
Time-Resolved Fluorescence Spectroscopy Study of Energy Transfer Dynamics in Phycobilisomes from Cyanobacteria Thermosynechococcus vulcanus NIES 2134 and Synechocystis sp. PCC 6803

 ,
, 
 ,
,  and
and 
Abstract
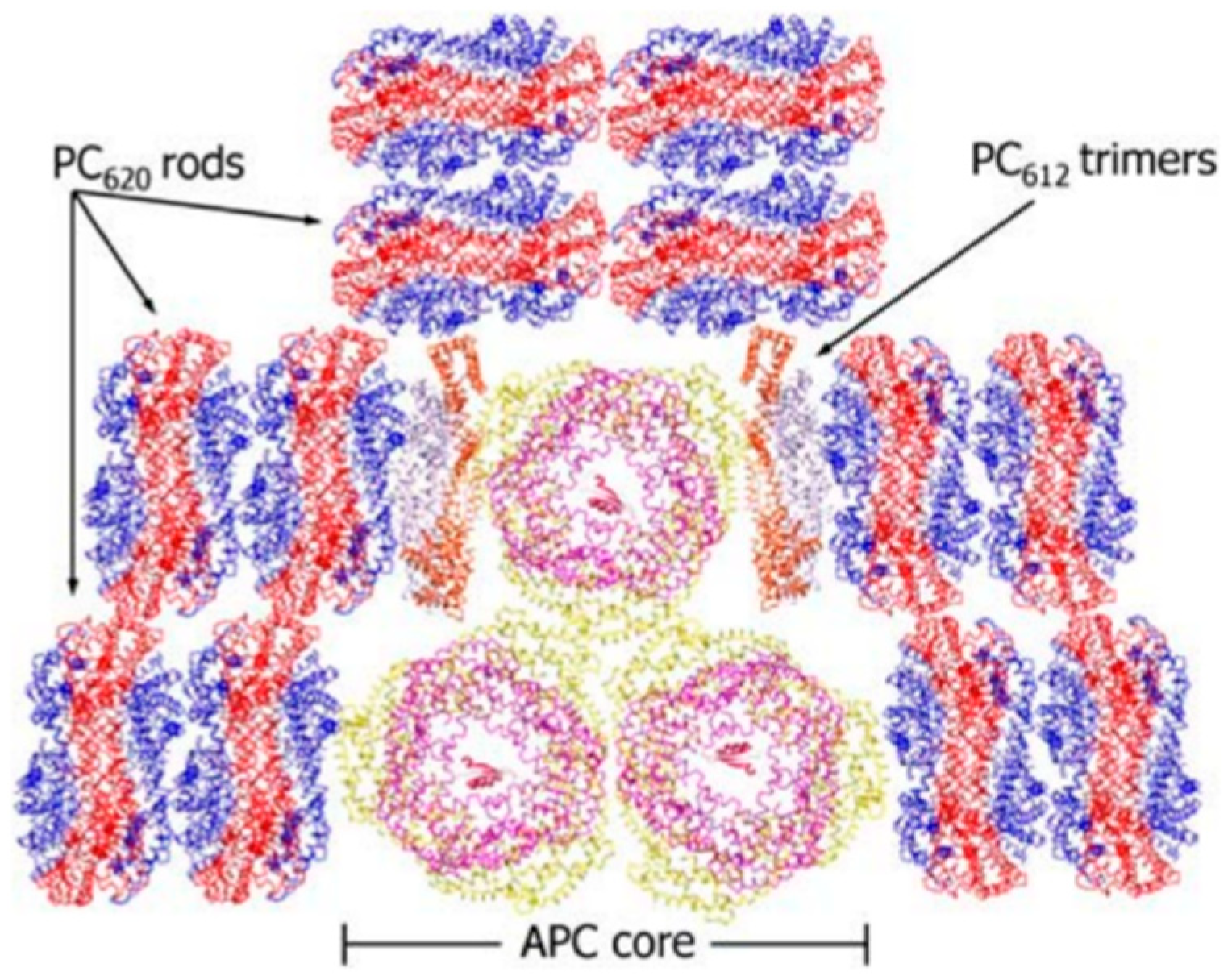
:1. Introduction
2. Materials and Methods
2.1. Growth of Cyanobacteria and Isolation of PBSs
2.2. Detection of Steady-State and TRFS
2.3. Analysis of the TRFS
3. Results
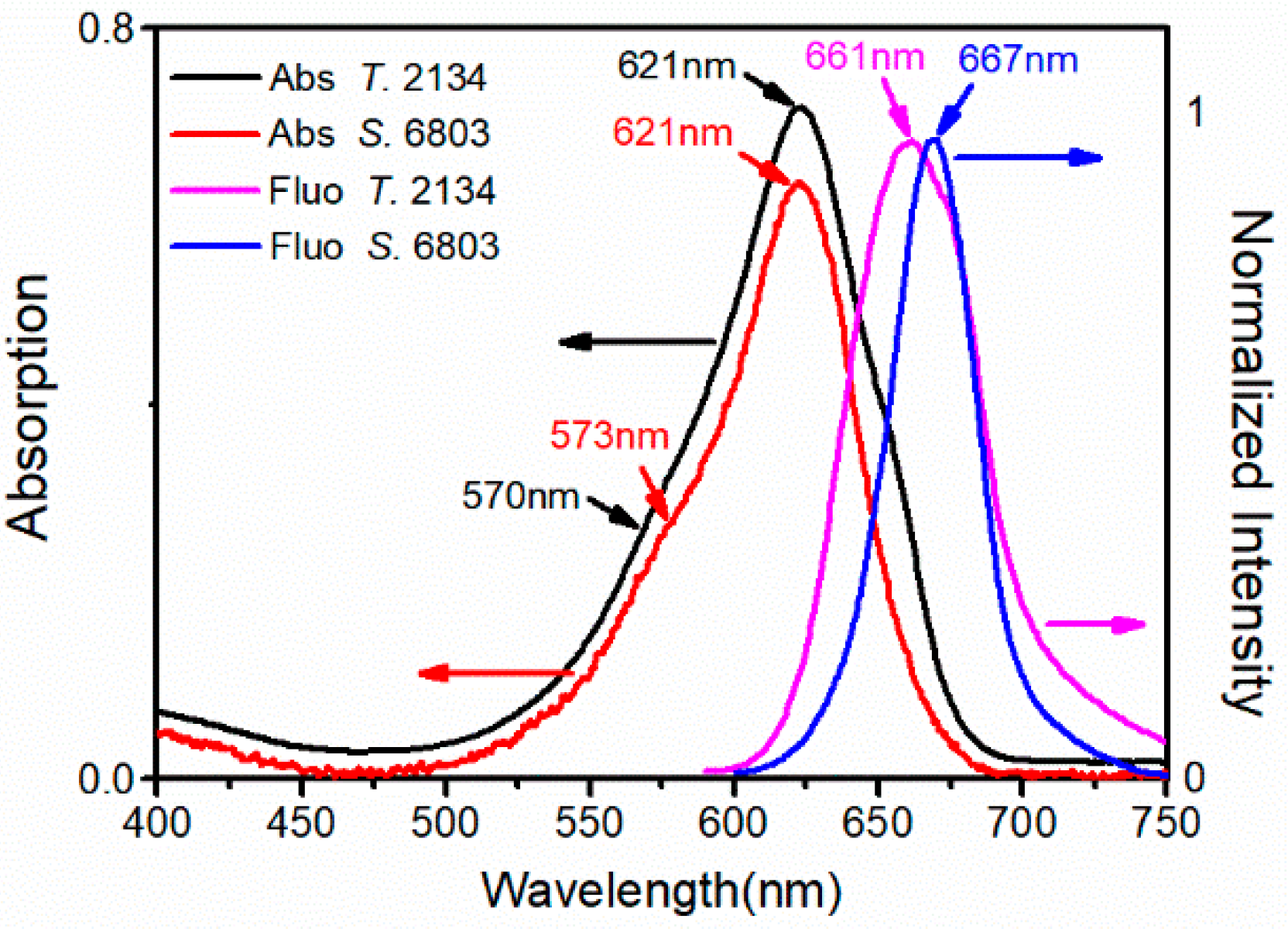
3.1. Steady-State and Transient-State Spectroscopy Results
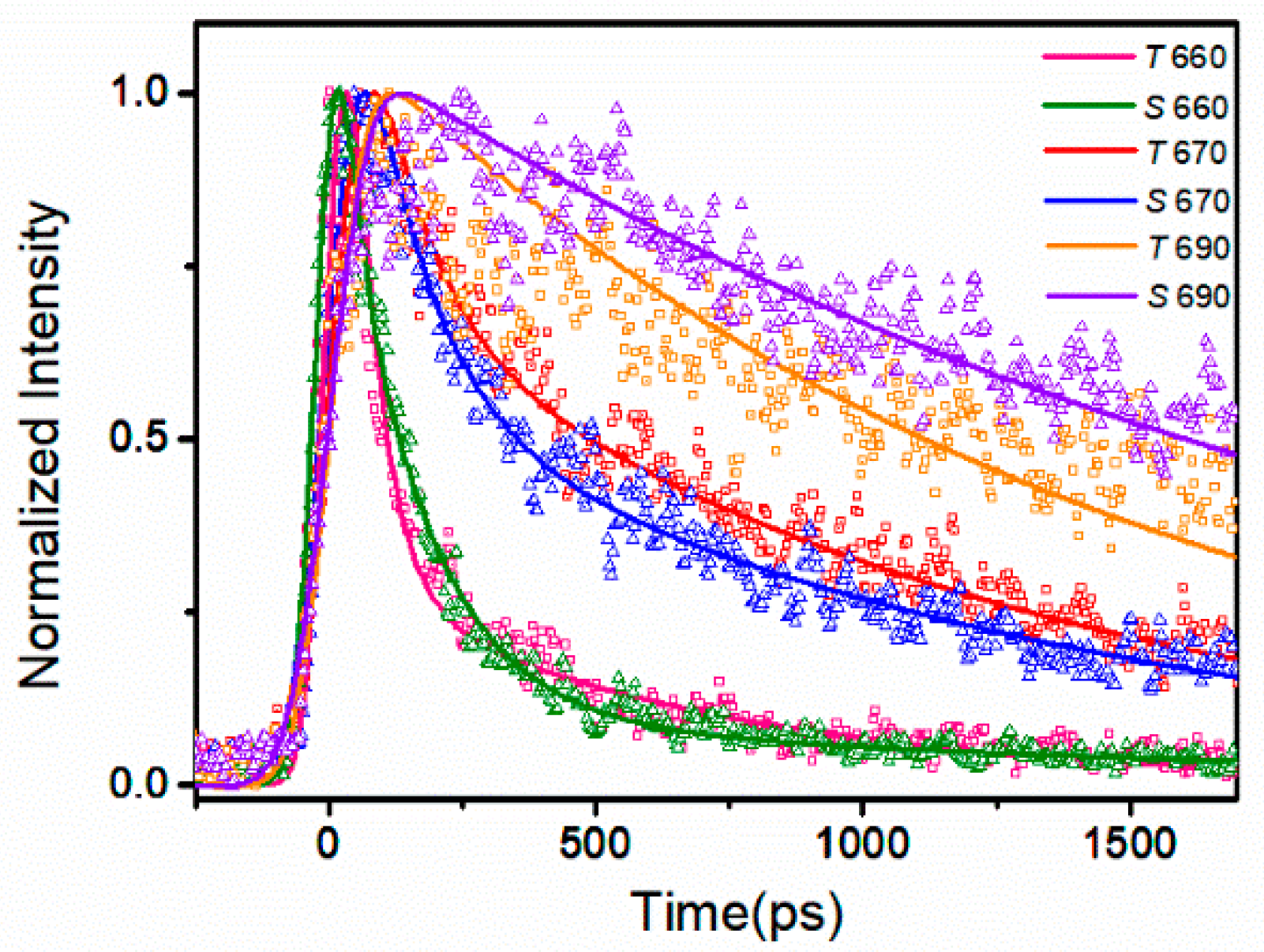
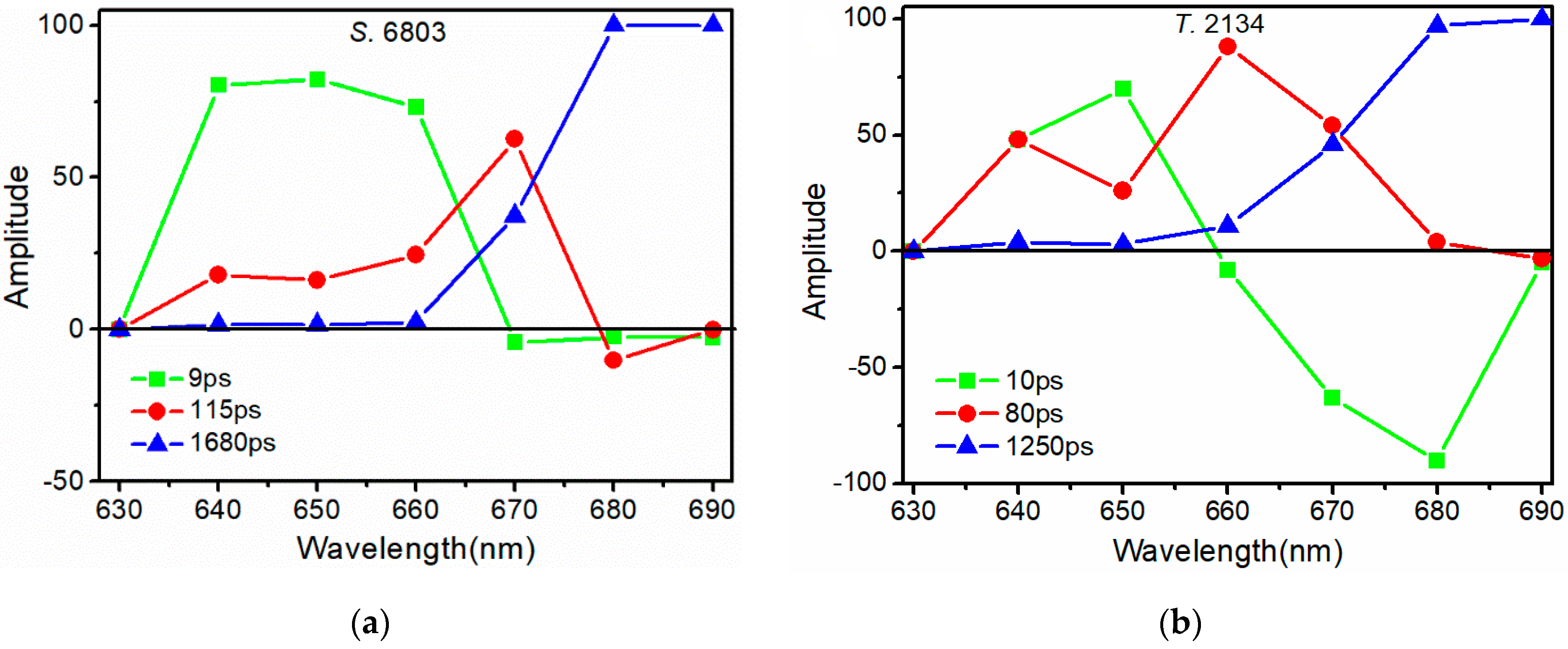
3.2. Time-Resolved Spectroscopy Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Garcia-Pichel, F. Cyanobacteria. Encyclopedia of Microbiology, 3rd ed.; Moselio, S., Ed.; Academic Press: San Diego, CA, USA, 2009; pp. 107–124. [Google Scholar]
- Glazer, A.N. Light harvesting by phycobilisomes. Annu. Rev. Biophys. Biophys. Chem. 1985, 14, 47–77. [Google Scholar] [CrossRef]
- Liu, L.; Chen, X.; Zhang, Y.; Zhou, B. Characterization, structure and function of linker polypeptides in phycobilisomes of cyanobacteria and red algae: An overview. Biochim. Biophys. Acta 2005, 1708, 133–142. [Google Scholar] [CrossRef] [Green Version]
