
Gamma Carbonic Anhydrases from Hydrothermal Vent Bacteria: Cases of Alternating Active Site Due to a Long Loop with Proton Shuttle Residue



Abstract
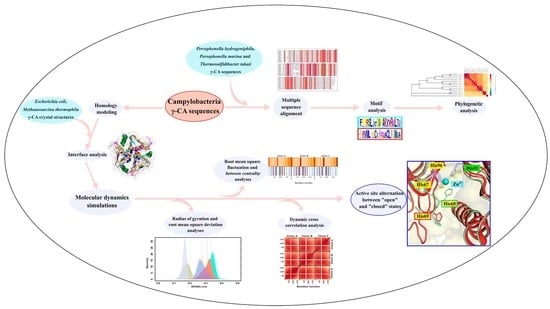
:
1. Introduction
2. Results and Discussion
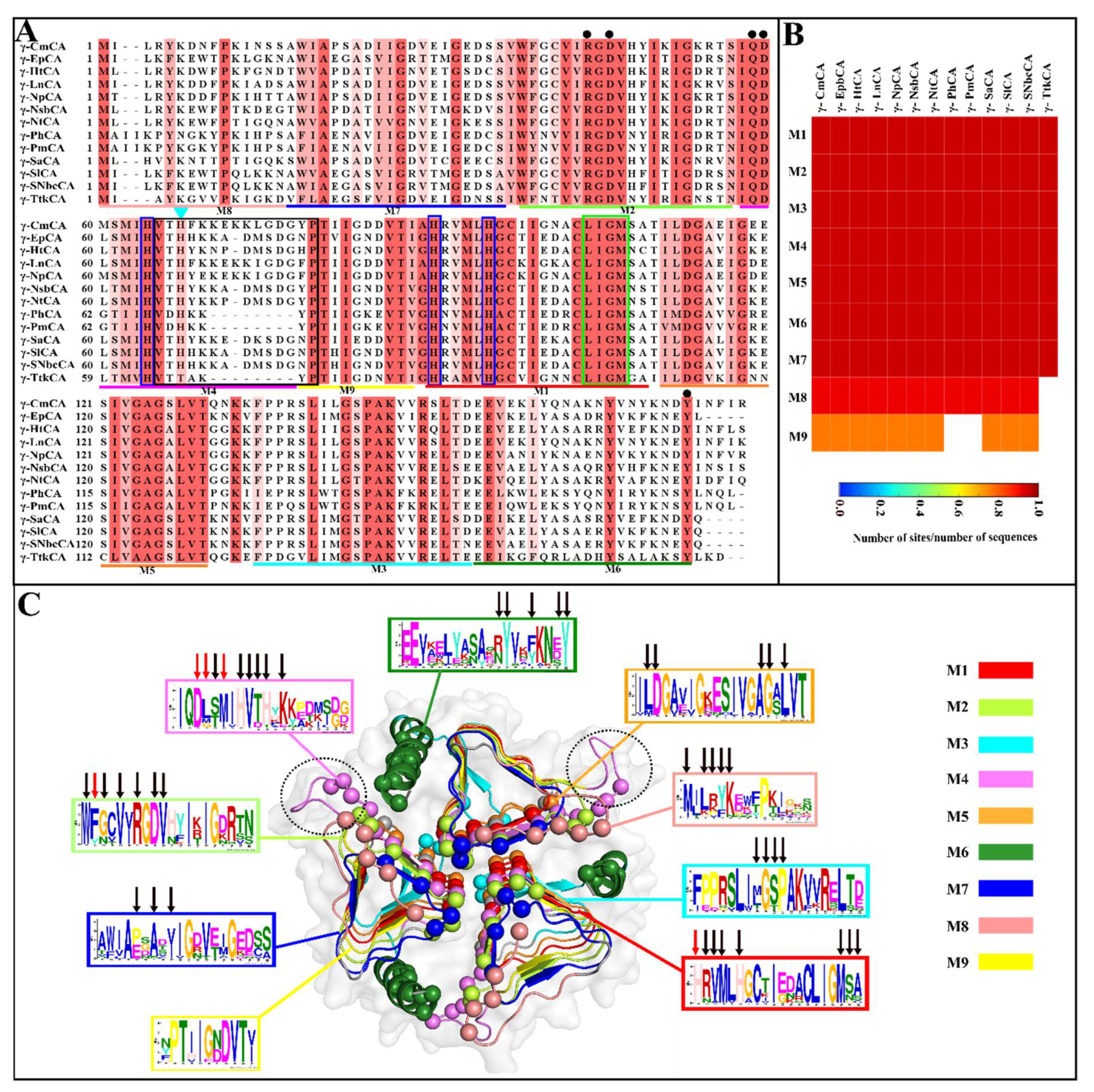
2.1. Sequence Analysis
2.1.1. Conserved Regions and Motifs Are Identified Using Multiple Sequence Alignment and Motif Analysis
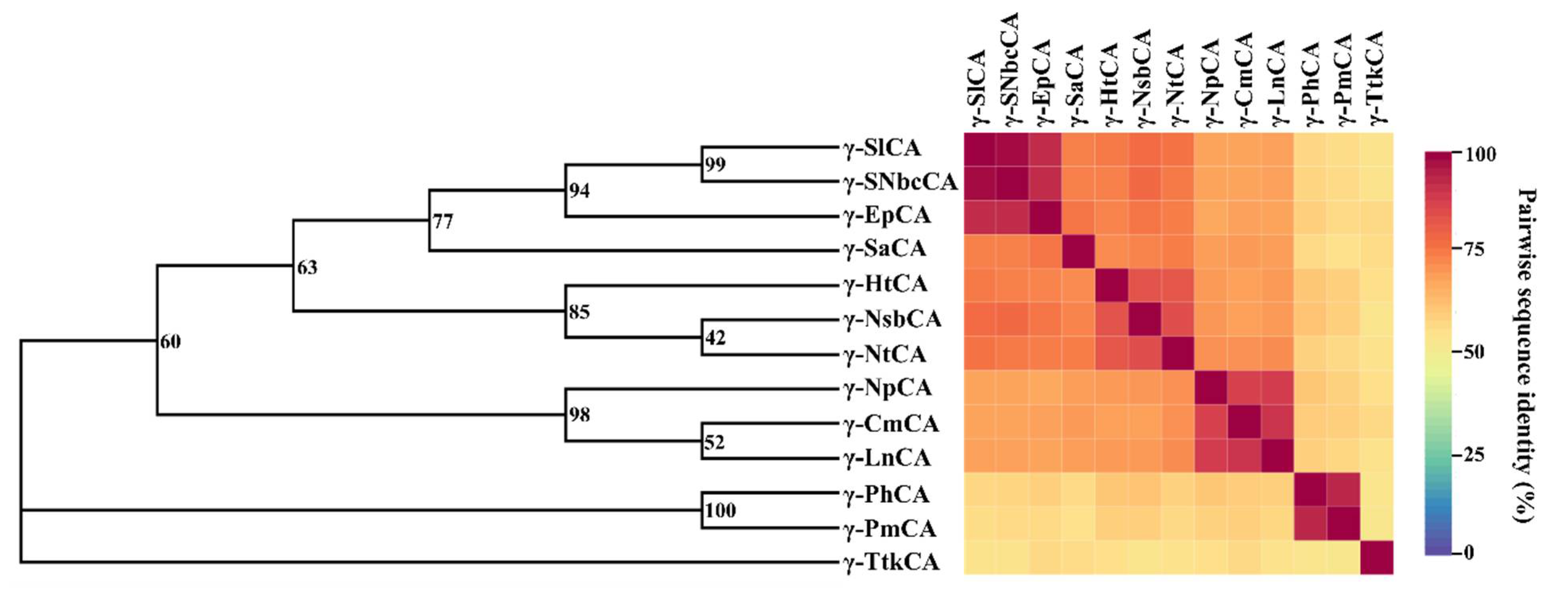
2.1.2. Evolutionary Relationships amongst the γ-CAs Are Investigated Using Phylogeny
2.2. Structure Analysis
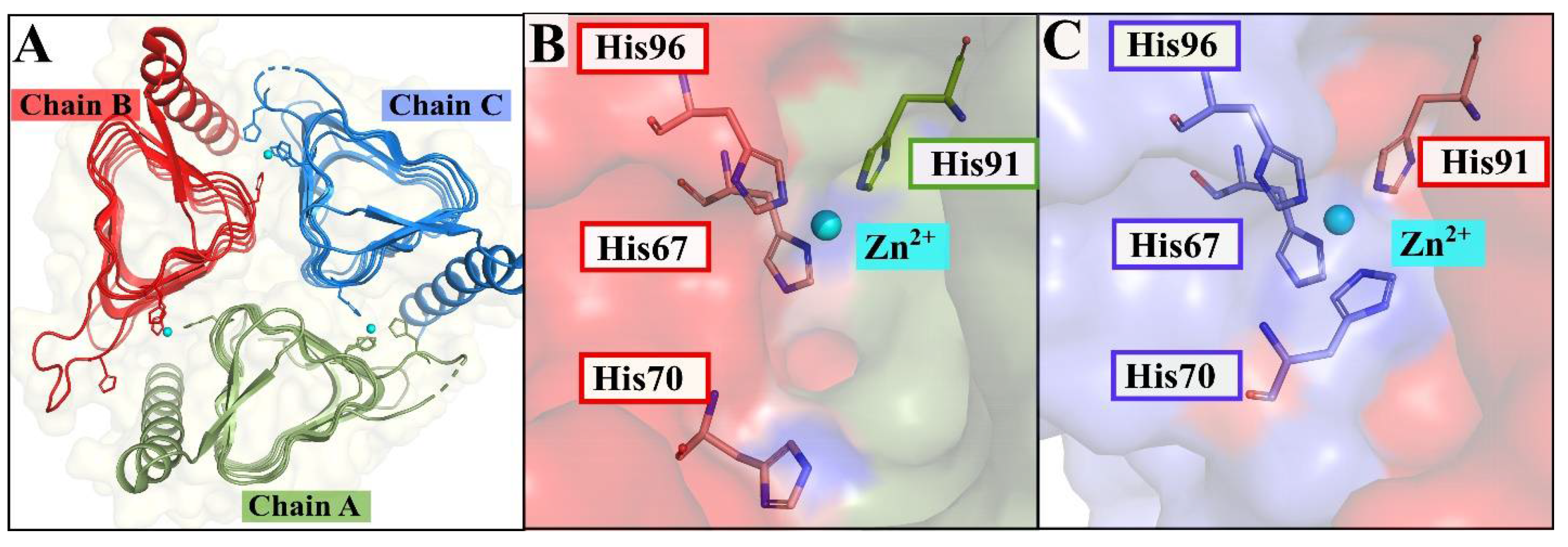
2.2.1. Interface and Hotspot Residues Are Probed in the Trimeric Structures
2.3. Molecular Dynamics Analysis
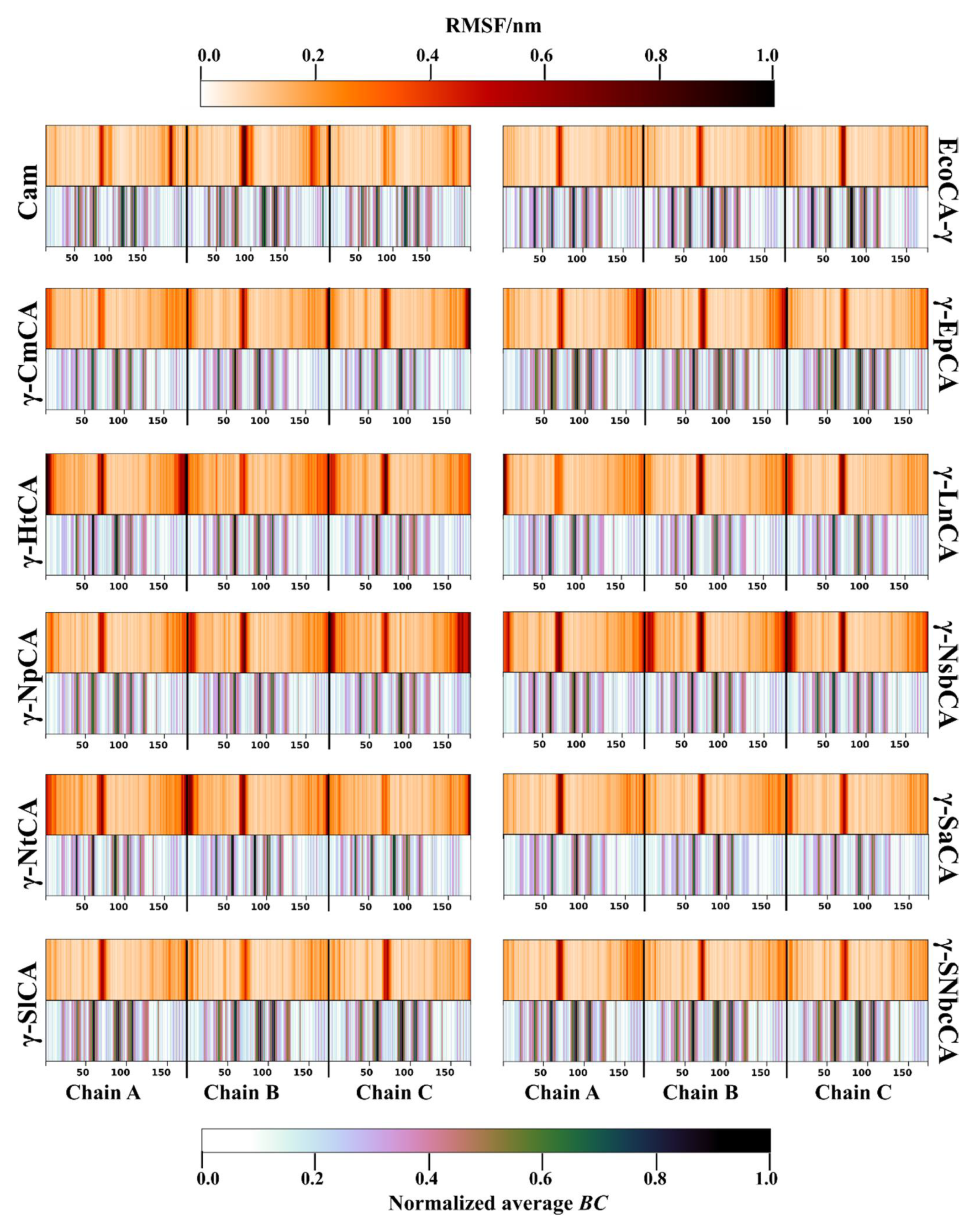
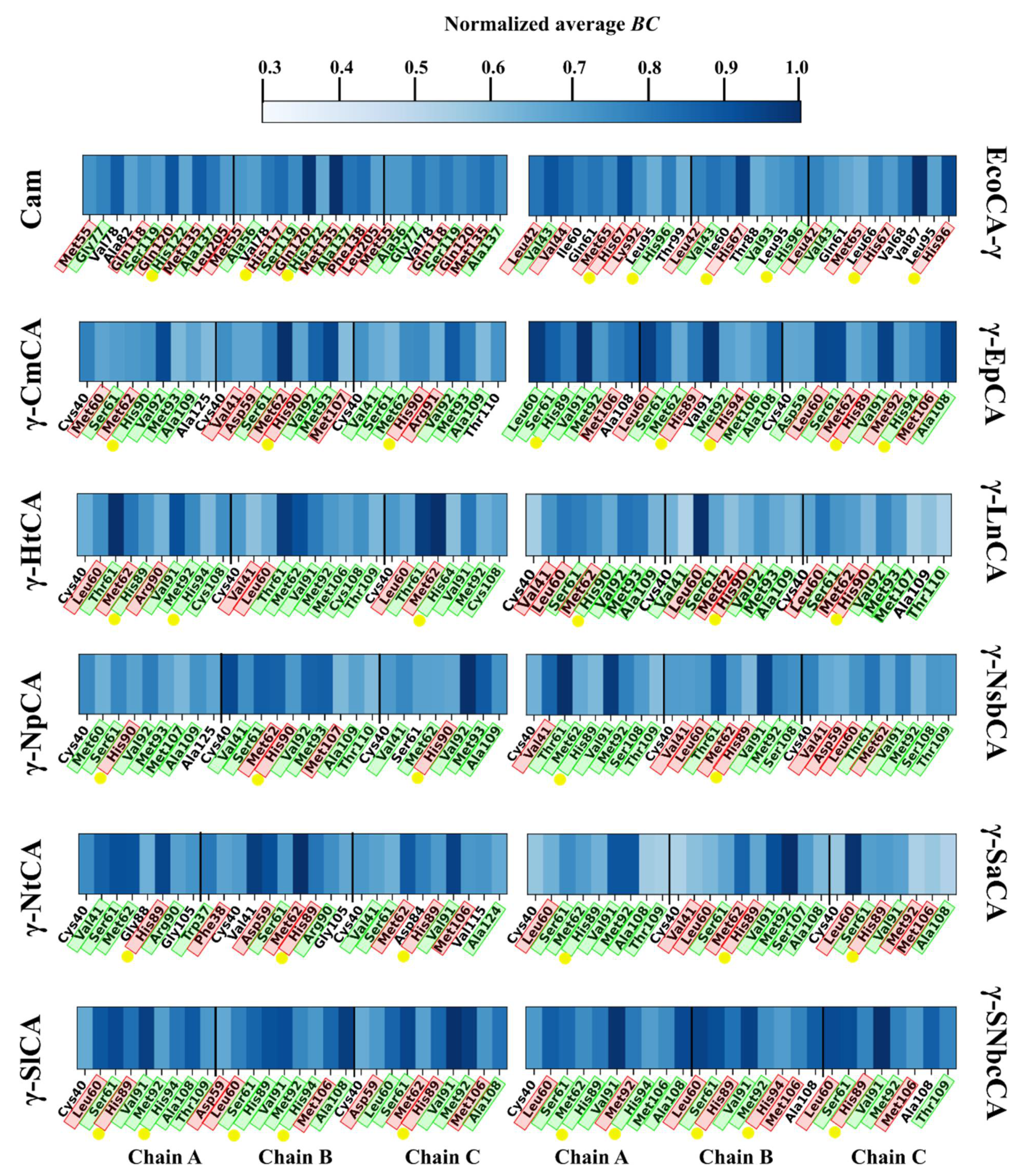
2.3.1. Residue Fluctuation and Functionality of High Communication Residues Is Investigated
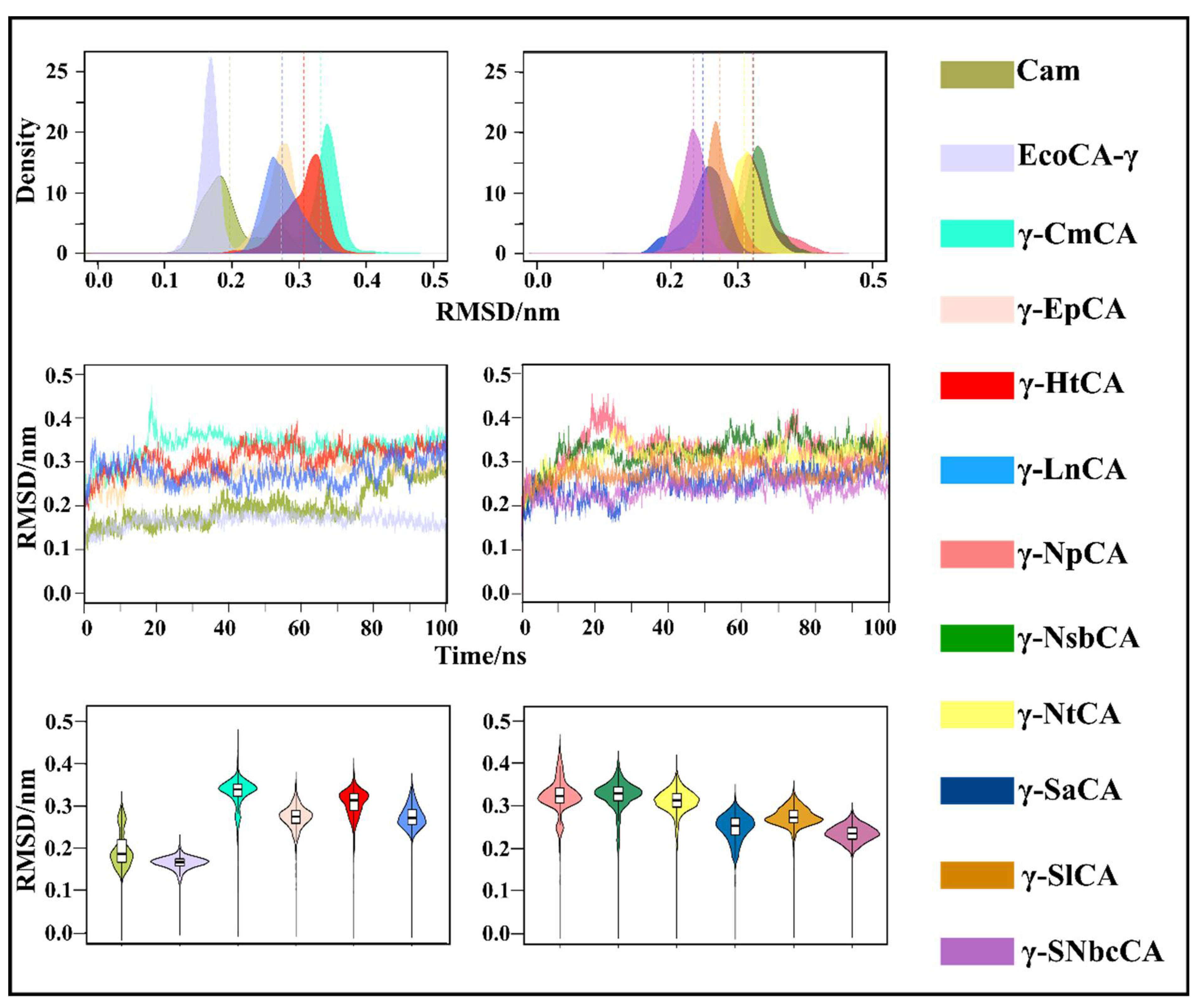
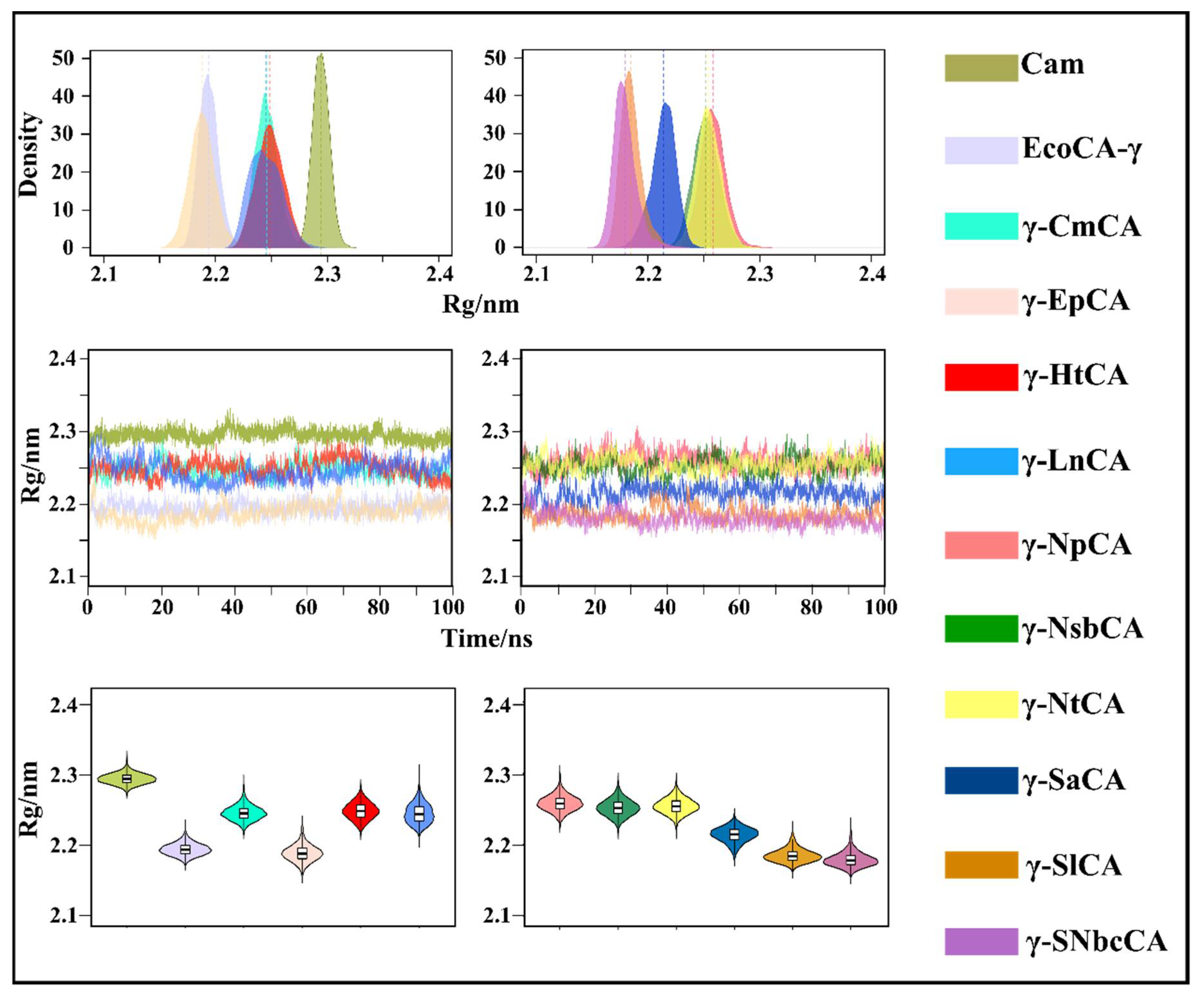
2.3.2. Conformational Changes Are Assessed Using RMSD and Rg Analysis
2.3.3. A Conformational Switch in Proton Shuttling His Close to the Active Site Is Observed
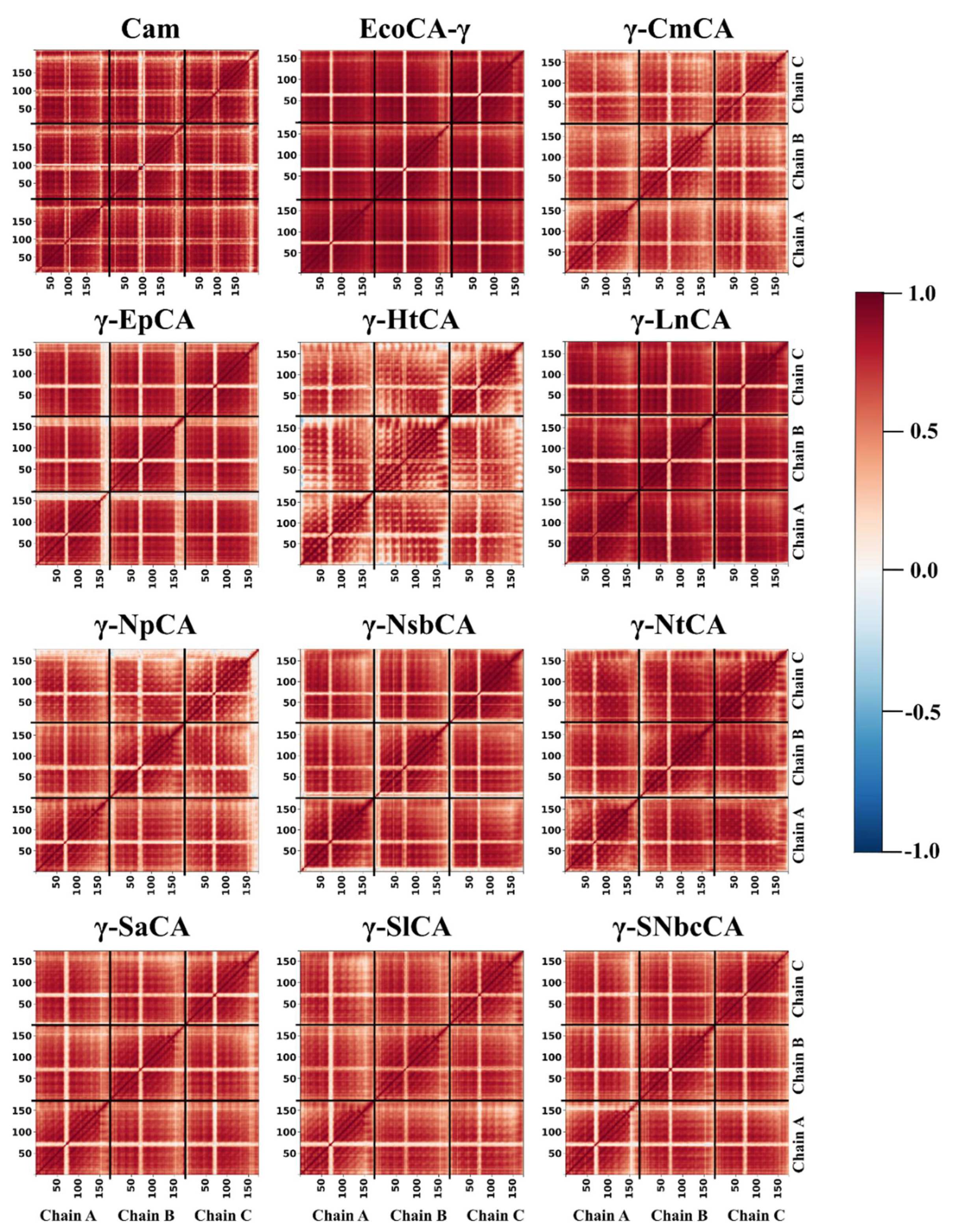
2.3.4. High Residue Correlations Are Perceived Using Dynamic Cross Correlation Analysis
3. Materials and Methods
3.1. Sequence Retrieval and Alignment
3.2. Sequence Alignments and Motif Analysis
3.3. Phylogenetic Analysis
3.4. Homology Modeling, Interface, and Hotspot Residue Identification
3.5. Molecular Dynamics Simulations
3.6. Average Betweenness Centrality Analysis
3.7. Dynamic Cross Correlation Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| BC | Betweenness centrality |
| CA | Carbonic anhydrase |
| Cam | Methanosarcina thermophila gamma carbonic anhydrase |
| EcoCA-γ | Escherichia coli gamma carbonic anhydrase |
| DCC | Dynamic cross correlation |
| γ-CmCA | Caminibacter mediatlanticus γ-carbonic anhydrase |
| γ-EpCA | Epsilonproteobacteria bacterium 4484_65 γ-carbonic anhydrase |
| γ-HtCA | Hydrogenimonas thermophila γ-carbonic anhydrase |
| γ-LnCA | Lebetimonas natsushimae γ-carbonic anhydrase |
| γ-NpCA | Nautilia profundicola γ-carbonic anhydrase |
| γ-NsbCA | Nitratiruptor sp. SB155-2 γ-carbonic anhydrase |
| γ-NtCA | Nitratiruptor tergarcus γ-carbonic anhydrase |
| γ-PhCA | Persephonella hydrogeniphila γ-carbonic anhydrase |
| γ-PmCA | Persephonella marina γ-carbonic anhydrase |
| γ-SaCA | Sulfurimonas autotrophica γ-carbonic anhydrase |
| γ-SlCA | Sulfurovum lithotrophicum γ-carbonic anhydrase |
| γ-SNbcCA | Sulfurovum sp. NBC37-1 γ-carbonic anhydrase |
| γ-TtkCA | Thermosulfidibacter takaii gamma carbonic anhydrase |
| MD | Molecular dynamics |
| MSA | Multiple sequence alignment |
| Rg | Radius of gyration |
| RMSD | Root mean square deviation |
| RMSF | Root mean square fluctuation |
References
- Tripp, B.C.; Smith, K.; Ferry, J.G. Carbonic anhydrase: New insights for an ancient enzyme. J. Biol. Chem. 2001, 276, 48615–48618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Fiore, A.; Alterio, V.; Monti, S.M.; De Simone, G.; D’Ambrosio, K. Thermostable carbonic anhydrases in biotechnological applications. Int. J. Mol. Sci. 2015, 16, 15456–15480. [Google Scholar] [CrossRef] [PubMed]
