Truncated Prosequence of Rhizopus oryzae Lipase: Key Factor for Production Improvement and Biocatalyst Stability



Abstract
1. Introduction
2. Results and Discussion
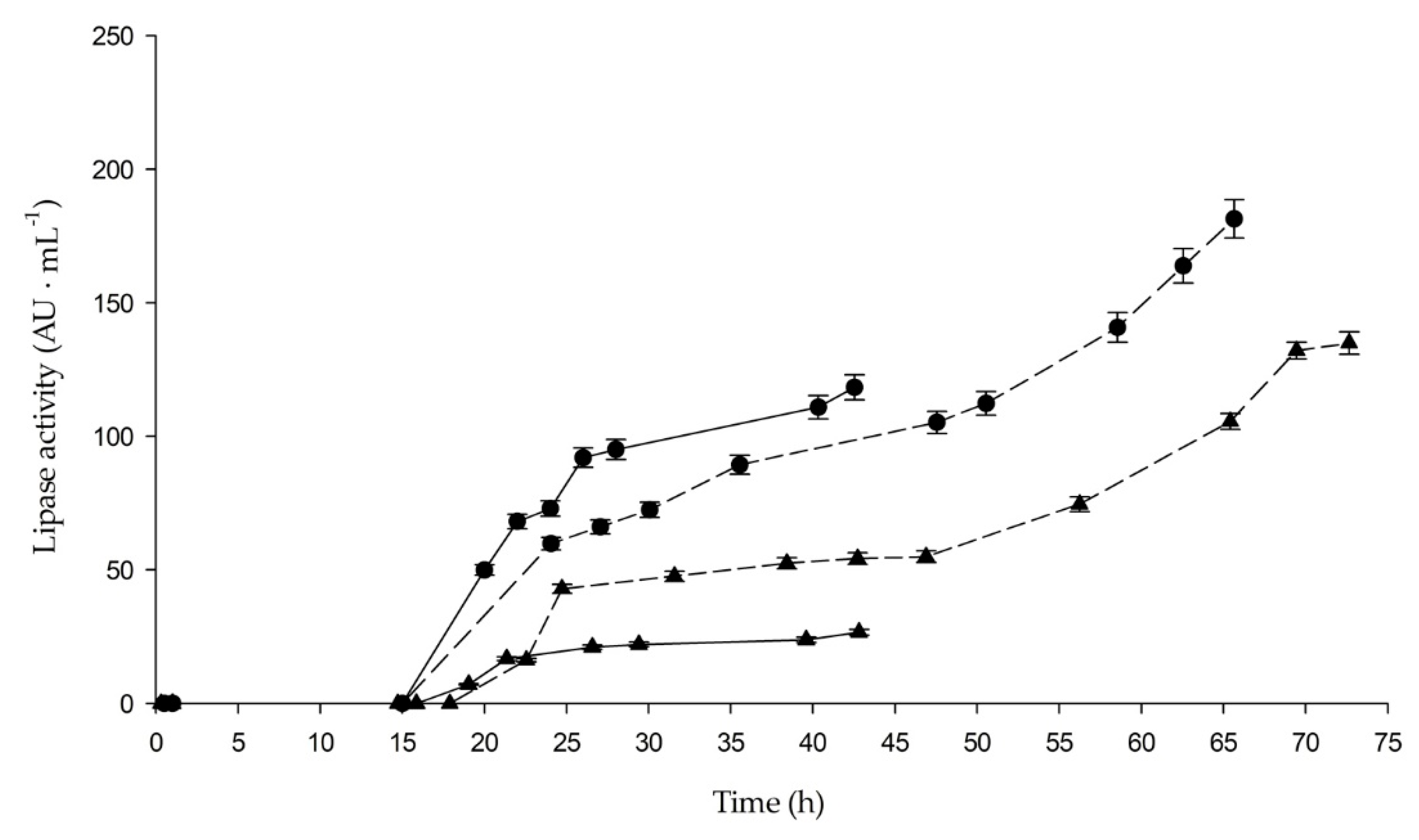
2.1. Batch and Fed-Batch Production of proROL and rROL
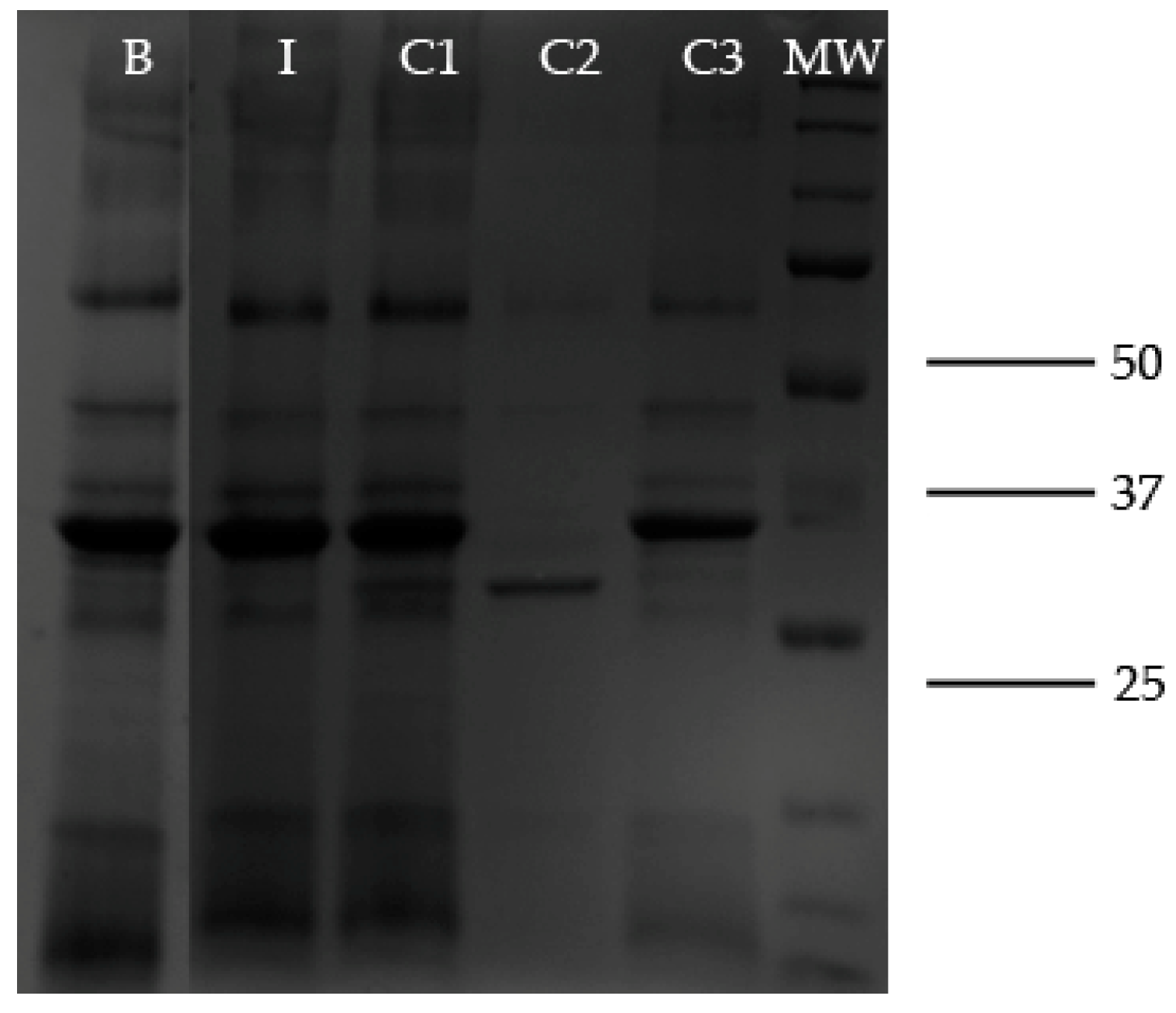
2.2. Electrophoretic Studies
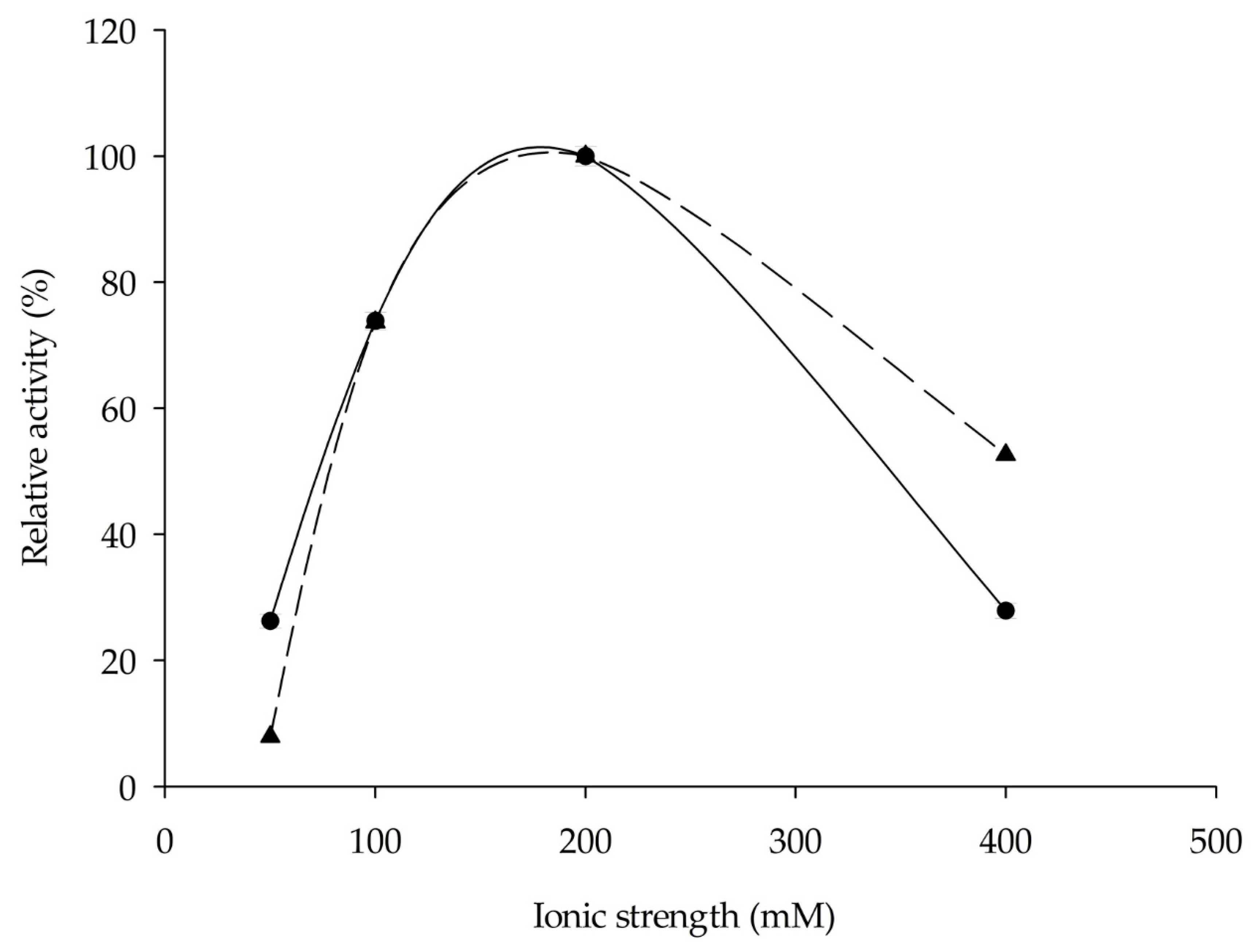
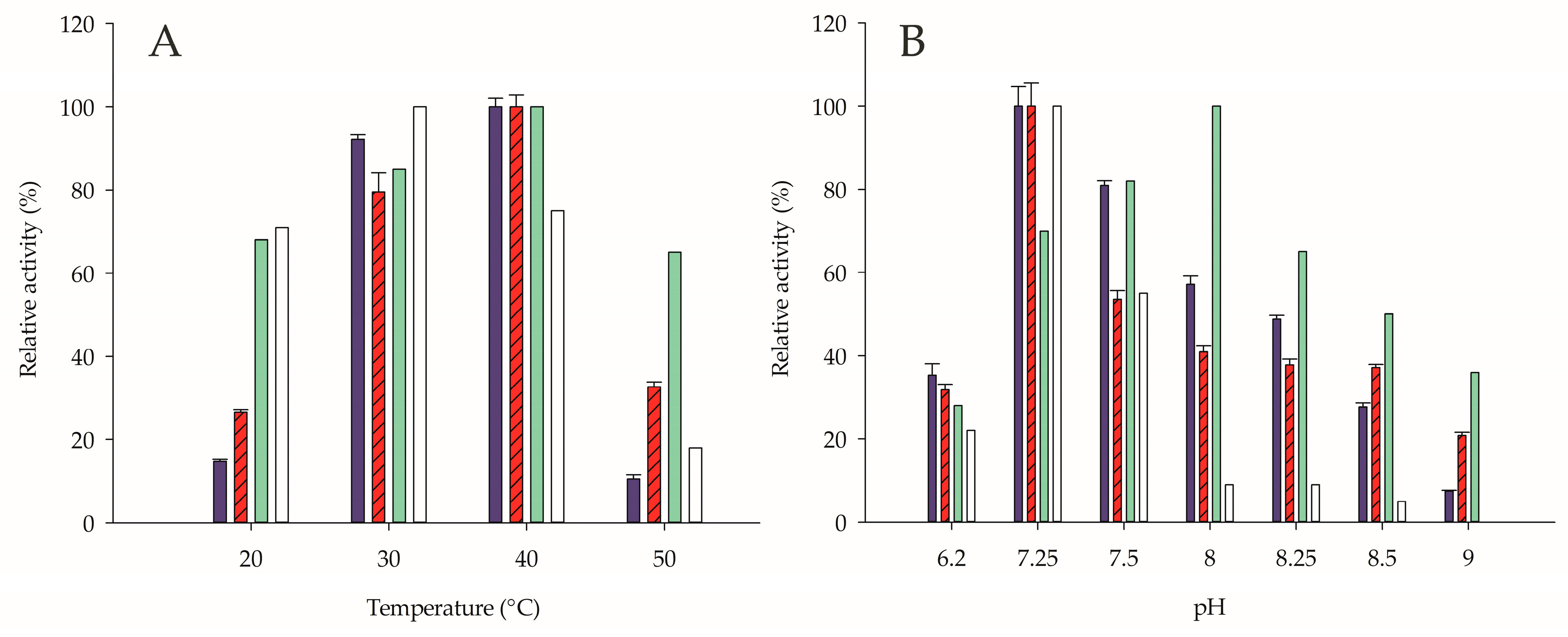
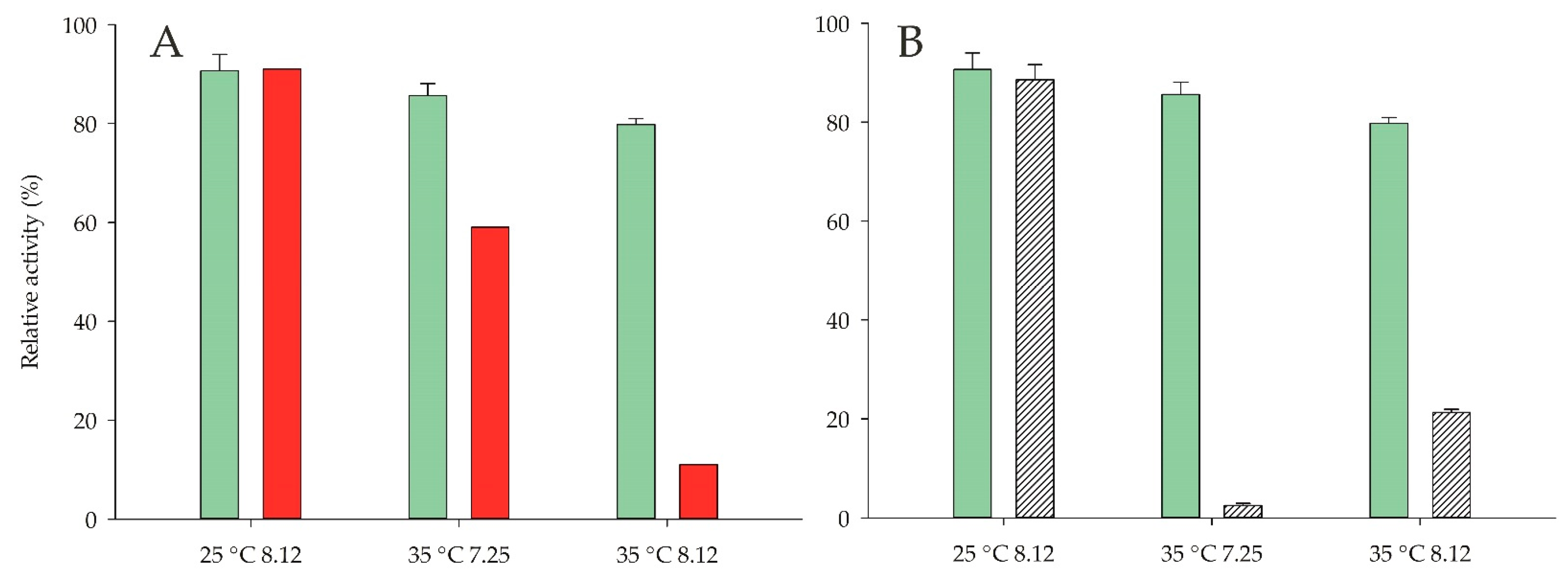
2.3. Influence of Ionic Strength, Temperature, and pH on Enzyme Activity
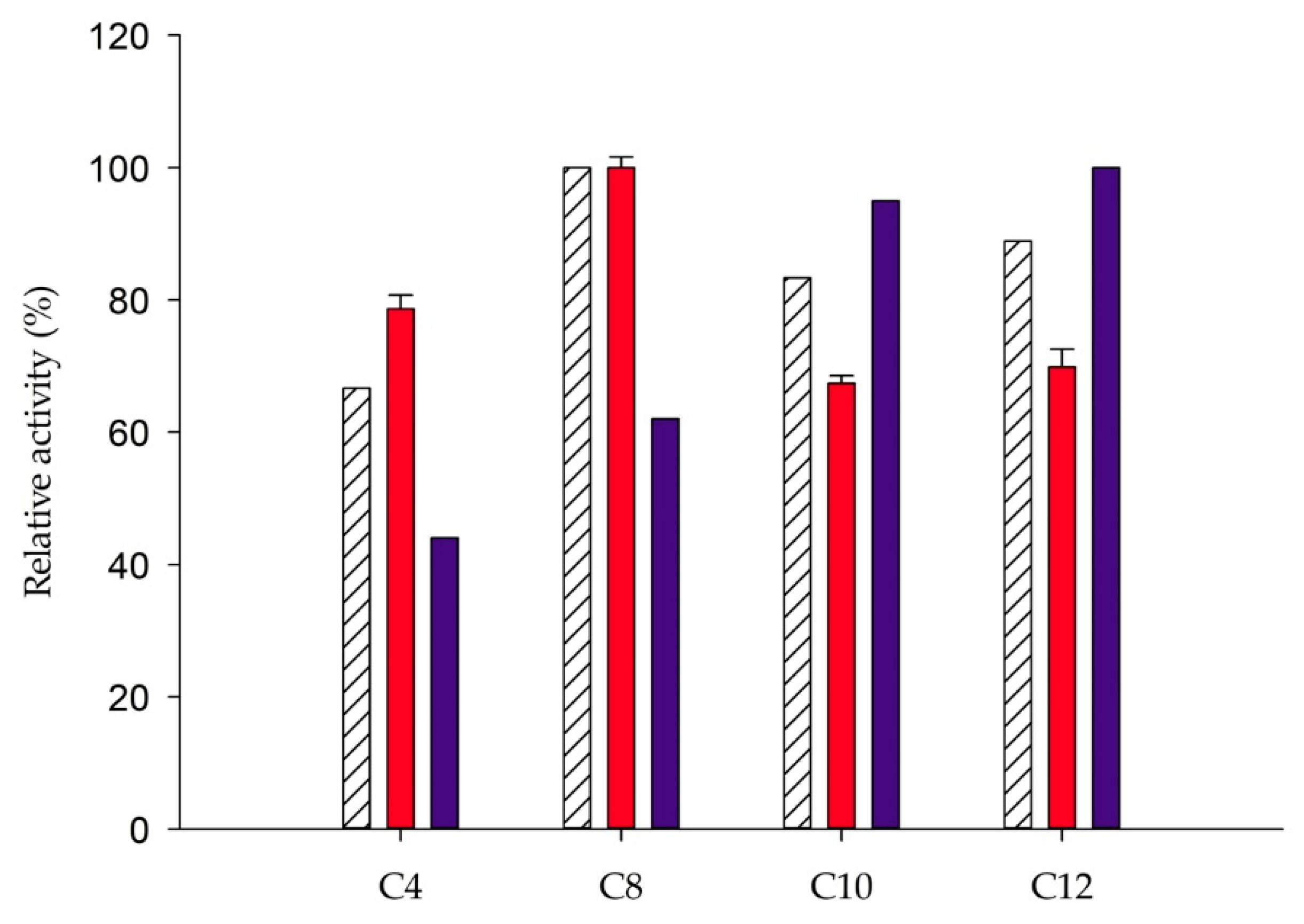
2.4. Substrate Specificity
2.5. proROL Lipase Proteolysis
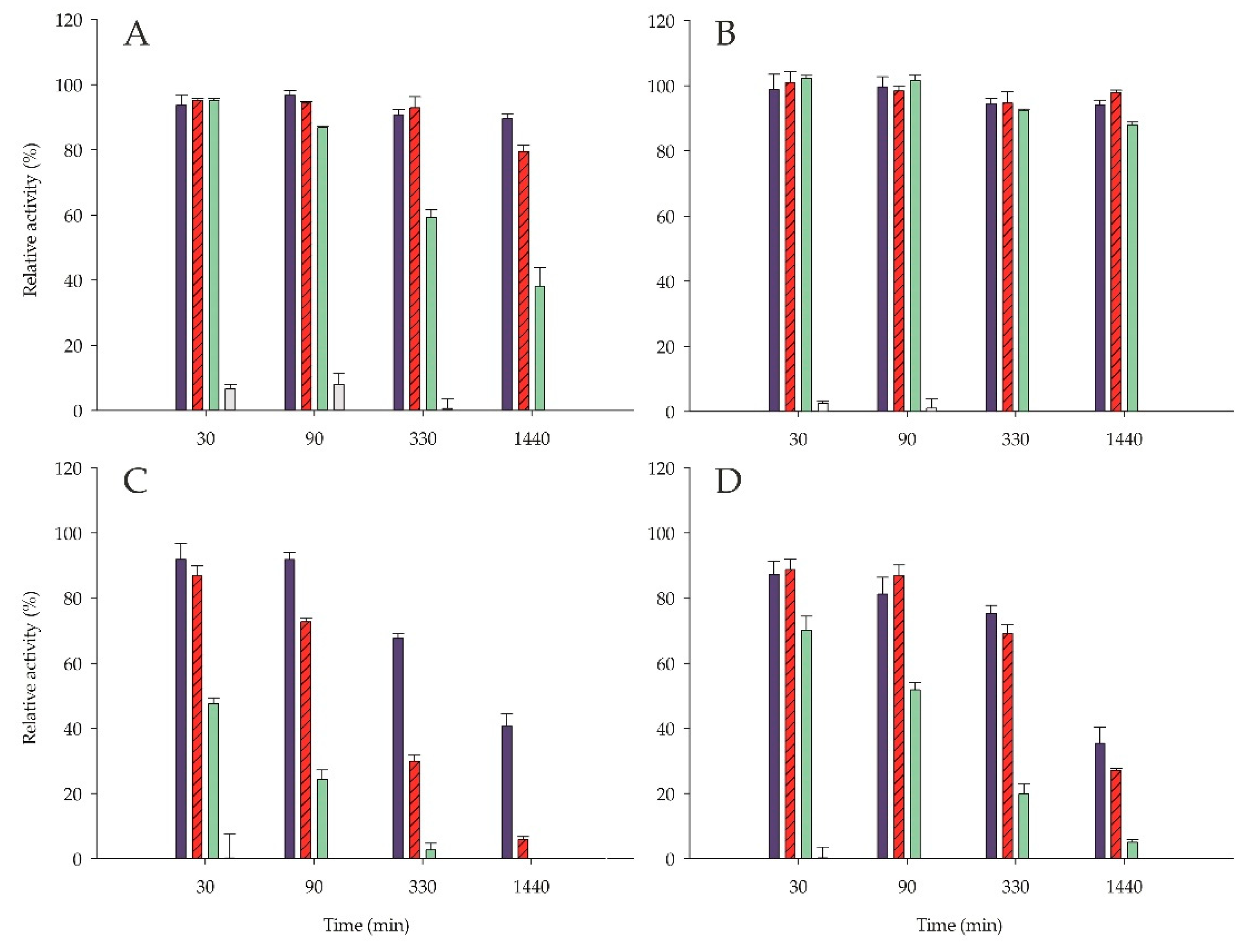
2.6. Stability
3. Materials and Methods
3.1. Strains and Derivative Plasmids
3.2. Batch Cultures
3.3. Fed-Batch Cultures
3.4. Lipases
3.5. Chemicals
3.6. Electrophoresis
3.7. Lipolytic Activity
3.8. Total Protein
3.9. Substrate Specificity
3.10. Proteolytic Activity Inhibition
3.11. Enzyme Stability
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ROL | Rhizopus oryzae lipase |
| rROL | Recombinant Rhizopus oryzae lipase formed by the mature sequence |
| nROL | Native Rhizopus oryzae lipase |
| proROL | Lipase formed by 28 amino acids of the C-terminal of the natural prosequence of ROL fused to the N-terminal of the mature sequence. |
| rROL-strain | Genetically modified Pichia pastoris strain to produce rROL |
| proROL-strain | Genetically modified Pichia pastoris strain to produce proROL |
| MeOH | Methanol |
| MNLF | Methanol non-limiting fed-batch |
| MLFB | Methanol limiting fed-batch |
| Y(P/X) | Product-biomass yield (AU·gX−1) |
| Y(X/S) | Biomass-substrate yield (gX·gS−1) |
| µ | specific growth rate (h−1) |
| µmax | maximum specific growth rate (h−1) |
| qp | specific production rate (AU·gX−1·h−1) |
| UPR | Unfolded protein response |
| PAOX1 | Inducible promoter alcohol oxidase 1 |
References
- Sandoval, G.; Casas-Godoy, L.; Bonet-Ragel, K.; Rodrigues, J.; Ferreira-Dias, S.; Valero, F. Enzyme-Catalyzed Production of Biodiesel as Alternative to Chemical-Catalyzed Processes: Advantages and Constraints. Curr. Biochem. Eng. 2017, 4, 109–141. [Google Scholar] [CrossRef]
