
Efficient Production of (R)-3-Aminobutyric Acid by Biotransformation of Recombinant E. coli

 ,
,  , , and
, , and
Abstract

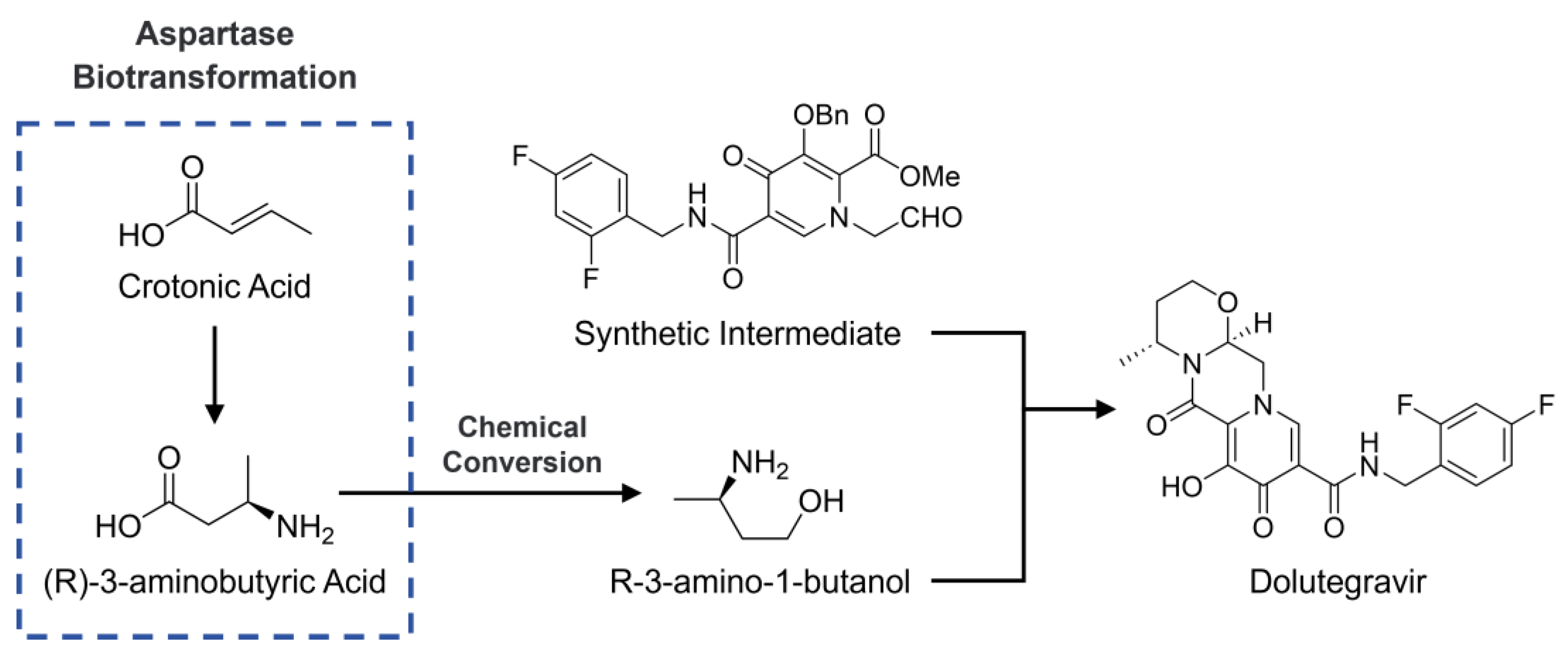
1. Introduction
2. Results and Discussion
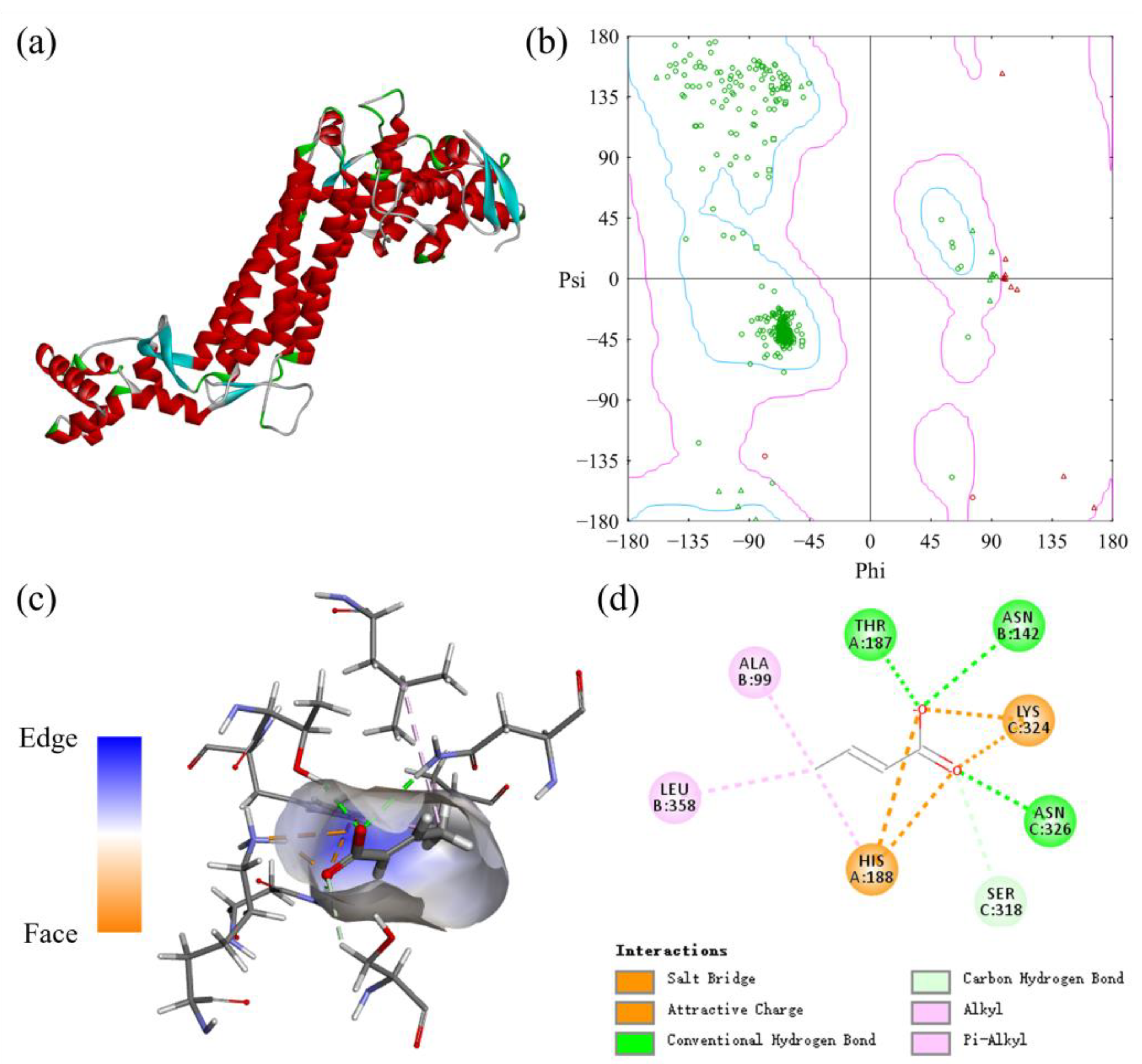
2.1. Structure and Catalytic Mechanism Analysis of Aspartase
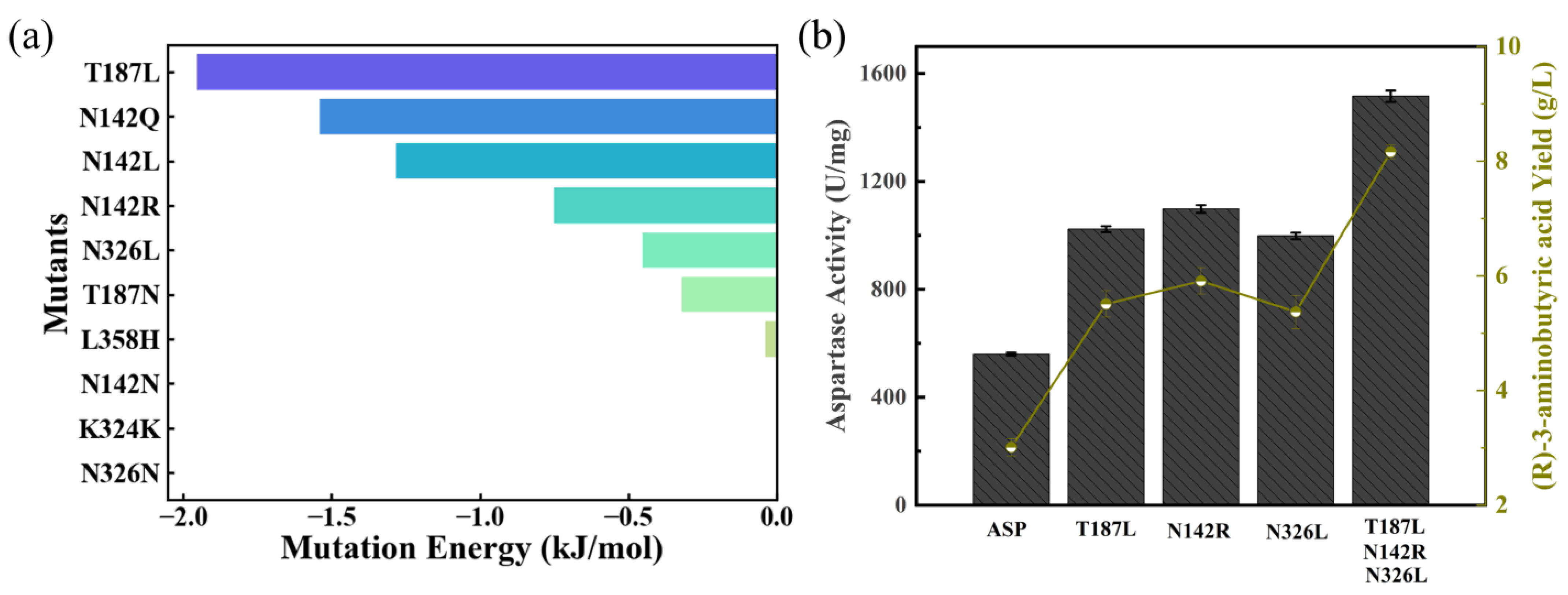
2.2. Rational Design of Aspartase to Enhance Catalytic Activity
2.3. Catalysis of (R)-3-Aminobutyric Acid Production by Engineered Aspartase Mutant
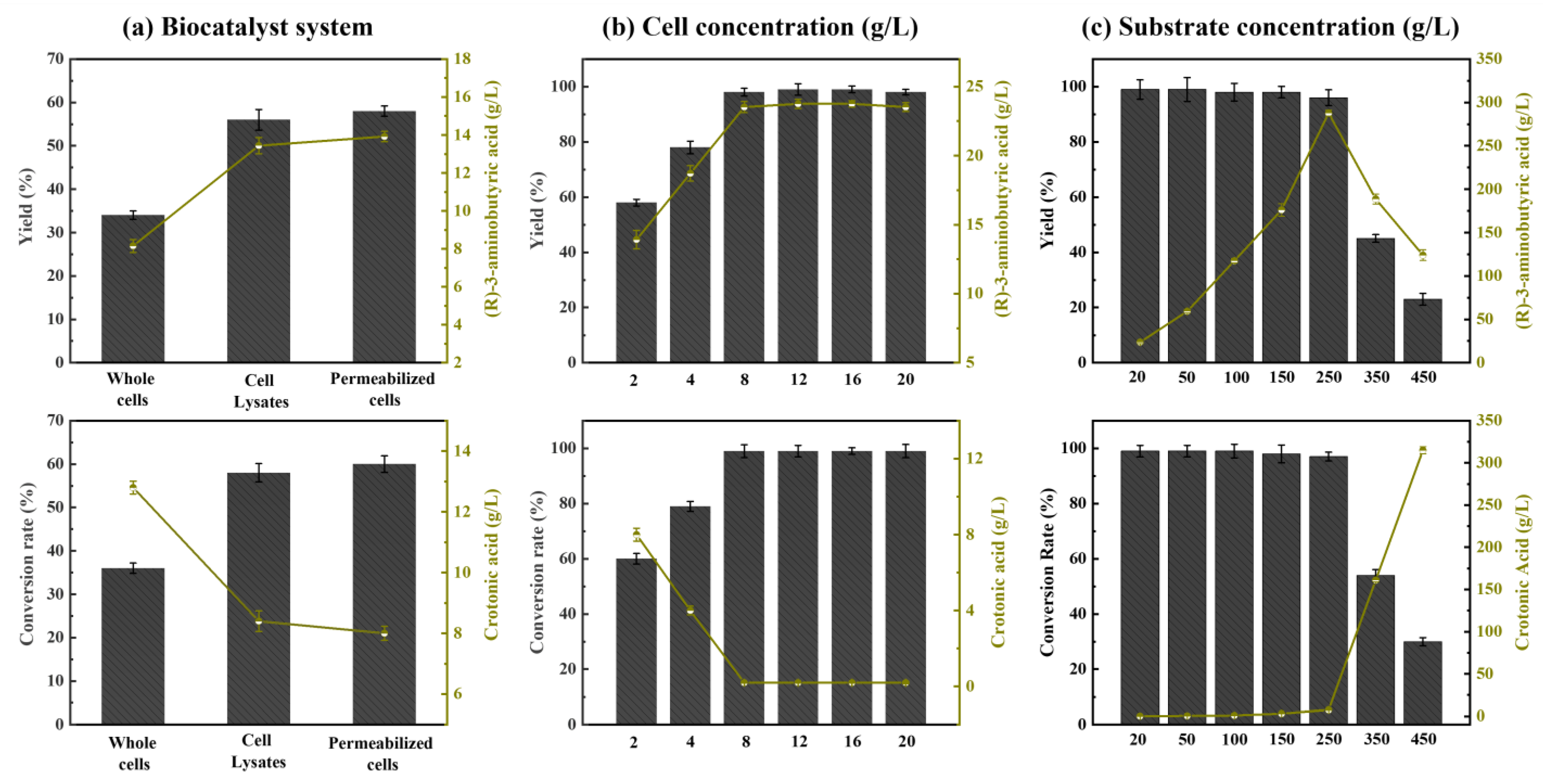
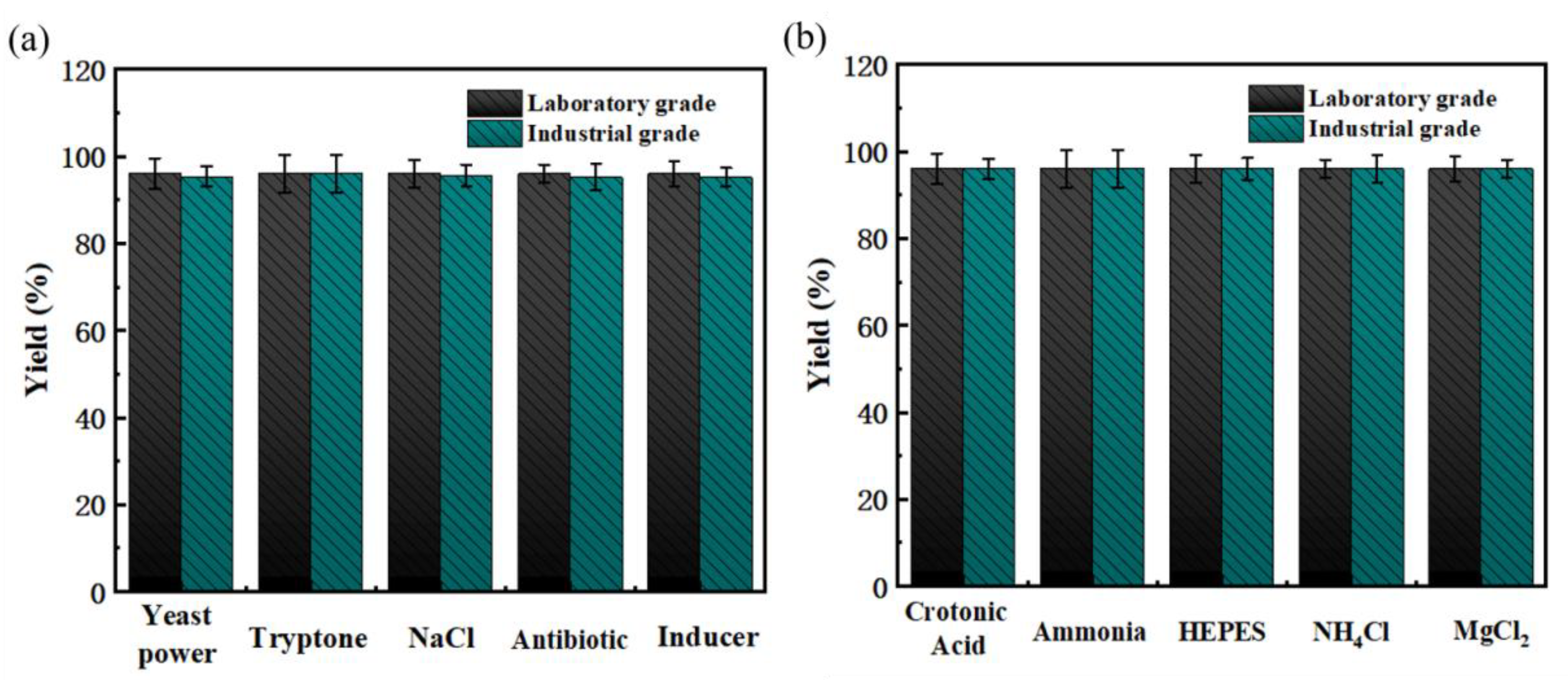
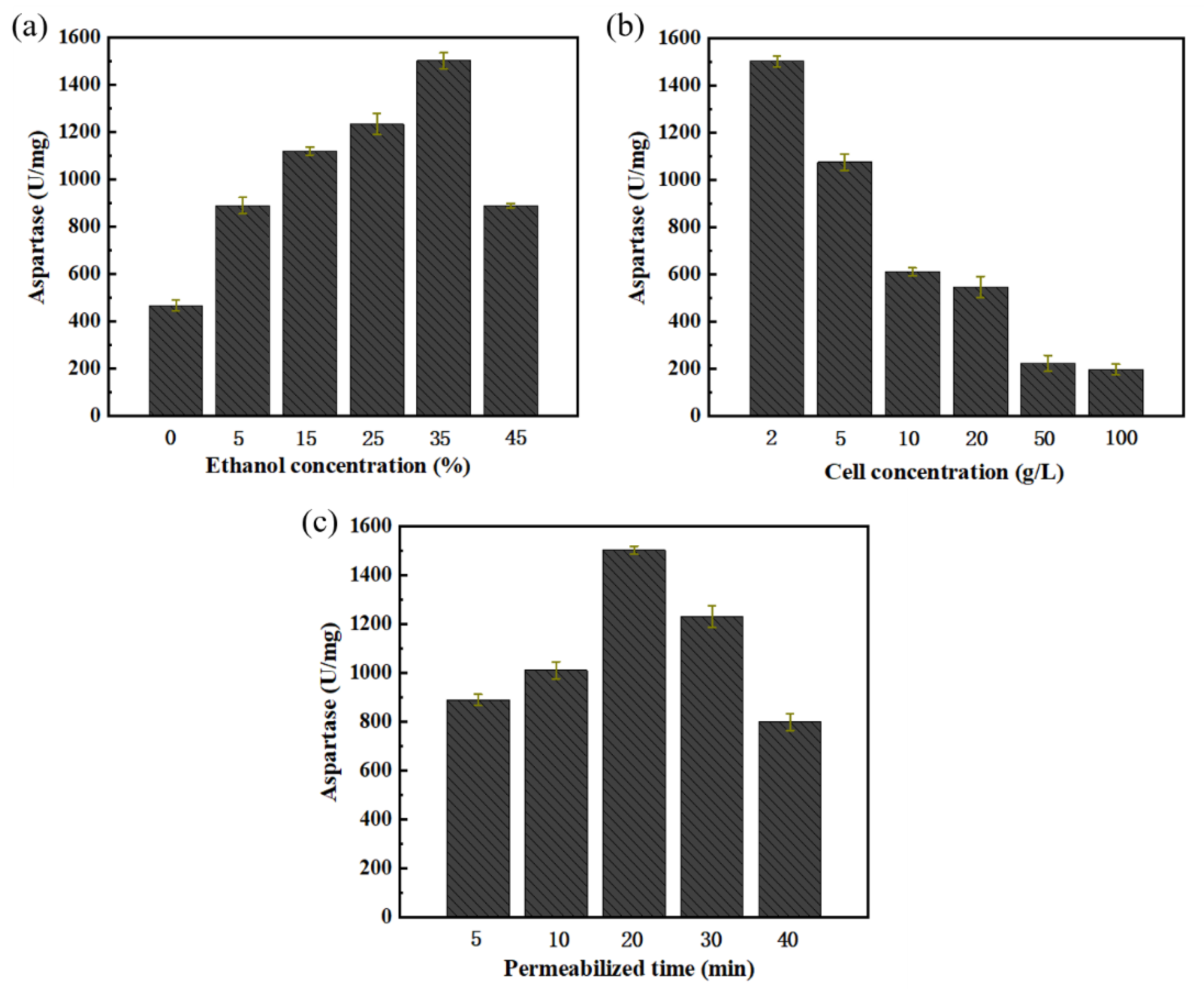
2.4. Optimization of Biocatalytic Process and Industrial Raw Material Substitution
2.5. High-Efficiency and Low-Cost (R)-3-Aminobutyric Acid Production in the Liter-Scale Integrated Strategy
3. Materials and Methods
3.1. Strains, Media, and Reagents
3.2. Rational Design of Aspartase
3.3. Construction of Engineered E. coli Strain for (R)-3-Aminobutyric Acid Production
3.4. Cultivation in Shaken Flasks
3.5. Preparation of Biocatalyst
3.6. Biocatalytic Production of (R)-3-Aminobutyric in Shaken Flasks
3.7. Optimization of Catalytic Reaction Conditions
3.8. Replacement of Industrial Materials
3.9. Product Preparation in Small-Scale Process
3.10. Activity Assay
3.11. Product Detection Method
3.12. Data Processing and Statistical Validation
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| LB | Luria–Bertani |
| IPTG | Isopropyl-beta-D-thiogalactopyranoside |
| AIDS | Acquired immune deficiency syndrome |
References
- Mbhele, N.; Chimukangara, B.; Gordon, M. HIV-1 integrase strand transfer inhibitors: A review of current drugs, recent advances and drug resistance. Int. J. Antimicrob. Agents 2021, 57, 106343. [Google Scholar] [CrossRef]
