1. Introduction
Population structure refers to consistent demographic differences among individuals as a function of some other attribute, such as geographic location, age, size, gender or physiological state. Over the last few years, structured population models have become a central modelling formalism in theoretical biology and game dynamics, as well as being one of the most widely used.
An important question is the effect of migration on evolution. Nagylaki [
1] studied the strong migration limit in a geographically-structured population, which occurs when migration dominates all other evolutionary forces in the limit of a large population, by means of a diffusion approximation. He considers a finite number of demes represented by the integers
. Deme
i is composed of
diploid individuals considered at a single multi-allelic locus, for
. Here,
N and
denote the whole population size and the proportion of deme
i in the whole population, for
, with
. Time is discrete with non-overlapping generations, and the reproduction scheme in each deme follows the Wright–Fisher model as a result of random mating. Following the production of a very large number of offspring and selection among offspring, there is migration. The probability that an individual in deme
i comes from deme
j is represented by
, for
. The backward migration matrix
is assumed to be constant and ergodic. Following migration and mutation, there is random sampling within demes to restore the deme sizes. In the limit of a large population size (
), the stochastic dynamics in this structured population is described by a Wright–Fisher diffusion as in a well-mixed population with an effective population size
taken as the unit of time, where:
Here,
is the stationary distribution associated with the backward migration matrix
. Under the same assumptions, but for a haploid population and in the absence of selection, Notohara [
2] showed that the genealogical process, known in this context as the structured coalescent (Herbots [
3]), is described in the limit of a large population size by the standard Kingman coalescent [
4,
5,
6], which is such that each pair of lineages coalesces backward in time at rate one independently of all others.
Diffusion approximations and genealogical processes are very important tools to address questions related to the effect of selection on the evolution of strategies in game dynamics. Among these questions, how cooperation can emerge and persist from interactions between individuals is of prime interest. This question has attracted increasing attention from mathematical or theoretical biologists (Axelrod and Hamilton [
7], Szabò and Töke [
8], Traulsen
et al. [
9], Santos and Pacheco [
10], Hauert and Szabò [
11], Nowak [
12,
13], Nowak and Sigmund [
14], Ohtsuki
et al. [
15], Szabò and Fáth [
16]). In order to study this question, a simple game named the prisoner’s dilemma has been considered. The simplest form of this game has payoffs in additive form with the following parameters: a donor pays a cost
c to a recipient to get a benefit
b, where
.
The first studies on the evolution of cooperation in structured populations assumed symmetric interactions between all members of the population. This means that the payoffs depend only on the strategies used by the interacting individuals (Hamilton [
17,
18], Trivers [
19], Frank [
20], Nowak and Sigmund [
21,
22], Nowak [
23], Traulsen and Nowak [
24], Kroumi and Lessard [
25]). Even in this case, interactions can form a complex system as in the set-structured population introduced in Tarnita
et al. [
26]: every individual of the population belongs to exactly
K sets among
M sets, and a cooperator cooperates only with individuals that belong to at least
L of the
K sets, and defects otherwise.
In living groups, repetitive interactions for access to limited resources or mating opportunities can lead to the creation of a social order. A hierarchy dominance that may depend on differences in resource holding power (Hammerstein [
27], Wilson [
28]) can be established so that individuals are dominant over those below them and submissive to those above them. This motivates the study of asymmetric interactions with a cost for not conforming to the established hierarchy in structured populations with linear or cyclic dominance (Tao
et al. [
29], Kroumi and Lessard [
30]).
In this paper, we consider a Moran-type model for games played in a population structured into
d colonies of different finite sizes. In pairwise interactions, the individuals can adopt one of two strategies,
or
. An individual from colony
i interacts with an individual from colony
j with probability
for
, where
for
. The expected payoff that an individual receives determines the probability for this individual to produce an offspring. One offspring is produced at a time, and following migration, this offspring replaces one individual locally chosen at random. More precisely, if the offspring is produced in colony
i and migrates to colony
j with probability
, then the offspring replaces an individual chosen at random in colony
j, for
. Finally, there is mutation of the strategy used by each individual independently of all others with probability
u from one time step to the next, and when this occurs, the mutant strategy is chosen at random among
and
. We will find a condition for
to be more abundant on average than
at equilibrium in the limit of a large population under weak selection and weak mutation. This result relies on the strong migration limit of the genealogical process in the absence of selection, which is proven using a lemma for two-time scale Markov chains due to Möhle [
31]. The condition will be express in terms of expected weighted payoffs using reproductive values as weights, which is an alternative to the use of structure coefficients (Nowak
et al. [
32]) for games in structured populations. This will allow us to give an intuitive interpretation of this condition.