- DiMario, R.J.; Machingura, M.C.; Waldrop, G.L.; Moroney, J.V. The many types of carbonic anhydrases in photosynthetic organisms. Plant Sci. 2018, 268, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Jensen, E.L.; Clement, R.; Kosta, A.; Maberly, S.C.; Gontero, B. A new widespread subclass of carbonic anhydrase in marine phytoplankton. ISME J. 2019, 13, 2094–2106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kikutani, S.; Nakajima, K.; Nagasato, C.; Tsuji, Y.; Miyatake, A.; Matsuda, Y. Thylakoid luminal θ-carbonic anhydrase critical for growth and photosynthesis in the marine diatom Phaeodactylum tricornutum. Proc. Natl. Acad. Sci. USA 2016, 113, 9828–9833. [Google Scholar] [CrossRef] [Green Version]
- Tan, S.I.; Han, Y.L.; Yu, Y.J.; Chiu, C.Y.; Chang, Y.K.; Ouyang, S.; Fan, K.C.; Lo, K.H.; Ng, I.S. Efficient carbon dioxide sequestration by using recombinant carbonic anhydrase. Process Biochem. 2018, 73, 38–46. [Google Scholar] [CrossRef]
- Manyumwa, C.V.; Emameh, R.Z.; Tastan Bishop, Ö. Alpha-Carbonic Anhydrases from Hydrothermal Vent Sources as Potential Carbon Dioxide Sequestration Agents: In Silico Sequence, Structure and Dynamics Analyses. Int. J. Mol. Sci. 2020, 21, 8066. [Google Scholar] [CrossRef]
- Ferry, J.G. The γ class of carbonic anhydrases. Biochim. Biophys. Acta—Proteins Proteom. 2010, 1804, 374–381. [Google Scholar] [CrossRef] [Green Version]
- Lane, T.W.; Saito, M.A.; George, G.N.; Pickering, I.J.; Prince, R.C.; Morel, F.M. Biochemistry: A cadmium enzyme from a marine diatom. Nature 2005, 435, 42. [Google Scholar] [CrossRef]
- Tripp, B.C.; Bell, C.B.; Cruz, F.; Krebs, C.; Ferry, J.G. A role for iron in an ancient carbonic anhydrase. J. Biol. Chem. 2004, 279, 6683–6687. [Google Scholar] [CrossRef] [Green Version]
- Alterio, V.; Langella, E.; Viparelli, F.; Vullo, D.; Ascione, G.; Dathan, N.A.; Morel, F.M.; Supuran, C.T.; De Simone, G.; Monti, S.M. Structural and inhibition insights into carbonic anhydrase CDCA1 from the marine diatom Thalassiosira weissflogii. Biochimie 2012, 94, 1232–1241. [Google Scholar] [CrossRef]
- Xu, Y.; Feng, L.; Jeffrey, P.D.; Shi, Y.; Morel, F.M. Structure and metal exchange in the cadmium carbonic anhydrase of marine diatoms. Nature 2008, 452, 56–61. [Google Scholar] [CrossRef]
- Liljas, A.; Kannan, K.K.; Bergsten, P.C.; Waara, I.; Fridborg, K.; Strandberg, B.; Carlbom, U.; Järup, L.; Lövgren, S.; Petef, M. Crystal structure of human carbonic anhydrase C. Nat. New Biol. 1972, 235, 131–137. [Google Scholar] [CrossRef]
- Cox, E.H.; McLendon, G.L.; Morel, F.M.; Lane, T.W.; Prince, R.C.; Pickering, I.J.; George, G.N. The active site structure of Thalassiosira weissflogii carbonic anhydrase 1. Biochemistry 2000, 39, 12128–12130. [Google Scholar] [CrossRef]
- Jeyakanthan, J.; Rangarajan, S.; Mridula, P.; Kanaujia, S.P.; Shiro, Y.; Kuramitsu, S.; Yokoyama, S.; Sekar, K. Observation of a calcium-binding site in the γ-class carbonic anhydrase from Pyrococcus horikoshii Horikoshii. Acta Crystallogr. Sect. D Biol. Crystallogr. 2008, 64, 1012–1019. [Google Scholar] [CrossRef]
- Zimmerman, S.A.; Tomb, J.-F.; Ferry, J.G. Characterization of CamH from Methanosarcina thermophila, founding member of a subclass of the γ class of carbonic anhydrases. J. Bacteriol. 2010, 192, 1353–1360. [Google Scholar] [CrossRef] [Green Version]
- Coleman, J.E. Mechanism of action of carbonic anhydrase substrate, sulfonamide, and anion binding. J. Biol. Chem. 1967, 242, 5212–5219. [Google Scholar] [CrossRef]
