3.1. Braid Configuration under External Force
Minimizing the energy Equation (
1), we obtain a phase diagram, which is shown in
Figure 2 spanned by the applied torque
and the dimensionless parameter
that quantifies the propensity of the braided bundle to form a kink. The kink angle is given by
. Depending on
and the applied torque, the braid may or may not cause a localized bend of the filament or kink, as can be seen in the figure where, at large applied torques, we predict straight, braided bundles, as is intuitively reasonable. If the braids move outward along the bundle, or if we decrease the applied force, the torque acting on the kinks is reduced. In the first case, this is due to the decreasing moment arm produced by the kinked bundle, while in the second case, it is simply due to the reduced applied force.
The nonmonotonic behavior of the phase boundary is perhaps the most striking feature of the phase diagram, which was also discussed in Ref. [
6]. If
is small, the binding energy of the linkers is also small, allowing the braid to extend along the bundle, thereby reducing its effective bending modulus. As
is increased, the braids become shorter and stiffer. When
is sufficiently large, however, the braid kinks even in the absence of an applied force in order to minimize the energy of the bundle. Because of this effect, the large-
limit also exhibits kinks. We note that this large-
limit occurs around
, which we believe is obtainable in some biopolymer systems, including condensed DNA [
9,
10], cross-linked intermediate filaments [
11,
12] and perhaps for collagen [
5,
13,
14,
15]. We should note that we introduced the torque with the opposite sign from that used here in Ref. [
6], i.e., extension is positive, and compression is negative.
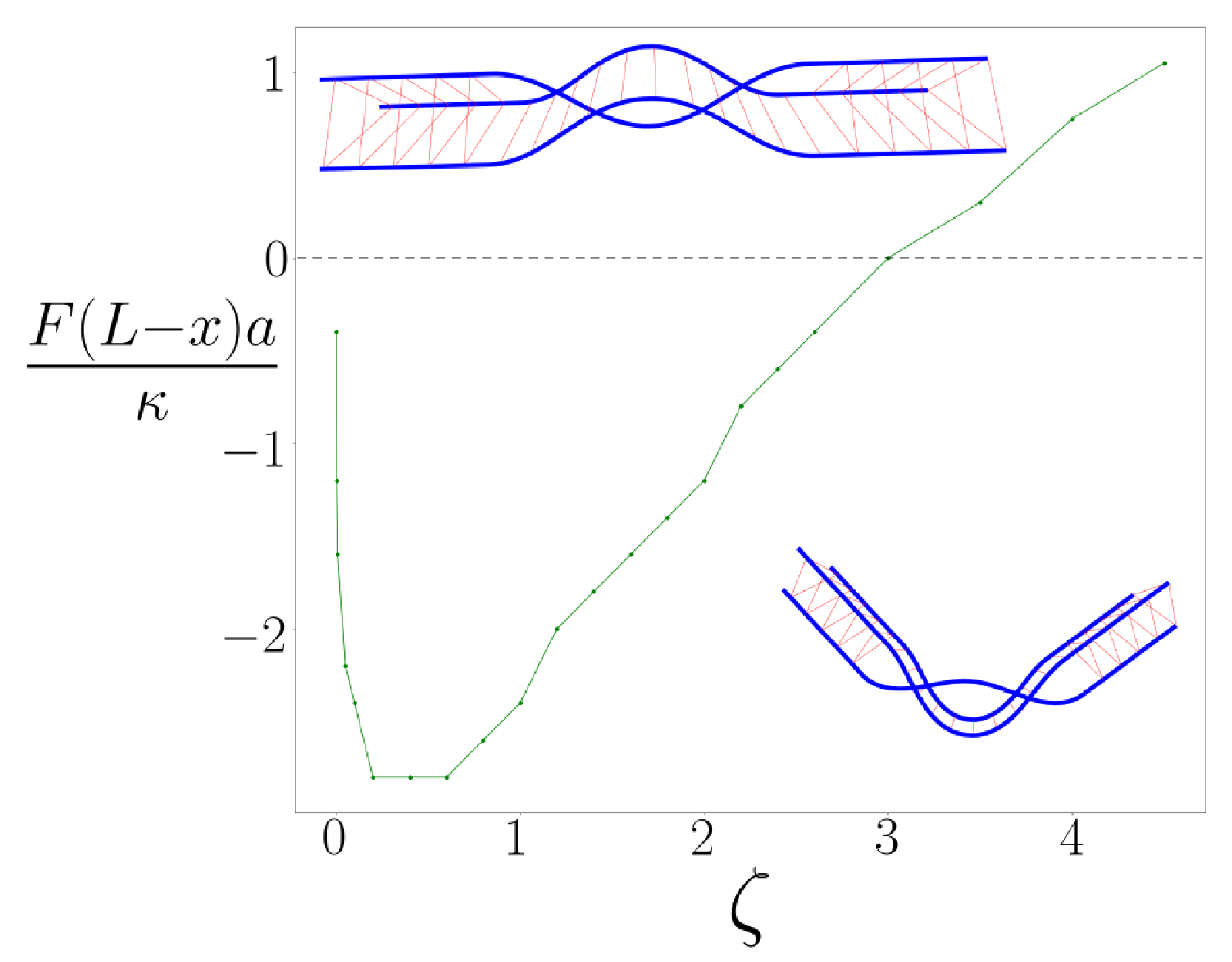
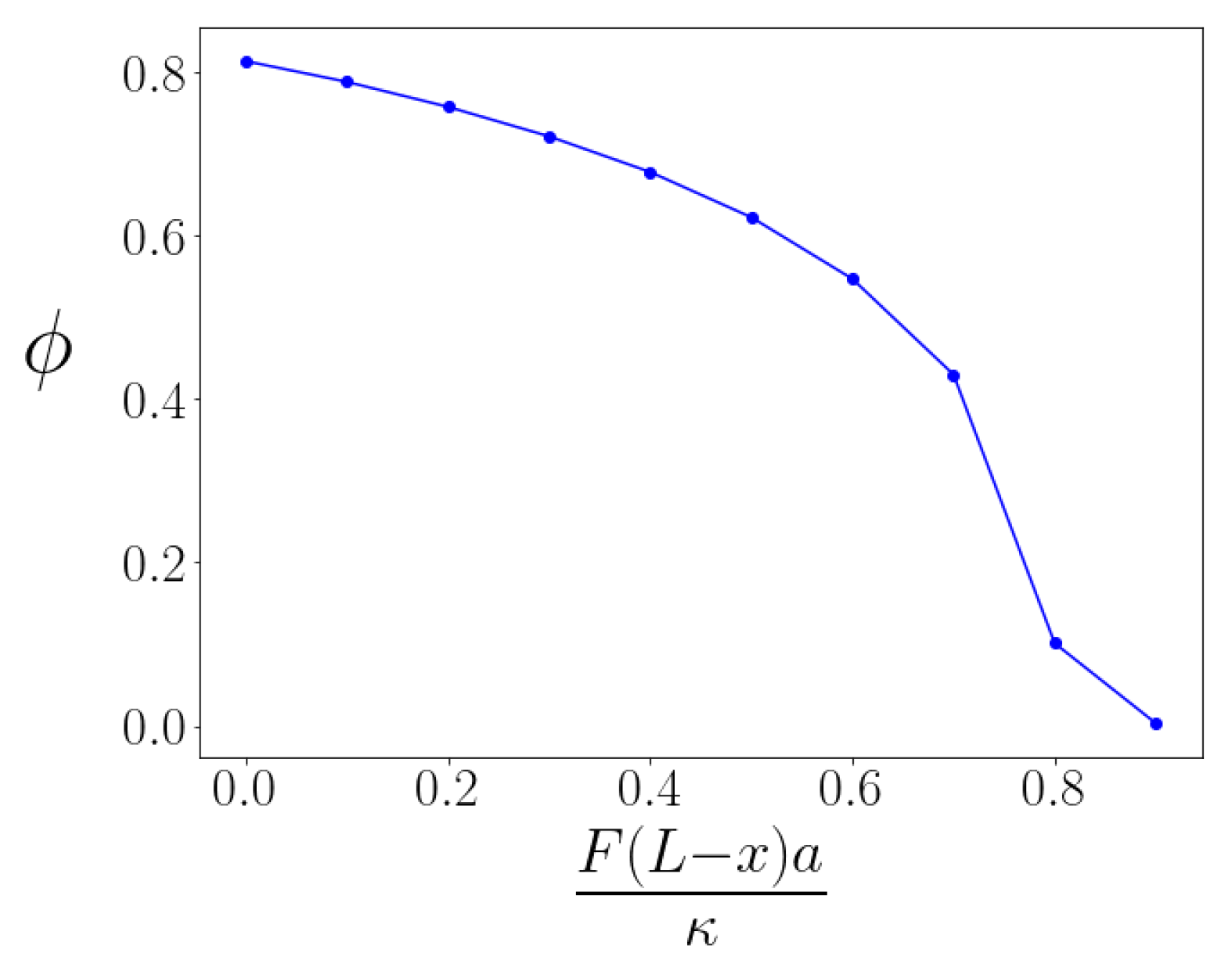
When we consider the case of sufficiently large values of the
parameter, so that kinks exist even under non-zero tensile force, we may then investigate how the kink angle changes in response to that loading by determining the energy-minimized configuration of each kinked braid. As expected, the kink is straightened—the kink angle
decreases—under tension. The dependence of the kink angle
versus applied tension is shown in
Figure 3 for a fixed
. We observe that the angle versus torque of the kinked bundle is nonlinear; the torque response of the bundle is non-Hookean. We understand this effect to occur because the structure of the energy-minimizing braid itself changes with applied torque so the braided bundle does not bend simply like an effectively thicker filament.