- Manaf, I.S.A.; Embong, N.H.; Khazaai, S.N.M.; Rahim, M.H.A.; Yusoff, M.M.; Lee, K.T.; Maniam, G.P. A review for key challenges of the development of biodiesel industry. Energy Convers. Manag. 2019, 185, 508–517. [Google Scholar] [CrossRef]
- Guldhe, A.; Singh, B.; Mutanda, T.; Permaul, K.; Bux, F. Advances in synthesis of biodiesel via enzyme catalysis: Novel and sustainable approaches. Renew. Sustain. Energy Rev. 2015, 41, 1447–1464. [Google Scholar] [CrossRef]
- Medina, A.R.; González-Moreno, P.; Esteban-Cerdán, L.; Grima, E.M. Biocatalysis: Towards ever greener biodiesel production. Biotechnol. Adv. 2009, 27, 398–408. [Google Scholar] [CrossRef]
- Guillén, M.; Benaiges, M.D.; Valero, F. Immobilization and stability of a Rhizopus oryzae lipase expressed in Pichia pastoris: Comparison between native and recombinant variants. Biotechnol. Prog. 2011, 27, 1232–1241. [Google Scholar] [CrossRef]
- Lotti, M.; Pleiss, J.; Valero, F.; Ferrer, P. Effects of methanol on lipases: Molecular, kinetic and process issues in the production of biodiesel. Biotechnol. J. 2015, 10, 22–30. [Google Scholar] [CrossRef]
- Singh, R.K.; Tiwari, M.K.; Singh, R.; Lee, J.-K. From Protein Engineering to Immobilization: Promising Strategies for the Upgrade of Industrial Enzymes. Int. J. Mol. Sci. 2013, 14, 1232–1277. [Google Scholar] [CrossRef]
- Korman, T.P.; Sahachartsiri, B.; Charbonneau, D.M.; Huang, G.L.; Beauregard, M.; Bowie, J.U. Dieselzymes: Development of a stable and methanol tolerant lipase for biodiesel production by directed evolution. Biotechnol. Biofuels 2013, 6, 70. [Google Scholar] [CrossRef]
- Wang, Z.; Lv, P.; Luo, W.; Yuan, Z.; He, D. Expression in Pichia pastoris and characterization of Rhizomucor miehei lipases containing a new propeptide region. J. Gen. Appl. Microbiol. 2016, 62, 25–30. [Google Scholar] [CrossRef]
