Abstract
Fossil fuels are energy recourses that fulfill most of the world’s energy requirements. However, their production and use cause severe health and environmental problems including global warming and pollution. Consequently, plant and animal-based fuels (also termed as biofuels), such as biogas, biodiesel, and many others, have been introduced as alternatives to fossil fuels. Despite the advantages of biofuels, such as being renewable, environmentally friendly, easy to source, and reducing the dependency on foreign oil, there are several drawbacks of using biofuels including high cost, and other factors discussed in the fuel vs. food debate. Therefore, it is imperative to produce novel biofuels while also developing suitable manufacturing processes that ease the aforementioned problems. Polyhydroxyalkanoates (PHAs) are structurally diverse microbial polyesters synthesized by numerous bacteria. Moreover, this structural diversity allows PHAs to readily undergo methyl esterification and to be used as biofuels, which further extends the application value of PHAs. PHA-based biofuels are similar to biodiesel except for having a high oxygen content and no nitrogen or sulfur. In this article, we review the microbial production of PHAs, biofuel production from PHAs, parameters affecting the production of fuel from PHAs, and PHAs biorefineries. In addition, future work on the production of biofuels from PHAs is also discussed.
1. Introduction
Polyhydroxyalkanoates (PHAs) are bacterial polyesters synthesized by several prokaryotic microorganisms under unbalanced nutrition conditions, e.g., carbon and energy storage conditions. Lemoigne first discovered these thermoplastics in 1926, and since then, PHAs have attracted significant commercial and research interest from the eco-friendly polymer market due to their favorable properties such as high biocompatibility, excellent biodegradability, diverse chemical structure, and manufacturing from renewable carbon resources [1,2,3,4,5].
Bio-based polyesters are considered alternates for petrochemical plastics. Moreover, among the various biopolymers, PHAs symbolize a sustainable future alternative for fossil fuel-based plastics. PHAs have mostly been synthesized by microbial cultures grown on renewable materials in clean environments [6,7,8,9]. Fossil fuels are energy recourses that fulfill most of the world’s energy requirements. However, their production and use cause severe health and environmental problems including global warming and pollution. Consequently, plant and animal-based fuels (also termed as biofuels), such as biogas, biodiesel, and many others, have been introduced as alternatives to fossil fuels. Despite the advantages of biofuels, such as being renewable, environmentally friendly, easy to source, and reducing the dependency on foreign oil, there are several drawbacks to using biofuels including high cost, and other factors discussed in the fuel vs. food debate. Therefore, it is imperative to produce novel biofuels while also developing suitable manufacturing processes that ease the aforementioned problems. Polyhydroxyalkanoates (PHAs) are structurally diverse microbial polyesters synthesized by numerous prokaryotic microorganisms. PHAs are naturally synthesized bio-based materials. Since PHAs are biocompatible, bioresorbable, and biodegradable; hence, they have no negative effect on the environment. When PHAs-based products are left in the environment, they degraded into CO2, H2O, and CH4, which facilitate the natural cycle of circulatory and renewability. In addition, the spent PHAs can undergo hydrolysis and produce optically pure building blocks which can be used as synthons for pharmaceuticals, organic synthesis, and fragrances [10]. Moreover, the structural diversity of PHAs allows them to readily undergo methyl esterification and to be used as biofuels, which further extends the application value of PHAs.
2. Sources of PHAs
PHAs are long polymeric molecules that exhibit ester linkages in their structures. PHAs are synthesized by various types of prokaryotic microorganisms as intracellular storage materials comprising significant amounts of carbon and energy [11]. A PHA molecule is typically made up of 600 to 35,000 fatty acid monomer units. Each monomer unit consists of a side chain (R) group that is usually a saturated alkyl group but can also be unsaturated, branched, or substituted alkyl group [12,13,14]. Depending on the number of carbon atoms in the PHA monomer, these biopolymers can be classified into short-chain length (scl) PHAs (generally 3–5 carbon atoms), medium-chain length (mcl) PHAs (comprising 6–14 carbon atoms), and long-chain length (lcl) PHAs (contains 15 or more carbon atoms). More than 150 PHA monomers have been recorded, where the chemical or physical modification of existing PHAs led to the production of new types of PHAs. These features of PHAs gave rise to a variety of PHA properties suitable for a number of applications, such as biodegradable packaging materials and various medical products, as shown in Figure 1 (Lignocellulose (biomass-derived) PHAs and their applications) [2,15,16].
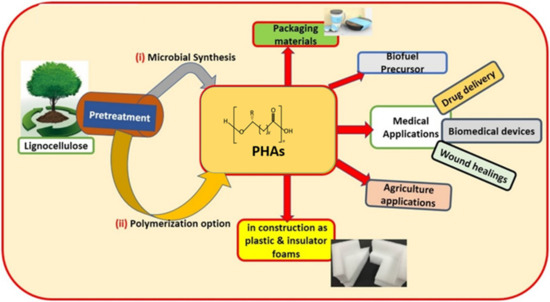
Figure 1.
Applications of polyhydroxyalkanoates (PHAs) in various fields. Reproduced with permission from [17]. Copyright 2020 Elsevier Ltd.
The synthesis of PHAs is a simple biosynthetic process consisting of only three genes and three enzymes. Figure 2 shows the PHAs biosynthetic pathway in bacteria. The Halobactericeae, a family of bacteria, contains more than 300 bacteria sources for PHA synthesis. Pseudomonas oleovorans and Pseudomonas fragi are bacteria that can produce mcl PHAs through the beta (β) oxidation of alkanoic acids by producing hydroxyalkanoyl-CoA as a substrate [17,18]. Furthermore, PHAs containing 2-hydroxypropionate (2HP), 3-hydroxypropionate (3HP), 4-hydroxybutyrate (4HB), 3-hydroxyvalerate (3HV), and mcl 3-hydroxyalkanoate (3-HA) monomer units can all be produced by Escherichia coli via genes or exogenous and endogenous processes [19,20,21]. In addition, the synthesis of PHAs was also studied in other eukaryotes, namely microalgae (both recombinant and even wild type). However, the bacterial synthesis of PHAs is not cost-effective compared to petroleum-based plastics; therefore; there is great interest in discovering a synthetic pathway for PHA production in eukaryotic cells, especially in crops [22,23,24].
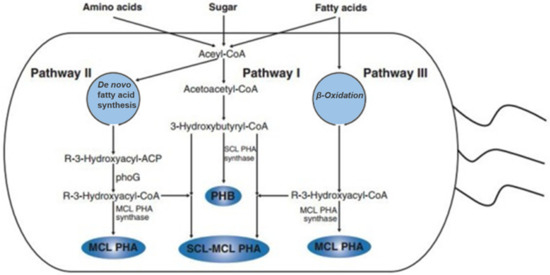
Figure 2.
Biosynthesis of PHAs in bacteria. Reproduced with permission from [25]. Copyright 2015 Elsevier Ltd.
2.1. Production of PHAs
In the 1980s, numerous companies tried to produce PHAs on an industrial scale based on the expectations that petroleum prices would increase over time. People would prefer to use eco-friendly biodegradable plastics or green plastics instead. Moreover, industries such as Chemie Linz AG, Austria; ICI, UK; and TianAn, China, successfully produced poly-(R)-3-hydroxybutyrate (P3HB) and several copolymers such as P3HBHHx of (R)-3-hydroxybutyrate (3HB), (R)-3-hydroxyhexanoate (3HHx), P3HBV of (R)-3-hydroxybutyrate (3HB), and (R)-3-hydroxyvalerate (3HV) [1]. However, up until the year 2000, petroleum prices did not increase as significantly as expected and many PHA production-related projects were discontinued. Surprisingly, after 2001 a sharp increase in the petroleum price was observed and in 2008, a barrel of oil reached $140, which reignited the search for petroleum-free plastics (Figure 3 shows the production of PHAs). Industry focused on two polyesters: polylactic acid (PLA) and PHAs, where PLA is economical and available in bulk but PHAs are expensive. However, the applications and properties of PHAs can be tailored by varying the co-monomer and the content of the copolyesters [26,27,28,29]. The advantages and disadvantages of both PLA and PHAs are compiled in Table 1.
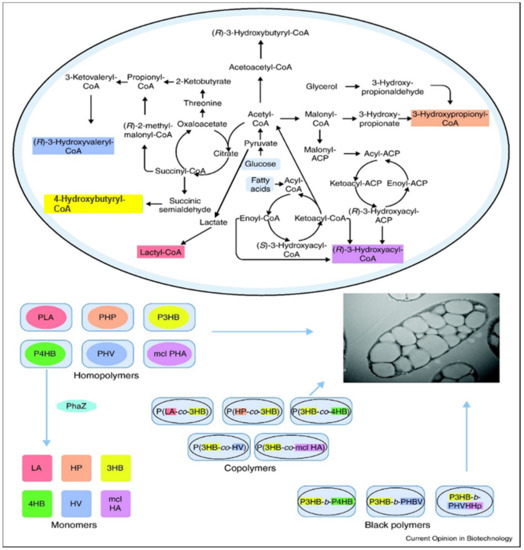
Figure 3.
Microbial synthesis of PHAs. Key metabolic paths for the production of precursors for PHAs Reproduced with permission from [30]. Copyright 2011 Elsevier Ltd.

Table 1.
Comparison of polyhydroxyalkanoates (PHA) and polylactic acid (PLA).
2.2. Fermentation Industry
Both wild type and recombinant bacteria are used to produce PHA (Table 2). The production of PHAs at an industrial scale has several prerequisites such as strain development, shake flask optimization, lab and pilot fermenter studies, and industrial production scale-up [31]. Furthermore, several factors affect the microbial production of PHAs including growth rate, cell density, percentage of PHAs in cell dry weight, the time required to attain final cell density, choice of substrate, price of the substrate, and the use of an economical method for PHAs extraction and purification [32,33,34,35].
Table 2.
Bacterial strains for pilot and large-scale production of PHAs.