- Dow, D.E.; Bartlett, J.A. Dolutegravir, the Second-Generation of Integrase Strand Transfer Inhibitors (INSTIs) for the Treatment of HIV. Infect. Dis. Ther. 2014, 3, 83–102. [Google Scholar] [CrossRef]
- Cook, N.J.; Li, W.; Berta, D.; Badaoui, M.; Ballandras-Colas, A.; Nans, A.; Kotecha, A.; Rosta, E.; Engelman, A.N.; Cherepanov, P. Structural basis of second-generation HIV integrase inhibitor action and viral resistance. Science 2020, 367, 806–810. [Google Scholar] [CrossRef] [PubMed]
- Gandhi, R.T.; Landovitz, R.J.; Sax, P.E.; Smith, D.M.; Springer, S.A.; Günthard, H.F.; Thompson, M.A.; Bedimo, R.J.; Benson, C.A.; Buchbinder, S.P.; et al. Antiretroviral Drugs for Treatment and Prevention of HIV in Adults: 2024 Recommendations of the International Antiviral Society–USA Panel. JAMA 2025, 333, 609–628. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Ran, X.; Qiang, F.; Xiaojing, S.; Yang, H.; Xiaoli, D.; Li, T. A preliminary study on plasma concentration, short-term efficacy, and safety profile of dolutegravir in Chinese people with HIV. Expert Rev. Clin. Pharmacol. 2024, 17, 625–631. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.-M.; Alharbi, N.S.; Sun, B.; Shantharam, C.S.; Rakesh, K.P.; Qin, H.-L. Synthetic routes and structure-activity relationships (SAR) of anti-HIV agents: A key review. Eur. J. Med. Chem. 2019, 181, 111566. [Google Scholar] [CrossRef]
- Slagman, S.; Fessner, W.-D. Biocatalytic routes to anti-viral agents and their synthetic intermediates. Chem. Soc. Rev. 2021, 50, 1968–2009. [Google Scholar] [CrossRef]
- Hughes, D.L. Review of Synthetic Routes and Final Forms of Integrase Inhibitors Dolutegravir, Cabotegravir, and Bictegravir. Org. Process Res. Dev. 2019, 23, 716–729. [Google Scholar] [CrossRef]
