined with Aeration in Biosynthesis of Bacterial Cellulose. Polymers. 2021; 13(23):4241. https://doi.org/10.3390/polym13234241
Chicago/Turabian StyleShavyrkina, Nadezhda A., Ekaterina A. Skiba, Anastasia E. Kazantseva, Evgenia K. Gladysheva, Vera V. Budaeva, Nikolay V. Bychin, Yulia A. Gismatulina, Ekaterina I. Kashcheyeva, Galina F. Mironova, Anna A. Korchagina, and et al. 2021. "Static Culture Combined with Aeration in Biosynthesis of Bacterial Cellulose" Polymers 13, no. 23: 4241. https://doi.org/10.3390/polym13234241
APA StyleShavyrkina, N. A., Skiba, E. A., Kazantseva, A. E., Gladysheva, E. K., Budaeva, V. V., Bychin, N. V., Gismatulina, Y. A., Kashcheyeva, E. I., Mironova, G. F., Korchagina, A. A., Pavlov, I. N., & Sakovich, G. V. (2021). Static Culture Combined with Aeration in Biosynthesis of Bacterial Cellulose. Polymers, 13(23), 4241. https://doi.org/10.3390/polym13234241