Abstract
Chiral compounds are valuable industrial products and intermediates, and the production of chemicals with high enantiopurity is one of the major objects in asymmetric catalysis. Compared with traditional chemical synthesis, enzymatic synthesis can produce chiral molecules under sustainable conditions which are much greener, more economical, and more environmentally friendly. The superfamily of α/β hydrolases includes a lot of diverse enzymes showing excellent chemo-, regio-, and enantio-selectivity in asymmetric synthesis and many of them are biocatalysts in industry. This review outlines the current knowledge of the structures and reaction mechanism of α/β hydrolases and summarizes the screening and protein engineering efforts to develop biocatalysts for chiral chemicals production in recent years. Other strategies such as whole-cell catalysis and protein immobilization to improve the performance of α/β hydrolases are also discussed. The progress in biocatalyst development based on α/β hydrolases will promote the biosynthesis of chiral compounds, thus contributing to the green and sustainable development of the chemical and pharmaceutical industry.
1. Introduction
Chiral chemicals are important stocks in the manufacturing industry of pharmaceuticals, foods, and materials [1]. Chiral chemicals of different enantiomers can be resolved by gas or liquid chromatography [2,3,4], capillary electrophoresis [5], spectroscopy [6,7], mass spectrometry [8], and sensor recognition [9,10]. Many pharmaceuticals are chiral compounds, and their different enantiomers usually show different physiological activities [11,12,13]. Although the structures of enantiomeric drugs are remarkably similar, in many cases only one enantiomer shows a beneficial effect, while the other has no effect or even is harmful. Therefore, enantiomeric pure chiral drugs used in clinical treatment can eliminate the negative side effects of the undesirable chiral partner, reduce the drug dose and metabolic burden of the human body, and improve medicine potency. The demands on the synthesis of chiral compounds with high enantiopurity are continuously increasing and the preparation methods for high enantiopurity chemicals are the major topics in asymmetric catalysis [12,13].
The family of α/β hydrolases is one of the largest known protein families, and many of them are widely used as biocatalysts in the synthesis of chiral chemicals [14]. Epoxide hydrolase (EH; E.C.3.3.2.x), for example, can catalyze the kinetic resolution and enantio-convergent hydrolysis of racemic epoxides to prepare enantiopure functional alcohols [15], which are important building blocks for optically active drug synthesis. In addition, optically pure (S)-2-pentanol prepared by haloalkane dehalogenase (HLD, EC 3.8.1.5) is an intermediate for the synthesis of potential anti-Alzheimer’s disease drugs [16]. With more and more enzymes discovered and identified from nature, enzymatic preparation of chiral compounds offers a useful, more economical, and environmentally friendly alternative to traditional chemical approaches [17].
Natural enzymes offer many advantages over traditional chemical catalysts and are increasingly used in the synthetic process of chiral compounds, but their properties such as activity, stability, and enantioselectivity often need to be improved for specific industrial processes. Recent developments in protein engineering techniques have provided efficient approaches to tailor enzymes and overcome the limitations of natural biocatalysts, and more improved or novel enzymes required in many industrial applications are obtained by directed evolution and rational design.
In this review, we focused on the applications of α/β hydrolases in producing enantiomerically pure chemicals. Although α/β hydrolases are very diverse and contain many important enzymes with various functions, we mainly reviewed the studies on several groups of α/β hydrolases including epoxide hydrolases, dehalogenases, hydroxynitrile lyases, and fungal lipases, which are widely used in chiral chemical production in scientific research and industrial processes. We discussed the high-throughput screening methods for novel enzymes and recent advances in protein engineering approach applied to generate desired variants, and summarized other effective strategies such as whole-cell catalysis and immobilization for expanding the practical usefulness of this enzyme family. It should be noted that this short review is intended to be broad and can therefore not be complete in the sense that all relevant work is mentioned.
2. Structure and Catalytic Mechanism of α/β Hydrolase
The family of α/β hydrolases is one of the largest known protein families, including a wide range of members such as epoxide hydrolases, dehalogenases, hydroxynitrile lyases, fungal lipases, amidases, dienelactone hydrolases, haloperoxidases, acetylcholine esterases, serine carboxypeptidases, serine carboxypeptidase-like acyltransferases and other enzymes with distinct functions. These enzymes have a characteristic conserved structure which consists of eight β-strands connected by α-helixes and folds into a mostly parallel central β-sheet surrounded by α-helixes [14]. The active sites of these enzymes usually contain a catalytic triad composed of three conserved residues in loops, including a nucleophile (serine, aspartate, or cysteine), a histidine, and a catalytic acid (aspartate or glutamate). Many α/β hydrolases also contain lid/cap domains located on top of the active site, and these domains are not conserved and vary considerably between different enzymes (Figure 1) [15,18]. The lid/cap domain also affects the properties of these enzymes. For example, in haloalkane dehalogenase, the conformational change of the cap domain is considered to be a key step to allow water to enter and solvate the halide ion [19]. In prolyl oligopeptidase, it is suggested that the lid domain helps to exclude bulk solvents [20]. Some studies revealed that the mutations in the lid/cap domains influence the substrate specificity of the enzymes [14,21].
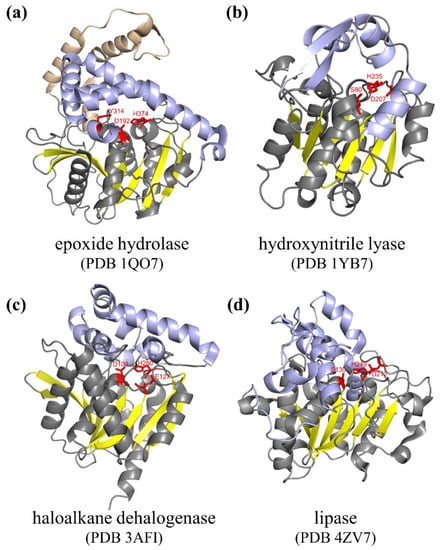
Figure 1.
Structures of α/β hydrolases. (a) Aspergillus niger epoxide hydrolase [22]. (b) Hevea brasiliensis hydroxynitrile lyase [23]. (c) haloalkane dehalogenase DbjA [24]. (d) Candida antarctica lipase B [25]. The central β-sheet, flank helices, the cap domain, and the N-terminal meander are shown in yellow, gray, slateblue, and brown, respectively. The active site residues are shown as red sticks.
Members of α/β hydrolases catalyze a broad variety of chemical reactions with substrates containing diverse functional groups, but all of these reactions utilize the common mechanism of nucleophilic catalysis because of their highly similar three-dimensional structures and active site residues. The mechanism of substrate selectivity, regioselectivity, and stereoselectivity of α/β hydrolases are diverse and case-by-case depending on the structures of these enzymes. The catalytic mechanism was illustrated by Aspergillus niger epoxide hydrolase (AnEH) (Figure 2) [22]. The substrate-binding cavity is between the α/β-hydrolase fold domain and the lid domain, and the substrate is located at the enzyme active center by the hydrogen bonds between the oxirane ring and the hydroxyl groups of Tyr251 and Tyr314 on the cap domain. The catalytic triad is constituted of two aspartates and one histidine on the loops of the α/β-hydrolase fold domain. The reaction is catalyzed through a two-step mechanism. In the first step, a nucleophilic attack of an aspartic residue toward the oxirane ring forms a covalent ester intermediate with the ring opened by the classical push-pull mechanism. In the second step, the covalent intermediate is hydrolyzed to release the final product by a water molecule in which the proton is extracted by the His374/Asp348 charge relay system (Figure 2).
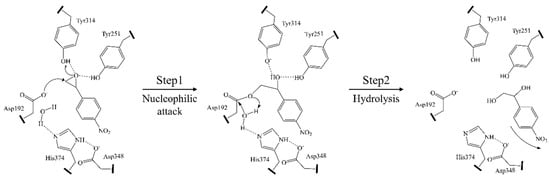
Figure 2.