- Wang, Y.; Xu, M.; Yang, T.; Zhang, X.; Rao, Z. Surface charge-based rational design of aspartase modifies the optimal pH for efficient β-aminobutyric acid production. Int. J. Biol. Macromol. 2020, 164, 4165–4172. [Google Scholar] [CrossRef]
- Jefford, C.W.; Wang, J. An enantiospecific synthesis of β-amino acids. Tetrahedron Lett. 1993, 34, 1111–1114. [Google Scholar] [CrossRef]
- Weiß, M.; Brinkmann, T.; Gröger, H. Towards a greener synthesis of (S)-3-aminobutanoic acid: Process development and environmental assessment. Green Chem. 2010, 12, 1580–1588. [Google Scholar] [CrossRef]
- Eissen, M.; Weiß, M.; Brinkmann, T.; Steinigeweg, S. Comparison of Two Alternative Routes to an Enantiomerically Pure β-Amino Acid. Chem. Eng. Technol. 2010, 33, 629–637. [Google Scholar] [CrossRef]
- Santaniello, E.; Ferraboschi, P.; Grisenti, P.; Manzocchi, A. The biocatalytic approach to the preparation of enantiomerically pure chiral building blocks. Chem. Rev. 1992, 92, 1071–1140. [Google Scholar] [CrossRef]
- Wohlgemuth, R. Asymmetric biocatalysis with microbial enzymes and cells. Curr. Opin. Microbiol. 2010, 13, 283–292. [Google Scholar] [CrossRef]
- Bhardwaj, K.K.; Gupta, R. Synthesis of Chirally Pure Enantiomers by Lipase. J. Oleo Sci. 2017, 66, 1073–1084. [Google Scholar] [CrossRef]
- Klotz, S.; Kaufmann, N.; Kuenz, A.; Prüße, U. Biotechnological production of enantiomerically pure d-lactic acid. Appl. Microbiol. Biotechnol. 2016, 100, 9423–9437. [Google Scholar] [CrossRef] [PubMed]
- Ren, Q.; Ruth, K.; Thöny-Meyer, L.; Zinn, M. Enatiomerically pure hydroxycarboxylic acids: Current approaches and future perspectives. Appl. Microbiol. Biotechnol. 2010, 87, 41–52. [Google Scholar] [CrossRef] [PubMed]