2.3. Microbial Synthesis of PHAs Homopolymers
A homopolymer can be defined as a polymer that comprises >99% by weight of one type of monomer and <1% by weight of another. PHAs can be produced as homopolymers, copolymers, or blends depending on the bacterial strain or growth substrate used, and more than 300 species, mainly of bacteria, are reported to produce these polymers [36]. The bacteria that produce PHAs are categorized into two groups based on the stressed conditions required for PHA synthesis. One group requires the limitation of essential nutrients such as nitrogen (N), sulfur (S), magnesium (Mg), or phosphorus (P) with excessive carbon source supply. A. eutrophus, Protomonas extorquens, and P. oleovorans are included in this group. On the other hand, the second group does not require the limitation of essential nutrients bacteria (commonly known as growth-associated PHA biosynthesis). Alcaligenes latus, a mutant strain of Azotobacter vinelandii and recombinant E. coli belongs to the second group of bacteria [37]. P3HB, the most common homopolymer of PHA, is synthesized by wild type bacteria. Other homopolymers includes P3HP [31,38], P4HP [39], poly(3-hydroxyvalerate) (PHV), [40] poly(3-hydroxy-5-phenylvaleric acid) (P3H5PV) [41] poly(3-hydroxyhexanoate) (PHHx) [42], poly(3-hydroxyheptanoate) (PHHp) [43], poly(3-hydroxyoctanoate) (PHO) [44], poly(3-hydroxynonanoate) (PHN [45]), poly(3-hydroxydodecanoate) (PHD) [46], and poly(3-hydroxydodecanoate) (PHDD) [30,47]. These homopolymers have not yet been fully investigated due to their low contents in cells. The mcl-PHA homopolyesters are typically produced only by recombinant strains, e.g., β-oxidation weakened mutants [48].
Cupriavidus necator and recombinant E. coli are the most used strains for the industrial production of PHAs. Pohlmann et al., discovered the production of PHA homopolymer and copolyers, including P3HB (over 80%), P3HB4HB (over 75%), and P3HBV (over 75%), by C. necator and E. coli. Moreover, these results led to further genetic manipulations of bacteria for the production of PHAs. A. latus is another strain (similar to R. eutropha) that was also used for the production of P3HB and P3HBV [49,50,51].
Wang et al., [52] and Liu et al., [53] used Pseudomonas putida KT2442, a well-studied producer of PHA, to synthesize PHA homopolymers including PHHx and PHHp. Moreover, they synthesized an almost homopolymer of poly(3-hydroxyoctanoate-co-2mol% 3-hydroxyhexanoate) (PHO*) when hexanoate, heptanoate, and octanoate were used in the growth medium. The phaG gene of P. putida KT2442, encoding for R-3-hydroxyacyl-ACP-CoA transacylase and β-oxidation related genes, had been deleted preventing fatty acid substrates’ shortening. This leads to the accumulation of mutant to produce mcl PHA homopolymers. On the other hand, when PHA synthase genes phaC1 and phaC2 were exchanged with PHA synthase operon phaPCJ from Aeromonas hydrophila 4AK4, or P3HBC from R. eutropha, PHV or P3HB were produced using valerate, butyrate, and γ-butyrolacton as substrates, respectively. In addition, Liu and Chan [54] successfully synthesized poly(3-hydroxytetradecanoate) (PHTD) using fadB and fadA knockout mutant of P. putida KT2442. In another study, Wang et al., [55] used an engineered strain of Pseudomonas entomophila LAC23 (designed from P. entomophila L48) and deficient β-oxidation pathways to produce mcl PHA homopolymers. The authors found that P. entomophila LAC23 could be used for the synthesis of different mcl PHAs when using appropriate fatty acids as carbon sources (Figure 4) 3-hydroxytetradecanoates (3HTD) was successfully synthesized when tetradecanoic acid was used as carbon source. Moreover, the authors were able to produce C7-C14 PHA homopolymers using P. entomophila LAC23 grown on appropriate C7-C14 fatty acids.
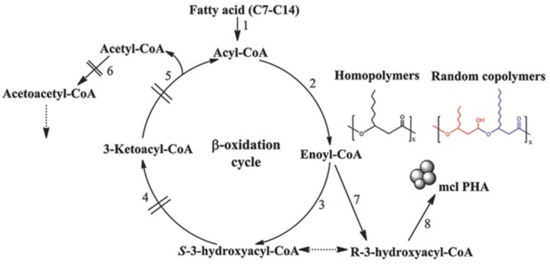
Figure 4.
Production of mcl-PHA homopolymers by the β-oxidation of impaired Pseudomonas entomophila LAC23. Reproduced with permission from [55]. Copyright 2017 John Wiley & Sons, Inc.
Sharma et al., [56] used recombinant P. putida LS461, with the phaC1pha ZphaC2 genes deleted, developed by involving JC123 carrying phaC116 PHA synthase. The resulting strain, P. putida LS46123, was able to produce P3HB or PHV PHA homopolymers when either glucose or free fatty acids were used as a carbon source. Wu et al., [57] demonstrated the production of P3HB Rhodopseudomonas palustris WP3-5 when acetate and propionate were used as carbon sources. Scheel et al., [58] developed recombinant E. coli strains by deleting arcA and ompR; two global regulators capable of preventing the uptake and activation of exogenous fatty acids. It was found that the yields of homopolymers P3HB, PHV, and PHHx increased significantly with only a modest increase in PHHp and PHD in ΔarcA mutant compared to the parental strain. Thus, P. putida KT2442, P. putida LS461, P. entomophila LAC23, R. palustris WP3-5, and E. coli-based derivatives can produce PHAs homopolymers. Table 3 summarizes the production of PHA homopolymers by different strains.
Table 3.
Microbial production of PHAs homopolymers.
2.4. Microbial Synthesis of PHAs Copolymers
When two or more different types of monomers are linked in the same polymer chain, copolymers are formed [84]. All the mcl PHAs are copolymers of C6–C12. P3HB is the most commonly used PHA; due to its exceptional properties, such as high thermal stability and excellent mechanical properties, P3HB based copolymers have attracted industrial interest [6]. Various copolymers such as P3HBV, P3HB4HB, and P3HBHHx have been produced on an industrial scale for several applications, but the non-3-hydroxybutyrate (3HB) monomer of these copolymers is not economical and adds additional cost to the overall synthesis. For example, to produce copolymers of 3-hydroxyvalerate (3HV), 4-hyroxybutyrate (4HB), and 3-hydroxyhexanoate (3HHx) based on 3HB, propionate, 1,4-butandiol, and lauric acid are required. Moreover, these non-3HB monomers are toxic and challenging to control in the cell growth processes. As a consequence, researchers started using genetically engineered metabolic pathways and low-cost non-fatty acid substrates, such as glycerol, gluconate, and glucose, for the production of PHA copolymers [48,85].
Koller et al. [86], reported the synthesis of PHBV from 3HV-unrelated resources glucose and glycerol without the genetic manipulation of Haloferax mediterranei strain. Aldor et al., [87] successfully synthesized P3HBV by adopting a metabolically engineered pathway. The authors used a strain of Salmonella enterica serovar Typhimurium to metabolize propionyl coenzyme A (propionyl-CoA). The S. enterica accumulated significant amounts of P3HBV when grown aerobically using glycerol as a glucose source. Li et al., [85] constructed a metabolically engineered E. coli strain for the synthesis of P3HB4HB using glucose as a carbon source. The genes responsible for succinate degradation and accumulation of P3HB in Clostridium kluyveri and R. eutropha, respectively, were co-expressed for the synthesis of P3HB. Additionally, the sad and gabD genes of E. coli were deleted for the synthesis of the 4HB monomer. In another study, Zheng et al., [88] genetically engineered A. hydrophila 4AK4 and P. putida GPp104 for the production of P3HBHHx copolymers using gluconate and glucose as carbon sources. A truncated gene (tesA) encoding for cytosolic thioesterase I of E. coli, which is used for the conversion of acyl-ACP into free fatty acids, was incorporated into A. hydrophila 4AK4. The recombinant strain was able to synthesize 10% and 19% (w/w) P3HBHHx when grown on gluconate and glucose, respectively. Table 4 summarizes the production of PHA copolymers by different recombinant strains. Sudo et al., reported the synthesis of (2-hydroxyalkanoate-co-3-hydroxybutyrate) (P2HA-co-P3HB) in engineered E. coli using glucose as a carbon source and hydrophobic amino acids as supplementation. Four different amino acids, i.e., leucine, valine (Val), isoleucine (Ile), and phenylalanine, were introduced into the growth medium and P2HB-co-P3HB production was observed when Val was used as supplementation. Figure 5 shows the biosynthetic pathway of P2HB-co-P3HB in E. coli from biomass-derived carbon sources.
Table 4.
Microbial production of PHAs copolymers.
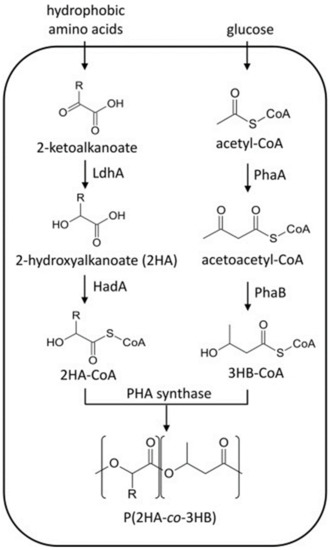
Figure 5.
Poly (2-hydroxyalkanoate-co-3-hydroxybutyrate) (P2HA-co-P3HB) biosynthesis pathways in engineered Escherichia coli. Reproduced with permission from [113]. Copyright 2020 Elsevier Ltd.