Mechanism of a typical α/β hydrolase, 4-nitrostyrene oxide hydrolase AnEH. Reprinted with permission from Ref. [22]. Copyright 2000, Elsevier.
3. Regio- and Stereo-Selectivity of α/β Hydrolases
Enzymes usually show significant regio- and/or stereo-selectivity. Although various mechanisms for selectivity may exist in different enzymes, the conserved catalytic mechanism of α/β hydrolases implies that their regio- and stereo-selectivity is achieved by a similar mechanism determined by the position of the substrate and the structure of the active site. This feature makes the engineering of regio- and stereo-selectivity of α/β hydrolases feasible by rational design based on high-resolution structures. Different types of α/β hydrolases showed different substrate preferences and their regio- and stereo-selectivity has been extensively studied.
In epoxide hydrolases, different substituents in epoxide substrate will affect the regioselectivity and direct different nucleophilic attacking sites in the ring-opening reaction, so that the enzyme can selectively attack one of the two carbons on the epoxy ring [26]. During ring opening of alkyl-substituted epoxides, electron-withdrawing groups usually direct the nucleophilic attack to the unsubstituted carbon with the least steric hindrance. In contrast, the reaction of electron-donating substituents (such as phenyl) can help to change the preferred attack site by stabilizing the positive charge on the attacked epoxide carbon, so that a ring-opening reaction can take place at the substituted carbon. In addition to the difference in substituents, space limitations of binding sites and environmental factors such as pH and temperature can also affect the selectivity of enzymes [27,28].
Regioselectivity of lipases contains two types according to the catalyzed carbon positions of the glycerol backbone of a triglyceride molecule [29,30]. Lang et al. reported a crystallographic study of the complex of a Pseudomonas cepacia lipase and a triglyceride analog. Four triacylglycerol binding pockets including an oxyanion hole and three pockets for accommodating different fatty acid chains were detected, and the different sizes of pockets and the hydrophobic/hydrophilic ratios can affect the regioselectivity of the lipase [31]. A study using chemical modification and molecular dynamics simulations for P. cepacia lipase indicated that the enantioselectivity of the lipase depends on the hydrogen bond between the Onon-α (oxygen at non-α-position of the acyloxy group) of the substrates and Tyr29 located in the catalytic cavity [32]. Furthermore, environmental factors such as the polarity and temperature of the reaction system can also influence the kinetic resolution of primary and secondary alcohols [33,34,35]. A recent study revealed that the key regions in lipase that affect the kinetic resolution depend on the type of chiral substrates. When carboxylic acids are substrates, the pivotal areas are acyl-binding pockets, the hydrophobic cavity, and the oxyanion hole, while the hydrophilic cavity, the hydrophobic cavity, and the oxyanion hole play key roles in enantioselectivity for the substrates of chiral alcohols [36].
Studies on haloalkane dehalogenase DbjA showed that the enzyme could kinetically distinguish the different substrate enantiomers α-bromoester and β-bromoalkane, and the reaction enantioselectivity was controlled by different molecular bases [24]. Methyl 2-bromobutyrate as a representative substrate for α-bromoesters has different orientations for the R and S enantiomers, which are stabilized by different hydrogen bonds with the side chain of Asn38 or Trp104 in the active site, and these differences favor binding of the R enantiomer over the S enantiomer. Differently, both enantiomers of 2-bromopentane as a representative substrate for β-bromoalkanes have similar hydrophobic interaction with the same wall of the active site, and the enantiopreference is determined by the entropy favoring the S enantiomer. Due to these different molecular bases, mutations in a surface loop can modulate the enantioselectivity differently for the two types of substrates [24]. However, another haloalkane dehalogenase DhaA does not have the same enzymatic activity towards β-bromoalkane [37]. Comparing the enzyme structures, it is found that the fragment 138-HHTEVAEQDH-150 termed ERB fragment in DbjA seems to be the reason for this difference [37]. The enantioselectivity with inverse temperature dependence can be applied to β-bromoalkanes rather than α-bromoesters by mutating the residues located in the surface ring or flank of DbjA [24]. A directed evolution study of DhaA showed that the residues around the active site can be successfully engineered to obtain enantio-complementary enzymes [38].
The FAD-independent hydroxynitrile lyases from plants are also α/β hydrolase members. They usually exist in the form of homodimer or heterodimer in solution and catalyze cyanohydrins with various (R) or (S) configurations as substrates. Studies discovered that the enzyme enantioselectivity can be adjusted by changing the structure of the binding site [14,39,40].
In summary, the stereoselectivity mechanism of enzymes is associated with both protein structures and reaction environments. The enantioselectivity can be improved or even altered by mutating amino acids located in specific regions such as the active site pocket or the entrance tunnel of the enzyme [41], while environmental conditions can also be optimized by other methods such as immobilizing enzymes on solid supports [42,43].
4. Screening of New α/β Hydrolases from Nature
The diversity of natural environment and species provides an abundant source to discover novel enzymes with regio- and stereo-selectivity. In the past decades, tremendous efforts have been made to screen and characterize new α/β hydrolases from animals, plants, bacteria, and other species.
Traditional screening methods are usually focusing on known species or available gene sequences to explore potentially valuable enzymes. Two plants derived EHs, VrEH1 and VrEH2, which can be applied to the enantiomeric convergent hydrolysis of p-nitrostyrene oxide, were successfully screened from mung bean (Vigna radiata) [44,45]. The gene of another EH VrEH3 in V. radiata was also cloned and expressed in Escherichia coli, and the recombinant VrEH3 showed different stereoselectivity towards a variety of epoxides and is considered to be an advantageous industrial biocatalyst [46]. Similarly, an EH gene Aueh2 cloned from Aspergillus usamii E001 was expressed in E. coli and the recombinant AuEH2 showed superior enzymatic properties in the resolution of racemic styrene oxide [47]. Zhang et al. screened a new EH enzyme GmEH3 from soybean (Glycine max) using the computer-aided analysis and they found that recombinant GmEH3 showed high regio-complementarity towards (S)- and (R)-m-chlorostyrene oxide and the highest enantioselectivity to rac-phenyl glycidyl ether [48]. An EH gene amEH was successfully identified from coastal wetland soil bacteria Agromyces mediolanus ZJB120203 and the recombinant AmEH showed advantages for optically active epichlorohydrin preparation [49].
In addition, metagenomics is another emerging method for discovering new enzymes recently. It is an approach based on microbial genomics, aiming at revealing microbial biodiversity and offering the potential to discover new natural functional chemicals or genes [50]. This strategy involves the isolation of metagenomic DNA directly from environmental samples, the construction of metagenomic libraries, the analysis of the genetic composition of all microorganisms in the samples by genomics research techniques, and the screening of novel physiological active products.
In metagenomics, total genetic materials from natural samples are directly studied, which is quite different from traditional microbiology relying on cultivated microorganisms. Therefore, metagenomics enables us to recognize and investigate those unknown microorganisms that cannot be, or have not been cultivated. Metagenomic libraries can be screened and analyzed for detecting information on the target enzymes in soil, ocean, and extreme environments. Previously, 29 potentials EH genes were identified by analyzing the genomes of bacteria in soil, five of which have been confirmed with EH activity. Two of these enzymes showed wide substrate scope, good regio-selectivity in the hydrolysis of styrene oxide, and high enantio-selectivity to obtain the (R)- and (S)-diols from the same enantiopure epoxide [51]. Two new thermophilic EHs were successfully identified from metagenomic samples from China and Russia and both enzymes demonstrated a wide range of substrate preferences. One of them CH65-EH can tolerate high temperatures and remain at about 80% of initial activity after 3 h heat treatment at 70 °C [52]. So far, more and more high-performance natural enzymes have been discovered and identified by metagenomics, which has enriched the protein library [53,54,55].