- Park, H.M.; Park, J.H.; Choi, J.W.; Lee, J.; Kim, B.Y.; Jung, C.H.; Kim, J.S. Structures of the γ-class carbonic anhydrase homologue YrdA suggest a possible allosteric switch. Acta Crystallogr. Sect. D Biol. Crystallogr. 2012, 68, 920–926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sawaya, M.R.; Cannon, G.C.; Heinhorst, S.; Tanaka, S.; Williams, E.B.; Yeates, T.O.; Kerfeld, C.A. The structure of β-carbonic anhydrase from the carboxysomal shell reveals a distinct subclass with one active site for the price of two. J. Biol. Chem. 2006, 281, 7546–7555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Covarrubias, A.S.; Larsson, A.M.; Högbom, M.; Lindberg, J.; Bergfors, T.; Björkelid, C.; Mowbray, S.L.; Unge, T.; Jones, T.A. Structure and function of carbonic anhydrases from Mycobacterium tuberculosis. J. Biol. Chem. 2005, 280, 18782–18789. [Google Scholar] [CrossRef] [Green Version]
- Covarrubias, A.S.; Bergfors, T.; Jones, T.A.; Hogbom, M. Structural mechanics of the pH- dependent activity of β-carbonic anhydrase from Mycobacterium tuberculosis. J. Biol. Chem. 2006, 281, 4993–4999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferraroni, M.; Del Prete, S.; Vullo, D.; Capasso, C.; Supuran, C.T. Crystal structure and kinetic studies of a tetrameric type II β-carbonic anhydrase from the pathogenic bacterium Vibrio cholerae. Acta Crystallogr. Sect. D Biol. Crystallogr. 2015, 71, 2449–2456. [Google Scholar] [CrossRef] [PubMed]
- Del Prete, S.; Bua, S.; Supuran, C.T.; Capasso, C. Escherichia coli γ-carbonic anhydrase: Characterisation and effects of simple aromatic/heterocyclic sulphonamide inhibitors. J. Enzym. Inhib. Med. Chem. 2020, 35, 1545–1554. [Google Scholar] [CrossRef] [PubMed]
- Manyumwa, C.V.; Tastan Bishop, Ö. In Silico Investigation of Potential Applications of Gamma Carbonic Anhydrases as Catalysts of CO2 Biomineralization Processes: A Visit to the Thermophilic Bacteria Persephonella hydrogeniphila, Persephonella marina, Thermosulfidibacter takaii, and Thermus thermophilus. Int. J. Mol. Sci. 2021, 22, 2861. [Google Scholar] [CrossRef] [PubMed]
- Waite, D.W.; Vanwonterghem, I.; Rinke, C.; Parks, D.H.; Zhang, Y.; Takai, K.; Sievert, S.M.; Simon, J.; Campbell, B.J.; Hanson, T.E.; et al. Comparative genomic analysis of the class Epsilonproteobacteria and proposed reclassification to Epsilonbacteraeota (phyl. nov.). Front. Microbiol. 2017, 8, 682. [Google Scholar] [CrossRef] [PubMed]
- Waite, D.W.; Vanwonterghem, I.; Rinke, C.; Parks, D.H.; Zhang, Y.; Takai, K.; Sievert, S.M.; Simon, J.; Campbell, B.J.; Hanson, T.E.; et al. Addendum: Comparative genomic analysis of the class Epsilonproteobacteria and proposed reclassification to Epsilonbacteraeota (phyl. nov.). Front. Microbiol. 2018, 9, 772. [Google Scholar] [CrossRef] [PubMed]
- Brown, D.K.; Penkler, D.L.; Sheik Amamuddy, O.; Ross, C.; Atilgan, A.R.; Atilgan, C.; Tastan Bishop, Ö. MD-TASK: A software suite for analyzing molecular dynamics trajectories. Bioinformatics 2017, 33, 2768–2771. [Google Scholar] [CrossRef] [Green Version]
- Alber, B.E.; Ferry, J.G. Characterization of heterologously produced carbonic anhydrase from Methanosarcina thermophila. J. Bacteriol. 1996, 178, 3270–3274. [Google Scholar] [CrossRef] [Green Version]