We will later see that the force-induced straightening of the kinks allows for the nonmonotonic behavior of the bundle’s extension versus time under a constant tensile loading. This is discussed in more detail in
Section 3.4, but first, in the following section, we assume the torque to change weakly, so we can assume the kink angle to be constant, which sufficiently simplifies the study of the braid/anti-braid pair production for our analysis.
3.2. The Piece-Wise Linear Defect Potential and the Defect Distribution
We showed that the production of braid/anti-braid pairs in a compressed bundle can be mapped onto the Kramers escape problem in one dimension, using a single reaction coordinate [
6,
16]. The two defects must be first produced together in the form of defect/anti-defect pairs, which requires energy
. The formation of one defect introduces a length mismatch between the filaments involved that is then compensated by the second anti-defect. As a result, defect pair production entails only local rearrangements of cross linkers. During this point in the thermally-activated production of the defect pair, we may take the single reaction coordinate to represent the total length “exchanged” between defects. Once the defects separate so that a region of cross-linked bundle appears between them, the defects can no longer exchange length but they may separate along the bundle by repetitive motion. Most importantly, since the defects generate localized bends or kinks under external loading, the motion of the defects changes the end-to-end distance of the bundle under load. By separating the braids, the energy of the system decreases in response to the applied tension and increases in response to the applied compression. During this separation (under external loading) the distance between the defects plays the role of the reaction coordinate.
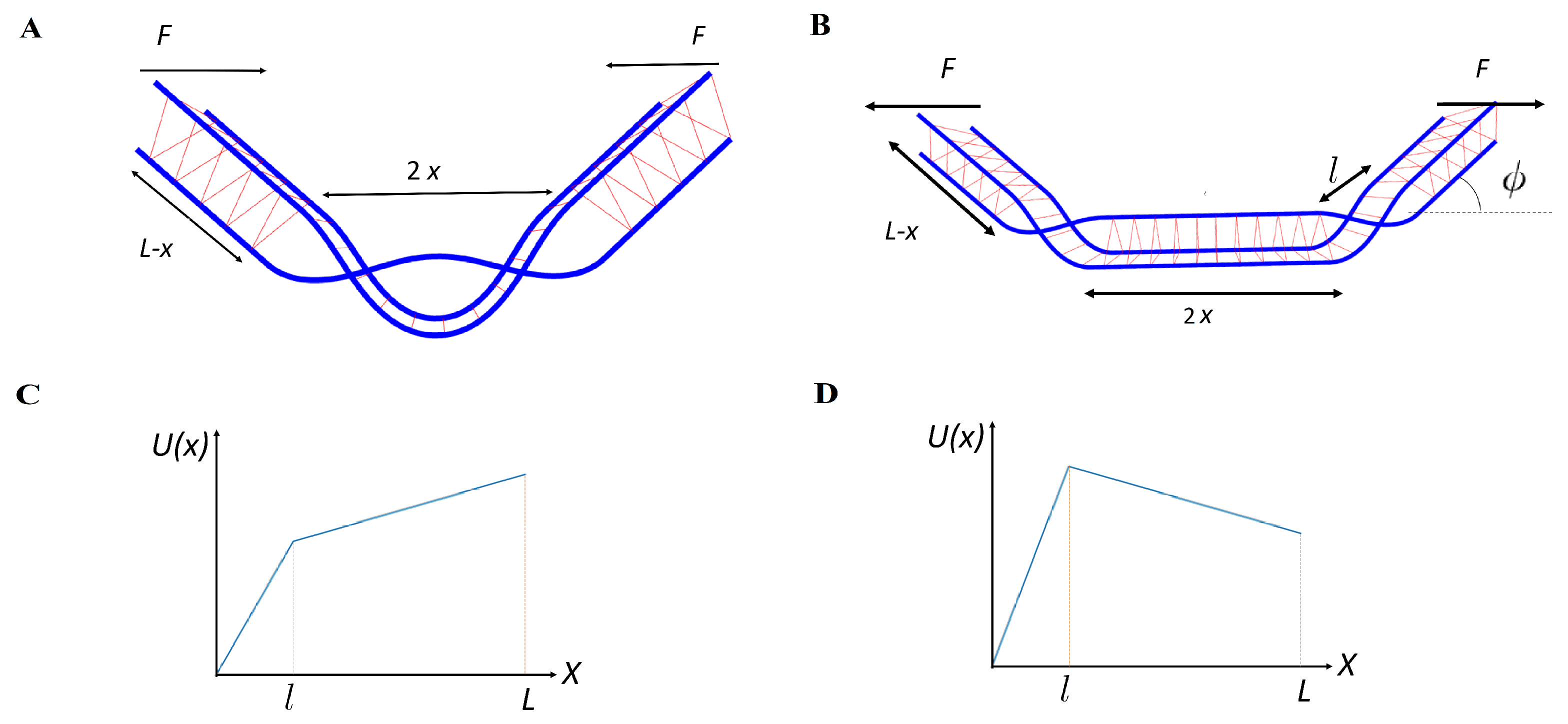
Taking these two aspects of the problem together, we may consider the stochastic pair production process as the thermal escape of a single fictitious particle, representing the reaction coordinate
x in an approximately piece-wise linear potential. Before defect pair separation,
, where
l is the size of the defect at the moment of the separation, the potential increases linearly as more length is exchanged between the defect pair and thus more cross links are removed. Understanding the exact form of this potential would require taking into account all different pathways leading from the properly cross-linked bundle to the bundle with braids. We make the simplest approximation, i.e., a linear potential, motivated by the fact that we need to remove the number of cross links proportional to the size of the uncross-linked region. Then, the effective potential is