5. Engineering of Natural α/β Hydrolases
Although many natural enzymes have been screened as biocatalysts with excellent performance, most of them are still unable to meet the needs of industrial applications. Low catalytic activity, thermostability, and enantioselectivity under complex and harsh industrial process conditions are still the main limitations for the large-scale application of natural enzymes. With the development of protein engineering technology, functional improvements have been achieved for existing α/β hydrolases, specifically in key enzyme characteristics such as their enantioselectivity and stability, in order to tailor these enzymes for specific industrial applications (Table 1).

Table 1.
Native and engineered α/β hydrolases for chiral chemical production.
Two major approaches have been developed in protein engineering. One is directed evolution. A library of variants with random mutations in the protein-encoding gene is created through sequential error-prone PCR (epPCR) or in vitro DNA recombination, and then screening methods are used to isolate functionally improved mutant proteins for further study or the next round of improvement. Directed evolution does not rely on the knowledge of the enzyme structure and catalytic mechanism, but large libraries generated by directed evolution need to be screened for the best mutants with desired properties, which is considered to be time-consuming and laborious. The development of new efficient and convenient screening methods is the key to obtaining novel biocatalysts by directed evolution [74,75]. The second approach is the rational design which generates beneficial mutants based on protein sequences, structural information, and the relationship between enzyme structure and function. Advanced computational methods are often adopted to introduce specific mutagenesis in conserved functional domains, catalytic active sites, and certain amino acid positions for yielding remarkable desired properties (Figure 3). The combination of directed evolution and rational design strategies, named semi-rational design, with the benefits of both strategies has also been developed for improving substrate specificity, enantioselectivity, and stability.
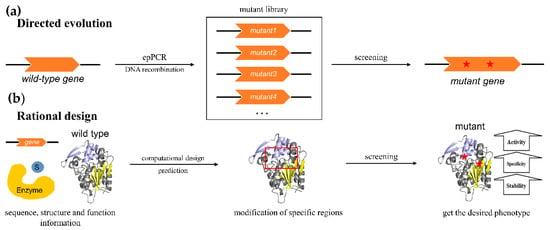
Figure 3.
Protein engineering strategies. (a) Directed evolution. (b) Rational design. “S”, substrate; red stars: mutation sites. The enzyme structure shown in the figure is hydroxynitrile lyase HbHNL from Hevea brasiliensis (PDB ID: 1YB7) [23].
5.1. Engineering of α/β Hydrolases to Catalyze Different Reactions
Many members of α/β hydrolase family show “catalytic promiscuity” and catalyze diverse chemical transformations. Enzyme promiscuity plays a key role in protein engineering because enzyme activities can be changed by only altering a few amino acid residues [76,77]. SABP2-G12T-M239K, the variant of plant esterase salicylic acid binding protein 2 (SABP2) can be converted into a hydroxynitrile lyase only with the replacement of two amino acid residues. This mutant can also catalyze reverse reactions to form mandelonitrile with low enantio-specificity [66]. Nedrud et al. substituted the active site residues of hydroxynitrile lyase HbHNL with the corresponding residues in esterase SABP2 and found key residues to generate the esterase activity [72]. On the contrary, Jochens et al. attempted to convert the esterase of Pseudomonas into epoxide hydrolase by replacing the residues of active site loops in the esterase with the residues in the epoxide hydrolases. After residues in multiple sites were carefully chosen and replaced, the mutant enzyme showed an epoxide hydrolase activity with enantioselectivity for R-type enantiomers, but it was widely inhibited by substrates and had a limited turnover number [63]. These examples provide evidence for specificity engineering by utilizing the catalytic promiscuity of enzymes, although their performance, compared with natural enzymes, still needs further improvement.
Beier et al. reported a study on haloalkane dehalogenase which provides a new way to expand the application of α/β hydrolases in chiral preparations [67]. By mutating the catalytic residue histidine and blocking the hydrolysis process, the variant DbjA H280F could catalyze a transhalogenation reaction and realize the replacement of halogen substitutes. This strategy was further demonstrated in several different haloalkane dehalogenases [67].
5.2. Catalytic Activity Enhancement of α/β Hydrolases
High catalytic activities are required for enzymes in commercial applications. It is possible to increase the activities of natural enzymes through rational design and directed evolution approaches. Kotik et al. significantly improved the activity of Aspergillus niger M200 epoxide hydrolase through single site mutagenesis of Ala217 at the substrate entrance. For example, A217C enhanced the activity of the enzyme by three times towards allyl glycidyl ether and styrene oxide, while A217V increased the activity of the enzyme to allyl glycidyl ether by six times [57]. A completely different approach was used by Yu et al. to improve the activity of Candida antarctica lipase B. A mutant cp283 obtained through circular permutation showed higher catalytic activity, compared with the wild-type enzyme in the transesterification reaction with different substrates using 1-butanol and ethyl acetate as acyl receptors [73]. Langermann et al. replaced the active site residue in hydroxynitrile lyase HbHNL with the corresponding site in esterase SABP2, thereby improving the catalytic activity of HbHNL on an unnatural substrate mandelonitrile [71]. More recently, Marek et al. transplanted a nine-residue-long extension of L9 loop and α4 helix from DbjA into the corresponding site of another haloalkane dehalogenase DbeA. The mutation not only altered its enantio-preference with several linear β-bromoalkanes but also enhanced the catalytic activity of DbeA towards various halogenated substrates [68].
5.3. Regio- and Stereo-Selectivity Engineering of α/β Hydrolases
Since the resolution process of racemates is costly and often difficult, enzymatic catalytic synthesis of enantiopure compounds has gradually become a useful alternative to various chemical preparation routes. While enantio-convergent bioconversions by natural enzymes are usually not able to meet industrial requirements, it is necessary to improve or alter the enantioselectivity of biocatalysts by protein engineering.
Substrate binding sites are frequently engineered for the improvement of regio- and stereo-selectivity because they contain the catalytic residues and determine the location of substrates during the catalysis. By mutating the residues of the atypical oxirane oxygen hole (Trp186/Trp187/Tyr247) around the active site in epoxide hydrolase Alp1U, Zhang et al. obtained mutants Y247F and W187F/Y247F with high regioselectivity, respectively, towards C-2 and C-3 of fluostatin C [56]. Recently, Wen et al. reported the improvement of the enantio-specificity of AuEH2 towards racemic ortho-trifluoromethyl styrene oxide by tuning the substrate binding pocket [60]. With a similar method, Hu et al. performed the molecular docking of AuEH2 with (R)-styrene oxide by AutoDock Vina and screened suitable sites for site-directed saturation mutagenesis and combinatorial mutagenesis. A mutant A214C/A250I exhibited a 12.6-times increase of the enantiomeric ratio towards the substate rac-styrene oxide [59]. Accompanied by the deep understanding of the relationship between the structure and activity of enzymes, more successful studies have been reported to enhance the enantioselectivity by the design and engineering of active site residues [58,61,62,78].
In addition to the active site, the entrance tunnel of the enzyme has also been targeted to modulate the selectivity. For example, Kotik et al. reported that a single amino acid mutation at the entrance site of the AnEH tunnel can notably increase its enantioselectivity [57].
5.4. Stability Enhancement of α/β Hydrolases
Enzymes with high thermostabilities are required in many industrial production processes because the elevated temperature has positive effects on many bioconversion processes such as improving the solubility of substrates, increasing the catalytic rate, and reducing microbial contaminants [79]. Through a combination of computational design and experimental screening, Floor et al. established a method named FRESCO, which based on the calculation of folding energies for all possible substitutions and the integration of conformational sampling in disulfide-bond designs, utilized the molecular dynamics as a fast-screening tool. Using this method, a multisite mutant of a large monomeric protein, haloalkane dehalogenase LinB, was obtained with drastically improved thermostability (a 23 °C increase in apparent melting temperature and an over 200-fold longer half-life at 60 °C) and retained moderate enantioselectivity [70]. Similarly, Bednar et al. established another method, FireProt, which is a computational strategy aimed at the prediction of highly stable multipoint mutations and significantly increased the thermostability of haloalkane dehalogenase DhaA [69]. Utilizing the circular permutation strategy to analyze the three characteristic regions of typical esterase RhEst1, Li et al. obtained three mutants CP-20, CP-22, and CP-24 with ameliorated thermostability [64]. A systematic study by Jones et al. used and compared five protein engineering strategies (random mutagenesis, two computational methods Rosetta and FoldX, consensus mutation, and homoproline mutation) to enhance the stability of an α/β-hydrolase fold enzyme, salicylic acid binding protein 2. The results showed that all five methods could obtain mutants with enhanced stability, but consensus mutation and homoproline mutation seemed to be the best choices. This research may provide a reference for the subsequent engineering strategy of increasing the stability of α/β hydrolases and other enzymes [65].