- Takahashi, S.; Ueda, M.; Atomi, H.; Beer, H.D.; Bornscheuer, U.T.; Schmid, R.D.; Tanaka, A. Extracellular production of active Rhizopus oryzae lipase by Saccharomyces cerevisiae. J. Ferment. Bioeng. 1998, 86, 164–168. [Google Scholar] [CrossRef]
- Satomura, A.; Kuroda, K.; Ueda, M. Generation of a Functionally Distinct Rhizopus oryzae Lipase through Protein Folding Memory. PLoS ONE 2015, 10, e0124545. [Google Scholar] [CrossRef] [PubMed]
- Canet, A.; Bonet-Ragel, K.; Benaiges, M.D.; Valero, F. Lipase-catalysed transesterification: Viewpoint of the mechanism and influence of free fatty acids. Biomass Bioenergy 2016, 85, 94–99. [Google Scholar] [CrossRef]
- Bonet-Ragel, K.; Canet, A.; Benaiges, M.D.; Valero, F. Synthesis of biodiesel from high FFA alperujo oil catalysed by immobilised lipase. Fuel 2015, 161, 12–17. [Google Scholar] [CrossRef]
- Picó, E.A.; López, C.; Cruz-Izquierdo, Á.; Munarriz, M.; Iruretagoyena, F.J.; Serra, J.L.; Llama, M.J. Easy reuse of magnetic cross-linked enzyme aggregates of lipase B from Candida antarctica to obtain biodiesel from Chlorella vulgaris lipids. J. Biosci. Bioeng. 2018, 126, 451–457. [Google Scholar] [CrossRef]
- Bonet-Ragel, K.; Canet, A.; Benaiges, M.D.; Valero, F. Effect of acyl-acceptor stepwise addition strategy using alperujo oil as a substrate in enzymatic biodiesel synthesis. J. Chem. Technol. Biotechnol. 2018, 93, 541–547. [Google Scholar] [CrossRef]
- Beer, H.; McCarthy, J.E.; Bornscheuer, U.T.; Schmid, R.D. Cloning, expression, characterization and role of the leader sequence of a lipase from Rhizopus oryzae. Biochim. Biophys. Acta BBA Bioenergy 1998, 1399, 173–180. [Google Scholar] [CrossRef]
- Sayari, A.; Frikha, F.; Miled, N.; Mtibaa, H.; Ali, Y.B.; Verger, R.; Gargouri, Y. N-terminal peptide of Rhizopus oryzae lipase is important for its catalytic properties. FEBS Lett. 2005, 579, 976–982. [Google Scholar] [CrossRef]
- Beer, H.D.; Wohlfahrt, G.; Schmid, R.D.; McCarthy, J.E.G. The folding and activity of the extracellular lipase of Rhizopus oryzae are modulated by a prosequence. Biochem. J. 1996, 319, 351–359. [Google Scholar] [CrossRef]