The remainder of this paper is organized as follows. We present the details of the model in
Section 2. We use a two-time scale convergence result that is established in
Appendix A to derive the limiting genealogical process in a neutral structured population in
Section 3. The equilibrium state under weak selection is studied in
Section 4. In
Section 5, a condition for a weighted average equilibrium frequency of
to increase as the selection intensity increases from zero is deduced. This condition is applied to situations with linear or cyclic dominance hierarchy in
Section 6 and to games in set-structured populations in
Section 7. The results are interpreted and discussed in
Section 8.
2. Model
We assume a population subdivided into
d colonies represented by the integers
. Each colony is made of a finite number
of haploid individuals, for
. Each individual in the population adopts one of two strategies,
or
. We assume pairwise interactions between individuals within each colony and between individuals from different colonies. More precisely, an individual from colony
i interacts with an individual chosen at random from colony
j with probability
, for
, with
, for
. Then, the payoff that the individual from
i receives is given by the entries of the matrix:
according to the strategy adopted by the individual from
i, corresponding to row
or
, and the strategy used by the individual from
j, corresponding to column
or
. The expected payoffs of strategies
and
played by individuals in colony
i are denoted by
and
, respectively. It is assumed that these expected payoffs translate into fertilities, or reproductive successes, in the form:
and:
respectively, where
represents the intensity of selection. It is assumed throughout the paper that selection is weak, actually that the intensity of selection
s is small compared to the inverse of the population size
, where:
The case
corresponds to neutrality.
Time is discrete. At each time step, an individual is chosen in the whole population with probability proportional to its fertility to produce an offspring. This offspring inherits the strategy used by its parent. If the parent is from colony i, then the offspring stays in colony i with probability or migrates to colony with probability , with , for . In both cases, the offspring replaces an individual chosen at random in the same colony. It is assumed throughout that the forward migration matrix is irreducible and aperiodic, that is ergodic. In other words, there exists some power of this migration matrix, for some integer , with all positive entries. Finally, strategy mutation occurs with probability u for each individual independently of all others, so that the strategy used by the individual at the next time step is chosen at random among and with probability u and remains the same with the complementary probability .
3. Genealogical Process in the Neutral Model
In this section, we derive the genealogical process of a sample taken from the population structured into d colonies under the neutral model in the limit of a large population size. Every individual of the population has the same fertility, which is given by one.
Consider a sample of size
at a given time step. Looking backward in time at the genealogy of this sample, the distribution of the ancestors in the
d colonies at any previous time step can be described by a vector:
where
denotes the number of ancestors in colony
i, for
. Then:
is the total number of ancestors.
Let
be the distribution of the ancestors
time steps back. Given an initial sample of
n individuals, this is a discrete-time Markov chain with state space:
Let
be the transition probability from state
to state
. A possible transition is from
to
, for
, such that
. Here,
is a
d-dimensional unit vector with the
i-th component equal to one and all other components equal to zero. This transition is obtained if one of the
ancestors in colony
i produced an offspring who stayed in colony
i and is one of the
other ancestors in this colony, or if one of the
ancestors in colony
produced an offspring who migrated to colony
i and is one of the
ancestors in this colony. All this occurs with probability:
Here,
is the proportion of colony
i in the whole population, for
. Another possible transition is from
to
, for
, such that
and
. This occurs if an individual in colony
j other than the
ancestors in this colony produced an offspring who migrated to colony
i and is one of the
ancestors in this colony. The probability of this event is:
The last transition with positive probability is to stay in the same state, for which we have:
From Equations (
9)–(
11), the transition matrix
can be decomposed into the form:
Here,
is an identity matrix of a size given by the number of elements in the state space
, and
is an infinitesimal generator whose non-null entries are given by:
Moreover, the non-null entries of
are given by:
Now, let
be the subset of all possible states with
k ancestors, namely:
for
. Note that the set
is the disjoint union of the subsets
,
,
. With respect to these subsets in this order, the matrices
and
whose non-null entries
and
are given by Equations (
13) and (
14), respectively, can be expressed in the block forms:
and:
Here,
denotes a zero matrix of any dimension.
The exponential matrix
for every integer
takes the block form:
Note that
is the infinitesimal generator of an irreducible Markov chain on a finite state space, which is
, for
. Consequently, the limit matrix:
exists for
, and so does:
Note also that
is actually a rank one matrix with the stationary distribution associated with
in every row, for
. It remains to find this distribution.
Let
be the stationary distribution associated with
. By definition, this distribution satisfies the equation:
with:
Note that
, where
is the backward migration matrix, whose entries are given by:
This is obtained from Equation (
13) in the case where
for
. The entry
is the probability that an individual chosen at random in colony
i comes from colony
j one time step back. Moreover,
which means that
is the stationary distribution associated with the backward migration matrix
. The stationary distribution associated with
, for
, can be expressed with respect to this distribution. For
, we have:
where:
The proof of Equation (
25) is given in
Appendix B.