2.5. Microbial Production of PHA Block Copolymers
A polymer that contains alternating segments of different polymer compositions, covalently linked via their reactive ends, is called block copolymers. Block copolymers can be in the form of diblock, triblock or repeating multiblocks. Studies have shown that PHA homopolymers and copolymers exhibit many unfavorable properties and a common setback in the commercialization of these polymers is their brittleness, which limits their end-use applications [114]. Therefore, significant research focus has been placed on developing a suitable combination of these PHA polymers to achieve desirable properties [115]. Pederson et al., [116] and McChalicher et al., [115] successfully synthesized P3HB-b-P3HBV block copolymers in C. necator with the periodic addition of substrates (fructose and pentanoic acid). The P3HB and P3HBV blocks were synthesized during the fructose utilization and pulse feed of pentanoic acid. Though, the P3HB-b-P3HBV block copolymers still exhibited brittleness due to the brittle nature of scl PHAs. However, the combination of scl and mcl block copolymers could result in better properties compared to pure scl or mcl PHAs copolymers. In this context, Li et al., [117] reported the synthesis of P3HB-b-PHVHHp (P3HB-b-poly(3-hydroxyvalerate-co-3-hydroxyheptanoate) (PHVHHp)) block copolymers by recombinant P. putida KTOY06ΔC (phaPCJ A.c ). The authors demonstrated the formation of block copolymers when butyrate (C4) and heptanoate (C7) were introduced into the growth media. Keeping this in view, Tripathi et al., [118] adopted the same strategy and microbially linked scl-P3HB and mcl-HHx to synthesize P3HB-b-PHHx diblock copolymers in a β-oxidation deficient recombinant P. putida KT2442. The β-oxidation cycle was deleted to its maximum and glycerol was used as a carbon source. Recently, Sudo et al., [113] were able to synthesize poly(2-hydroxybutyrate) (P2HB)-b-P3HB polymers in E. coli using glucose as carbon source and hydrophobic amino acids as supplementation. The authors demonstrated that block copolymers successfully accumulated in genetically engineered E. coli by expressing the chimeric PHA synthase (PhaCAR) gene. It was demonstrated that PhaCAR exhibits strict substrate specificity and that it can synthesize P2HB-b-P3HB in engineered E. coli from exogenous 2HB and 3HB. Moreover, 2-hydroxyalkanoate (2HA) units were incorporated using PhaCAR, the lactate dehydrogenase (LdhA), and CoA transferase (HadA). Four hydrophobic amino acids, leucine, Val, Ile, and phenylalanine, were introduced into the culture as supplementation and the required block copolymer was produced. Figure 6 shows the biosynthetic pathway of P2HB-b-P3HB in E. coli from biomass-derived carbon sources.
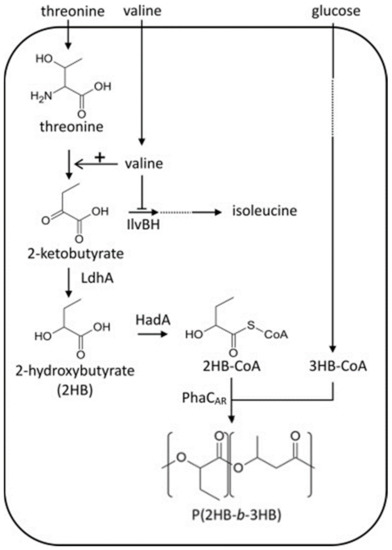
Figure 6.
Poly(2-hydroxybutyrate-b-3-hydroxybutyrate) (P2HB-b-P3HB) biosynthesis pathways in engineered E. coli with amino acid supplementation. Amino acids are also supplied through the endogenous pathways. Reproduced with permission from [113]. Copyright 2020 Elsevier Ltd.
3. Biofuels Based on PHAs
Due to the scarceness of traditional fossil fuels, increased emissions of combustion pollutants, and their costs, biomass-based fuels have become an attractive alternative option. Biofuels refer to the fuels produced from organic materials such as plants and animals. Table 5 lists the types of biofuels used for energy generation [119]. Studies have shown that the depletion of fossil fuels and environmental concerns increased the interest among scientists to explore renewable biofuels that are environmentally friendly and more acceptable. More importantly, in recent years the worldwide interest in biofuels has increased enormously [120]. To date, several biofuels derived from biomass have been explored including hydrogen (H2), methane (CH4), methanol (CH3OH), ethanol (C2H5OH), biodiesel, acetone (C3H6O), and several others. Globally, the transportation industry is a major source of carbon dioxide (CO2) emission and energy consumption. Liquid biofuels can replace fossil fuels in the transportation industry [121]. Liquid biofuels, such as biodiesel and bioethanol, have numerous advantages including ease of production from common biomass, eco-friendly, biodegradable, and sustainability. However, the extensive use of these biofuels created the food vs. fuel controversy, while their high production costs restricted their large-scale application in the transportation industry.
Table 5.
Types of biofuels for energy.
In 2009, Zhang et al., [122] introduced a novel biofuel, hydroxyalkanoates methyl ester (HAME) and hydroxybutyrate methyl ester (HBME), derived from bacterial PHAs (Figure 7). HA esters have chemical structures similar to those of biofuels, especially biodiesel, consisting of the methyl ester of long-chain fatty acids. HA esters are different from petroleum: petroleum is enriched with low oxygen (O2) and high nitrogen (N2) and sulfur (S) contents leading to the generation of environmental pollutants upon combustion. In contrast, HA esters have a high O2 content without any N2 or S. In general, oxygenated additives can reduce the exhaust smoke for diesel as well as decrease combustion times and ignition delay. However, oxygenated additives, like HA esters, have a low cetane number and a high heat of vaporization which makes it difficult to completely fuel diesel engines. Therefore, the introduction of HA esters as sustainable fuels or fuel additives contributed to the divergence of the biofuel market [122]. In contrast, the use of PHAs as biofuel sources becomes very promising, as highly purified PHAs are not needed. Thus, PHAs can be produced from the sewage or industrial wastewater, which is not in competition with human or animal food sources, leading to cost reduction.
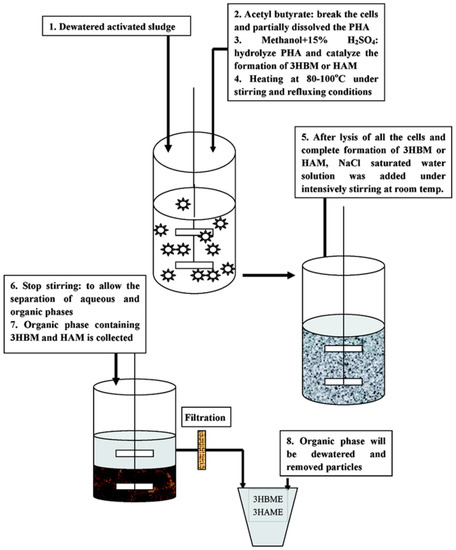
Figure 7.
Production of PHA based biofuels hydroxyalkanoates methyl ester (HAME) or hydroxybutyrate methyl ester (HBME). Reproduced with permission from [122]. Copyright 2009 American Chemical Society.
Zhang et al., [122] reported the production of HAME or HBME via the esterification of mcl PHAs and P3HB by acid-catalyzed hydrolysis (Figure 8). Combustion heat is one of the most important benchmarks to assess the quality of a fuel. The combustion heat of HAME, HBME, and their blends with diesel, gasoline, and C2H5OH were compared. The results demonstrated that when HBME was blended with C2H5OH, the combustion heat of C2H5OH improved by up to 30%. However, the combustion heat was reduced when HBME was blended with gasoline and diesel. Furthermore, HBME and HAME were obtained via acid-catalyzed hydrolysis and the most commonly used acid catalyst is sulfuric acid (H2SO4). Moreover, until now, different concentrations of H2SO4 with methanol were used to reach the maximum yield of HBME or HAME.
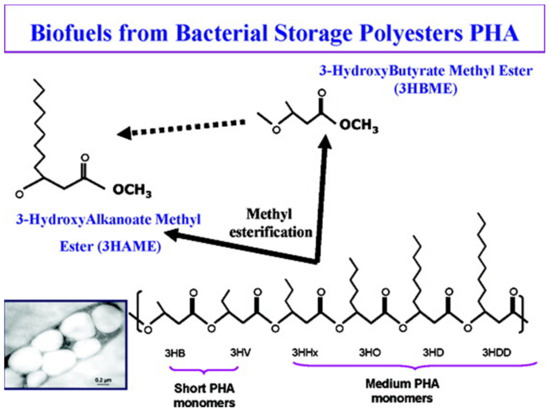
Figure 8.
Methyl esterification of various PHA monomers to produce PHA-based biofuels. Reproduced with permission from [122]. Copyright 2009 American Chemical Society.
Choonut et al., [123] documented the optimized conditions for HBME production from P3HB by acid-catalyzed hydrolysis. They produce P3HB from A. eutrophus TISTR 1107 and used it as a substrate to produce HBME using three acids (H2SO4, HCl, and H3PO4) at three different concentrations (5, 10, and 15%) and with different types of solvents including methanol and ethanol. The production of HBME was performed at various reaction times of 10, 20, 30, 40, 50, 60, and 70 h. Moreover, the optimal conditions to produce HBME were methanol with 10% (v/v) H2SO4 at 67 °C for 50 h. The maximum yield of HBME (70.7%) was obtained under these specific conditions. Sangkharak et al., [124] reported the production of HBME from P3HB. P3HB was isolated from Bacillus licheniformis M2-12 and used as a biofuel substrate. P3HB underwent esterification under acid- and base-catalyzed reaction conditions. The acid-catalyzed reaction proceeded using methanol with 10% (v/v) H2SO4 at 67 °C for 60 h. In contrast, the base-catalyzed hydrolysis of P3HB was performed using methanol with 2% (w/w) potassium hydroxide at 67 °C for 60 h and exhibited a high recovery of 68%. The fuel-related properties and purity (>95%) of the HBME produced from the acid-catalyzed hydrolysis of P3HB were investigated. The high molar mass, carbon and oxygen content, freezing point, and density of HBME showed that it can be potentially used as novel biofuel or as a fuel additive. The increasing need for biofuels may also increase the demand for biofuel additives or lubricants. Keunun et al., [125] reported the accumulation of P3HB in C. necator, by a two-step fermentation process that utilizes this P3HB as a novel substrate, for the synthesis of biolubricants. The lubricant was produced via a two-step transesterification process.
4. Parameters Affecting HAME-Based Biofuels Production
Several factors that affect the yield of HAME biofuels have been identified. These include reaction time, temperature, type and content of alcohol, as well as catalyst type and concentration.
4.1. Reaction Time
The conversion of fatty acids to esters is directly proportional to the reaction time. At the start, the dispersion of alcohols into the PHAs slows down the reaction rate, however, the reaction rate increases after thorough mixing. In general, the product yield reaches a maximum in < 90 min, after which it remains relatively constant upon a further increase in reaction time. Choonut et al., [123] studied the effect of reaction time (i.e., 10, 20, 30, 40, 50, 60, and 70 h) on the conversion of P3HB into HBME. Results showed that while the yield of HBME increased from 12.8 to 70.7% with increasing reaction time from 10 h to 50 h, the yield decreased upon a further increase in reaction time (60 and 70 h). Hence, a long reaction time is needed to produce HBME from P3HB under optimum reaction conditions. Okwundu et al., [126] reported the production of biodiesel from low free fatty acid beef tallow using homogenous and heterogeneous base-catalyzed reactions while documenting the effect of reaction time on the yield. The homogeneous and heterogeneous catalysts give a maximum biodiesel yield of 94.2% and 87.5% at 1 h and 4 h reaction time, respectively.