- Iverson, T.M.; Alber, B.E.; Kisker, C.; Ferry, J.G.; Rees, D.C. A closer look at the active site of γ-class carbonic anhydrases: High-resolution crystallographic studies of the carbonic anhydrase from Methanosarcina thermophila. Biochemistry 2000, 39, 9222–9231. [Google Scholar] [CrossRef]
- Le, S.Q.; Gascuel, O. An improved general amino acid replacement matrix. Mol. Biol. Evol. 2008, 25, 1307–1320. [Google Scholar] [CrossRef] [Green Version]
- Herrou, J.; Crosson, S. Molecular structure of the Brucella abortus metalloprotein RicA, a Rab2-binding virulence effector. Biochemistry 2013, 52, 9020–9028. [Google Scholar] [CrossRef] [Green Version]
- Eisenberg, D.; Lüthy, R.; Bowie, J.U. [20] VERIFY3D: Assessment of protein models with three-dimensional profiles. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1997; Volume 277, pp. 396–404. [Google Scholar] [CrossRef]
- Wiederstein, M.; Sippl, M.J. ProSA-web: Interactive web service for the recognition of errors in three-dimensional structures of proteins. Nucleic Acids Res. 2007, 35, W407–W410. [Google Scholar] [CrossRef] [Green Version]
- Laskowski, R.A.; MacArthur, M.W.; Moss, D.S.; Thornton, J.M. PROCHECK: A program to check the stereochemical quality of protein structures. J. Appl. Crystallogr. 1993, 26, 283–291. [Google Scholar] [CrossRef]
- DeLano, W. Pymol: An open-source molecular graphics tool. CCP4 Newsl. Protein Crystallogr. 2002, 40, 82–92. [Google Scholar]
- Krissinel, E.; Henrick, K. Inference of macromolecular assemblies from crystalline state. J. Mol. Biol. 2007, 372, 774–797. [Google Scholar] [CrossRef] [PubMed]
- Kortemme, T.; Kim, D.E.; Baker, D. Computational alanine scanning of protein-protein interfaces. Sci. STKE 2004, 219, l2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tina, K.; Bhadra, R.; Srinivasan, N. PIC: Protein interactions calculator. Nucleic Acids Res. 2007, 35, W473–W476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Penkler, D.L.; Atilgan, C.; Tastan Bishop, Ö. Allosteric modulation of human Hsp90 α conformational dynamics. J. Chem. Inf. Model. 2018, 58, 383–404. [Google Scholar] [CrossRef] [Green Version]
- Laitaoja, M.; Valjakka, J.; Jänis, J. Zinc coordination spheres in protein structures. Inorg. Chem. 2013, 52, 10983–10991. [Google Scholar] [CrossRef]
- Borchert, M.; Saunders, P. Heat-Stable Carbonic Anhydrases and Their Use. U.S. Patent 8,945,826, 29 December 2010. [Google Scholar]
- Voordeckers, J.W.; Starovoytov, V.; Vetriani, C. Caminibacter mediatlanticus sp. nov., a thermophilic, chemolithoautotrophic, nitrate-ammonifying bacterium isolated from a deep-sea hydrothermal vent on the Mid-Atlantic Ridge. Int. J. Syst. Evol. Microbiol. 2005, 55, 773–779. [Google Scholar] [CrossRef]
- Takai, K.; Nealson, K.H.; Horikoshi, K. Hydrogenimonas thermophila gen. nov., sp. nov., a novel thermophilic, hydrogen-oxidizing chemolithoautotroph within the ε-Proteobacteria, isolated from a black smoker in a Central Indian Ridge hydrothermal field. Int. J. Syst. Evol. Microbiol. 2004, 54, 25–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagata, R.; Takaki, Y.; Tame, A.; Nunoura, T.; Muto, H.; Mino, S.; Sawayama, S.; Takai, K.; Nakagawa, S. Lebetimonas natsushimae sp. nov., a novel strictly anaerobic, moderately thermophilic chemoautotroph isolated from a deep-sea hydrothermal vent polychaete nest in the Mid-Okinawa Trough. Syst. Appl. Microbiol. 2017, 40, 352–356. [Google Scholar] [CrossRef] [PubMed]
