
Entrapment of the Fastest Known Carbonic Anhydrase with Biomimetic Silica and Its Application for CO2 Sequestration

Abstract
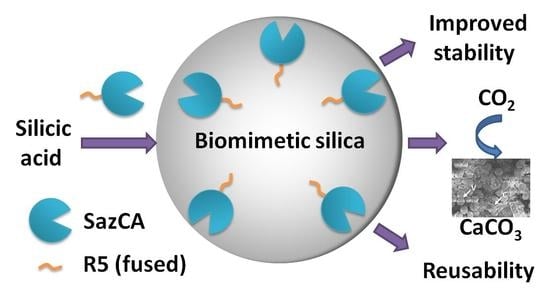
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Vector Construction
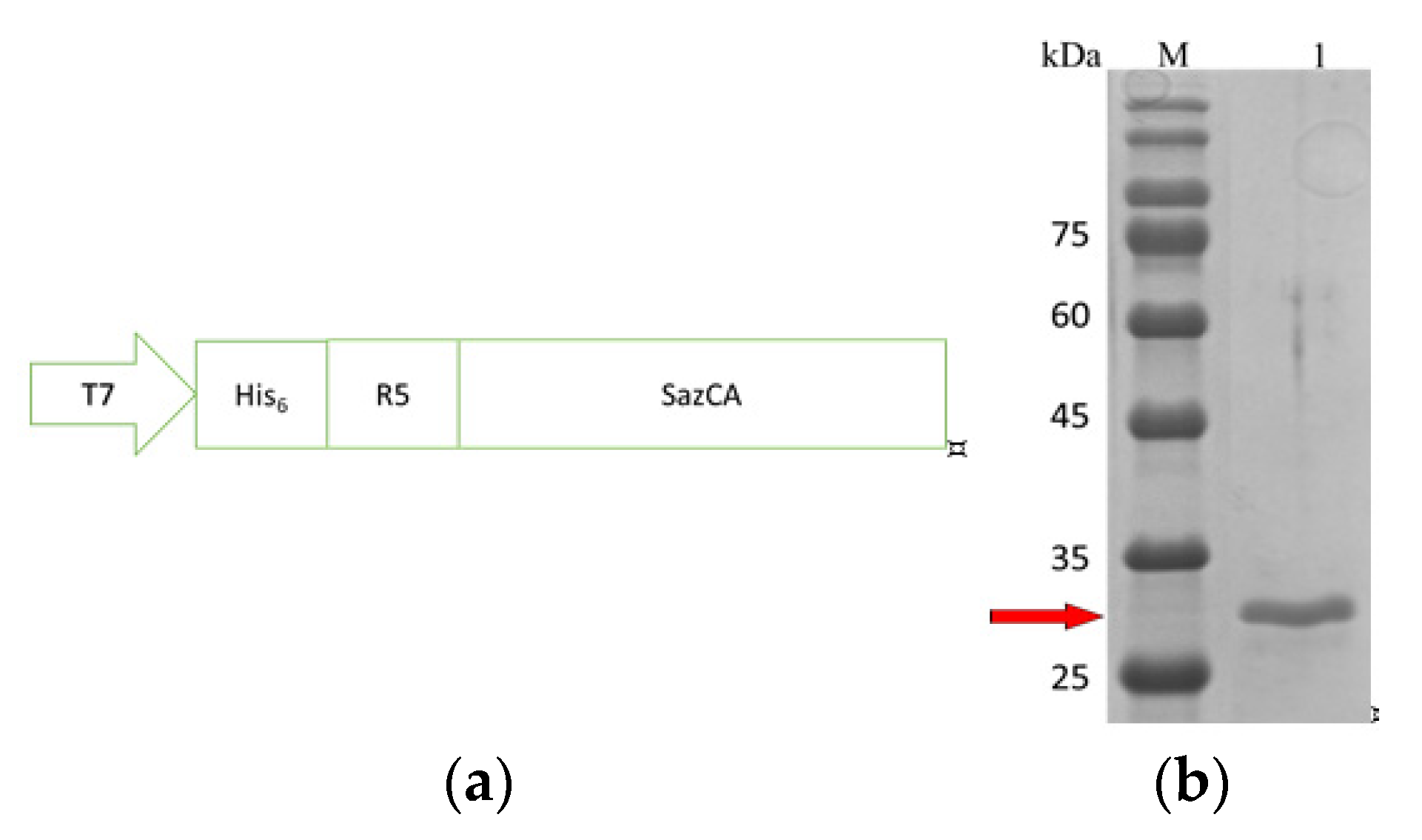
2.2. Expression and Purification of R5-SazCA
2.3. Hydratase Activity Assay
2.4. Esterase Activity Assay
2.5. Entrapment of R5-SazCA in Silica Nanoparticles
2.6. CO2 Sequestration
3. Results and Discussion
3.1. Expression and Purification of R5-SazCA
3.2. Encasuplation of R5-SazCA
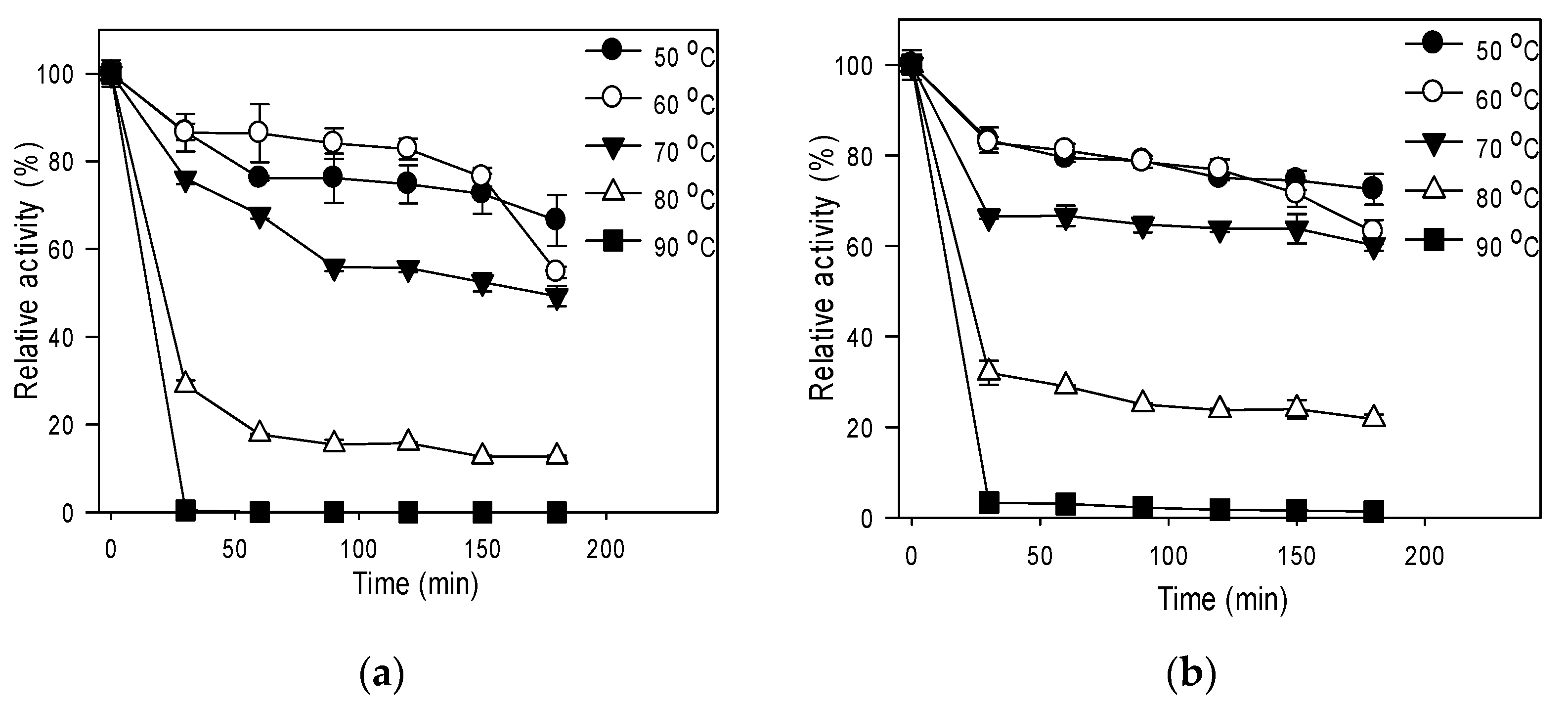
3.3. Effect of Temperature on Activity and Stability
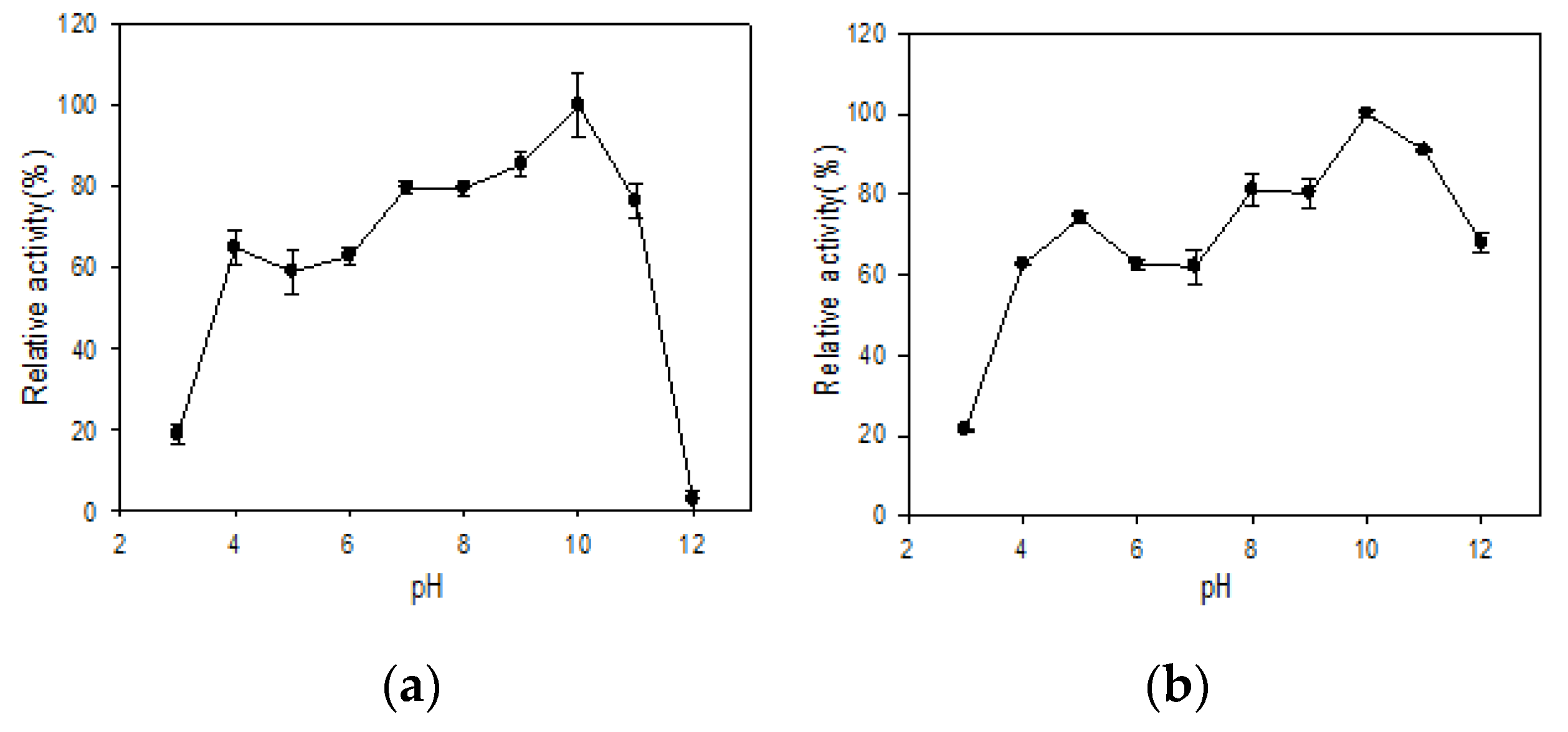
3.4. Effect of pH on Stability
3.5. Cation and Salinity Tolerance
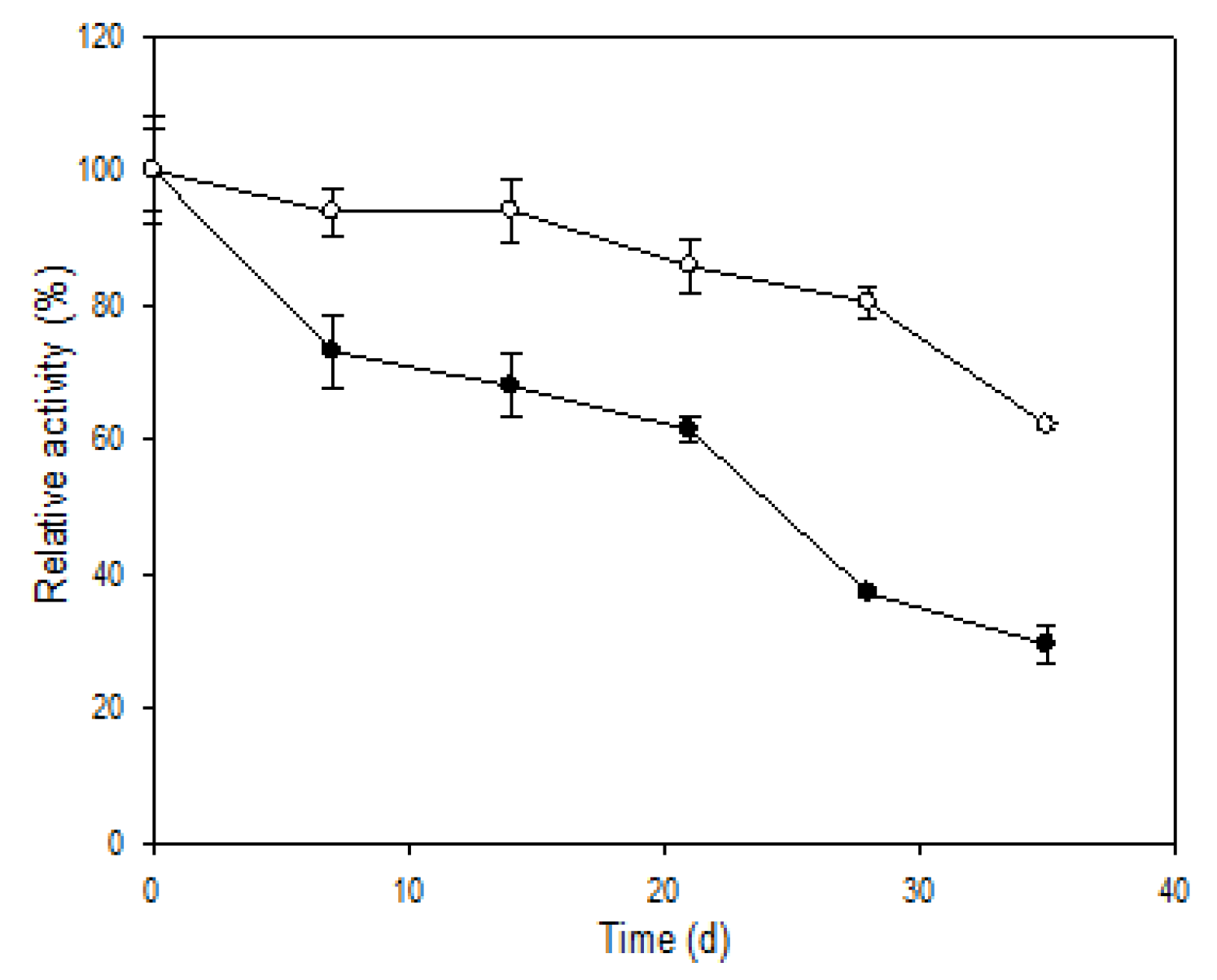
3.6. Storage Stability and Reusability