, with
. The energy of the defect incorporates the bending energy of the filaments, the missing binding energy of the cross links absent in the defected region, and the work of the applied force:
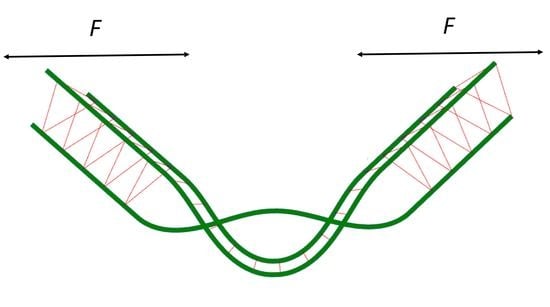
where
is the angle formed by a single braid (see
Figure 1),
F is a force acting on the ends of the bundle (positive sign is chosen for the extension), and
is a length of the bundle. The angle
is determined by energy minimization with respect to it. As the defects separate,
x grows. The angle also changes but as soon as
, this change contributes to the energy at the next order in the small parameter
. We address the explicit dependence of the angle on interbraid separation in
Section 3.4. Omitting this effect, we can again write an effective potential
with
. Thus, we may explore the pair production process using the following potential:
Before considering dynamics, we use this potential to consider the equilibrium distribution of defects on a bundle of length
L. Specifically, we consider the equilibrium separation of two defects. Using Equation (
6), it is trivial to write the probability distribution in this potential:
where the partition function
Z is given by the following:
and
is the inverse temperature. A straightforward calculation arrives at the partition function written as the sum of two parts corresponding to the two pieces of the potential as follows:
with
and
Taking the ratio of these partition sums, we obtain the ratio of observable separated braid pairs to strongly interacting and co-localized braids:
From the above, we see that in the thermal equilibrium, we expect there to be a low density of separated braids, at least at low temperatures (
K). In particular, if the thermal energy is much lower than the cross-linker binding energy (which is typically true in biopolymer filament systems) we expect an exponentially small density of braids (
, where for known filaments
. In fact, it appears that the smallest value of
is found for DNA bundles condensed by polyvalent ions where
[
6,
9,
10]. Braid pairs, however, can be generated either during bundle formation or via cycles of compression and expansion, as would be expected in a bundle network under reciprocal shear.