- Dombrowski, N.; Seitz, K.W.; Teske, A.P.; Baker, B.J. Genomic insights into potential interdependencies in microbial hydrocarbon and nutrient cycling in hydrothermal sediments. Microbiome 2017, 5, 106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, J.L.; Campbell, B.J.; Hanson, T.E.; Zhang, C.L.; Cary, S.C. Nautilia profundicola sp. nov., a thermophilic, sulfur-reducing epsilonproteobacterium from deep-sea hydrothermal vents. Int. J. Syst. Evol. Microbiol. 2008, 58, 1598–1602. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, S.; Takai, K.; Inagaki, F.; Horikoshi, K.; Sako, Y. Nitratiruptor tergarcus gen. nov., sp. nov. and Nitratifractor salsuginis gen. nov., sp. nov., nitrate-reducing chemolithoautotrophs of the ε-Proteobacteria isolated from a deep-sea hydrothermal system in the Mid-Okinawa Trough. Int. J. Syst. Evol. Microbiol. 2005, 55, 925–933. [Google Scholar] [CrossRef] [PubMed]
- Inagaki, F.; Takai, K.; Nealson, K.H.; Horikoshi, K. Sulfurovum lithotrophicum gen. nov., sp. nov., a novel sulfur-oxidizing chemolithoautotroph within the ε-Proteobacteria isolated from Okinawa Trough hydrothermal sediments. Int. J. Syst. Evol. Microbiol. 2004, 54, 1477–1482. [Google Scholar] [CrossRef]
- Inagaki, F.; Takai, K.; Kobayashi, H.; Nealson, K.H.; Horikoshi, K. Sulfurimonas autotrophica gen. nov., sp. nov., a novel sulfur-oxidizing ε-proteobacterium isolated from hydrothermal sediments in the Mid-Okinawa Trough. Int. J. Syst. Evol. Microbiol. 2003, 53, 1801–1805. [Google Scholar] [CrossRef] [Green Version]
- Sikorski, J.; Munk, C.; Lapidus, A.; Djao, O.D.N.; Lucas, S.; Del Rio, T.G.; Nolan, M.; Tice, H.; Han, C.; Cheng, J.F.; et al. Complete genome sequence of Sulfurimonas autotrophica type strain (OK10T). Stand. Genom. Sci. 2010, 3, 194. [Google Scholar] [CrossRef] [Green Version]
- Nakagawa, S.; Takaki, Y.; Shimamura, S.; Reysenbach, A.L.; Takai, K.; Horikoshi, K. Deep-sea vent ε-proteobacterial genomes provide insights into emergence of pathogens. Proc. Natl. Acad. Sci. USA 2007, 104, 12146–12150. [Google Scholar] [CrossRef] [Green Version]
- Nakagawa, S.; Takai, K.; Inagaki, F.; Hirayama, H.; Nunoura, T.; Horikoshi, K.; Sako, Y. Distribution, phylogenetic diversity and physiological characteristics of epsilon-Proteobacteria in a deep-sea hydrothermal field. Environ. Microbiol. 2005, 7, 1619–1632. [Google Scholar] [CrossRef]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Söding, J.; et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef]
- Katoh, K.; Misawa, K.; Kuma, K.I.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [Green Version]
- Notredame, C.; Higgins, D.G.; Heringa, J. T-coffee: A novel method for fast and accurate multiple sequence alignment1. J. Mol. Biol. 2000, 302, 205–217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Tommaso, P.; Moretti, S.; Xenarios, I.; Orobitg, M.; Montanyola, A.; Chang, J.M.; Taly, J.F.; Notredame, C. T-Coffee: A web server for the multiple sequence alignment of protein and RNA sequences using structural information and homology extension. Nucleic Acids Res. 2011, 39, W13–W17. [Google Scholar] [CrossRef] [PubMed]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME SUITE: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, W202–W208. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whelan, S.; Goldman, N. A general empirical model of protein evolution derived from multiple protein families using a maximum-likelihood approach. Mol. Biol. Evol. 2001, 18, 691–699. [Google Scholar] [CrossRef] [Green Version]