4.2. Reaction Temperature
Studies have shown that a high reaction temperature decreases the viscosity of fatty acids, which leads to an increase in reaction rate and a reduced reaction time. However, increasing the reaction temperature above a certain level results in a decrease in yield due to the saponification of triglycerides. In general, the reaction temperature must be lower than the boiling point of alcohol used. Base-catalyzed alcoholysis is normally performed near the boiling point of the alcohol or even at room temperature [74]. Zhang et al., [69] obtained a maximum yield (52%) of HBME from P3HB at 100 °C. In contrast, Wang et al., [75], Keunun et al., [72], Sangkharak et al., [71], and Choonut et al., [70] perform the esterification reaction of P3HB at 67 °C and obtained maximum yields for HBME of 40, 65, 68, and 70%, respectively. Overall, the optimal temperature ranges for the esterification of P3HB ranges from 67 to 100 °C, depending on other reaction parameters.
4.3. Type and Content of Alcohol
The alcohols used in the esterification reactions include CH3OH, C2H5OH, propanol (C3H7OH), and butanol (C4H9OH), of which CH3OH and C2H5OH are the most used. These two alcohols are the first choice for laboratory-scale research owing to their low cost and physical and chemical properties. In general, a higher amount of alcohol leads to a higher esterification conversion of the polyesters to biofuels. Weerachanchai et al., [127] perform the acid-catalyzed esterification of palm shell oil and investigated the effect of alcohol type and content on the esterification conversion. The results demonstrated that a 3.25:1 mole ratio of CH3OH or C2H5OH to acid under optimum conditions led to a 73.39% and 54.80% esterification conversion, respectively. However, upon a twofold increase in the mole ratio of alcohol, the conversion only increased by 2.42%. CH3OH gave higher esterification conversion compared to C2H5OH, which could be ascribed to the length of the alkyl group (C1 vs. C2) where the higher solubility and activity of the scl alcohol (CH3OH) promotes a faster reaction rate. Long-chain alcohols result in longer reaction times at the same reaction temperature of the scl alcohols, or shorter reaction time at higher reaction temperatures [128]. Choonut et al., [123] studied the effect of the alcohol used (CH3OH and C2H5OH) on the esterification of P3HB to produce HBME and found that CH3OH with 10% acid catalyst gives the highest yield.
4.4. Catalyst Type and Concentration
In general, acid- and base-catalyzed systems are used for the esterification of polyesters. Acid-catalyzed systems are mostly used to produce HAME biofuels, for which H2SO4 is the most frequently used catalyst. However, other acids such as H3PO4, organic sulfonic acids, and HCl are also used. Zhang et al., [69], Wang et al., [75], Keunun et al., [72], and Sangkharak et al., [71] investigated the esterification of PHAs to produce HAME using H2SO4 as catalyst. Choonut et al., studied the effect of different types of acids, including H3PO4, H2SO4, and HCl, on the conversion of P3HB to HBME. Their findings suggested that an acid-catalyzed system containing H2SO4 as catalyst gives maximum yield.
Catalyst concentration affects the yield of HAME biofuels. In general, the product yield increases when a 10% (v/v) acid concentration is used, but decrease upon a further increase in catalyst concentration. Zhang et al., [69], studied PHAs conversion to HAME using 15% H2SO4 and reported a 52% conversion. In contrast, Wang et al., [75], Keunun et al., [72], Sangkharak et al., [71], and Choonut et al., [70] carried the conversion of P3HB to HBME and reported maximum yield at this concentration.
5. PHAs Biorefineries
The United Nations identified numerous crops worldwide that can be used as bioenergy feedstocks. Many of these crops have the potential for coproducing PHAs. In the crop-based synthesis approach of PHAs, a metabolically engineered system is established in one crop and, at the same time, generic technology is created which can be transferred to a different biomass crop. In this way, additional crops will be harvested and commercialized in different regions based on need [129,130]. The sugarcane industry was the first PHAs producing biorefinery. This industry faced many challenges, which were clearly relevant to dedicated bioenergy crops where the residual bagasse is combusted to produce steam and power for the process while excess power is returned to the power grid [80]. The major advantage of sugarcane is that 42% of the total dry weight of the stalk can be extracted to fermentable sugar. However, another bioenergy crop, switchgrass, holds potential for multigene expression systems to coproduce PHAs. Moreover, switchgrass holds the advantage over sugarcane as it can be grown in less moist environments and can be processed year-round, leading to overall improved capital efficiency. Moreover, switchgrass is a high yield crop, meaning it can be harvested on land set aside for marginal use of other crops, needs few chemical supplies, can potentially reduce the runoff of topsoil, pesticide, and fertilizers, and fixes CO2 in its roots systems [81] (Figure 9).
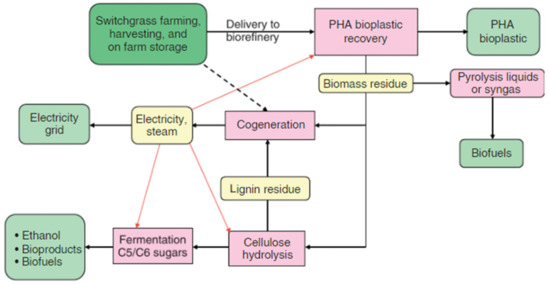
Figure 9.
Biorefineries for the production of PHAs from switchgrass. Reproduced with permission from [131]. Copyright 2009 Society of Chemical Industry and John Wiley & Sons, Ltd.
In summary, the solvent extraction process and residual dry biomass of switchgrass are used to recover PHAs and to produce bioenergy in cogeneration (cogeneration offers efficient generation of both steam and electricity while the excess steam can be used in other biorefinery processes) [82]. Unlike fully integrated switchgrass-based PHAs biorefinery, PHAs biorefinery can be as simple as the installation of a recovery system near an existing power plant which provides a reliable economical supply of dry and easily handled feedstock. Over time, additional capital, such as bioproduct capacity, biofuel, and cellulose hydrolysis, can be added to this emerging integrated PHA biorefinery [132,133]. In addition, biomass residues and fresh biomass could be converted to biofuel via thermochemical processes. According to the literature, the greenhouse gas emissions from switchgrass-based cellulosic C2H5OH is 94% less than that of gasoline [84]. Hence, PHAs are value-added coproducts obtained from the large scale production of biofuels and bioenergy from plants. Therefore, the synthesis of these biopolymers from biomass crops can considerably improve the economics of biomass biorefineries to produce liquid biofuels and bioenergy [131,134,135].
6. Conclusions and Future Directions
In conclusion, it has become clear that PHA-based biofuels are creating industrial value in energy, and the development of PHAs can address three basic issues:
- Petroleum shortage for plastic materials.
- Reduced CO2 emissions.
- Environmental protection.
As a source of biofuels, PHAs are very promising biopolyesters since they do not need to be of high purity. Thus, PHAs can be obtained from crops, activated sludge, or nutritious wastewater making it cost-effective while also addressing the food vs. fuel and fuel vs. land controversies. Hence, PHAs are novel substrates for biofuel production where recent developments in PHAs synthesis concerning open and continuous mixed cultures will produce cost-effective PHAs for the biofuel applications. After being used as bioplastics, PHAs can undergo methyl esterification to biofuels, which further extends their application value. However, much work is still needed to make PHAs cost-effective so that PHAs-based biofuels can be used as an alternative to existing biofuels such as biodiesel, ethanol, methane gas, and hydrogen.
In the future, the production costs of PHAs should be taken into consideration to enhance their commercialization. The production of PHAs in activated sludge or wastewater should be improved. So far, P3HB is mostly used in the production of HAME biofuels, thus, more monomers should be explored and tested for esterification. It would be worthwhile to develop HAME into biolubricants or fuel additives for the diversification of the biofuel or fuel additive industry. In addition, upcoming research should also focus to improve the quality of HAME biofuels.