- Tian, L.; Gwezdala, M.; van Stokkum, I.H.M.; Koehorst, R.B.M.; Kirilovsky, D.; van Amerongen, H. Picosecond kinetics of light harvesting and photoprotective quenching in wild-type and mutant phycobilisomes isolated from the cyanobacterium synechocystis PCC 6803. Biophys. J. 2012, 102, 1692–1700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Falamas, A.; Porav, S.A.; Tosa, V. Investigations of the energy transfer in the phycobilisome antenna of arthrospira platensis using femtosecond spectroscopy. Appl. Sci. 2020, 10, 4045. [Google Scholar] [CrossRef]
- Kolodny, Y.; Zer, H.; Propper, M.; Yochelis, S.; Paltiel, Y.; Keren, N. Marine cyanobacteria tune energy transfer efficiency in their light-harvesting antennae by modifying pigment coupling. FEBS. J. 2021, 288, 980–994. [Google Scholar] [CrossRef] [PubMed]
- Adir, N. Elucidation of the molecular structures of components of the phycobilisome: Reconstructing a giant. Photosynth. Res. 2005, 85, 15–32. [Google Scholar] [CrossRef] [PubMed]
- Gantt, E. Phycobilisomes. Annu. Rev. Plant Physiol. 1981, 32, 327–347. [Google Scholar] [CrossRef]
- David, L.; Marx, A.; Adir, N. High-resolution crystal structures of trimeric and rod phycocyanin. J. Mol. Biol. 2011, 405, 201–213. [Google Scholar] [CrossRef]
- Schirmer, T.; Bode, W.; Huber, R.; Sidler, W.; Zuber, H. X-ray crystallographic structure of the light-harvesting biliprotein c-phycocyanin from the thermophilic cyanobacterium mastigocladus laminosus and its resemblance to globin structures. J. Mol. Biol. 1985, 184, 257–277. [Google Scholar] [CrossRef]