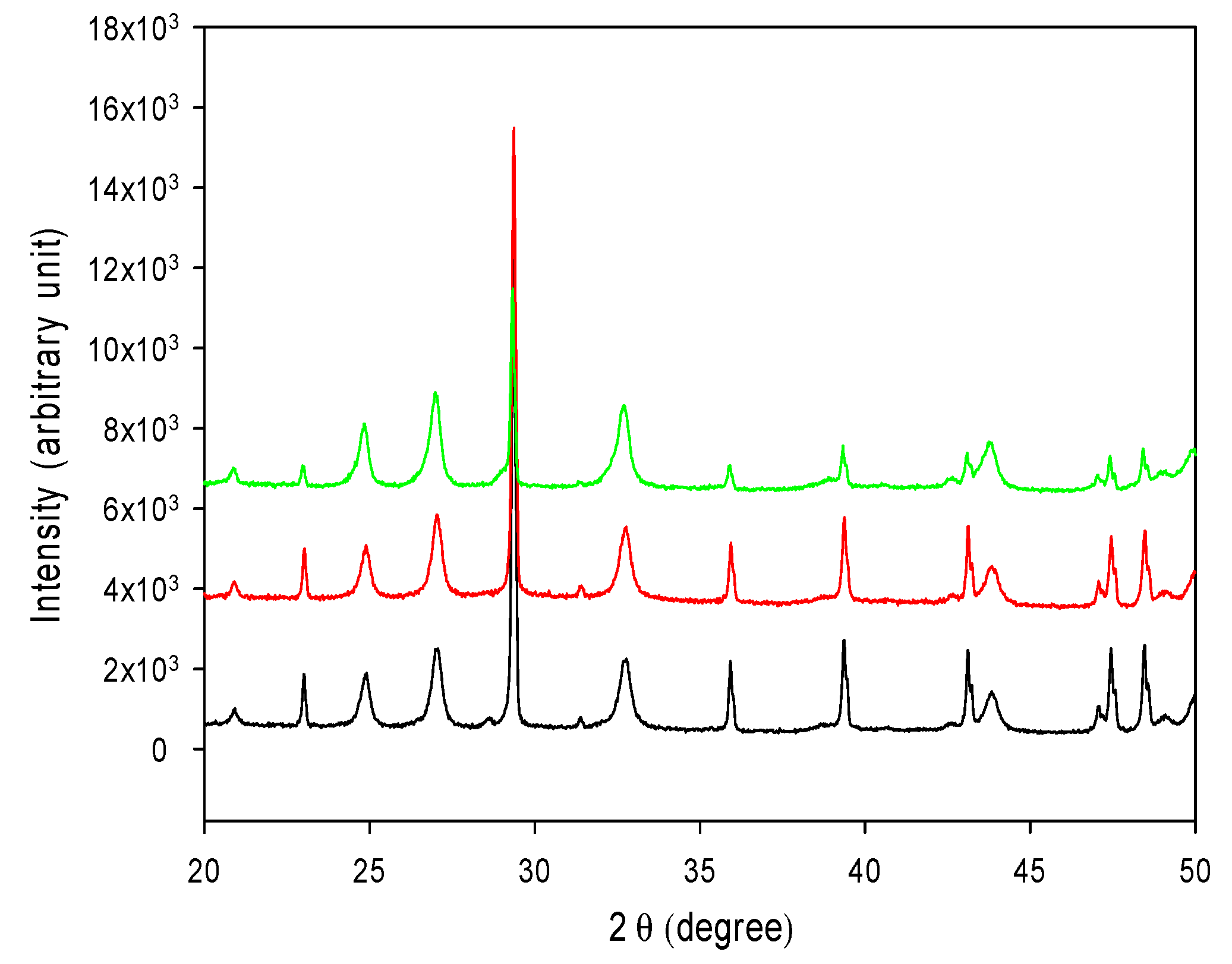
3.7. CO2 Sequestration in CaCO3
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Crippa, M.; Guizzardi, D.; Muntean, M.; Schaaf, E.; Solazzo, E.; Monforti-Ferrario, F.; Olivier, J.G.J.; Vignati, E. Fossil CO2 Emissions of All World Countries—2020 Report, EUR 30358 EN; European Commission: Luxembourg, 2020; pp. 1–244. [Google Scholar]
- D’Alessandro, D.M.; Smit, B.; Long, J.R. Carbon dioxide capture: Prospects for new materials. Angew. Chem. Int. Ed. 2010, 49, 6058–6082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bose, H.; Satyanarayana, T. Microbial carbonic anhydrases in biomimetic carbon sequestration for mitigating global warming: Prospects and perspectives. Front. Microbiol. 2017, 8, 1615. [Google Scholar] [CrossRef]
- Smith, K.S.; Ferry, J.G. Prokaryotic carbonic anhydrases. FEMS Microbiol. Rev. 2000, 24, 335–366. [Google Scholar] [CrossRef]