Author Contributions
Conceptualization, S.R. and S.J.P.; writing—original draft preparation, S.R.; writing—review and editing, S.J.P. and K.Y.R.; supervision, S.J.P.; project administration, S.J.P.; funding acquisition, K.Y.R. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Technology Innovation Program (or Industrial Strategic Technology Development Program- Development of technology on materials and components) (20010106, Adhesives with low water permeability and low outgassing) funded By the Mistry of Trade, Industry & Energy (MOTIE, Korea) and supported by the Technological Innovation R&D Program (S2829590) funded by the Small and Medium Business Administration (SMBA, Korea).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Chen, G.-Q. A microbial polyhydroxyalkanoates (PHA) based bio-and materials industry. Chem. Soc. Rev. 2009, 38, 2434–2446. [Google Scholar] [CrossRef] [PubMed]
- Sudesh, K.; Abe, H.; Doi, Y. Synthesis, structure and properties of polyhydroxyalkanoates: Biological polyesters. Prog. Polym. Sci. 2000, 25, 1503–1555. [Google Scholar] [CrossRef]
- Chanprateep, S. Current trends in biodegradable polyhydroxyalkanoates. J. Biosci. Bioeng. 2010, 110, 621–632. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.-Q.; Patel, M.K. Plastics derived from biological sources: Present and future: A technical and environmental review. Chem. Rev. 2012, 112, 2082–2099. [Google Scholar] [CrossRef] [PubMed]
- Lemoigne, M. Produits de deshydration et de polymerisation de l’acide β= oxybutyrique. Bull. Soc. Chim. Biol. 1926, 8, 770–782. [Google Scholar]
- Raza, Z.A.; Abid, S.; Banat, I.M. Polyhydroxyalkanoates: Characteristics, production, recent developments and applications. Int. Biodeterior. Biodegrad. 2018, 126, 45–56. [Google Scholar] [CrossRef]
- Ma, Y.-M.; Wei, D.-X.; Yao, H.; Wu, L.-P.; Chen, G.-Q. Synthesis, characterization and application of thermoresponsive polyhydroxyalkanoate-graft-poly (N-isopropylacrylamide). Biomacromolecules 2016, 17, 2680–2690. [Google Scholar] [CrossRef]
- Matsumoto, K.i.; Hori, C.; Fujii, R.; Takaya, M.; Ooba, T.; Ooi, T.; Isono, T.; Satoh, T.; Taguchi, S. Dynamic changes of intracellular monomer levels regulate block sequence of polyhydroxyalkanoates in engineered Escherichia coli. Biomacromolecules 2018, 19, 662–671. [Google Scholar] [CrossRef]
- Mizuno, S.; Enda, Y.; Saika, A.; Hiroe, A.; Tsuge, T. Biosynthesis of polyhydroxyalkanoates containing 2-hydroxy-4-methylvalerate and 2-hydroxy-3-phenylpropionate units from a related or unrelated carbon source. J. Biosci. Bioeng. 2018, 125, 295–300. [Google Scholar] [CrossRef]
- Koller, M.; Mukherjee, A. Polyhydroxyalkanoates—Linking properties, applications, and end-of-life options. Chem. Biochem. Eng. Q. 2020, 34, 115–129. [Google Scholar] [CrossRef]
- David, Y.; Joo, J.C.; Yang, J.E.; Oh, Y.H.; Lee, S.Y.; Park, S.J. Biosynthesis of 2-hydroxyacid-containing polyhydroxyalkanoates by employing butyryl-CoA transferases in metabolically engineered Escherichia coli. Biotechnol. J. 2017, 12, 1700116. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.; Hu, D.; Che, X.; Jiang, X.; Li, T.; Chen, J.; Zhang, H.M.; Chen, G.-Q. Engineering of Halomonas bluephagenesis for low cost production of poly (3-hydroxybutyrate-co-4-hydroxybutyrate) from glucose. Metab. Eng. 2018, 47, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.J.; Chae, C.G.; Kang, K.H.; Oh, Y.H.; Joo, J.C.; Song, B.K.; Lee, S.Y.; Park, S.J. Biosynthesis of Lactate-containing polyhydroxyalkanoates in recombinant Escherichia coli by employing new CoA transferases. KSBB J. 2016, 31, 27–32. [Google Scholar] [CrossRef][Green Version]
- Reddy, C.; Ghai, R.; Kalia, V.C. Polyhydroxyalkanoates: An overview. Bioresour. Technol. 2003, 87, 137–146. [Google Scholar] [CrossRef]
- Verlinden, R.A.; Hill, D.J.; Kenward, M.; Williams, C.D.; Radecka, I. Bacterial synthesis of biodegradable polyhydroxyalkanoates. J. Appl. Microbiol. 2007, 102, 1437–1449. [Google Scholar] [CrossRef]
- Khanna, S.; Srivastava, A.K. Recent advances in microbial polyhydroxyalkanoates. Process. Biochem. 2005, 40, 607–619. [Google Scholar] [CrossRef]
- Adeleye, A.T.; Odoh, C.K.; Enudi, O.C.; Banjoko, O.O.; Osigbeminiyi, O.O.; Toluwalope, O.E.; Louis, H. Sustainable synthesis and applications of polyhydroxyalkanoates (PHAs) from biomass. Process. Biochem. 2020, 96, 174–193. [Google Scholar] [CrossRef]
- Tamang, P.; Banerjee, R.; Köster, S.; Nogueira, R. Comparative study of polyhydroxyalkanoates production from acidified and anaerobically treated brewery wastewater using enriched mixed microbial culture. J. Environ. Sci. 2019, 78, 137–146. [Google Scholar] [CrossRef]
- Matias, F.; Rodrigues, M.F.D.A. New PHA products using unrelated carbon sources. Braz. J. Microbiol. 2011, 42, 1354–1363. [Google Scholar] [CrossRef]
- Pavan, F.A.; Junqueira, T.L.; Watanabe, M.D.; Bonomi, A.; Quines, L.K.; Schmidell, W.; de Aragao, G.M. Economic analysis of polyhydroxybutyrate production by Cupriavidus necator using different routes for product recovery. Biochem. Eng. J. 2019, 146, 97–104. [Google Scholar] [CrossRef]
- Harmaen, A.; Khalina, A.; Ali, H.M.; Azowa, I. Thermal, morphological, and biodegradability properties of bioplastic fertilizer composites made of oil palm biomass, fertilizer, and poly (hydroxybutyrate-co-valerate). Int. J. Polym. Sci. 2016, 2016, 3230109. [Google Scholar] [CrossRef]
- Gebelein, C.G. New and traditional polymers from biotechnology. In Biotechnology and Polymers; Springer: Berlin/Heidelberg, Germany, 1991; pp. 1–9. [Google Scholar]
- Francis, L.; Keshavarz, T.; Roy, I. Biosynthesis of polyhydroxyalkanoates and their applications. In Proceedings of the International Conference on Biodegradable Polymers: Their Production, Characterisation and Application, London, UK, 10 December 2007. [Google Scholar]
- Moradali, M.F.; Rehm, B.H. Bacterial biopolymers: From pathogenesis to advanced materials. Nat. Rev. Genet. 2020, 18, 195–210. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.-Q.; Hajnal, I.; Wu, H.; Lv, L.; Ye, J. Engineering biosynthesis mechanisms for diversifying polyhydroxyalkanoates. Trends Biotechnol. 2015, 33, 565–574. [Google Scholar] [CrossRef] [PubMed]
- Long, J.-Y.; Song, K.-L.; He, X.; Zhang, B.; Cui, X.-F.; Song, C.-F. Mutagenesis of phaR, a regulator gene of polyhydroxyalkanoate biosynthesis of Xanthomonas oryzae pv. oryzae caused pleiotropic phenotype changes. Front. Microbiol. 2018, 9, 3046. [Google Scholar] [CrossRef]
- Coats, E.R.; Watson, B.S.; Brinkman, C.K. Polyhydroxyalkanoate synthesis by mixed microbial consortia cultured on fermented dairy manure: Effect of aeration on process rates/yields and the associated microbial ecology. Water Res. 2016, 106, 26–40. [Google Scholar] [CrossRef] [PubMed]
- Pham, T.H.; Webb, J.S.; Rehm, B.H. The role of polyhydroxyalkanoate biosynthesis by Pseudomonas aeruginosa in rhamnolipid and alginate production as well as stress tolerance and biofilm formation. Microbiology 2004, 150, 3405–3413. [Google Scholar] [CrossRef]
- Gonzalez-Miro, M.; Chen, S.; Gonzaga, Z.J.; Evert, B.; Wibowo, D.; Rehm, B.H. Polyester as antigen carrier toward particulate vaccines. Biomacromolecules 2019, 20, 3213–3232. [Google Scholar] [CrossRef]
- Gao, X.; Chen, J.-C.; Wu, Q.; Chen, G.-Q. Polyhydroxyalkanoates as a source of chemicals, polymers, and biofuels. Curr. Opin. Biotechnol. 2011, 22, 768–774. [Google Scholar] [CrossRef]
- Wang, Q.; Liu, C.; Xian, M.; Zhang, Y.; Zhao, G. Biosynthetic pathway for poly (3-hydroxypropionate) in recombinant Escherichia coli. J. Microbiol. 2012, 50, 693–697. [Google Scholar] [CrossRef]
- Castilho, L.R.; Mitchell, D.A.; Freire, D.M. Production of polyhydroxyalkanoates (PHAs) from waste materials and by-products by submerged and solid-state fermentation. Bioresour. Technol. 2009, 100, 5996–6009. [Google Scholar] [CrossRef]
- Carvalho, G.; Oehmen, A.; Albuquerque, M.G.; Reis, M.A. The relationship between mixed microbial culture composition and PHA production performance from fermented molasses. New. Bitechnol. 2014, 31, 257–263. [Google Scholar] [CrossRef] [PubMed]
- Bengtsson, S.; Hallquist, J.; Werker, A.; Welander, T. Acidogenic fermentation of industrial wastewaters: Effects of chemostat retention time and pH on volatile fatty acids production. Biochem. Eng. J. 2008, 40, 492–499. [Google Scholar] [CrossRef]
- Bhubalan, K.; Lee, W.-H.; Loo, C.-Y.; Yamamoto, T.; Tsuge, T.; Doi, Y.; Sudesh, K. Controlled biosynthesis and characterization of poly (3-hydroxybutyrate-co-3-hydroxyvalerate-co-3-hydroxyhexanoate) from mixtures of palm kernel oil and 3HV-precursors. Polym. Degrad. Stab. 2008, 93, 17–23. [Google Scholar] [CrossRef]