- Berns, D.S.; MacColl, R. Phycocyanin in Physical Chemical Studies. Chem. Rev. 1989, 89, 807–825. [Google Scholar] [CrossRef]
- Adir, N.; Dobrovetsky, Y.; Lerner, N. Structure of c-phycocyanin from the thermophilic cyanobacterium synechococcus vulcanus at 2.5 Å: Structural implications for thermal stability in phycobilisome sssembly. J. Mol. Biol. 2001, 313, 71–81. [Google Scholar] [CrossRef] [Green Version]
- Brejc, K.; Ficner, R.; Huber, R.; Steinbacher, S. Isolation, crystallization, crystal structure analysis and refinement of allophycocyanin from the cyanobacterium spirulina platensis at 2.3 Å resolution. J. Mol. Biol. 1995, 249, 424–440. [Google Scholar] [CrossRef] [PubMed]
- Ying, L.; Xie, X. Fluorescence spectroscopy, exciton dynamics, and photochemistry of single allophycocyanin trimers. J. Phys. Chem. B 1998, 102, 10399–10409. [Google Scholar] [CrossRef]
- McGregor, A.; Klartag, M.; David, L.; Adir, N. Allophycocyanin trimer stability and functionality are primarily due to polar enhanced hydrophobicity of the phycocyanobilin binding pocket. J. Mol. Biol. 2008, 384, 406–421. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Moerner, W.E. Dissecting pigment architecture of individual photosynthetic antenna complexes in solution. Proc. Natl. Acad. Sci. USA 2015, 112, 13880–13885. [Google Scholar] [CrossRef] [Green Version]