- Hama, S.; Tamalampudi, S.; Shindo, N.; Numata, T.; Yamaji, H.; Fukuda, H.; Kondo, A. Role of N-terminal 28-amino-acid region of Rhizopus oryzae lipase in directing proteins to secretory pathway of Aspergillus oryzae. Appl. Microbiol. Biotechnol. 2008, 79, 1009–1018. [Google Scholar] [CrossRef]
- Hiol, A.; Jonzo, M.D.; Rugani, N.; Druet, D.; Sarda, L.; Comeau, L.C. Purification and characterization of an extracellular lipase from a thermophilic Rhizopus oryzae strain isolated from palm fruit. Enzym. Microb. Technol. 2000, 26, 421–430. [Google Scholar] [CrossRef]
- Ben Salah, R.; Mosbah, H.; Fendri, A.; Gargouri, A.; Gargouri, Y.; Mejdoub, H. Biochemical and molecular characterization of a lipase produced by Rhizopus oryzae. FEMS Microbiol. Lett. 2006, 260, 241–248. [Google Scholar] [CrossRef] [PubMed]
- López, E.; Deive, F.J.; Longo, M.A.; Sanromán, M.Á.; Braga, M.; Ángeles, S. Lipolytic Enzyme Production by Immobilized Rhizopus oryzae. Chem. Eng. Technol. 2008, 31, 1555–1560. [Google Scholar] [CrossRef]
- Hama, S.; Tamalampudi, S.; Fukumizu, T.; Miura, K.; Yamaji, H.; Kondo, A.; Fukuda, H. Lipase localization in Rhizopus oryzae cells immobilized within biomass support particles for use as whole-cell biocatalysts in biodiesel-fuel production. J. Biosci. Bioeng. 2006, 101, 328–333. [Google Scholar] [CrossRef] [PubMed]
- Ueda, M.; Takahashi, S.; Washida, M.; Shiraga, S.; Tanaka, A. Expression of Rhizopus oryzae lipase gene in Saccharomyces cerevisiae. J. Mol. Catal. B Enzym. 2002, 17, 113–124. [Google Scholar] [CrossRef]
- Takahashi, S.; Ueda, M.; Tanaka, A. Independent production of two molecular forms of a recombinant Rhizopus oryzae lipase by KEX2-engineered strains of Saccharomyces cerevisiae. Appl. Microbiol. Biotechnol. 1999, 52, 534–540. [Google Scholar] [CrossRef]
- Takahashi, S.; Ueda, M.; Tanaka, A. Function of the prosequence for in vivo folding and secretion of active Rhizopus oryzae lipase in Saccharomyces cerevisiae. Appl. Microbiol. Biotechnol. 2001, 55, 454–462. [Google Scholar] [CrossRef]