- Olivera, E.R.; Carnicero, D.; Jodra, R.; Miñambres, B.; García, B.; Abraham, G.A.; Gallardo, A.; Román, J.S.; García, J.L.; Naharro, G. Genetically engineered Pseudomonas: A factory of new bioplastics with broad applications. Environ. Microbiol. 2001, 3, 612–618. [Google Scholar] [CrossRef]
- Anjum, A.; Zuber, M.; Zia, K.M.; Noreen, A.; Anjum, M.N.; Tabasum, S. Microbial production of polyhydroxyalkanoates (PHAs) and its copolymers: A review of recent advancements. Int. J. Biol. Macromol. 2016, 89, 161–174. [Google Scholar] [CrossRef]
- Linares-Pastén, J.A.; Sabet-Azad, R.; Pessina, L.; Sardari, R.R.; Ibrahim, M.H.; Hatti-Kaul, R. Efficient poly (3-hydroxypropionate) production from glycerol using Lactobacillus reuteri and recombinant Escherichia coli harboring L. reuteri propionaldehyde dehydrogenase and Chromobacterium sp. PHA synthase genes. Bioresour. Technol. 2015, 180, 172–176. [Google Scholar] [CrossRef]
- Valentin, H.E.; Schönebaum, A.; Steinbüchel, A. Identification of 4-hydroxyvaleric acid as a constituent of biosynthetic polyhydroxyalkanoic acids from bacteria. Appl. Microbiol. Biotechnol. 1992, 36, 507–514. [Google Scholar] [CrossRef]
- Steinbüchel, A.; Schmack, G. Large-scale production of poly (3-hydroxyvaleric acid) by fermentation of Chromobacterium violaceum, processing, and characterization of the homopolyester. J. Environ. Polym. Degrad. 1995, 3, 243–258. [Google Scholar] [CrossRef]
- Haywood, G.W.; Anderson, A.J.; Ewing, D.F.; Dawes, E.A. Accumulation of a polyhydroxyalkanoate containing primarily 3-hydroxydecanoate from simple carbohydrate substrates by Pseudomonas sp. strain NCIMB 40135. Appl. Environ. Microbiol. 1990, 56, 3354–3359. [Google Scholar] [CrossRef]
- Jian, J.; Li, Z.-J.; Ye, H.-M.; Yuan, M.-Q.; Chen, G.-Q. Metabolic engineering for microbial production of polyhydroxyalkanoates consisting of high 3-hydroxyhexanoate content by recombinant Aeromonas hydrophila. Bioresour. Technol. 2010, 101, 6096–6102. [Google Scholar] [CrossRef]
- Steinbüchel, A.; Valentin, H.E.; Schönebaum, A. Application of recombinant gene technology for production of polyhydroxyalkanoic acids: Biosynthesis of poly (4-hydroxybutyric acid) homopolyester. J. Environ. Polym. Degrad. 1994, 2, 67–74. [Google Scholar] [CrossRef]
- Anderson, A.J.; Haywood, G.W.; Dawes, E.A. Biosynthesis and composition of bacterial poly (hydroxyalkanoates). Int. J. Biol. Macromol. 1990, 12, 102–105. [Google Scholar] [CrossRef]
- Chung, C.-W.; Kim, Y.-S.; Kim, Y.-B.; Bae, K.-S.; Rhee, Y.-H. Isolation of a Pseudomonas sp. strain exhibiting unusual behavior of poly (3-hydroxyalkanoates) biosynthesis and characterization of synthesized polyesters. J. Microbiol. Biotechnol. 1999, 9, 847–853. [Google Scholar]
- Gumel, A.M.; Annuar, M.S.M.; Heidelberg, T. Biosynthesis and characterization of polyhydroxyalkanoates copolymers produced by Pseudomonas putida Bet001 isolated from palm oil mill effluent. PLoS ONE 2012, 7, e45214. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, L.; Wu, L.-P.; Dechuan, M.; Chen, J.; Wu, Q.; Chen, G.-Q. Pseudomonas putida KT2442 as a platform for the biosynthesis of polyhydroxyalkanoates with adjustable monomer contents and compositions. Bioresour. Technol. 2013, 142, 225–231. [Google Scholar] [CrossRef]
- Koller, M. Advances in Polyhydroxyalkanoate (PHA) Production; MDPI: Basel, Switzerland, 2017. [Google Scholar]
- Motallebi-Veshareh, M.; Balzer, D.; Lanka, E.; Jagura-Burdzy, G.; Thomas, C.M. Conjugative transfer functions of broad-host-range plasmid RK2 are coregulated with vegetative replication. Mol. Microbiol. 1992, 6, 907–920. [Google Scholar] [CrossRef]
- Chen, G.-Q.; König, K.-H.; Lafferty, R.M. Production of poly-D (-)-3-hydroxybutyrate and poly-D (-)-3-hydroxyvalerate by strains ofAlcaligenes latus. Antonie Van Leeuwenhoek 1991, 60, 61–66. [Google Scholar] [CrossRef]
- Pohlmann, A.; Fricke, W.F.; Reinecke, F.; Kusian, B.; Liesegang, H.; Cramm, R.; Eitinger, T.; Ewering, C.; Pötter, M.; Schwartz, E. Genome sequence of the bioplastic-producing “Knallgas” bacterium Ralstonia eutropha H16. Nat. Biotechnol. 2006, 24, 1257–1262. [Google Scholar] [CrossRef]
- Liu, Q.; Luo, G.; Zhou, X.R.; Chen, G.-Q. Biosynthesis of poly (3-hydroxydecanoate) and 3-hydroxydodecanoate dominating polyhydroxyalkanoates by β-oxidation pathway inhibited Pseudomonas putida. Metab. Eng. 2011, 13, 11–17. [Google Scholar] [CrossRef]
- Wang, H.-H.; Zhou, X.-R.; Liu, Q.; Chen, G.-Q. Biosynthesis of polyhydroxyalkanoate homopolymers by Pseudomonas putida. Appl. Microbiol. Biotechnol. 2011, 89, 1497–1507. [Google Scholar] [CrossRef]
- Liu, W.; Chen, G.-Q. Production and characterization of medium-chain-length polyhydroxyalkanoate with high 3-hydroxytetradecanoate monomer content by fadB and fadA knockout mutant of Pseudomonas putida KT2442. Appl. Microbiol. Biotechnol. 2007, 76, 1153–1159. [Google Scholar] [CrossRef]
- Wang, Y.; Chung, A.; Chen, G.Q. Synthesis of medium-chain-length polyhydroxyalkanoate homopolymers, random copolymers, and block copolymers by an engineered strain of Pseudomonas entomophila. Adv. Healthc. Mater. 2017, 6, 1601017. [Google Scholar] [CrossRef]
- Sharma, P.K.; Munir, R.I.; Blunt, W.; Dartiailh, C.; Cheng, J.; Charles, T.C.; Levin, D.B. Synthesis and physical properties of polyhydroxyalkanoate polymers with different monomer compositions by recombinant Pseudomonas putida LS46 expressing a novel PHA synthase (PhaC116) enzyme. Appl. Sci. 2017, 7, 242. [Google Scholar] [CrossRef]
- Scheel, R.A.; Ji, L.; Lundgren, B.R.; Nomura, C.T. Enhancing poly (3-hydroxyalkanoate) production in Escherichia coli by the removal of the regulatory gene arcA. AMB Express 2016, 6, 120. [Google Scholar] [CrossRef]
- Wu, S.C.; Liou, S.Z.; Lee, C.M. Correlation between bio-hydrogen production and polyhydroxybutyrate (PHB) synthesis by Rhodopseudomonas palustris WP3-5. Bioresour. Technol. 2012, 113, 44–50. [Google Scholar] [CrossRef]
- De Andrade Rodrigues, M.; Valentin, H.; Berger, P.; Tran, M.; Asrar, J.; Gruys, K.; Steinbüchel, A. Polyhydroxyalkanoate accumulation in Burkholderia sp.: A molecular approach to elucidate the genes involved in the formation of two homopolymers consisting of short-chain-length 3-hydroxyalkanoic acids. Appl. Microbiol. Biotechnol. 2000, 53, 453–460. [Google Scholar] [CrossRef]
- Yao, J.; Zhang, G.; Wu, Q.; Chen, G.-Q.; Zhang, R. Production of polyhydroxyalkanoates by Pseudomonas nitroreducens. Antonie Van Leeuwenhoek 1999, 75, 345–349. [Google Scholar] [CrossRef]
- Kato, M.; Bao, H.; Kang, C.-K.; Fukui, T.; Doi, Y. Production of a novel copolyester of 3-hydroxybutyric acid and medium-chain-length 3-hydroxyalkanoic acids by Pseudomonas sp. 61-3 from sugars. Appl. Microbiol. Biotechnol. 1996, 45, 363–370. [Google Scholar] [CrossRef]
- Kimura, H.; Ohura, T.; Takeishi, M.; Nakamura, S.; Doi, Y. Effective microbial production of poly (4-hydroxybutyrate) homopolymer by Ralstonia eutropha H16. Polym. Int. 1999, 48, 1073–1079. [Google Scholar] [CrossRef]
- Wang, H.-H.; Li, X.-T.; Chen, G.-Q. Production and characterization of homopolymer polyhydroxyheptanoate (P3HHp) by a fadBA knockout mutant Pseudomonas putida KTOY06 derived from P. putida KT2442. Process. Biochem. 2009, 44, 106–111. [Google Scholar] [CrossRef]
- Shen, X.-W.; Yang, Y.; Jian, J.; Wu, Q.; Chen, G.-Q. Production and characterization of homopolymer poly (3-hydroxyvalerate)(PHV) accumulated by wild type and recombinant Aeromonas hydrophila strain 4AK4. Bioresour. Technol. 2009, 100, 4296–4299. [Google Scholar] [CrossRef]
- Ramachander, T.; Rohini, D.; Belhekar, A.; Rawal, S. Synthesis of PHB by recombinant E. coli harboring an approximately 5 kb genomic DNA fragment from Streptomyces aureofaciens NRRL 2209. Int. J. Biol. Macromol. 2002, 31, 63–69. [Google Scholar] [CrossRef]
- Valappil, S.P.; Peiris, D.; Langley, G.; Herniman, J.; Boccaccini, A.R.; Bucke, C.; Roy, I. Polyhydroxyalkanoate (PHA) biosynthesis from structurally unrelated carbon sources by a newly characterized Bacillus spp. J. Biotechnol. 2007, 127, 475–487. [Google Scholar] [CrossRef]
- Rodríguez-Contreras, A.; Koller, M.; Miranda-de Sousa Dias, M.; Calafell-Monfort, M.; Braunegg, G.; Marqués-Calvo, M.S. Influence of glycerol on poly (3-hydroxybutyrate) production by Cupriavidus necator and Burkholderia sacchari. Biochem. Eng. J. 2015, 94, 50–57. [Google Scholar] [CrossRef]