- Xue, Y.-P.; Cao, C.-H.; Zheng, Y.-G. Enzymatic asymmetric synthesis of chiral amino acids. Chem. Soc. Rev. 2018, 47, 1516–1561. [Google Scholar] [CrossRef]
- Li, R.; Wijma, H.J.; Song, L.; Cui, Y.; Otzen, M.; Tian, Y.e.; Du, J.; Li, T.; Niu, D.; Chen, Y.; et al. Computational redesign of enzymes for regio- and enantioselective hydroamination. Nat. Chem. Biol. 2018, 14, 664–670. [Google Scholar] [CrossRef]
- Liu, M.; Sibi, M.P. Recent advances in the stereoselective synthesis of β-amino acids. Tetrahedron 2002, 58, 7991–8035. [Google Scholar] [CrossRef]
- Kim, J.; Kyung, D.; Yun, H.; Cho, B.-K.; Seo, J.-H.; Cha, M.; Kim, B.-G. Cloning and Characterization of a Novel β-Transaminase from Mesorhizobium sp. Strain LUK: A New Biocatalyst for the Synthesis of Enantiomerically Pure β-Amino Acids. Appl. Environ. Microbiol. 2007, 73, 1772–1782. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Liu, Y. A QM/MM study of the catalytic mechanism of aspartate ammonia lyase. J. Mol. Graph. Model. 2014, 51, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Puthan Veetil, V.; Fibriansah, G.; Raj, H.; Thunnissen, A.-M.W.H.; Poelarends, G.J. Aspartase/Fumarase Superfamily: A Common Catalytic Strategy Involving General Base-Catalyzed Formation of a Highly Stabilized aci-Carboxylate Intermediate. Biochemistry 2012, 51, 4237–4243. [Google Scholar] [CrossRef] [PubMed]
- Fujii, T.; Sakai, H.; Kawata, Y.; Hata, Y. Crystal Structure of Thermostable Aspartase from Bacillus sp. YM55-1: Structure-based Exploration of Functional Sites in the Aspartase Family. J. Mol. Biol. 2003, 328, 635–654. [Google Scholar] [CrossRef]