- Joerger, R.D.; Haas, M.J. Overexpression of aRhizopus delemar lipase gene inEscherichia coli. Lipids 1993, 28, 81–88. [Google Scholar] [CrossRef]
- Di Lorenzo, M.; Hidalgo, A.; Haas, M.; Bornscheuer, U.T. Heterologous Production of Functional Forms of Rhizopus oryzae Lipase in Escherichia coli. Appl. Environ. Microbiol. 2005, 71, 8974–8977. [Google Scholar] [CrossRef]
- Minning, S.; Schmidt-Dannert, C.; Schmid, R.D. Functional expression of Rhizopus oryzae lipase in Pichia pastoris: High-level production and some properties. J. Biotechnol. 1998, 66, 147–156. [Google Scholar] [CrossRef]
- Potvin, G.; Ahmad, A.; Zhang, Z. Bioprocess engineering aspects of heterologous protein production in Pichia pastoris: A review. Biochem. Eng. J. 2012, 64, 91–105. [Google Scholar] [CrossRef]
- Ben Salah, R.; Gargouri, A.; Verger, R.; Gargouri, Y.; Mejdoub, H. Expression in Pichia pastoris X33 of His-tagged lipase from a novel strain of Rhizopus oryzae and its mutant Asn 134 His: Purification and characterization. World J. Microbiol. Biotechnol. 2009, 25, 1375–1384. [Google Scholar] [CrossRef]
- Yu, X.W.; Yang, M.; Jiang, C.; Zhang, X.; Xu, Y. N-Glycosylation Engineering to Improve the Constitutive Expression of Rhizopus oryzae Lipase in Komagataella phaffii. J. Agric. Food Chem. 2017, 65, 6009–6015. [Google Scholar] [CrossRef] [PubMed]
- Jiao, L.; Zhou, Q.; Su, Z.; Xu, L.; Yan, Y. High-level extracellular production of Rhizopus oryzae lipase in Pichia pastoris via a strategy combining optimization of gene-copy number with co-expression of ERAD-related proteins. Protein Expr. Purif. 2018, 147, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; He, X.; Li, Z.; Wang, F. Combined Strategies for Improving the Production of Recombinant Rhizopus oryzae Lipase in Pichia pastoris. Bioresour. 2013, 8, 2867–2880. [Google Scholar] [CrossRef]
- Canales, C.; Altamirano, C.; Berrios, J. Effect of dilution rate and methanol-glycerol mixed feeding on heterologous Rhizopus oryzae lipase production with Pichia pastoris Mut+ phenotype in continuous culture. Biotechnol. Prog. 2015, 31, 707–714. [Google Scholar] [CrossRef]