6. Other Progress in Biocatalyst Development Based on α/β Hydrolases
In addition to protein engineering, many other strategies have also been used to improve enzyme performance in industrial production (Figure 4).
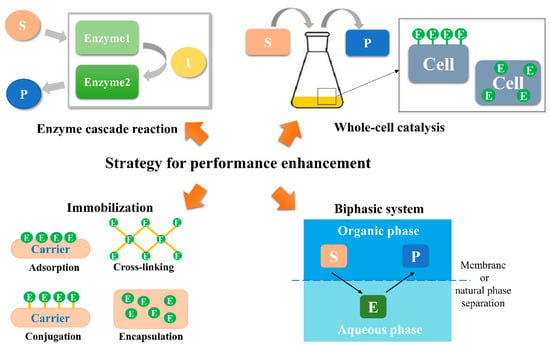
Figure 4.
Strategies for performance enhancement based on α/β hydrolases. “S”, substrate; “I”, intermediate; “P”, product; “E”, enzyme.
Inspired by natural multi-enzymatic metabolic pathways, a cascade biocatalytic system combining multiple catalytic steps in one pot provides a more economical and efficient technique. In recent years, more and more cascaded biocatalytic processes have been developed with multiple enzymes for industrial applications. Söderlund et al. achieved rapid production of 2-hydroxyacetophenone from racemic styrene oxide by co-expressing epoxide hydrolase StEH1 and alcohol dehydrogenase in host cells which metabolize, supply, and regenerate precious nicotinamide dinucleotide coenzyme [80]. Recently, Yu et al. reported a unique mutually beneficial P450 monooxygenase-peroxygenase system. Two enzymes in the system can not only be used as catalysts for key reactions but also ensure the internal in situ H2O2 recycle to avoid the toxic effects caused by its accumulation. By using this system, (R)-phenyl glycol and its 9 derivatives were synthesized with high enantioselectivity [81]. Yuan et al. constructed a system for artificial de novo synthesis of rac-phenylethanediol in Saccharomyces cerevisiae using styrene as an intermediate and cascade enzymes as catalysts [82]. Similarly, Wang et al. constructed a dual-enzyme system of epoxide hydrolases to prepare (R)-phenyl-1,2-ethanediol with high enantiomeric excess and yield from high concentration racemic styrene oxide [83].
Purified enzymes often have insufficient stability for industrial production. Whole-cell biocatalysts offer a convenient solution and avoid costly and tedious enzyme purification processes. Therefore, the development of whole-cell catalysts is a prevalent economic method for industrial applications [84,85]. Zhang et al. used the E. coli whole-cell catalyst expressing EH enzyme GmEH3 from soybean (Glycine max) and achieved gram-scale enantioconvergent hydrolysis of rac-epoxides with high enantiopurity and yield [48]. In addition, whole-cell catalysis can change the substrate specificity of the enzymes in some cases. Bisogo et al. revealed that, compared with the commercial epoxide hydrolase from Aspergillus niger, the whole cells of A. niger possessed another new hydrolysis ability that can accept trisubstituted oxiranic steroids as substrates, while the commercial enzymes cannot accept this kind of substrates [86].
Immobilization is also a widely used strategy that can not only improve the functional properties of enzymes but also enhance the reusability of enzymes for various industrial processes. Lots of immobilization methods have been developed for various α/β hydrolases, and the immobilization techniques have been optimized to achieve the best performance according to the nature of the enzymes and the demands of industrial applications.
Adsorption is a simple and cheap physical method for enzyme immobilization. Enzymes bind to the surface of water-insoluble carriers by hydrogen bonds, multiple salt bonds, and van der Waal’s forces. For example, Zheng et al. utilized amine-functionalized meso-cellular foam (MCF) to adsorb DhaA, remarkably improving its stability [87]. Rao et al. used inexpensive celite®545 to physically adsorb Arabidopsis thaliana HNL(AtHNL) to optimize its catalytic efficiency and pH stability, and the immobilized enzymes showed wider substrate selectivity in the retro-Henry reaction [42].
Covalent conjugation is also a frequently used technique to immobilize enzymes. Enzymes are attached to carriers by multi-point covalent bonds, leaving enzyme active sites unaffected and improving enzyme stability [88,89,90]. Based on this principle, Wang et al. reported that the conjugation of DhaA with Arabinogalactan, a hydrophilic polysaccharide, can form a hydration layer and reduce the solvent-accessible area of DhaA, thus slow down the protonation process and improve the pH and high salt stability [43].
Encapsulation is another effective immobilization technique. Enzymes are encapsulated into various spherical polymeric particles or nanoparticles for biocatalysts immobilization. By this means, Huang et al. encapsulated epoxide hydrolase SpEH from Sphingomonas sp. HXN-200 and BSA in organic-inorganic capsules, and improved the activity, stability, and reusability of SpEH [91]. Salvi et al. developed a new enzyme immobilization method using organic-inorganic hybrid nanoflowers with soybean epoxide hydrolase as an organic component and calcium ion as an inorganic component. The epoxide hydrolase nanoflowers showed high activity and stability in the hydrolysis of styrene oxide [92].
In the cross-linking method, enzymes are aggregated and cross-linked with covalent bonds by chemical regents, which form three-dimensional cross-linked enzyme aggregates. Jangir et al. reported the immobilization of Baliospermium montanum hydroxynitrile lyase BmHNL through cross-linking. The immobilized enzymes showed improved enantio-preference in (S)-mandelonitrile synthesis [93].
Recently, Pei et al. reported an interesting immobilization method, catalytic-active inclusion bodies (CatIBs), without additional carriers and chemicals to immobilize enzymes. Two self-assembling peptides, NSPdoT from rotavirus and HVdoT from human Vasodilator-stimulated phosphoprotein, can induce the formation of CatIBs of an (R)-hydroxynitrile lyase AtHNL from Arabidopsis thaliana in the expression host E. coli cells, which significantly improves the thermostability and pH stability of the enzyme [94].
Many substrates catalyzed by α/β hydrolase are non-polar hydrophobic substances with poor solubility and some substrates display spontaneous hydrolysis in water, resulting in low catalytic efficiency. Generally, adding organic solvents to the enzyme reaction system can partially relieve the problem, but organic solvents, particularly in high concentrates, are often toxic to the enzymes, which limits their application [18]. A biphasic system, constructed by natural phase separation or by a membrane, can offer a promising solution to overcome this problem. Hu et al. constructed an n-hexanol/buffer biphasic system for recombinant A. usamii epoxide hydrolase reAuEH2 to solve the difficulty caused by product inhibition. In this biphasic system, efficient resolution of racemic styrene oxide was achieved, and the substrate concentration and volumetric productivity significantly increased [95]. Gao et al. established an aqueous/organic biphasic system based on a hollow fiber membrane bioreactor, which solved the problem of substrate toxicity. The membrane-based biphasic system prevents the foaming and emulsification in the traditional biphasic system, and the two-phase separation of substrate and product reduced the burden of downstream product recovery [96].