- Womick, J.M.; Moran, A.M. Exciton coherence and energy transport in the light-harvesting dimers of allophycocyanin. J. Phys. Chem. B 2009, 113, 15747–15959. [Google Scholar] [CrossRef]
- Zhang, J.; Zhao, J.; Jiang, L.; Zheng, X.; Zhao, F.; Wang, H. Studies on the energy transfer among the rod-core complex from phycobilisome of Anabaena variabilis by time resolved fluorescence emission and anisotropy spectra. Biochim. Biophys. Acta Bioener. 1997, 1320, 285–296. [Google Scholar] [CrossRef] [Green Version]
- Hirota, Y.; Serikawa, H.; Kawakami, K.; Ueno, M.; Kamiya, N.; Kosumi, D. Ultrafast energy transfer dynamics of phycobilisome from Thermosynechococcus vulcanus, as revealed by ps fluorescence and fs pump-probe spectroscopies. Photosynth. Res. 2021, 148, 181–190. [Google Scholar] [CrossRef]
- Adir, N.; Lerner, N. The crystal structure of a novel unmethylated form of c-phycocyanin, a possible connector between cores and rods in phycobilisomes. J. Biol. Chem. 2003, 278, 25926–25932. [Google Scholar] [CrossRef] [Green Version]
- Marx, A.; Adir, N. Allophycocyanin and phycocyanin crystal structures reveal facets of phycobilisome assembly. Biochim. Biophys. Acta Bioener. 2013, 1827, 311–318. [Google Scholar] [CrossRef] [Green Version]