- Kawata, Y.; Tamura, K.; Kawamura, M.; Ikei, K.; Mizobata, T.; Nagai, J.; Fujita, M.; Yano, S.; Tokushige, M.; Yumoto, N. Cloning and over-expression of thermostable Bacillus sp. YM55-1 aspartase and site-directed mutagenesis for probing a catalytic residue. Eur. J. Biochem. 2000, 267, 1847–1857. [Google Scholar] [CrossRef]
- Fibriansah, G.; Veetil, V.P.; Poelarends, G.J.; Thunnissen, A.-M.W.H. Structural Basis for the Catalytic Mechanism of Aspartate Ammonia Lyase. Biochemistry 2011, 50, 6053–6062. [Google Scholar] [CrossRef]
- Puthan Veetil, V.; Raj, H.; Quax, W.J.; Janssen, D.B.; Poelarends, G.J. Site-directed mutagenesis, kinetic and inhibition studies of aspartate ammonia lyase from Bacillus sp. YM55-1. FEBS J. 2009, 276, 2994–3007. [Google Scholar] [CrossRef]
- de Villiers, M.; Puthan Veetil, V.; Raj, H.; de Villiers, J.; Poelarends, G.J. Catalytic Mechanisms and Biocatalytic Applications of Aspartate and Methylaspartate Ammonia Lyases. ACS Chem. Biol. 2012, 7, 1618–1628. [Google Scholar] [CrossRef]
- Zhou, L.; Tao, C.; Shen, X.; Sun, X.; Wang, J.; Yuan, Q. Unlocking the potential of enzyme engineering via rational computational design strategies. Biotechnol. Adv. 2024, 73, 108376. [Google Scholar] [CrossRef]
- Song, Z.; Zhang, Q.; Wu, W.; Pu, Z.; Yu, H. Rational design of enzyme activity and enantioselectivity. Front. Bioeng. Biotechnol. 2023, 11, 1129149. [Google Scholar] [CrossRef]
- Ding, Y.; Perez-Ortiz, G.; Peate, J.; Barry, S.M. Redesigning Enzymes for Biocatalysis: Exploiting Structural Understanding for Improved Selectivity. Front. Mol. Biosci. 2022, 9, 908285. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, P.; Fernandes, P.A.; Ramos, M.J. Modern computational methods for rational enzyme engineering. Chem Catal. 2022, 2, 2481–2498. [Google Scholar] [CrossRef]