- Niu, W.N.; Li, Z.P.; Tan, T. Secretion of Pro- and Mature Rhizopus arrhizus Lipases by Pichia pastoris and Properties of the Proteins. Mol. Biotechnol. 2006, 32, 73–82. [Google Scholar] [CrossRef]
- Wang, J.R.; Li, Y.Y.; Xu, S.D.; Li, P.; Liu, J.S.; Liu, D.N. High-Level Expression of Pro-Form Lipase from Rhizopus oryzae in Pichia pastoris and Its Purification and Characterization. Int. J. Mol. Sci. 2013, 15, 203–217. [Google Scholar] [CrossRef]
- Barrigón, J.M.; Montesinos, J.L.; Valero, F. Searching the best operational strategies for Rhizopus oryzae lipase production in Pichia pastoris Mut+phenotype: Methanol limited or methanol non-limited fed-batch cultures? Biochem. Eng. J. 2013, 75, 47–54. [Google Scholar] [CrossRef]
- Valkonen, M.; Penttilä, M.; Saloheimo, M. Effects of inactivation and constitutive expression of the unfolded-protein response pathway on protein production in the yeast Saccharomyces cerevisiae. Appl. Environ. Microbiol. 2003, 69, 2065–2072. [Google Scholar] [CrossRef]
- Guillén, M.; Benaiges, M.D.; Valero, F. Comparison of the biochemical properties of a recombinant lipase extract from Rhizopus oryzae expressed in Pichia pastoris with a native extract. Biochem. Eng. J. 2011, 54, 117–123. [Google Scholar] [CrossRef]
- Kohno, M.; Kugimiya, W.; Hashimoto, Y.; Morita, Y. Purification, Characterization, and Crystallization of Two Types of Lipase from Rhizopus niveus. Biosci. Biotechnol. Biochem. 1994, 58, 1007–1012. [Google Scholar] [CrossRef] [PubMed]
- Pashangeh, K.; Akhond, M.; Karbalaei-Heidari, H.; Absalan, G. Biochemical characterization and stability assessment of Rhizopus oryzae lipase covalently immobilized on amino-functionalized magnetic nanoparticles. Int. J. Biol. Macromol. 2017, 105, 300–307. [Google Scholar] [CrossRef] [PubMed]
- Ben Salah, A.; Sayari, A.; Verger, R.; Gargouri, Y. Kinetic studies of Rhizopus oryzae lipase using monomolecular film technique. Biochimie 2001, 83, 463–469. [Google Scholar] [CrossRef]