- Pais, J.; Farinha, I.; Freitas, F.; Serafim, L.S.; Martínez, V.; Martínez, J.C.; Arévalo-Rodríguez, M.; Prieto, M.A.; Reis, M.A. Improvement on the yield of polyhydroxyalkanotes production from cheese whey by a recombinant Escherichia coli strain using the proton suicide methodology. Enzyme Microb. Technol. 2014, 55, 151–158. [Google Scholar] [CrossRef]
- Kahar, P.; Tsuge, T.; Taguchi, K.; Doi, Y. High yield production of polyhydroxyalkanoates from soybean oil by Ralstonia eutropha and its recombinant strain. Polym. Degrad. Stab. 2004, 83, 79–86. [Google Scholar] [CrossRef]
- Chanprateep, S.; Buasri, K.; Muangwong, A.; Utiswannakul, P. Biosynthesis and biocompatibility of biodegradable poly (3-hydroxybutyrate-co-4-hydroxybutyrate). Polym. Degrad. Stab. 2010, 95, 2003–2012. [Google Scholar] [CrossRef]
- López-Cuellar, M.; Alba-Flores, J.; Rodríguez, J.G.; Pérez-Guevara, F. Production of polyhydroxyalkanoates (PHAs) with canola oil as carbon source. Int. J. Biol. Macromol. 2011, 48, 74–80. [Google Scholar] [CrossRef]
- Koller, M.; Bona, R.; Chiellini, E.; Fernandes, E.G.; Horvat, P.; Kutschera, C.; Hesse, P.; Braunegg, G. Polyhydroxyalkanoate production from whey by Pseudomonas hydrogenovora. Bioresour. Technol. 2008, 99, 4854–4863. [Google Scholar] [CrossRef]
- Althuri, A.; Mathew, J.; Sindhu, R.; Banerjee, R.; Pandey, A.; Binod, P. Microbial synthesis of poly-3-hydroxybutyrate and its application as targeted drug delivery vehicle. Bioresour. Technol. 2013, 145, 290–296. [Google Scholar] [CrossRef]
- Suwannasing, W.; Imai, T.; Kaewkannetra, P. Cost-effective defined medium for the production of polyhydroxyalkanoates using agricultural raw materials. Bioresour. Technol. 2015, 194, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, F.C.; Dias, M.L.; Castilho, L.R.; Freire, D.M. Characterization of poly (3-hydroxybutyrate) produced by Cupriavidus necator in solid-state fermentation. Bioresour. Technol. 2007, 98, 633–638. [Google Scholar] [CrossRef] [PubMed]
- Cavalheiro, J.M.; de Almeida, M.C.M.; Grandfils, C.; Da Fonseca, M. Poly (3-hydroxybutyrate) production by Cupriavidus necator using waste glycerol. Process. Biochem. 2009, 44, 509–515. [Google Scholar] [CrossRef]
- Xu, Y.; Wang, R.-H.; Koutinas, A.; Webb, C. Microbial biodegradable plastic production from a wheat-based biorefining strategy. Process. Biochem. 2010, 45, 153–163. [Google Scholar] [CrossRef]
- Jung, H.-R.; Choi, T.-R.; Han, Y.H.; Park, Y.-L.; Park, J.Y.; Song, H.-S.; Yang, S.-Y.; Bhatia, S.K.; Gurav, R.; Park, H. Production of blue-colored polyhydroxybutyrate (PHB) by one-pot production and coextraction of indigo and PHB from recombinant Escherichia coli. Dyes Pigment. 2020, 173, 107889. [Google Scholar] [CrossRef]
- Gu, J.J.; Zhou, Y.; Tong, Y.B.; Lu, J.J.; Ye, B.C. Efficient production and characterization of homopolymeric poly (3-hydroxyvalerate) produced by Bacillus strain PJC48. Biotechnol. Appl. Biochem. 2018, 65, 622–629. [Google Scholar] [CrossRef]
- Hiroe, A.; Watanabe, S.; Kobayashi, M.; Nomura, C.T.; Tsuge, T. Increased synthesis of poly (3-hydroxydodecanoate) by random mutagenesis of polyhydroxyalkanoate synthase. Appl. Microbiol. Biotechnol. 2018, 102, 7927–7934. [Google Scholar] [CrossRef]
- Chobchuenchom, W.; Tanadchangsaeng, N. Production of medium chain length polyhydroxyalkanoates by Pseudomonas putida ATCC47054 using glycerol and sodium octanoate as substrates. In Proceedings of the RSU International Research Conference, Riga, Latvia, 1–3 April 2019. [Google Scholar]
- Yamada, M.; Yukita, A.; Hanazumi, Y.; Yamahata, Y.; Moriya, H.; Miyazaki, M.; Yamashita, T.; Shimoi, H. Poly (3-hydroxybutyrate) production using mannitol as a sole carbon source by Burkholderia sp. AIU M5M02 isolated from a marine environment. Fish. Sci. 2018, 84, 405–412. [Google Scholar] [CrossRef]
- Kourmentza, C.; Costa, J.; Azevedo, Z.; Servin, C.; Grandfils, C.; De Freitas, V.; Reis, M. Burkholderia thailandensis as a microbial cell factory for the bioconversion of used cooking oil to polyhydroxyalkanoates and rhamnolipids. Bioresour. Technol. 2018, 247, 829–837. [Google Scholar] [CrossRef]
- Możejko-Ciesielska, J.; Kiewisz, R. Bacterial polyhydroxyalkanoates: Still fabulous? Microbiol. Res. 2016, 192, 271–282. [Google Scholar] [CrossRef]
- Li, Z.-J.; Shi, Z.-Y.; Jian, J.; Guo, Y.-Y.; Wu, Q.; Chen, G.-Q. Production of poly (3-hydroxybutyrate-co-4-hydroxybutyrate) from unrelated carbon sources by metabolically engineered Escherichia coli. Metab. Eng. 2010, 12, 352–359. [Google Scholar] [CrossRef] [PubMed]
- Koller, M. Polyhydroxyalkanoate biosynthesis at the edge of water activitiy-haloarchaea as biopolyester factories. Bioengineering 2019, 6, 34. [Google Scholar] [CrossRef] [PubMed]
- Aldor, I.S.; Kim, S.-W.; Prather, K.L.J.; Keasling, J.D. Metabolic engineering of a novel propionate-independent pathway for the production of poly (3-hydroxybutyrate-co-3-hydroxyvalerate) in recombinant Salmonella enterica serovar typhimurium. Appl. Environ. Microbiol. 2002, 68, 3848–3854. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Y.-Z.; Han, J.; Guo, J.-J.; Chen, G.-Q. Production of poly (3-hydroxybutyrate-co-3-hydroxyhexanoate) from gluconate and glucose by recombinant Aeromonas hydrophila and Pseudomonas putida. Biotechnol. Lett. 2005, 27, 1381–1386. [Google Scholar] [CrossRef] [PubMed]
- Matsusaki, H.; Abe, H.; Taguchi, K.; Fukui, T.; Doi, Y. Biosynthesis of poly (3-hydroxybutyrate-co-3-hydroxyalkanoates) by recombinant bacteria expressing the PHA synthase gene phaC1 from Pseudomonas sp. 61-3. Appl. Microbiol. Biotechnol. 2000, 53, 401–409. [Google Scholar] [CrossRef]
- Yim, K.S.; Lee, S.Y.; Chang, H.N. Synthesis of poly-(3-hydroxybutyrate-co-3-hydroxyvalerate) by recombinant Escherichia coli. Biotechnol. Bioeng. 1996, 49, 495–503. [Google Scholar] [CrossRef]
- Kichise, T.; Fukui, T.; Yoshida, Y.; Doi, Y. Biosynthesis of polyhydroxyalkanoates (PHA) by recombinant Ralstonia eutropha and effects of PHA synthase activity on in vivo PHA biosynthesis. Int. J. Biol. Macromol. 1999, 25, 69–77. [Google Scholar] [CrossRef]
- Tsuge, T.; Yano, K.; Imazu, S.i.; Numata, K.; Kikkawa, Y.; Abe, H.; Taguchi, S.; Doi, Y. Biosynthesis of polyhydroxyalkanoate (PHA) copolymer from fructose using wild-type and laboratory-evolved PHA synthases. Macromol. Biosci. 2005, 5, 112–117. [Google Scholar] [CrossRef]
- Lee, S.H.; Oh, D.H.; Ahn, W.S.; Lee, Y.; Choi, J.i.; Lee, S.Y. Production of poly (3-hydroxybutyrate-co-3-hydroxyhexanoate) by high-cell-density cultivation of Aeromonas hydrophila. Biotechnol. Bioeng. 2000, 67, 240–244. [Google Scholar] [CrossRef]
- Han, J.; Qiu, Y.-Z.; Liu, D.-C.; Chen, G.-Q. Engineered Aeromonas hydrophila for enhanced production of poly (3-hydroxybutyrate-co-3-hydroxyhexanoate) with alterable monomers composition. FEMS Microbiol. Lett. 2004, 239, 195–201. [Google Scholar] [CrossRef]
- Kim, J.S.; Lee, B.H.; Kim, B.S. Production of poly (3-hydroxybutyrate-co-4-hydroxybutyrate) by Ralstonia eutropha. Biochem. Eng. J. 2005, 23, 169–174. [Google Scholar] [CrossRef]
- Rao, U.; Sridhar, R.; Sehgal, P. Biosynthesis and biocompatibility of poly (3-hydroxybutyrate-co-4-hydroxybutyrate) produced by Cupriavidus necator from spent palm oil. Biochem. Eng. J. 2010, 49, 13–20. [Google Scholar] [CrossRef]
- Zhao, W.; Chen, G.-Q. Production and characterization of terpolyester poly (3-hydroxybutyrate-co-3-hydroxyvalerate-co-3-hydroxyhexanoate) by recombinant Aeromonas hydrophila 4AK4 harboring genes phaAB. Process. Biochem. 2007, 42, 1342–1347. [Google Scholar] [CrossRef]
- Lee, W.-H.; Azizan, M.N.; Sudesh, K. Effects of culture conditions on the composition of poly (3-hydroxybutyrate-co-4-hydroxybutyrate) synthesized by Comamonas acidovorans. Polym. Degrad. Stab. 2004, 84, 129–134. [Google Scholar] [CrossRef]
- Lee, W.-H.; Loo, C.-Y.; Nomura, C.T.; Sudesh, K. Biosynthesis of polyhydroxyalkanoate copolymers from mixtures of plant oils and 3-hydroxyvalerate precursors. Bioresour. Technol. 2008, 99, 6844–6851. [Google Scholar] [CrossRef]
- Meng, D.-C.; Shi, Z.-Y.; Wu, L.-P.; Zhou, Q.; Wu, Q.; Chen, J.-C.; Chen, G.-Q. Production and characterization of poly (3-hydroxypropionate-co-4-hydroxybutyrate) with fully controllable structures by recombinant Escherichia coli containing an engineered pathway. Metab. Eng. 2012, 14, 317–324. [Google Scholar] [CrossRef]
- Reddy, S.V.; Thirumala, M.; Mahmood, S. A novel Bacillus sp. accumulating poly (3-hydroxybutyrate-co-3-hydroxyvalerate) from a single carbon substrate. J. Ind. Microbiol. Biotechnol. 2009, 36, 837–843. [Google Scholar] [CrossRef] [PubMed]