- Lutz, S. Beyond directed evolution—Semi-rational protein engineering and design. Curr. Opin. Biotechnol. 2010, 21, 734–743. [Google Scholar] [CrossRef] [PubMed]
- Ding, J.; You, S.; Zhang, J.; Zhang, H.; Wang, H.; Zhang, W.; Qi, W.; Su, R.; He, Z. Rational design of 17β-hydroxysteroid dehydrogenase type3 for improving testosterone production with an engineered Pichia pastoris. Bioresour. Technol. 2021, 341, 125833. [Google Scholar] [CrossRef] [PubMed]
- Gran-Scheuch, A.; Wijma, H.J.; Capra, N.; van Beek, H.L.; Trajkovic, M.; Baldenius, K.; Breuer, M.; Thunnissen, A.-M.W.H.; Janssen, D.B. Bioinformatics and Computationally Supported Redesign of Aspartase for β-Alanine Synthesis by Acrylic Acid Hydroamination. ACS Catal. 2025, 15, 928–938. [Google Scholar] [CrossRef]
- Asano, Y.; Kira, I.; Yokozeki, K. Alteration of substrate specificity of aspartase by directed evolution. Biomol. Eng. 2005, 22, 95–101. [Google Scholar] [CrossRef]
- Sun, C.; Cheng, Z.; Jiao, J.; Ding, S.; Tian, Z. Method for Enzymatic Preparation of R-3-Aminobutyric. Acid. Patent WO2019062222A1, 4 April 2019. [Google Scholar]
- Vogel, A.; Schmiedel, R.; Hofmann, U.; Gruber, K.; Zangger, K. Converting Aspartase into a β-Amino Acid Lyase by Cluster Screening. ChemCatChem 2014, 6, 965–968. [Google Scholar] [CrossRef]
- Lin, W.; Zheng, Y.; Zhang, J.; Zhou, Y.; Wang, M.; You, S.; Su, R.; Qi, W. Enhanced catalytic activity of polyethylene terephthalate hydrolase by structure-guided loop-focused iterative mutagenesis strategy. J. Hazard. Mater. 2025, 490, 137837. [Google Scholar] [CrossRef]