7. Conclusions
Most bioactive compounds usually have chirality, which is crucial for their physiological activities, reflecting in the different isomers of racemates may have different activities. Chiral compounds play a key role in fine chemicals, medicine, material science, and other fields, so the demand for single enantiomers is growing over time. Enzymes, an excellent catalyst in nature, have become the best tool for asymmetric synthesis because of their advantages such as high catalytic efficiency, mild reaction conditions, high selectivity, and green regeneration. As one of the largest protein families α/β hydrolases, many members show significant potential in chiral synthesis. This review shows that with our deeper and deeper understanding of the structure and catalytic mechanism of α/β hydrolases, techniques to enhance the performance of these enzymes through protein engineering and other means become more effective and practical. At the same time, the diverse function and manifold substates of α/β hydrolases and their wide distribution in nature indicate that these enzymes can be modified or engineered to catalyze tremendous reactions to generate chiral products that are difficult to synthesize originally. In recent years, thanks to the development of computer science and artificial intelligence, great progress has been made in structural biology, functional genomics, protein design, and protein engineering technology. The application of these new technologies to the customization and improvement of α/β hydrolases will further promote the biosynthesis of chiral compounds, thus contributing to the green and sustainable development of the chemical and pharmaceutical industry.
Author Contributions
Conceptualization, Y.F. and J.X.; writing—original draft preparation, M.Q.; writing—review and editing, S.D., Q.C., Y.F. and J.X. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by the Fundamental Research Funds for the Central Universities (FRF-DF-20-09), Training Program for Young Teaching Backbone Talents, USTB (2302020JXGGRC-005), Major Education and Teaching Reform Research Project, USTB (JG2021ZD01), National Natural Science Foundation of China (32070125), and Shandong Energy Institute (SEI I202114 and SEI S202106).
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Hilterhaus, L.; Liese, A. Building blocks. Adv. Biochem. Eng. Biotechnol. 2007, 105, 133–173. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Ryoo, D.H.; Lee, J.M.; Ryoo, J.J. Synthesis and application of C2 and C3 symmetric (R)-phenylglycinol-derived chiral stationary phases. Chirality 2016, 28, 186–191. [Google Scholar] [CrossRef]
- Hu, X.; Guo, X.; Sun, S.; Zhu, B.; Yu, J.; Guo, X. Enantioseparation of nine indanone and tetralone derivatives by HPLC using carboxymethyl-β-cyclodextrin as the mobile phase additive. Chirality 2017, 29, 38–47. [Google Scholar] [CrossRef]
- Xie, S.M.; Fu, N.; Li, L.; Yuan, B.Y.; Zhang, J.H.; Li, Y.X.; Yuan, L.M. Homochiral metal-organic cage for gas chromatographic separations. Anal. Chem. 2018, 90, 9182–9188. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Mu, X.; Qiao, J.; Su, Y.; Qi, L. New chiral ligand exchange capillary electrophoresis system with chiral amino amide ionic liquids as ligands. Talanta 2017, 175, 451–456. [Google Scholar] [CrossRef]
- Lynch, C.C.; De Los Santos, Z.A.; Wolf, C. Chiroptical sensing of unprotected amino acids, hydroxy acids, amino alcohols, amines and carboxylic acids with metal salts. Chem. Commun. 2019, 55, 6297–6300. [Google Scholar] [CrossRef]
- Wu, S.; Yin, Z.Z.; Wu, D.; Tao, Y.; Kong, Y. Chiral enantioselective assemblies induced from achiral porphyrin by L- and D-lysine. Langmuir 2019, 35, 16761–16769. [Google Scholar] [CrossRef]
- Wang, L.; Jin, Z.; Wang, X.; Zeng, S.; Sun, C.; Pan, Y. Pair of stereodynamic chiral benzylicaldehyde probes for determination of absolute configuration of amino acid residues in peptides by mass spectrometry. Anal. Chem. 2017, 89, 11902–11907. [Google Scholar] [CrossRef]
- Wu, D.; Yu, Y.; Zhang, J.; Guo, L.; Kong, Y. Chiral poly(ionic liquid) with nonconjugated backbone as a fluorescent enantioselective sensor for phenylalaninol and tryptophan. ACS Appl. Mater. Interfaces 2018, 10, 23362–23368. [Google Scholar] [CrossRef] [PubMed]
- Temel, F.; Erdemir, S.; Tabakci, B.; Akpinar, M.; Tabakci, M. Selective chiral recognition of alanine enantiomers by chiral calix [4]arene coated quartz crystal microbalance sensors. Anal. Bioanal. Chem. 2019, 411, 2675–2685. [Google Scholar] [CrossRef]
- Jiang, W.; Fang, B. Synthesizing chiral drug intermediates by biocatalysis. Appl. Biochem. Biotechnol. 2020, 192, 146–179. [Google Scholar] [CrossRef]
- Patel, R.N. Biocatalysis for synthesis of pharmaceuticals. Bioorg. Med. Chem. 2018, 26, 1252–1274. [Google Scholar] [CrossRef]
- Sun, H.; Zhang, H.; Ang, E.L.; Zhao, H. Biocatalysis for the synthesis of pharmaceuticals and pharmaceutical intermediates. Bioorg. Med. Chem. 2018, 26, 1275–1284. [Google Scholar] [CrossRef] [PubMed]
- Holmquist, M. Alpha/Beta-hydrolase fold enzymes: Structures, functions and mechanisms. Curr. Protein Pept. Sci. 2000, 1, 209–235. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.Y. Epoxide hydrolase-mediated enantioconvergent bioconversions to prepare chiral epoxides and alcohols. Biotechnol. Lett. 2008, 30, 1509–1514. [Google Scholar] [CrossRef]
- Koudelakova, T.; Bidmanova, S.; Dvorak, P.; Pavelka, A.; Chaloupkova, R.; Prokop, Z.; Damborsky, J. Haloalkane dehalogenases: Biotechnological applications. Biotechnol. J. 2013, 8, 32–45. [Google Scholar] [CrossRef]
- Wu, S.; Snajdrova, R.; Moore, J.C.; Baldenius, K.; Bornscheuer, U.T. Biocatalysis: Enzymatic synthesis for industrial applications. Angew. Chem. Int. Ed. 2021, 60, 88–119. [Google Scholar] [CrossRef]
- Saini, P.; Sareen, D. An overview on the enhancement of enantioselectivity and stability of microbial epoxide hydrolases. Mol. Biotechnol. 2017, 59, 98–116. [Google Scholar] [CrossRef]
- Schanstra, J.P.; Janssen, D.B. Kinetics of halide release of haloalkane dehalogenase: Evidence for a slow conformational change. Biochemistry 1996, 35, 5624–5632. [Google Scholar] [CrossRef] [PubMed]
- Rea, D.; Fülöp, V. Prolyl oligopeptidase structure and dynamics. CNS Neurol. Disord. Drug Targets 2011, 10, 306–310. [Google Scholar] [CrossRef]
- Pries, F.; van den Wijngaard, A.J.; Bos, R.; Pentenga, M.; Janssen, D.B. The role of spontaneous cap domain mutations in haloalkane dehalogenase specificity and evolution. J. Biol. Chem. 1994, 269, 17490–17494. [Google Scholar] [CrossRef]