- Dreybrodt, W.; Lauckner, J.; Zaihua, L.; Svensson, U.; Buhmann, D. The kinetics of the reaction CO2 + H2O → H+ + HCO3− as one of the rate limiting steps for the dissolution of calcite in the system H2O-CO2-CaCO3. Geochim. Cosmochim. Acta 1996, 60, 3375–3381. [Google Scholar] [CrossRef]
- Di Fiore, A.; Alterio, V.; Monti, S.M.; De Simone, G.; D’Ambrosio, K. Thermostable carbonic anhydrases in biotechnological applications. Int. J. Mol. Sci. 2015, 16, 15456–15480. [Google Scholar] [CrossRef] [PubMed]
- Capasso, C.; De Luca, V.; Carginale, V.; Cannio, R.; Rossi, M. Biochemical properties of a novel and highly thermostable bacterial alpha-carbonic anhydrase from Sulfurihydrogenibium yellowstonense YO3AOP1. J. Enzyme. Inhib. Med. Chem. 2012, 27, 892–897. [Google Scholar] [CrossRef] [Green Version]
- Luca, V.D.; Vullo, D.; Scozzafava, A.; Carginale, V.; Rossi, M.; Supuran, C.T.; Capasso, C. An α-carbonic anhydrase from the thermophilic bacterium Sulphurihydrogenibium azorense is the fastest enzyme known for the CO2 hydration reaction. Bioorg. Med. Chem. 2013, 21, 1465–1469. [Google Scholar] [CrossRef]
- Vullo, D.; De Luca, V.; Scozzafava, A.; Carginale, V.; Rossi, M.; Supuran, C.T.; Capasso, C. Anion inhibition studies of the fastest carbonic anhydrase (CA) known, the extremo-CA from the bacterium Sulfurihydrogenibium azorense. Bioorg. Med. Chem. Lett. 2012, 22, 7142–7145. [Google Scholar] [CrossRef] [Green Version]
- Yoshimoto, M.; Walde, P. Immobilized carbonic anhydrase: Preparation, characteristics and biotechnological applications. World J. Microbiol. Biotechnol. 2018, 34, 151. [Google Scholar] [CrossRef]