- Zheng, Y.; Zhang, J.; You, S.; Lin, W.; Su, R.; Qi, W. Efficient thermophilic polyethylene terephthalate hydrolase enhanced by cross correlation-based accumulated mutagenesis strategy. Bioresour. Technol. 2024, 406, 130929. [Google Scholar] [CrossRef]
- Falzone, C.J.; Karsten, W.E.; Conley, J.D.; Viola, R.E. L-Aspartase from Escherichia coli: Substrate specificity and role of divalent metal ions. Biochemistry 1988, 27, 9089–9093. [Google Scholar] [CrossRef]
- Talukder, M.M.R.; Min, P.S.; Jae, C.W. Integration of cell permeabilization and medium engineering for enhanced enantioselective synthesis of ethyl-S-3-hydroxy-3-phenylpropanoate (S-EHPP). Biochem. Eng. J. 2019, 148, 24–28. [Google Scholar] [CrossRef]
- Catania, C.; Ajo-Franklin, C.M.; Bazan, G.C. Membrane permeabilization by conjugated oligoelectrolytes accelerates whole-cell catalysis. RSC Adv. 2016, 6, 100300–100306. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fermentation | Bioproduction by Permeabilized Cells | ||||
|---|---|---|---|---|---|
| Type | OD600 | Aspartase (U/mg) | Reaction Time (h) | (R)-3-Aminobutyric Acid (g/L) | Yield (%) |
| Lab scale (Reagents) | 4.43 ± 0.37 | 1516 ± 19 | 24 | 287 ± 1.25 | 95.9 ± 0.55 |
| Lab scale (Industrial materials) | 4.35 ± 0.21 | 1503 ± 12 | 24 | 281 ± 0.65 | 93.9 ± 0.25 |
| Bioreactor scale (Industrial materials) | 4.29 ± 0.21 | 1496 ± 16 | 48 | 215 ± 0.94 | 71.8 ± 0.42 |
| Bioreactor scale after the process optimization (Industrial materials) | 4.32 ± 0.28 | 1502 ± 23 | 24 | 284 ± 1.07 | 94.9 ± 0.87 |
| Reagents | Industrial Materials | ||
|---|---|---|---|
| Component | Cost (¥) | Component | Cost (¥) |
| Tryptone (35 g) | 17.50 | Tryptone (35 g) | 0.35 |
| Yeast extract (17.5 g) | 4.55 | Yeast extract (17.5 g) | 0.16 |