- Zou, J.; Hallberg, B.M.; Bergfors, T.; Oesch, F.; Arand, M.; Mowbray, S.L.; Jones, T.A. Structure of Aspergillus niger epoxide hydrolase at 1.8 Å resolution: Implications for the structure and function of the mammalian microsomal class of epoxide hydrolases. Structure 2000, 15, 111–122. [Google Scholar] [CrossRef] [PubMed]
- Gartler, G.; Kratky, C.; Gruber, K. Structural determinants of the enantioselectivity of the hydroxynitrile lyase from Hevea brasiliensis. J. Biotechnol. 2007, 129, 87–97. [Google Scholar] [CrossRef]
- Prokop, Z.; Sato, Y.; Brezovsky, J.; Mozga, T.; Chaloupkova, R.; Koudelakova, T.; Jerabek, P.; Stepankova, V.; Natsume, R.; van Leeuwen, J.G.; et al. Enantioselectivity of haloalkane dehalogenases and its modulation by surface loop engineering. Angew. Chem. Int. Ed. 2010, 49, 6111–6115. [Google Scholar] [CrossRef] [PubMed]
- Strzelczyk, P.; Bujacz, G.D.; Kielbasinski, P.; Blaszczyk, J. Crystal and molecular structure of hexagonal form of lipase B from Candida antarctica. Acta Biochim. Pol. 2016, 63, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Lau, E.Y.; Newby, Z.E.; Bruice, T.C. A theoretical examination of the acid-catalyzed and noncatalyzed ring-opening reaction of an oxirane by nucleophilic addition of acetate. Implications to epoxide hydrolases. J. Am. Chem. Soc. 2001, 123, 3350–3357. [Google Scholar] [CrossRef] [PubMed]
- Widersten, M.; Gurell, A.; Lindberg, D. Structure-function relationships of epoxide hydrolases and their potential use in biocatalysis. Biochim. Biophys. Acta 2010, 1800, 316–326. [Google Scholar] [CrossRef]
- Orru, R.V.; Faber, K. Stereoselectivities of microbial epoxide hydrolases. Curr. Opin. Chem. Biol. 1999, 3, 16–21. [Google Scholar] [CrossRef] [PubMed]
- Casas-Godoy, L.; Gasteazoro, F.; Duquesne, S.; Bordes, F.; Marty, A.; Sandoval, G. Lipases: An overview. Methods Mol. Biol. 2018, 1835, 3–38. [Google Scholar] [CrossRef]
- Jaeger, K.E.; Dijkstra, B.W.; Reetz, M.T. Bacterial biocatalysts: Molecular biology, three-dimensional structures, and biotechnological applications of lipases. Annu. Rev. Microbiol. 1999, 53, 315–351. [Google Scholar] [CrossRef]
- Lang, D.A.; Mannesse, M.L.; de Haas, G.H.; Verheij, H.M.; Dijkstra, B.W. Structural basis of the chiral selectivity of Pseudomonas cepacia lipase. Eur. J. Biochem. 1998, 254, 333–340. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.; Guo, L.; Xu, G.; Wu, J.-P.; Yang, L.-R. A new mechanism of enantioselectivity toward chiral primary alcohol by lipase from Pseudomonas cepacia. J. Mol. Catal. B Enzym. 2014, 109, 109–115. [Google Scholar] [CrossRef]
- Bordes, F.; Cambon, E.; Dossat-Letisse, V.; Andre, I.; Croux, C.; Nicaud, J.M.; Marty, A. Improvement of Yarrowia lipolytica lipase enantioselectivity by using mutagenesis targeted to the substrate binding site. ChemBioChem 2009, 10, 1705–1713. [Google Scholar] [CrossRef]
- Kim, C.; Lee, J.; Cho, J.; Oh, Y.; Choi, Y.K.; Choi, E.; Park, J.; Kim, M.J. Kinetic and dynamic kinetic resolution of secondary alcohols with ionic-surfactant-coated Burkholderia cepacia lipase: Substrate scope and enantioselectivity. J. Org. Chem. 2013, 78, 2571–2578. [Google Scholar] [CrossRef] [PubMed]
- Kitamoto, Y.; Kuruma, Y.; Suzuki, K.; Hattori, T. Effect of solvent polarity on enantioselectivity in Candida antarctica lipase B catalyzed kinetic resolution of primary and secondary alcohols. J. Org. Chem. 2015, 80, 521–527. [Google Scholar] [CrossRef] [PubMed]
- Maldonado, M.R.; Alnoch, R.C.; de Almeida, J.M.; dos Santos, L.A.; Andretta, A.T.; Ropaín, R.d.P.C.; de Souza, E.M.; Mitchell, D.A.; Krieger, N. Key mutation sites for improvement of the enantioselectivity of lipases through protein engineering. Biochem. Eng. J. 2021, 172, 108047. [Google Scholar] [CrossRef]
- Adamu, A.; Wahab, R.A.; Aliyu, F.; Aminu, A.H.; Hamza, M.M.; Huyop, F. Haloacid dehalogenases of Rhizobium sp. and related enzymes: Catalytic properties and mechanistic analysis. Process. Biochem. 2020, 92, 437–446. [Google Scholar] [CrossRef]
- van Leeuwen, J.G.; Wijma, H.J.; Floor, R.J.; van der Laan, J.M.; Janssen, D.B. Directed evolution strategies for enantiocomplementary haloalkane dehalogenases: From chemical waste to enantiopure building blocks. ChemBioChem 2012, 13, 137–148. [Google Scholar] [CrossRef]
- Nuylert, A.; Nakabayashi, M.; Yamaguchi, T.; Asano, Y. Discovery and structural analysis to improve the enantioselectivity of hydroxynitrile lyase from Parafontaria laminata millipedes for (R)-2-chloromandelonitrile synthesis. ACS Omega 2020, 5, 27896–27908. [Google Scholar] [CrossRef]
- Vishnu Priya, B.; Sreenivasa Rao, D.H.; Gilani, R.; Lata, S.; Rai, N.; Akif, M.; Kumar Padhi, S. Enzyme engineering improves catalytic efficiency and enantioselectivity of hydroxynitrile lyase for promiscuous retro-nitroaldolase activity. Bioorg. Chem. 2022, 120, 105594. [Google Scholar] [CrossRef]
- Ding, Y.; Perez-Ortiz, G.; Peate, J.; Barry, S.M. Redesigning enzymes for biocatalysis: Exploiting structural understanding for improved selectivity. Front. Mol. Biosci. 2022, 9, 908285. [Google Scholar] [CrossRef] [PubMed]
- Rao, D.H.S.; Shivani, K.; Padhi, S.K. Immobilized Arabidopsis thaliana hydroxynitrile lyase-catalyzed retro-Henry reaction in the synthesis of (S)-β-nitroalcohols. Appl. Biochem. Biotechnol. 2021, 193, 560–576. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Yu, W.; Shen, L.; Zheng, H.; Guo, X.; Zhong, J.; Hu, T. Conjugation of haloalkane dehalogenase DhaA with arabinogalactan to increase its stability. J. Biotechnol. 2021, 335, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.Q.; He, W.H.; Kong, X.D.; Fan, L.Q.; Zhao, J.; Li, S.X.; Xu, J.H. Heterologous overexpression of Vigna radiata epoxide hydrolase in Escherichia coli and its catalytic performance in enantioconvergent hydrolysis of p-nitrostyrene oxide into (R)-p-nitrophenyl glycol. Appl. Microbiol. Biotechnol. 2014, 98, 207–218. [Google Scholar] [CrossRef]
- Xu, W.; Xu, J.-H.; Pan, J.; Gu, Q.; Wu, X.-Y. Enantioconvergent hydrolysis of styrene epoxides by newly discovered epoxide hydrolases in mung bean. Org. Lett. 2006, 8, 1737–1740. [Google Scholar] [CrossRef]
- Hu, D.; Tang, C.; Li, C.; Kan, T.; Shi, X.; Feng, L.; Wu, M. Stereoselective hydrolysis of epoxides by reVrEH3, a novel Vigna radiata epoxide hydrolase with high enantioselectivity or high and complementary regioselectivity. J. Agric. Food Chem. 2017, 65, 9861–9870. [Google Scholar] [CrossRef]