- David, L.; Prado, M.; Arteni, A.; Elmlund, D.; Blankenship, R.; Adir, N. Structural studies show energy transfer within stabilized phycobilisomes independent of the mode of rod-core assembly. Biochim. Biophys. Acta 2014, 1837, 385–395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adir, N.; Dines, M.; Klartag, M.; McGregor, A.; Melamed-Frank, M. Assembly and Disassembly of Phycobilisomes. In Complex Intracellular Structures in Prokaryotes. Microbiology Monographs, 1st ed.; Shively, J.M., Ed.; Springer: Berlin/Heidelberg, Germany, 2006; Volume 2, pp. 47–77. [Google Scholar]
- Ma, J.; You, X.; Sun, S.; Wang, X.; Qin, S.; Sui, S. Structural basis of energy transfer in Porphyridium purpureum phycobilisome. Nature 2020, 579, 146–151. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zhao, J.; Jiang, L. Energy transfer processes in phycobilisome model complex at 77 K. China Ser. B-Chem. 2000, 43, 233–239. [Google Scholar] [CrossRef]
- Zhao, F.; Zheng, X.; Zhang, J.; Wang, H.; Yu, Z.; Zhao, J.; Jiang, L. Energy transfer in allophycocyanin hexamer from Anabaena variabilis by time-resolved spectroscopy. J. Photochem. Photobiol. B 1998, 45, 144–149. [Google Scholar] [CrossRef]
- Sandstrom, A.; Gillbro, T.; Sandstrom, U.; Fischer, R.; Scheer, H. Picosecond study of energy transfer within 18-S particles of AN 112 (a mutant of Synechococcus 6301) phycobilisomes. Biochim. Biophys. Acta Bioener. 1988, 933, 54–64. [Google Scholar] [CrossRef]
- Zhang, J.; Zhao, F.; Zheng, X.; Wang, H. Direct measurement of excitation transfer dynamics between two trimers in C-phycocyanin hexamer from cyanobacterium Anabaena variabilis. Chem. Phys. Lett. 1999, 304, 357–364. [Google Scholar] [CrossRef]