- Sumper, M.; Kröger, N. Silica formation in diatoms: The function of long chain polyamines and silaffins. J. Mater. Chem. 2004, 14, 2059–2065. [Google Scholar] [CrossRef]
- Knecht, M.R.; Wright, D.W. Functional analysis of the biomimetic silica precipitating activity of the R5 peptide from Cylindrotheca fusiformis. Chem. Comm. 2003, 24, 3038–3039. [Google Scholar] [CrossRef]
- Chien, L.-J.; Lee, C.-K. Biosilicification of dual-fusion enzyme immobilized on magnetic nanoparticle. Biotechnol. Bioeng. 2007, 100, 223–230. [Google Scholar] [CrossRef] [PubMed]
- Luckarift, H.R.; Spain, J.C.; Naik, R.R.; Stone, M.O. Enzyme immobilization in a biomimetic silica support. Nat. Biotech. 2004, 22, 211–213. [Google Scholar] [CrossRef] [PubMed]
- Betancor, L.; Luckarift, H.R. Bioinspired enzyme encapsulation for biocatalysis. Trends Biotechnol. 2008, 26, 566–572. [Google Scholar] [CrossRef]
- Lutz, H.; Jaeger, V.; Schmüser, L.; Bonn, M.; Pfaendtner, J.; Weidner, T. The structure of the diatom silaffin peptide R5 within freestanding two-dimensional biosilica sheets. Angew Chem. Int. Ed. 2017, 56, 8277–8280. [Google Scholar] [CrossRef]
- Senior, L.; Crump, M.P.; Williams, C.; Booth, P.J.; Mann, S.; Perriman, A.W.; Curnow, P. Structure and function of the silicifying peptide R5. J. Mater. Chem. B 2015, 3, 2607–2614. [Google Scholar] [CrossRef] [Green Version]
- Jo, B.H.; Seo, J.H.; Yang, Y.J.; Baek, K.; Choi, Y.S.; Pack, S.P.; Oh, S.H.; Cha, H.J. Bioinspired silica nanocomposite with autoencapsulated carbonic anhydrase as a robust biocatalyst for CO2 sequestration. ACS Catal. 2014, 4, 4332–4340. [Google Scholar] [CrossRef]
- Expasy. Available online: Web.expasy.org/compute_pi/ (accessed on 23 March 2021).