3.3. The Nonequilibrium Braid Distribution in a Time-Dependent Potential
When one applies a time-varying force, the effective potential controlling the production and motion of the braids also changes in time. As a result, we cannot rely on the equilibrium distribution discussed in the previous section. Instead, we have to solve the Smoluchowski diffusion equation for defect density
:
where the force now takes the following form:
During compression and . During expansion , (note that is negative). In the above, D is the defect diffusion constant. We cannot solve this equation analytically; however, we can provide a qualitative analysis. To simplify, we assume that A and B are fixed during each period of compression and expansion. We explore how the defect production rate depends on the lengths of these periods of compression and extension. We also estimate the maximal defect production rate.
The transport time from 0 to
l in the potential is controlled by the constant
A. This is the braid pair production rate when the braids have a stored length of
l. This problem is simply the first passage time [
16] to reach
l in the linearly increasing potential, which we may estimate to be as follows:
Similarly, we estimate the transport time from
L to
l. This gives an approximate value of the lifetime of the braid pair since when their separation returns to
l, they will likely annihilate. Here, we must distinguish between two limiting cases. In the first case, we consider purely diffusive braid motion and in the second, we look at the deterministic transport of the braids under an applied force, using a mobility set by the diffusion constant and the Einstein relation. We find the following:
In general, where we expect there to be both diffusive and advective defect motion, we find that the time for defects to recombine is the following:
In the limit of large L, so that the time for distant defect pairs to come together and potentially annihilate is much greater than their production time. If braids are able to separate sufficiently, we expect this ordering of time scales to be valid and thus predict braid proliferation on the bundle.
Because the production time
has an exponential dependence on
A,
since the
A parameter is much larger under stretching than it is under compression,
. If we choose the time dependence of the applied force so that the stretching time
and compression
satisfy the inequalities
we may analyze the dynamics of the system using a few approximations.
Since the braid production time during compression is small compared to the compression time:
, then, during the compression period, the density on the left
equilibrates. Since
, during the stretching period, the probability density
decreases near the potential maximum at
, but it is highly unlikely that thermally excited hopping over the barrier at
l occurs. Since
, the applied force is changing sufficiently fast that the density of the right of the potential
may be replaced by its time-averaged value. Moreover, since
, density on the right of the potential is effectively determined by the dynamics during the stretching period. Because the compression period is shorter, the already produced braids are unlikely to be driven together and annihilate. As a result, we conclude that the density distribution on the left is effectively determined by the compression period and the density on the right by the stretching period. Finally, we note that the Smoluchowski diffusion equation requires the continuity of both the probability density and its current at the boundary
. From these conditions, we obtain a value of the averaged density on the right as a function of the equilibrium density on the left. This implies that with this sequence of inequalities, the braid production and separation may be considered to take place in a time-averaged, effective potential where the production part is set by the compression forces and braids separate under a force related to extension. Specifically, we consider the following:
In this potential, the probability of braids on the left
will be proportional to the following:
while on the right,
The ratio is as follows:
which exceeds the case of only compression by a factor of
, and the case of only expansion by a factor of
—see Equation (
11).
3.4. Constant Force Stretching Dynamics of a Braided Bundle
We now consider an experiment in which one stretches a previously compressed bundle (by laser tweezers or other means) at a fixed force and determines the time rate of change of the bundle’s length. This is akin to a step force rheological measurement, and is closely related to determining the force-extension curve of a filament or filament bundle. Typically, in such force extension measurements, one considers the limit of slow extension so that the observed length corresponds the thermal equilibrium prediction under a fixed force [
17]. In this case, however, the extension of the bundle will be time dependent, even though the force is constant.