- Khan, F.I.; Lan, D.; Durrani, R.; Huan, W.; Zhao, Z.; Wang, Y. The Lid Domain in Lipases: Structural and Functional Determinant of Enzymatic Properties. Front. Bioeng. Biotechnol. 2017, 5, 16. [Google Scholar] [CrossRef] [PubMed]
- Barrero, J.J.; Casler, J.C.; Valero, F.; Ferrer, P.; Glick, B.S. An improved secretion signal enhances the secretion of model proteins from Pichia pastoris. Microb. Cell Factories 2018, 17, 161–174. [Google Scholar] [CrossRef]
- Cámara, E.; Albiol, J.; Ferrer, P. Droplet digital PCR-aided screening and characterization of Pichia pastoris multiple gene copy strains. Biotechnol. Bioeng. 2016, 113, 1542–1551. [Google Scholar] [CrossRef]
- Cos, O.; Serrano, A.; Montesinos, J.L.; Ferrer, P.; Cregg, J.M.; Valero, F. Combined effect of the methanol utilization (Mut) phenotype and gene dosage on recombinant protein production in Pichia pastoris fed-batch cultures. J. Biotechnol. 2005, 116, 321–335. [Google Scholar] [CrossRef]
- Ponte, X.; Montesinos-Seguí, J.L.; Valero, F. Bioprocess efficiency in Rhizopus oryzae lipase production by Pichia pastoris under the control of PAOX1 is oxygen tension dependent. Process. Biochem. 2016, 51, 1954–1963. [Google Scholar] [CrossRef]
- Guillén, M.; Benaiges, M.D.; Valero, F. Biosynthesis of ethyl butyrate by immobilized recombinant Rhizopus oryzae lipase expressed in Pichia pastoris. Biochem. Eng. J. 2012, 65, 1–9. [Google Scholar] [CrossRef]
- Diaz, P.; Prim, N.; Pastor, F.J. Direct fluorescence-based lipase activity assay. Biotech. 1999, 27, 696–700. [Google Scholar] [CrossRef]
- Cámara, E.; Landes, N.; Albiol, J.; Gasser, B.; Mattanovich, D.; Ferrer, P. Increased dosage of AOX1 promoter-regulated expression cassettes leads to transcription attenuation of the methanol metabolism in Pichia pastoris. Sci. Rep. 2017, 7, 696–700. [Google Scholar] [CrossRef] [PubMed]
- Resina, D. Expression of a Rhizopus oryzae lipase in Pichia pastoris under control of the nitrogen source-regulated formaldehyde dehydrogenase promoter. J. Biotechnol. 2004, 109, 103–113. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | rROL | proROL |