- Huson, D.H.; Richter, D.C.; Rausch, C.; Dezulian, T.; Franz, M.; Rupp, R. Dendroscope: An interactive viewer for large phylogenetic trees. BMC Bioinf. 2007, 8, 460. [Google Scholar] [CrossRef] [Green Version]
- Parisi, G.; Perales, M.; Fornasari, M.; Colaneri, A.; Schain, N.; Casati, D.; Zimmermann, S.; Brennicke, A.; Araya, A.; Ferry, J.; et al. Gamma carbonic anhydrases in plant mitochondria. Plant Mol. Biol. 2004, 55, 193–207. [Google Scholar] [CrossRef]
- Vogler, M.; Karan, R.; Renn, D.; Vancea, A.; Vielberg, M.T.; Grötzinger, S.W.; DasSarma, P.; DasSarma, S.; Eppinger, J.; Groll, M.; et al. Crystal structure and active site engineering of a halophilic γ-carbonic anhydrase. Front. Microbiol. 2020, 11, 742. [Google Scholar] [CrossRef]
- Pena, K.L.; Castel, S.E.; de Araujo, C.; Espie, G.S.; Kimber, M.S. Structural basis of the oxidative activation of the carboxysomal γ-carbonic anhydrase, CcmM. Proc. Natl. Acad. Sci. USA 2010, 107, 2455–2460. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Zhang, Y.; Wang, L.; Jing, Q.; Wang, X.; Xi, X.; Zhao, X.; Wang, H. Molecular structure of thermostable and zinc-ion-binding γ-class carbonic anhydrases. BioMetals 2019, 32, 317–328. [Google Scholar] [CrossRef] [PubMed]
- Hatherley, R.; Brown, D.K.; Glenister, M.; Bishop, Ö.T. PRIMO: An interactive homology modeling pipeline. PLoS ONE 2016, 11, e0166698. [Google Scholar] [CrossRef] [Green Version]
- Eswar, N.; Webb, B.; Marti-Renom, M.A.; Madhusudhan, M.S.; Eramian, D.; Shen, M.Y.; Pieper, U.; Sali, A. Comparative protein structure modeling using Modeller. Curr. Protoc. Bioinform. 2006, 15, 5–6. [Google Scholar] [CrossRef] [Green Version]
- Cukuroglu, E.; Gursoy, A.; Keskin, O. HotRegion: A database of predicted hot spot clusters. Nucleic Acids Res. 2012, 40, D829–D833. [Google Scholar] [CrossRef] [PubMed]
- Darnell, S.J.; LeGault, L.; Mitchell, J.C. KFC Server: Interactive forecasting of protein interaction hot spots. Nucleic Acids Res. 2008, 36, W265–W269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sukhwal, A.; Sowdhamini, R. PPCheck: A webserver for the quantitative analysis of protein-protein interfaces and prediction of residue hotspots. Bioinform. Biol. Insights 2015, 9, BBI-S25928. [Google Scholar] [CrossRef]
- Gordon, J.C.; Myers, J.B.; Folta, T.; Shoja, V.; Heath, L.S.; Onufriev, A. H++: A server for estimating pKas and adding missing hydrogens to macromolecules. Nucleic Acids Res. 2005, 33 (Suppl. 2), W368–W371. [Google Scholar] [CrossRef]
- Sanyanga, T.A.; Nizami, B.; Tastan Bishop, Ö. Mechanism of Action of Non-Synonymous Single Nucleotide Variations Associated with α-Carbonic Anhydrase II Deficiency. Molecules 2019, 24, 3987. [Google Scholar] [CrossRef] [Green Version]
- Case, D.A.; Belfon, K.; Ben-Shalom, I.; Brozell, S.R.; Cerutti, D.; Cheatham, T.; Cruzeiro, V.W.; Darden, T.; Duke, R.E.; Giambasu, G.; et al. Amber 2020. 2020. Available online: https://ambermd.org/doc12/Amber20.pdf (accessed on 14 March 2021).