- Hu, D.; Tang, C.-D.; Yang, B.; Liu, J.-C.; Yu, T.; Deng, C.; Wu, M.-C. Expression of a novel epoxide hydrolase of Aspergillus usamii E001 in Escherichia coli and its performance in resolution of racemic styrene oxide. J. Ind. Microbiol. Biotechnol. 2015, 42, 671–680. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Li, C.; Zhu, X.X.; Liu, Y.Y.; Zhao, J.; Wu, M.C. Highly regio- and enantio-selective hydrolysis of two racemic epoxides by GmEH3, a novel epoxide hydrolase from Glycine max. Int. J. Biol. Macromol. 2020, 164, 2795–2803. [Google Scholar] [CrossRef]
- Xue, F.; Liu, Z.-Q.; Zou, S.-P.; Wan, N.-W.; Zhu, W.-Y.; Zhu, Q.; Zheng, Y.-G. A novel enantioselective epoxide hydrolase from Agromyces mediolanus ZJB120203: Cloning, characterization and application. Process. Biochem. 2014, 49, 409–417. [Google Scholar] [CrossRef]
- Handelsmanl, J.; Rondon, M.R.; Brady, S.F.; Clardy, J.; Goodman, R.M. Molecular biological access to the chemistry of unknown soil microbes: A new frontier for natural products. Chem. Biol. 1998, 5, R245–R249. [Google Scholar] [CrossRef]
- Stojanovski, G.; Dobrijevic, D.; Hailes, H.C.; Ward, J.M. Identification and catalytic properties of new epoxide hydrolases from the genomic data of soil bacteria. Enzyme Microb. Technol. 2020, 139, 109592. [Google Scholar] [CrossRef]
- Ferrandi, E.E.; Sayer, C.; De Rose, S.A.; Guazzelli, E.; Marchesi, C.; Saneei, V.; Isupov, M.N.; Littlechild, J.A.; Monti, D. New thermophilic α/β class epoxide hydrolases found in metagenomes from hot environments. Front. Bioeng. Biotechnol. 2018, 6, 144. [Google Scholar] [CrossRef] [PubMed]
- Jimenez, D.J.; Dini-Andreote, F.; Ottoni, J.R.; de Oliveira, V.M.; van Elsas, J.D.; Andreote, F.D. Compositional profile of α/β-hydrolase fold proteins in mangrove soil metagenomes: Prevalence of epoxide hydrolases and haloalkane dehalogenases in oil-contaminated sites. Microb. Biotechnol. 2015, 8, 604–613. [Google Scholar] [CrossRef] [PubMed]
- Wu, K.; Wang, H.; Sun, H.; Wei, D. Efficient kinetic resolution of phenyl glycidyl ether by a novel epoxide hydrolase from Tsukamurella paurometabola. Appl. Microbiol. Biotechnol. 2015, 99, 9511–9521. [Google Scholar] [CrossRef] [PubMed]
- Ottoni, J.R.; Cabral, L.; de Sousa, S.T.P.; Junior, G.V.L.; Domingos, D.F.; Soares Junior, F.L.; da Silva, M.C.P.; Marcon, J.; Dias, A.C.F.; de Melo, I.S.; et al. Functional metagenomics of oil-impacted mangrove sediments reveals high abundance of hydrolases of biotechnological interest. World J. Microbiol. Biotechnol. 2017, 33, 141. [Google Scholar] [CrossRef]
- Zhang, L.; De, B.C.; Zhang, W.; Mandi, A.; Fang, Z.; Yang, C.; Zhu, Y.; Kurtan, T.; Zhang, C. Mutation of an atypical oxirane oxyanion hole improves regioselectivity of the α/β-fold epoxide hydrolase Alp1U. J. Biol. Chem. 2020, 295, 16987–16997. [Google Scholar] [CrossRef] [PubMed]
- Kotik, M.; Stepanek, V.; Kyslik, P.; Maresova, H. Cloning of an epoxide hydrolase-encoding gene from Aspergillus niger M200, overexpression in E. coli, and modification of activity and enantioselectivity of the enzyme by protein engineering. J. Biotechnol. 2007, 132, 8–15. [Google Scholar] [CrossRef]
- Zou, S.P.; Zheng, Y.G.; Wu, Q.; Wang, Z.C.; Xue, Y.P.; Liu, Z.Q. Enhanced catalytic efficiency and enantioselectivity of epoxide hydrolase from Agrobacterium radiobacter AD1 by iterative saturation mutagenesis for (R)-epichlorohydrin synthesis. Appl. Microbiol. Biotechnol. 2018, 102, 733–742. [Google Scholar] [CrossRef]
- Hu, D.; Hu, B.C.; Hou, X.D.; Zhang, D.; Lei, Y.Q.; Rao, Y.J.; Wu, M.C. Structure-guided regulation in the enantioselectivity of an epoxide hydrolase to produce enantiomeric monosubstituted epoxides and vicinal diols via kinetic resolution. Org. Lett. 2022, 24, 1757–1761. [Google Scholar] [CrossRef]
- Wen, Z.; Hu, D.; Hu, B.C.; Zhang, D.; Huang, J.F.; Wu, M.C. Structure-guided improvement in the enantioselectivity of an Aspergillus usamii epoxide hydrolase for the gram-scale kinetic resolution of ortho-trifluoromethyl styrene oxide. Enzym. Microb. Technol. 2021, 146, 109778. [Google Scholar] [CrossRef]
- Rui, L.; Cao, L.; Chen, W.; Reardon, K.F.; Wood, T.K. Protein engineering of epoxide hydrolase from Agrobacterium radiobacter AD1 for enhanced activity and enantioselective production of (R)-1-phenylethane-1,2-diol. Appl. Environ. Microbiol. 2005, 71, 3995–4003. [Google Scholar] [CrossRef]
- Horsman, G.P.; Lechner, A.; Ohnishi, Y.; Moore, B.S.; Shen, B. Predictive model for epoxide hydrolase-generated stereochemistry in the biosynthesis of nine-membered enediyne antitumor antibiotics. Biochemistry 2013, 52, 5217–5224. [Google Scholar] [CrossRef] [PubMed]
- Jochens, H.; Stiba, K.; Savile, C.; Fujii, R.; Yu, J.G.; Gerassenkov, T.; Kazlauskas, R.J.; Bornscheuer, U.T. Converting an esterase into an epoxide hydrolase. Angew. Chem. Int. Ed. 2009, 48, 3532–3535. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Luan, Z.; Chen, Q.; Xu, J.; Yu, H. Rational selection of circular permutation sites in characteristic regions of the α/β-hydrolase fold enzyme RhEst1. J. Mol. Catal. B: Enzym. 2016, 125, 75–80. [Google Scholar] [CrossRef]
- Jones, B.J.; Lim, H.Y.; Huang, J.; Kazlauskas, R.J. Comparison of five protein engineering strategies for stabilizing an α/β-hydrolase. Biochemistry 2017, 56, 6521–6532. [Google Scholar] [CrossRef]
- Padhi, S.K.; Fujii, R.; Legatt, G.A.; Fossum, S.L.; Berchtold, R.; Kazlauskas, R.J. Switching from an esterase to a hydroxynitrile lyase mechanism requires only two amino acid substitutions. Chem. Biol. 2010, 17, 863–871. [Google Scholar] [CrossRef]
- Beier, A.; Damborsky, J.; Prokop, Z. Transhalogenation catalysed by haloalkane dehalogenases engineered to stop natural pathway at intermediate. Adv. Synth. Catal. 2019, 361, 2438–2442. [Google Scholar] [CrossRef]
- Marek, M.; Chaloupkova, R.; Prudnikova, T.; Sato, Y.; Rezacova, P.; Nagata, Y.; Kuta Smatanova, I.; Damborsky, J. Structural and catalytic effects of surface loop-helix transplantation within haloalkane dehalogenase family. Comput. Struct. Biotechnol. J. 2020, 18, 1352–1362. [Google Scholar] [CrossRef]
- Bednar, D.; Beerens, K.; Sebestova, E.; Bendl, J.; Khare, S.; Chaloupkova, R.; Prokop, Z.; Brezovsky, J.; Baker, D.; Damborsky, J. FireProt: Energy- and evolution-based computational design of thermostable multiple-point mutants. PLoS Comput. Biol. 2015, 11, e1004556. [Google Scholar] [CrossRef]