|---|---|---|
| µmax (h−1) | 0.045 ± 0.002 | 0.073 ± 0.004 |
| Final activity (AU mL−1) | 10.51 ± 1.15 | 12.38 ± 1.05 |
| YX/S (gX gMeOH−1) | 0.28 ± 0.02 | 0.335 ± 0.005 |
| YP/X (AU gX−1) | 3753 ± 240 | 5017 ± 320 |
| MLFB | MNLFB | |||||
|---|---|---|---|---|---|---|
| Parameter | proROL 0.015 h−1 | rROL 0.015 h−1 | proROL 0.045 h−1 | rROL 0.045 h−1 | proROL 3 gL−1 | rROL 3 gL−1 |
| Final activity (AU mL−1) | 219 | 135 | 147 | 27 | 358 | 280 |
| YP/X (total AU total gX−1) | 5264 | 2644 | 1908 | 479 | 4972 | 5282 |
| µ (h−1) | 0.011 | 0.014 | 0.038 | 0.043 | 0.065 | 0.046 |
| qp (AU gX−1 h−1) | 57 | 46 | 68.5 | 18 | 308 | 322 |
| Specific productivity * (AU gX−1 h−1) | 49 | 36 | 44 | 11 | 99 | 102 |
| Volumetric productivity * (AU L−1 h−1) | 2763 | 1857 | 2782 | 623 | 7160 | 5406 |
| Enzyme | Specific Activity (AU genzyme−1) | %Protein/Lyophilized Powder | % Lipase/Protein |
|---|---|---|---|
| rROL | 17.8 ± 0.6 | 7.2 | 28.5 |
| proROL | 16.1 ± 0.3 | 8.5 | 25.2 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
López-Fernández, J.; Barrero, J.J.; Benaiges, M.D.; Valero, F. Truncated Prosequence of Rhizopus oryzae Lipase: Key Factor for Production Improvement and Biocatalyst Stability. Catalysts 2019, 9, 961. https://doi.org/10.3390/catal9110961
López-Fernández J, Barrero JJ, Benaiges MD, Valero F. Truncated Prosequence of Rhizopus oryzae Lipase: Key Factor for Production Improvement and Biocatalyst Stability. Catalysts. 2019; 9(11):961. https://doi.org/10.3390/catal9110961
Chicago/Turabian StyleLópez-Fernández, Josu, Juan J. Barrero, M. Dolors Benaiges, and Francisco Valero. 2019. "Truncated Prosequence of Rhizopus oryzae Lipase: Key Factor for Production Improvement and Biocatalyst Stability" Catalysts 9, no. 11: 961. https://doi.org/10.3390/catal9110961
APA StyleLópez-Fernández, J., Barrero, J. J., Benaiges, M. D., & Valero, F. (2019). Truncated Prosequence of Rhizopus oryzae Lipase: Key Factor for Production Improvement and Biocatalyst Stability. Catalysts, 9(11), 961. https://doi.org/10.3390/catal9110961