- Masood, F.; Hasan, F.; Ahmed, S.; Hameed, A. Biosynthesis and characterization of poly (3-hydroxybutyrate-co-3-hydroxyvalerate) from Bacillus cereus FA11 isolated from TNT-contaminated soil. Ann. Microbiol. 2012, 62, 1377–1384. [Google Scholar] [CrossRef]
- Wong, Y.-M.; Brigham, C.J.; Rha, C.; Sinskey, A.J.; Sudesh, K. Biosynthesis and characterization of polyhydroxyalkanoate containing high 3-hydroxyhexanoate monomer fraction from crude palm kernel oil by recombinant Cupriavidus necator. Bioresour. Technol. 2012, 121, 320–327. [Google Scholar] [CrossRef]
- Tsuge, T.; Yamamoto, T.; Yano, K.; Abe, H.; Doi, Y.; Taguchi, S. Evaluating the ability of polyhydroxyalkanoate synthase mutants to produce P (3HB-co-3HA) from soybean oil. Macromol. Biosci. 2009, 9, 71–78. [Google Scholar] [CrossRef]
- Al-Kaddo, K.B.; Mohamad, F.; Murugan, P.; Tan, J.S.; Sudesh, K.; Samian, M.R. Production of P (3HB-co-4HB) copolymer with high 4HB molar fraction by Burkholderia contaminans Kad1 PHA synthase. Biochem. Eng. J. 2020, 153, 107394. [Google Scholar] [CrossRef]
- Norhafini, H.; Huong, K.-H.; Amirul, A. High PHA density fed-batch cultivation strategies for 4HB-rich P (3HB-co-4HB) copolymer production by transformant Cupriavidus malaysiensis USMAA1020. Int. J. Biol. Macromol. 2019, 125, 1024–1032. [Google Scholar] [CrossRef]
- Bhatia, S.K.; Gurav, R.; Choi, T.-R.; Jung, H.-R.; Yang, S.-Y.; Song, H.-S.; Jeon, J.-M.; Kim, J.-S.; Lee, Y.-K.; Yang, Y.-H. Poly (3-hydroxybutyrate-co-3-hydroxyhexanoate) production from engineered Ralstonia eutropha using synthetic and anaerobically digested food waste derived volatile fatty acids. Int. J. Biol. Macromol. 2019, 133, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Jeon, J.-M.; Brigham, C.J.; Kim, Y.-H.; Kim, H.-J.; Yi, D.-H.; Kim, H.; Rha, C.; Sinskey, A.J.; Yang, Y.-H. Biosynthesis of poly (3-hydroxybutyrate-co-3-hydroxyhexanoate)(P (HB-co-HHx)) from butyrate using engineered Ralstonia eutropha. Appl. Microbiol. Biotechnol. 2014, 98, 5461–5469. [Google Scholar] [CrossRef]
- Jeon, J.-M.; Kim, H.-J.; Bhatia, S.K.; Sung, C.; Seo, H.-M.; Kim, J.-H.; Park, H.-Y.; Lee, D.; Brigham, C.J.; Yang, Y.-H. Application of acetyl-CoA acetyltransferase (AtoAD) in Escherichia coli to increase 3-hydroxyvalerate fraction in poly (3-hydroxybutyrate-co-3-hydroxyvalerate). Bioprocess. Biosyst. Eng. 2017, 40, 781–789. [Google Scholar] [CrossRef] [PubMed]
- Bhatia, S.K.; Kim, J.-H.; Kim, M.-S.; Kim, J.; Hong, J.W.; Hong, Y.G.; Kim, H.-J.; Jeon, J.-M.; Kim, S.-H.; Ahn, J. Production of (3-hydroxybutyrate-co-3-hydroxyhexanoate) copolymer from coffee waste oil using engineered Ralstonia eutropha. Bioprocess. Biosyst. Eng. 2018, 41, 229–235. [Google Scholar] [CrossRef]
- Bhatia, S.K.; Gurav, R.; Choi, T.-R.; Jung, H.-R.; Yang, S.-Y.; Moon, Y.-M.; Song, H.-S.; Jeon, J.-M.; Choi, K.-Y.; Yang, Y.-H. Bioconversion of plant biomass hydrolysate into bioplastic (polyhydroxyalkanoates) using Ralstonia eutropha 5119. Bioresour. Technol. 2019, 271, 306–315. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Kurita, S.; Orita, I.; Nakamura, S.; Fukui, T. Modification of acetoacetyl-CoA reduction step in Ralstonia eutropha for biosynthesis of poly (3-hydroxybutyrate-co-3-hydroxyhexanoate) from structurally unrelated compounds. Microb. Cell Fact. 2019, 18, 147. [Google Scholar] [CrossRef]
- Sudo, M.; Hori, C.; Ooi, T.; Mizuno, S.; Tsuge, T.; Matsumoto, K.i. Synergy of valine and threonine supplementation on poly (2-hydroxybutyrate-block-3-hydroxybutyrate) synthesis in engineered Escherichia coli expressing chimeric polyhydroxyalkanoate synthase. J. Biosci. Bioeng. 2020, 129, 302–306. [Google Scholar] [CrossRef]
- Steinbüchel, A.; Hofrichter, M.; Koyama, T.; Vandamme, E.; De Baets, S. Biopolymers: Biology, Chemistry, Biotechnology, Applications: Polysaccharides I: Polysaccharides from Prokaryotes; Wiley-VCH: Weinheim, Germany, 2001. [Google Scholar]
- McChalicher, C.W.; Srienc, F. Investigating the structure–property relationship of bacterial PHA block copolymers. J. Biotechnol. 2007, 132, 296–302. [Google Scholar] [CrossRef]
- Pederson, E.N.; McChalicher, C.W.; Srienc, F. Bacterial synthesis of PHA block copolymers. Biomacromolecules 2006, 7, 1904–1911. [Google Scholar] [CrossRef]
- Li, S.Y.; Dong, C.L.; Wang, S.Y.; Ye, H.M.; Chen, G.-Q. Microbial production of polyhydroxyalkanoate block copolymer by recombinant Pseudomonas putida. Appl. Microbiol. Biotechnol. 2011, 90, 659–669. [Google Scholar] [CrossRef]
- Tripathi, L.; Wu, L.-P.; Chen, J.; Chen, G.-Q. Synthesis of Diblock copolymer poly-3-hydroxybutyrate-block-poly-3-hydroxyhexanoate [PHB-b-PHHx] by a β-oxidation weakened Pseudomonas putida KT2442. Microb. Cell Fact. 2012, 11, 44. [Google Scholar] [CrossRef] [PubMed]
- Brännström, H.; Kumar, H.; Alén, R. Current and potential biofuel production from plant oils. BioEnergy Res. 2018, 11, 592–613. [Google Scholar] [CrossRef]
- Chhetri, A.B.; Tango, M.S.; Budge, S.M.; Watts, K.C.; Islam, M.R. Non-edible plant oils as new sources for biodiesel production. Int. J. Mol. Sci. 2008, 9, 169–180. [Google Scholar] [CrossRef] [PubMed]
- Demirbas, A.; Bafail, A.; Ahmad, W.; Sheikh, M. Biodiesel production from non-edible plant oils. Energy Explor. Exploit. 2016, 34, 290–318. [Google Scholar] [CrossRef]
- Zhang, X.; Luo, R.; Wang, Z.; Deng, Y.; Chen, G.-Q. Application of (R)-3-hydroxyalkanoate methyl esters derived from microbial polyhydroxyalkanoates as novel biofuels. Biomacromolecules 2009, 10, 707–711. [Google Scholar] [CrossRef]
- Choonut, A.; Yunu, T.; Pichid, N.; Sangkharak, K. The optimization conditions of polyhydroxybutyrate methyl ester from polyhydroxybutyrate via acid-catalyst. Energy Proc. 2017, 138, 435–440. [Google Scholar] [CrossRef]
- Sangkharak, K.; Pichid, N.; Yunu, T.; Srinak, K.; Sornnum, S.; Prasertsan, P. Biofuel production, characterization and degradation of 3-hydroxybutyate methyl ester from polyhydroxybutyrate. Chiang Mai J. Sci. 2016, 43, 808–817. [Google Scholar]
- Keunun, P.; Rakkarn, T.; Yunu, T.; Paichid, N.; Prasertsan, P.; Sangkharak, K. The production of polyhydroxybutyrate by two-step fermentation and the application of polyhydroxybutyrate as a novel substrate for a biolubricant. J. Polym. Environ. 2018, 26, 2459–2466. [Google Scholar] [CrossRef]
- Okwundu, O.S.; El-Shazly, A.H.; Elkady, M. Comparative effect of reaction time on biodiesel production from low free fatty acid beef tallow: A definition of product yield. SN Appl. Sci. 2019, 1, 140. [Google Scholar] [CrossRef]
- Weerachanchai, P.; Tangsathitkulchai, C.; Tangsathitkulchai, M. Effect of reaction conditions on the catalytic esterification of bio-oil. Korean J. Chem. Eng. 2012, 29, 182–189. [Google Scholar] [CrossRef]
- Farag, H.; El-Maghraby, A.; Taha, N.A. Optimization of factors affecting esterification of mixed oil with high percentage of free fatty acid. Fuel Process. Technol. 2011, 92, 507–510. [Google Scholar] [CrossRef]
- Villegas Calvo, M.; Colombo, B.; Corno, L.; Eisele, G.; Cosentino, C.; Papa, G.; Scaglia, B.; Pilu, R.; Simmons, B.; Adani, F. Bioconversion of giant cane for integrated production of biohydrogen, carboxylic acids, and polyhydroxyalkanoates (PHAs) in a multistage biorefinery approach. ACS Sustain. Chem. Eng. 2018, 6, 15361–15373. [Google Scholar] [CrossRef]
- De Jong, E.; Higson, A.; Walsh, P.; Wellisch, M. Bio-Based Chemicals Value Added Products from Biorefineries; IEA Bioenergy: Utrecht, The Netherlands, 2012; Volume 34. [Google Scholar]
- Snell, K.D.; Peoples, O.P. PHA bioplastic: A value-added coproduct for biomass biorefineries. Biofuels Bioprod. Biorefin. Innovat. Sustain. Econ. 2009, 3, 456–467. [Google Scholar] [CrossRef]
- Budzianowski, W.M. High-value low-volume bioproducts coupled to bioenergies with potential to enhance business development of sustainable biorefineries. Renew. Sustain. Energy Rev. 2017, 70, 793–804. [Google Scholar] [CrossRef]
- Jiang, G.; Hill, D.J.; Kowalczuk, M.; Johnston, B.; Adamus, G.; Irorere, V.; Radecka, I. Carbon sources for polyhydroxyalkanoates and an integrated biorefinery. Int. J. Mol. Sci. 2016, 17, 1157. [Google Scholar] [CrossRef]
- Liu, Z.-H.; Shinde, S.; Xie, S.; Hao, N.; Lin, F.; Li, M.; Yoo, C.G.; Ragauskas, A.J.; Yuan, J.S. Cooperative valorization of lignin and residual sugar to polyhydroxyalkanoate (PHA) for enhanced yield and carbon utilization in biorefineries. Sustain. Energy Fuels 2019, 3, 2024–2037. [Google Scholar] [CrossRef]
- Raza, Z.A.; Riaz, S.; Banat, I.M. Polyhydroxyalkanoates: Properties and chemical modification approaches for their functionalization. Biotechnol. Progr. 2018, 34, 29–41. [Google Scholar] [CrossRef]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).