- Zhao, F.; Zhang, J.; Zheng, X.; Wang, H. Energy transfer among proteins in the phycobilisome of red algae. II. Energy transfer in synthesized complex of R-PE/R-PC and R-PE/APC. Zhongshan Da Xue Xue Bao. Zi Ran Ke Xue Ban Acta Sci. Nat. Univ. Sunyatseni 1998, 37, 18–23. [Google Scholar]
- Holzwarth, A.R.; Wendler, J.; Suter, G.W. Studies on chromophore coupling in isolated phycobiliproteins: II. Picosecond energy transfer kinetics and time-resolved fluorescence spectra of C-phycocyanin from synechococcus 6301 as a function of the aggregation state. Biophys. J. 1987, 51, 1–12. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Eem/nm | τ1/ps | A1/% | τ2/ps | A2/% | τ3/ps | A3/% |
|---|---|---|---|---|---|---|
| 640 | 12 | 80 | 93 | 18 | 1692 | 2 |
| 650 | 11 | 82 | 127 | 16 | 1540 | 2 |
| 660 | 9 | 73 | 150 | 25 | 1613 | 2 |
| 670 | 8 | −4 | 120 | 62 | 1291 | 37 |
| 680 | 9 | −1 | 116 | −10 | 1869 | 100 |
| 690 | 8 | −3 | 88 | −1 | 2088 | 100 |
| Average | 9 | 115 | 1680 |
| Eem/nm | τ1/ps | A1/% | τ2/ps | A2/% | τ3/ps | A3/% |
|---|---|---|---|---|---|---|
| 640 | 14 | 48 | 76 | 48 | 1099 | 4 |
| 650 | 10 | 70 | 101 | 26 | 1123 | 3 |
| 660 | 9 | −8 | 63 | 88 | 929 | 11 |
| 670 | 11 | −63 | 104 | 54 | 1218 | 46 |
| 680 | 10 | −90 | 76 | 4 | 1744 | 97 |
| 690 | 8 | −5 | 66 | −3 | 1369 | 100 |