| Sodium chloride (35 g) | 0.70 | Sodium chloride (35 g) | 0.02 |
| Seed fermentation broth (100 mL) | 0.65 | Seed fermentation broth (100 mL) | 0.02 |
| Resistance (0.35 mL) | 3.96 | Resistance (0.35 mL) | 0.14 |
| IPTG (0.35 mL) | 9.8 | IPTG (0.35 mL) | 2.4 |
| 35% ethanol (3.5 L) | 36.75 | 35% ethanol (3.5 L) | 12.01 |
| Butenoic acid (225 g) | 42.3 | Butenoic acid (225 g) | 13.95 |
| Ammonia (230 mL) | 2.34 | Ammonia (230 mL) | 0.525 |
| HEPES buffer (0.9 L) | 1.21 | HEPES buffer (0.9 L) | 0.05 |
| MgCl2 (2 mL) | 0.05 | MgCl2 (2 mL) | 0.01 |
| NH4Cl (2 mL) | 2.9 | NH4Cl (2 mL) | 0.12 |
| Total | 122.71 | Total | 29.72 |
| (R)-3-aminobutyric acid (1 kg) | 479.76 | (R)-3-aminobutyric acid (1 kg) | 116.21 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, H.; Xu, Q.; Lv, J.; Zhang, J.; Dou, T.; You, S.; Su, R.; Qi, W. Efficient Production of (R)-3-Aminobutyric Acid by Biotransformation of Recombinant E. coli. Catalysts 2025, 15, 466. https://doi.org/10.3390/catal15050466
Zhang H, Xu Q, Lv J, Zhang J, Dou T, You S, Su R, Qi W. Efficient Production of (R)-3-Aminobutyric Acid by Biotransformation of Recombinant E. coli. Catalysts. 2025; 15(5):466. https://doi.org/10.3390/catal15050466
Chicago/Turabian StyleZhang, Hongtao, Qing Xu, Jiajia Lv, Jiaxing Zhang, Tongyi Dou, Shengping You, Rongxin Su, and Wei Qi. 2025. "Efficient Production of (R)-3-Aminobutyric Acid by Biotransformation of Recombinant E. coli" Catalysts 15, no. 5: 466. https://doi.org/10.3390/catal15050466
APA StyleZhang, H., Xu, Q., Lv, J., Zhang, J., Dou, T., You, S., Su, R., & Qi, W. (2025). Efficient Production of (R)-3-Aminobutyric Acid by Biotransformation of Recombinant E. coli. Catalysts, 15(5), 466. https://doi.org/10.3390/catal15050466