- Abraham, M.J.; Murtola, T.; Schulz, R.; Páll, S.; Smith, J.C.; Hess, B.; Lindahl, E. GROMACS: High performance molecular simulations through multi-level parallelism from laptops to supercomputers. SoftwareX 2015, 1, 19–25. [Google Scholar] [CrossRef] [Green Version]
- Schafmeister, C.; Ross, W.; Romanovski, V. LEaP; University of California: San Francisco, CA, USA, 1995. [Google Scholar]
- Maier, J.A.; Martinez, C.; Kasavajhala, K.; Wickstrom, L.; Hauser, K.E.; Simmerling, C. ff14SB: Improving the accuracy of protein side chain and backbone parameters from ff99SB. J. Chem. Theory Comput. 2015, 11, 3696–3713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Humphrey, W.; Dalke, A.; Schulten, K. VMD: Visual molecular dynamics. J. Mol. Graph. 1996, 14, 33–38. [Google Scholar] [CrossRef]
- Williams, T.; Kelley, C.; Broker, H.B.; John Campbell, R.; Cunningham, D.; Denholm, G.; Elber, R.; Fearick, C.; Grammes, L.; Hart, L.; et al. Gnuplot 5.2. 2: An Interactive Plotting Program; 2017; Available online: http://www.gnuplot.info (accessed on 25 March 2021).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CA Abbreviation | Organism Name | Organism Taxonomic Family | CA Accession Number | CA Sequence Length | Query Sequence | Query Coverage | Sequence Identity | E-Value |
|---|---|---|---|---|---|---|---|---|
| γ-CmCA | Caminibacter mediatlanticus | Nautiliaceae | WP_007473220.1 | 179 aa | γ-CmCA | 100% | 100% | 9 × 10−129 |
| γ-EpCA | Epsilonproteobacteria bacterium 4484_65 | -- | OQX76055.1 | 174 aa | γ-CmCA | 97% | 68.6% | 4 × 10−84 |
| γ-HtCA | Hydrogenimonas thermophila | Hydrogenimonaceae | WP_092914162.1 | 178 aa | γ-CmCA | 99% | 69.1% | 1 × 10−88 |
| γ-LnCA | Lebetimonas natsushimae | Nautiliaceae | WP_096258534.1 | 179 aa | γ-CmCA | 100% | 91.1% | 3 × 10−117 |
| γ-NpCA | Nautilia profundicola | Nautiliaceae | WP_012663737.1 | 179 aa | γ-CmCA | 100% | 88.3% | 1 × 10−114 |
| γ-NsbCA | Nitratiruptor sp. SB155-2 | Nitratiruptoraceae | WP_012081704.1 | 178 aa | γ-CmCA | 99% | 69.1% | 2 × 10−87 |
| γ-NtCA | Nitratiruptor tergarcus | Nitratiruptoraceae | WP_084274664.1 | 178 aa | γ-CmCA | 100% | 71.5% | 1 × 10−92 |
| γ-SaCA | Sulfurimonas autotrophica | Thiovulacea | WP_013326385.1 | 174 aa | γ-CmCA | 97% | 70.7% | 4 × 10−86 |
| γ-SlCA | Sulfurovum lithotrophicum | Sulfurovacea | WP_046550225.1 | 174 aa | γ-CmCA | 97% | 68.4% | 4 × 10−84 |
| γ-SNbcCA | Sulfurovum sp. NBC37-1 | Sulfurovacea | WP_012083921.1 | 174 aa | γ-CmCA | 97% | 68.4% | 1 × 10−83 |
| CA | Residues |
|---|---|
| Cam | Interface AB Chain A: F8, N10, R12, E13, N14, P15, V16, T17, P18, W19, P36, Q37, A38, S39, I41, M55, A56, S57, R59, D61, E62, G77, V79, H81, L83, E84, E88, Q118, S119, Q120, H122, Q136, A137, F138, F140, R153, S154, A155, I157, G170, V172, T174 Chain B: F8, Y33, D35, P36, Q37, S53, P54, M55, Q75, D76, G77, H117, Q118, M135, Q136, P152, R153, A169, G170, Y191, T194, A197, V198, V201, N202, H204, L205, G208, Y209, E211, T212 Interface BC Chain B: F8, N10, R12, E13, N14, P15, V16, T17, P18, W19, P36, Q37, A38, S39, I41, M55, A56, S57, R59, D61, E62, G77, V79, H81, L83, E84, E88, Q118, S119, Q120, H122, Q136, A137, F138, F140, R153, S154, A155, I157, G170, V172, T174 Chain C: F8, Y33, D35, P36, Q37, M51, S53, P54, M55, Q75, D76, G77, H117, Q118, M135, Q136, P152, R153, A169, G170, Y191, T194, A197, V198, V201, N202, H204, L205, G208, Y209, E211, T212 Interface AC Chain A: F8, Y33, D35, P36, Q37, M51, S53, P54, M55, Q75, D76, G77, H117, Q118, M135, Q136 P152, R153, A169, G170, Y191, T194, A197, V198, Y200, V201, N202, H204, L205, G208, Y209, E211, T212 Chain C: F8, S9, N10, R12, E13, N14, P15, V16, T17, P18, W19, P36, Q37, A38, S39, I41, M55, A56, S57, R59, D61, E62, G77, V79, H81, L83, E84, E88, Q118, S119, Q120, H122, Q136, A137, F138, F140, R153, S154, A155, I157, G170, V172, T174 |
| EcoCA-γ | Interface DE Chain D: D22, D23, S24, W40, P41, L42, D62, G63, H91, K92, M108, G109, A126, G127, N167, Y168, W171, E174, Y175 Chain E: H6, P7, Y8, R9, S24, V26, I28, L42, V44, R46, D48, V49, G63, M65, H67, H70, K92, V93, M94, H96, I111, L113, L129, S145 Interface EF Chain E: D22, S24, W40, P41, L42, Q61, D62, G63, H91, K92, M108, G109, A126, G127, S164, N167, Y168, W171, E174, Y175, Q178 Chain F: P7, Y8, R9, S24, V26, I28, L42, V43, V44, R46, D48, V49, G63, M65, H67, H70, K92, V93, M94, H96, I111, L113, L129, S145 Interface DF Chain D: P7, Y8, R9, S24, S25, V26, I28, L42, V43, V44, R46, D48, V49, G63, M65, H67, H70, Y74, K92, V93, M94, H96, I111, L113, L129, S145 Chain F: D22, S24, W40, P41, L42, D62, G63, H91, K92, M108, G109, A126, G127, N167, Y168, W171, E174, Y175, Q178 |
| γ-CmCA | Interface AB Chain A: I2, L3, R4, Y5, K6, D7, D23, I25, G39, V41, R43, D45, V46, M60, S61, M62, H64, H67, K69, K70, R91, V92, M93, H95, S108, A109, T110, L112, G126, L128 Chain B: P20, S21, W37, F38, G39, D59, M60, H90, R91, M107, S108, A125, G126, L142, P145, Q162, K165, N166, Y167, N169, Y170, D173, Y174, F177, I178, R179 Interface BC Chain B: I2, L3, R4, Y5, K6, S21, D23, I25, G39, V41, R43, D45, V46, S61, M62, H64, H67, K69, K70, K72, R91, V92, M93, H95, S108, A109, T110, L112, D113, L128, S144 Chain C: P20, S21, W37, F38, D59, M60, H90, R91, M107, S108, A125, G126, L142, E155, K159, N166, Y167, Y170, D173, Y174, F177, I178, R179 Interface AC Chain A: S21, W37, F38, G39, D59, M60, H90, R91, M107, S108, A125, L142, S144, P145, K159, Q162, K165, N166 Y167, Y170, D173, Y174, F177, I178, R179 Chain C: M1, I2, L3, R4, Y5, K6, D7, N8, F9, S21, D23, I25, G39, V41, R43, D45, V46, M60, S61, M62, H64, H67, K70, R91, V92, M93, H95, A109, T110, I111, L112, D113, L128, Q131, S144, P145 |
| γ-EpCA | Interface AB Chain A: L3, K4, F5, K6, S23, I25, G39, V41, R43, D45, V46, L60, S61, M62, H64, H67, R90, V91, M92, H94, S107, T109, L111, I110, D112, L127, S143, P144 Chain B: W37, F38, G39, D59, L60, M62, H89, R90, M106, S107, A124, G125, M141, G142 R165, Y166, F169, E172, Y173 Interface BC Chain B: M1, L3, K4, F5, K6, S23, I25, G39, V41, R43, D45, V46, L60, S61, M62, I63, H64, V65, T66, H67, R90, M92, H94, S107, A108, T109, L111, L127, S143, P144 Chain C: G21, W37, F38, G39, Q58, D59, L60, H89, R90, M106, S107, A124, M141, R165, Y166, F169, E172, Y173 Interface AC Chain A: W37, F38, D59, L60, H89, R90, M106, S107, A124, G125, M141 R165, Y166, F169, E172, Y173 Chain C: L3, F5, K6, S23, I25, G39, V41, R43, D45, V46, L60, S61, M62, H64, T66, H67, H68, R90, V91, M92, H94, S107, A108, T109, L111, D112, G125, L127, S143 |
| γ-HtCA | Interface AB Chain A: L2, L3, R4, Y5, K6, D7, F9, T23, I25, G39, V41, R43, D45, V46, L60, T61, M62, H64, T66, H67, Y68, K69, R90, V91, M92, H94, N107, C108, T109, L111, D112, L127, S143, P144 Chain B: P20, D21, W37, F38, D59, L60, H89, R90, M106, N107 A124, I141, R165, Y166, F169, D172, Y173, N175, F176, L177, S178 Interface BC Chain B: M1, L2, L3, R4, Y5, K6, A22, T23, I25, G39, V41, R43, D45, V46, L60, T61, M62, H64, T66, H67, Y68, K69, R90, V91, M92, H94, N107, C108, T109, L111, G125, L127, S143, P144 Chain C: P20, D21, W37, F38, G39, D59, L60, H89, R90, M106, N107, A124, G125, I141, A161, R165, Y166, F169, D172, Y173, F176, L177, S178 Interface AC Chain A: P20, D21, W37, F38, D59, L60, H89, R90, M106, N107, A124, G125, I141, G142, R164, R165, Y166, F169, D172, Y173, F176, L177, S178 Chain C: L2, L3, R4, Y5, K6, F9, T23, I25, G39, V41, R43, D45, V46, L60, T61, M62, H64, T66, H67, K69, R90, V91, M92, H94, N107, C108, T109, I110, L111, G125, L127, S143, P144 |