| Average | 10 | 80 | 1250 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xie, M.; Li, W.; Xiao, C.; Zhen, Z.; Ma, J.; Lin, H.; Qin, S.; Zhao, F. Time-Resolved Fluorescence Spectroscopy Study of Energy Transfer Dynamics in Phycobilisomes from Cyanobacteria Thermosynechococcus vulcanus NIES 2134 and Synechocystis sp. PCC 6803. Crystals 2021, 11, 1233. https://doi.org/10.3390/cryst11101233
Xie M, Li W, Xiao C, Zhen Z, Ma J, Lin H, Qin S, Zhao F. Time-Resolved Fluorescence Spectroscopy Study of Energy Transfer Dynamics in Phycobilisomes from Cyanobacteria Thermosynechococcus vulcanus NIES 2134 and Synechocystis sp. PCC 6803. Crystals. 2021; 11(10):1233. https://doi.org/10.3390/cryst11101233
Chicago/Turabian StyleXie, Mingyuan, Wenjun Li, Chao Xiao, Zhanghe Zhen, Jianfei Ma, Hanzhi Lin, Song Qin, and Fuli Zhao. 2021. "Time-Resolved Fluorescence Spectroscopy Study of Energy Transfer Dynamics in Phycobilisomes from Cyanobacteria Thermosynechococcus vulcanus NIES 2134 and Synechocystis sp. PCC 6803" Crystals 11, no. 10: 1233. https://doi.org/10.3390/cryst11101233
APA StyleXie, M., Li, W., Xiao, C., Zhen, Z., Ma, J., Lin, H., Qin, S., & Zhao, F. (2021). Time-Resolved Fluorescence Spectroscopy Study of Energy Transfer Dynamics in Phycobilisomes from Cyanobacteria Thermosynechococcus vulcanus NIES 2134 and Synechocystis sp. PCC 6803. Crystals, 11(10), 1233. https://doi.org/10.3390/cryst11101233