- De Simone, G.; Monti, S.M.; Alterio, V.; Buonanno, M.; De Luca, V.; Rossi, M.; Carginale, V.; Supuran, C.T.; Capasso, C.; Di Fiore, A. Crystal structure of the most catalytically effective carbonic anhydrase enzyme known, SazCA from the thermophilic bacterium Sulfurihydrogenibium azorense. Bioorg. Med. Chem. Lett. 2015, 25, 2002–2006. [Google Scholar] [CrossRef]
- Forsyth, C.; Yip, T.W.; Patwardhan, S.V. CO2 sequestration by enzyme immobilized onto bioinspired silica. Chem. Commun. 2013, 49, 3191–3193. [Google Scholar] [CrossRef]
- Chien, L.-J.; Sureshkumar, M.; Hsieh, H.-H.; Wang, J.-L. Biosequestration of carbon dioxide using a silicified carbonic anhydrase catalyst. Biotechnol. Bioprocess Eng. 2013, 18, 567–574. [Google Scholar] [CrossRef]
- Hsu, K.-P.; Tan, S.-I.; Chiu, C.-Y.; Chang, Y.-K.; Ng, I.-S. ARduino-pH Tracker and screening platform for characterization of recombinant carbonic anhydrase in Escherichia coli. Biotechnol. Prog. 2019, 35, e2834. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Zhao, X.; Li, P.; Huang, H. Impact of Zn, Cu, and Fe on the activity of carbonic anhydrase of erythrocytes in ducks. Biol. Trace Elem. Res. 2007, 118, 227–232. [Google Scholar] [CrossRef]
- Kumari, M.; Lee, J.; Lee, D.W.; Hwang, I. High-level production in a plant system of a thermostable carbonic anhydrase and its immobilization on microcrystalline cellulose beads for CO2 capture. Plant Cell Rep. 2020, 39, 1317–1329. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hsieh, C.-J.; Cheng, J.-C.; Hu, C.-J.; Yu, C.-Y. Entrapment of the Fastest Known Carbonic Anhydrase with Biomimetic Silica and Its Application for CO2 Sequestration. Polymers 2021, 13, 2452. https://doi.org/10.3390/polym13152452
Hsieh C-J, Cheng J-C, Hu C-J, Yu C-Y. Entrapment of the Fastest Known Carbonic Anhydrase with Biomimetic Silica and Its Application for CO2 Sequestration. Polymers. 2021; 13(15):2452. https://doi.org/10.3390/polym13152452
Chicago/Turabian StyleHsieh, Chia-Jung, Ju-Chuan Cheng, Chia-Jung Hu, and Chi-Yang Yu. 2021. "Entrapment of the Fastest Known Carbonic Anhydrase with Biomimetic Silica and Its Application for CO2 Sequestration" Polymers 13, no. 15: 2452. https://doi.org/10.3390/polym13152452
APA StyleHsieh, C.-J., Cheng, J.-C., Hu, C.-J., & Yu, C.-Y. (2021). Entrapment of the Fastest Known Carbonic Anhydrase with Biomimetic Silica and Its Application for CO2 Sequestration. Polymers, 13(15), 2452. https://doi.org/10.3390/polym13152452