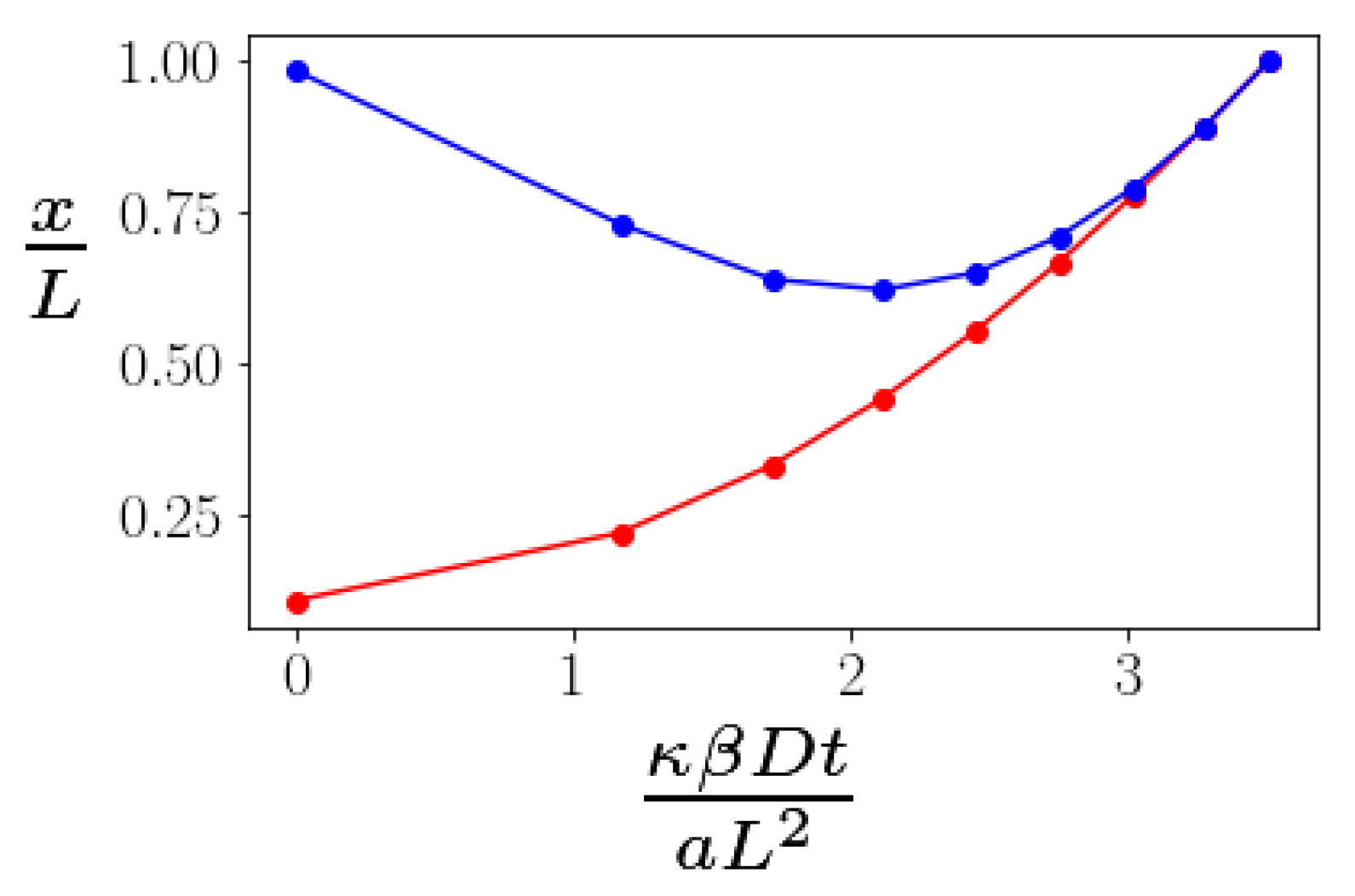
To study this problem, we minimize the energy of the bundle under a fixed stretching force. By doing so, we assume that the bending of the kink angles at the braids is fast compared to the time scale of measurement of the end-to-end distance. We do not, however, assume that the advection and diffusion of the braids is similarly fast. Doing this energy minimization numerically, we obtain the energy of the bundle as a function of the distance between two defects. Then, the average displacement will be controlled by a drift velocity
. Solving this equation numerically, we find time dependence shown in
Figure 4. The time dependence of the bundle’s extension is nonmonotonic: as defects diffuse from each other, the bundle initially becomes shorter. This happens due to the fact that as
x grows, the
becomes smaller; hence, the moment of the force decreases as well. Since the stretching torque decreases, the kinks become less stretched and their angles increase (see
Figure 3), decreasing the end-to-end distance
.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}