- Floor, R.J.; Wijma, H.J.; Colpa, D.I.; Ramos-Silva, A.; Jekel, P.A.; Szymanski, W.; Feringa, B.L.; Marrink, S.J.; Janssen, D.B. Computational library design for increasing haloalkane dehalogenase stability. ChemBioChem 2014, 15, 1660–1672. [Google Scholar] [CrossRef]
- von Langermann, J.; Nedrud, D.M.; Kazlauskas, R.J. Increasing the reaction rate of hydroxynitrile lyase from Hevea brasiliensis toward mandelonitrile by copying active site residues from an esterase that accepts aromatic esters. ChemBioChem 2014, 15, 1931–1938. [Google Scholar] [CrossRef] [PubMed]
- Nedrud, D.M.; Lin, H.; Lopez, G.; Padhi, S.K.; Legatt, G.A.; Kaz-Lauskas, R.J. Uncovering divergent evolution of α/β-hydrolases: A surprising residue substitution needed to convert Hevea brasiliensis hydroxynitrile lyase into an esterase. Chem. Sci. 2014, 5, 4265–4277. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Lutz, S. Improved triglyceride transesterification by circular permuted Candida antarctica lipase B. Biotechnol. Bioeng. 2010, 105, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Paye, M.F.; Rose, H.B.; Robbins, J.M.; Yunda, D.A.; Cho, S.; Bommarius, A.S. A high-throughput pH-based colorimetric assay: Application focus on alpha/beta hydrolases. Anal. Biochem. 2018, 549, 80–90. [Google Scholar] [CrossRef]
- Zheng, Y.C.; Ding, L.Y.; Jia, Q.; Lin, Z.; Hong, R.; Yu, H.L.; Xu, J.H. A high-throughput screening method for the directed evolution of hydroxynitrile lyase towards cyanohydrin synthesis. ChemBioChem 2021, 22, 996–1000. [Google Scholar] [CrossRef]
- Glasner, M.E.; Truong, D.P.; Morse, B.C. How enzyme promiscuity and horizontal gene transfer contribute to metabolic innovation. FEBS J. 2020, 287, 1323–1342. [Google Scholar] [CrossRef]
- Kapoor, M.; Gupta, M.N. Lipase promiscuity and its biochemical applications. Process. Biochem. 2012, 47, 555–569. [Google Scholar] [CrossRef]
- Cadet, F.; Fontaine, N.; Li, G.; Sanchis, J.; Ng Fuk Chong, M.; Pandjaitan, R.; Vetrivel, I.; Offmann, B.; Reetz, M.T. A machine learning approach for reliable prediction of amino acid interactions and its application in the directed evolution of enantioselective enzymes. Sci. Rep. 2018, 8, 16757. [Google Scholar] [CrossRef]
- Bommarius, A.S.; Paye, M.F. Stabilizing biocatalysts. Chem. Soc. Rev. 2013, 42, 6534–6565. [Google Scholar] [CrossRef]
- Soderlund, I.; Tjarnhage, E.; Hamnevik, E.; Widersten, M. Facile synthesis of 2-hydroxyacetophenone from racemic styrene oxide catalyzed by engineered enzymes. Biotechnol. Lett. 2022, 44, 985–990. [Google Scholar] [CrossRef]
- Yu, D.; Wang, J.B.; Reetz, M.T. Exploiting designed oxidase-peroxygenase mutual benefit system for asymmetric cascade reactions. J. Am. Chem. Soc. 2019, 141, 5655–5658. [Google Scholar] [CrossRef]
- Yuan, J.; Lukito, B.R.; Li, Z. De novo biosynthesis of (S)- and (R)-phenylethanediol in yeast via artificial enzyme cascades. ACS Synth. Biol. 2019, 8, 1801–1808. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Hu, D.; Zong, X.; Li, J.; Ding, L.; Wu, M.; Li, J. Enantioconvergent hydrolysis of racemic styrene oxide at high concentration by a pair of novel epoxide hydrolases into (R)-phenyl-1,2-ethanediol. Biotechnol. Lett. 2017, 39, 1917–1923. [Google Scholar] [CrossRef]
- Xuan, J.S.; Feng, Y.G. Enantiomeric tartaric acid production using cis-epoxysuccinate hydrolase: History and perspectives. Molecules 2019, 24, 903. [Google Scholar] [CrossRef] [PubMed]
- Zhou, R.; Dong, S.; Feng, Y.; Cui, Q.; Xuan, J. Development of highly efficient whole-cell catalysts of cis-epoxysuccinic acid hydrolase by surface display. Bioresour. Bioprocess. 2022, 9, 92. [Google Scholar] [CrossRef]
- Bisogno, F.R.; Orden, A.A.; Pranzoni, C.A.; Cifuente, D.A.; Giordano, O.S.; Kurina Sanz, M. Atypical regioselective biohydrolysis on steroidal oxiranes by Aspergillus niger whole cells: Some stereochemical features. Steroids 2007, 72, 643–652. [Google Scholar] [CrossRef]
- Zheng, H.; Yu, W.L.; Guo, X.; Zhao, Y.Z.; Cui, Y.; Hu, T.; Zhong, J.Y. An effective immobilized haloalkane dehalogenase DhaA from Rhodococcus rhodochrous by adsorption, crosslink and PEGylation on meso-cellular foam. Int. J. Biol. Macromol. 2019, 125, 1016–1023. [Google Scholar] [CrossRef]
- Zhao, Y.Z.; Yu, W.L.; Zheng, H.; Guo, X.; Guo, N.; Hu, T.; Zhong, J.Y. PEGylation with the thiosuccinimido butylamine linker significantly increases the stability of haloalkane dehalogenase DhaA. J. Biotechnol. 2017, 254, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Zou, S.P.; Xuan, X.L.; Wang, Z.J.; Zheng, Y.G. Conjugation of Agrobacterium radiobacter epoxide hydrolase with ficoll: Catalytic, kinetic and thermodynamic analysis. Int. J. Biol. Macromol. 2018, 119, 1098–1105. [Google Scholar] [CrossRef]
- Shan, Y.; Yu, W.; Shen, L.; Guo, X.; Zheng, H.; Zhong, J.; Hu, T.; Han, Y. Conjugation with inulin improves the environmental stability of haloalkane dehalogenase DhaA. Enzym. Microb. Technol. 2021, 149, 109832. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.; Wu, M.; Goldman, M.J.; Li, Z. Encapsulation of enzyme via one-step template-free formation of stable organic-inorganic capsules: A simple and efficient method for immobilizing enzyme with high activity and recyclability. Biotechnol. Bioeng. 2015, 112, 1092–1101. [Google Scholar] [CrossRef] [PubMed]
- Salvi, H.M.; Yadav, G.D. Organic-inorganic epoxide hydrolase hybrid nanoflowers with enhanced catalytic activity: Hydrolysis of styrene oxide to 1-phenyl-1,2-ethanediol. J. Biotechnol. 2021, 341, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Jangir, N.; Padhi, S.K. Immobilized Baliospermum montanum hydroxynitrile lyase catalyzed synthesis of chiral cyanohydrins. Bioorg. Chem. 2019, 84, 32–40. [Google Scholar] [CrossRef]
- Pei, X.; Wang, J.; Zheng, H.; Xiao, Q.; Wang, A.; Su, W. Catalytically active inclusion bodies (CatIBs) induced by terminally attached self-assembling coiled-coil domains: To enhance the stability of (R)-hydroxynitrile lyase. Enzym. Microb. Technol. 2022, 153, 109915. [Google Scholar] [CrossRef] [PubMed]
- Hu, D.; Wang, R.; Shi, X.L.; Ye, H.H.; Wu, Q.; Wu, M.C.; Chu, J.J. Kinetic resolution of racemic styrene oxide at a high concentration by recombinant Aspergillus usamii epoxide hydrolase in an n-hexanol/buffer biphasic system. J. Biotechnol. 2016, 236, 152–158. [Google Scholar] [CrossRef]
- Gao, P.; Wu, S.; Praveen, P.; Loh, K.C.; Li, Z. Enhancing productivity for cascade biotransformation of styrene to (S)-vicinal diol with biphasic system in hollow fiber membrane bioreactor. Appl. Microbiol. Biotechnol. 2017, 101, 1857–1868. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).