| γ-LnCA | Interface AB Chain A: M1, I2, L3, R4, Y5, K6, A22, D23, I25, G39, V41, R43, D45, V46, L60, S61, V92, M93 H95, R91 M62, H64, H67, S108, A109, T110, L112, L128, S144 Chain B: P20, S21, W37, F38, G39, D59, L60, H90, R91, M107, S108, A125, G126 N166, Y167, Y170, E173, Y174, F177, I178, K179 Interface BC Chain B: I2, L3, R4, Y5, K6, D23, I25, G39, V41, R43, D45, V46, L60, S61, M62, H64, H67, F68, K70, R91, V92, M93, H95, S108, A109, T110, L112, D113, G126, L128, S144 Chain C: S21, W37, F38, G39, D59, L60, H90, R91, M107, S108, A125, G126, L142 E155, E156, K159, Q162, N166, Y167, Y170, E173, Y174, F177, I178, K179 Interface AC Chain A: S21, W37, F38, D59, L60, H90, R91, M107, S108, A125, G126, L142, G143, N166, Y167, Y170, E173, Y174, F177, I178, K179 Chain C: I2, L3, R4, Y5, K6, D23, I25, G39, V41, R43, D45, V46, L60, S61, M62, H64, H67, R91, V92, M93, H95 S108, T110, I111, L112, D113, L128, S144, P145 |
| γ-NpCA | Interface AB Chain A: T2, L3, R4, Y5, K6, D7, D23, I25, G39, V41, R43, D45, M60, S61, M62, H64, H67, R91, V92 M93, H95, S108, T110, L112, D113, G126, L128, S144 Chain B: P20, S21, W37, F38, G39, Q58, D59, M60, H90, R91, M107, S108, A125 G126, L142, G143, K162, N166, Y167, Y170, D173, Y174, F177, V178, R179 Interface BC Chain B: M1, T2, L3, R4, Y5, K6, A22, D23, I25, G26, G39, V41, R43, D45, V46, M60, S61, M62, H64, T66, H67, E69, K70, R91, V92 M93, H95, S108, A109, T110, L112, D113, L128, S144 Chain C: W17, P20, S21, W37, F38, G39, D59, M60, H90, R91, A125 G126, G143, N159, K162, N163, N166, Y167, Y170, D173, Y174, F177, V178, R179 Interface AC Chain A: P20, S21, W37, F38, G39, D59, M60, H90, R91, M107, S108, A125, G126, L142, G143, P145, K162, N166 Y167, Y170, D173, Y174, F177, V178, R179 Chain C: T2, L3, R4, Y5, K6, D23, I25, G39, V41, R43, D45, V46, M60, M62, H64, T66, H67, E69, R91, V92 M93, H95, S108, A109, T110, L112, D113, G126, L128, S144, P145 |
| γ-NsbCA | Interface AB Chain A: L2, L3, R4, Y5, K6, F9, T23, I25, G39, V41, R43, D45, V46, L60, T61, M62, H64, T66, H67, Y68 R90, V91, M92, H94, N107, S108, T109, L111, D112, G125, L127, S143, P144 Chain B: P20, D21, W37, F38, G39, D59, L60, H89, R90, M106, A124, G125 L141, G142, E158, R165, Y166, F169, E172, Y173, N175, S176, I177, S178 Interface BC Chain B: L2, L3, R4, Y5, K6, E7, F9, T23, I25, G39, V41, R43, D45, V46, L60, 61, M62, H64, V65, T66, H67, Y68, R90, V91, M92, H94, N107, S108, T109, L111, G125, L127, S143, P144 Chain C: P20, D21, W37, F38, G39, D59, L60, H89, R90, M106, N107, A124, G125, L141, E158, A161, S162, R165, Y166, F169, E172, Y173, S176, I177, S178 Interface AC Chain A: W17, P20, D21, W37, F38, G39, D59, L60, H89, R90, M106, N107, I121, A124, G125, G142, E154, E155, E158, A161, R165, Y166, F169, E172, Y173, S176, I177, S178 Chain C: L2, L3, R4, Y5, K6, F9, T23, I25, G39, V41, R43, D45, V46, L60, T61, M62, H64, T66, H67, Y68, K70, R90, V91, M92, H94, N107, S108, T109, L111, L127, S143 |
| γ-NtCA | Interface AB Chain A: L2, L3, R4, Y5, K6, F9, T23, V25, G39, V41, R43, D45, V46, L60, S61, M62, H64, T66, H67, K69, R90, V91, M92, H94, N107, S108, T109, L111, D112, L127, T143, P144 Chain B: W17, P20, D21, W37, F38, G39, D59, L60, H89, R90, M106, N107, A124, G125, L141, G142, Q157, R165, Y166, F169, E172, Y173, F176, I177, Q178 Interface BC Chain B: M1, L2, L3, R4, Y5, K6, E7, F9, T23, V25, G39, V41, R43, D45, V46, L60, S61, M62, H64, T66, H67, Y68, R90, M92, H94, N107, S108, T109, L111, D112, G125, L127, T143, P144 Chain C: P20, D21, A22, W37, F38, G39, D59, L60, H89, R90, M106, N107, A124, G125, L141, G142, E158, R165, Y166, F169, E172, Y173, D175, F176, I177, Q178 Interface AC Chain A: P20, D21, W37, F38, G39, D59, L60, H89, R90, M106, N107, A124, G125, G142, K154, K164, R165, Y166, F169, E172, Y173, F176, I177, Q178 Chain C: L2, L3, R4, Y5, K6, F9, T23, V25, G39, V41, R43, D45, V46, L60, S61, M62, H64, T66, H67, K69, P71, D72, R90, V91, M92, H94, N107, T109, L111, D112, G125, L127, T143 |
| γ-SaCA | Interface AB Chain A: H3, V4, Y5, K6, D23, I25, G39, V41, R43, D45, V46, L60, S61, M62, H64, T66, H67, Y68, D75, R90, V91, M92, H94, S107, A108, T109, L111, D112, G125, L127, K130, T143, P144 Chain B: P20, S21, W37, F38, G39, D59, L60, H89, R90, M106, S107, A124, G125, M141, G142, E158, A161, S162, R165, Y166, F169, D172, Y173 Interface BC Chain B: L2, H3, V4, Y5, K6, D23, I25, G39, V41, R43, D45, V46, L60, S61, M62, H64, H67, K69, D75, R90, V91, M92, H94, S107, A108, T109, L111, D112, G125, L127, T143, P144 Chain C: P20, S21, W37, F38, G39, D59, L60, H89, R90, M106, S107, A124, G125, M141, G142, R165, Y166, F169, D172, Y173 Interface AC Chain A: P20, S21, W37, F38, G39, D59, L60, H89, R90, M106, S107, A124, G125, M141, G142, R165, Y166, F169, D172, Y173 Chain C: H3, V4, Y5, K6, D23, I25, G39, V41, R43, D45, V46, L60, S61, M62, H64, V65, T66, H67, R90, V91, M92, H94, S107, A108, T109, L111, D112, G125, L127, T143, P144 |
| γ-SlCA | Interface AB Chain A: M1, L3, K4, F5, K6, S23, I25, G39, V41, R43, D45, V46, L60, S61, M62, H64, V65, T66, H67, K69, R90, V91, M92, H94, S107, A108, T109, L111, D112, G125, L127, K130, S143, P144 Chain B: E20, G21, W37, F38, G39, D59, L60, H89, R90, M106, S107, A124, G125, M141, E158, A161, S162, R165, Y166, F169, E172, Y173 Interface BC Chain B: M1, L3, K4, F5, K6, S23, I25, G39, V41, R43, D45, V46, L60, S61, M62, H64, H67, H68, R90, V91, M92, H94, S107, A108, T109, I110, L111, D112, G125, L127, S143, P144 Chain C: G21, W37, F38, G39, D59, L60, H89, R90, M106, S107, A124, G125, M141, R165, Y166, F169, E172, Y173 Interface AC Chain A: E20, G21, W37, F38, G39, D59, L60, H89, R90, M106, S107, A124, G125, M141, E158, R165, Y166, F169, E172, Y173 Chain C: M1, L3, F5, K6, S23, I25, G39, V41, R43, D45, V46, L60, S61, M62, H64, H67, R90, V91, M92, H94, S107, A108, T109, L111, D112, G125, L127, K130, S143, P144 |
| γ-SNbcCA | Interface AB Chain A: I2, L3, K4, F5, K6, S23, I25, G39, V41, R43, D45, V46, L60, S61, M62, H64, H67, K69, R90, V91, M92, H94, S107, A108, T109, I110, L111, G125, L127, K130, S143, P144 Chain B: G21, W37, F38, G39, D59, L60, H89, R90, M106, S107, A124, G125, L127, K130, M141, G142, R165, Y166, F169, E172, Y173 Interface BC Chain B: M1, L3, K4, F5, K6, S23, I25, G39, V41, R43, D45, V46, L60, S61, M62, I63, H64, T66, H67, H68, K69, K70, R90, M92, H94, S107, T109, I110, L111, D112, S143, P144 Chain C: G21, W37, F38, G39, Q58, D59, L60, H89, R90, V91, M92, H94, M106, S107, A124, G125, M141, E154, E155, A157, E158, A161, R165, Y166, F169, E172, Y173 Interface AC Chain A: E20, G21, W37, F38, G39, Q58, D59, L60, H89, R90, M106, S107, A124, G125, M141, G142, R165, Y166, F169, E172, Y173 Chain C: L3, K4, F5, K6, S23, I25, G39, V41, R43, D45, V46, L60, S61, M62, H64, V65, H67, S107, T109, L111, D112, L127, S143 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Manyumwa, C.V.; Tastan Bishop, Ö. Gamma Carbonic Anhydrases from Hydrothermal Vent Bacteria: Cases of Alternating Active Site Due to a Long Loop with Proton Shuttle Residue. Catalysts 2021, 11, 1177. https://doi.org/10.3390/catal11101177
Manyumwa CV, Tastan Bishop Ö. Gamma Carbonic Anhydrases from Hydrothermal Vent Bacteria: Cases of Alternating Active Site Due to a Long Loop with Proton Shuttle Residue. Catalysts. 2021; 11(10):1177. https://doi.org/10.3390/catal11101177
Chicago/Turabian StyleManyumwa, Colleen Varaidzo, and Özlem Tastan Bishop. 2021. "Gamma Carbonic Anhydrases from Hydrothermal Vent Bacteria: Cases of Alternating Active Site Due to a Long Loop with Proton Shuttle Residue" Catalysts 11, no. 10: 1177. https://doi.org/10.3390/catal11101177
APA StyleManyumwa, C. V., & Tastan Bishop, Ö. (2021). Gamma Carbonic Anhydrases from Hydrothermal Vent Bacteria: Cases of Alternating Active Site Due to a Long Loop with Proton Shuttle Residue. Catalysts, 11(10), 1177. https://doi.org/10.3390/catal11101177