By definition of the stationary distribution
, we have:
for
, from which:
Owing to Proposition 1 in
Appendix A, we conclude that:
where
and
denotes the floor value of
, which is defined as the greatest integer less or equal to the real number
. Using the block forms of
and
given in Equations (
17) and (
20), respectively, we have:
The non-null entries of
are given by:
for
for
. We have:
The first term on the right-hand side of Equation (
32) is:
while the second term is:
These expressions into Equation (
32) give:
where:
Finally, using the fact that:
the non-null entries of
given in Equation (
32) take the form:
for
for
. Note that
for
and
. In summary, we have the following result in the limit of a large population size.
Theorem 1.
The strong migration limit of the genealogical process in a structured population, taking time steps as the unit of time as the population size , is given by:for , where has non-null entries given by:for , for . Remark 1 Equation (39) means that, in the limit as for , the ancestors are in state with probability as long as their number is k, while this number decreases by one at rate per time steps, for . In other words, after a scaled time of exponential distribution with parameter , the number of ancestors jumps from k to , and these ancestors are found in state , with probability for . Note that the number of ancestors is described by the standard Kingman coalescent in a well-mixed population (see Kingman [4,5,6]). Remark 2 The limiting process for the number of ancestors in the structured population of size N corresponds to the limiting process for the Moran model (see Moran [33,34]) in a well-mixed population of size with time steps as the unit of time as , where λ is given in Equation (36). The parameter λ is a measure of mixing, and is an effective population size that takes into account the population structure. 4. Equilibrium State
Suppose without loss of generality that the individuals in the population occupy ordered sites, such that the sites of colony 1 come first, then the sites of colony 2 come second, and so on, up to the sites of colony
d. The state of the population at a given time step is represented by the
N-dimensional vector
, where:
for
. With
being the probability for an individual from colony
i to interact with an individual chosen at random from colony
j, for
, and assuming that an individual can interact with itself, the expected payoffs of strategies
and
in colony
i for
are given by:
and:
respectively, where:
is the frequency of
in colony
j, for
(with the convention that
).
An offspring is produced according to the corresponding fertilities
and
given in Equations (
3) and (
4). This offspring migrates from colony
i to colony
j with probability
and replaces an individual chosen at random in colony
j, for
. Moreover, the strategy of each individual mutates into a strategy chosen at random among
and
with probability
u, and this occurs independently for all individuals. Then, the conditional expected value of the new state of the population takes the form:
where
is an all-ones
N-dimensional vector, while:
where
is a diagonal matrix with the vector:
on the main diagonal. Here,
denotes an all-ones
-matrix and
an all-ones
-dimensional vector. Moreover,
is the probability for a given individual in colony
i to be replaced by an offspring produced by an individual playing
in colony
j, if there is any, while:
is the total probability for a given individual in colony
i to be replaced by an offspring.
Note that:
and:
are the average fertilities in colony
j and in the whole population, respectively, where:
and:
are the corresponding average expected payoffs. Therefore,
and:
with:
and:
Note that the stochastic matrix:
has a stationary distribution given by:
with
T for transpose, where
is the stationary distribution of the backward migration matrix
defined in Equation (
23). Moreover,
and:
where
stands here for a matrix or a vector whose entries or components are functions little-
o with respect to
s as
.
At equilibrium, we have:
The scalar product with
yields:
Note that this equilibrium equation in the neutral case (
with
denoting expectation under this condition) gives:
from which:
Under weak selection, it follows from Equations (
60) and (
63) that:
This equation gives an approximation in the case of weak selection, that is for
small enough.
5. Condition for Weak Selection to Favour a Strategy over Another
In this section, we will prove the main result below.
Theorem 2.
Strategy is favoured by weak selection in the strong migration limit of a structured population with payoff matrices given by (2) for strategies and under weak mutation, in the sense that is more abundant than in expected weighted average equilibrium frequency for a weak enough intensity of selection, if:with being the proportion of individuals in colony i, the probability that an offspring migrates from colony j to colony i, the stationary proportion of ancestors of an individual that are in colony i in the neutral model and the probability for an individual in colony j to interact with an individual in colony k.
Proof.
The equilibrium Equation (
66) implies that:
Note that:
is a weighted expected frequency of strategy
at equilibrium. If the intensity of selection
is small enough, then:
if:
This is a condition for weak selection to favour
in the sense that strategy
is more abundant in weighted average frequency at equilibrium than strategy
. If the inequality is reversed, then weak selection favours
in the same sense.
With the assumptions of the model, the vector
takes the expression given in Equation (
59) and:
where
and
are given in Equations (
52) and (
53), respectively. Therefore, we have:
Moreover, Equation (24) entails that:
with the last equality obtained by permuting the indices
i and
j. Therefore, the condition for weak selection to favour
becomes:
Using the expressions given in Equations (
42) and (
43) for the expected payoffs of
and
in colony
j, we find that:
In the neutral model, a permutation of strategies
and
does not change the expected value of a product of their equilibrium frequencies. Consequently, we have:
and:
Moreover, in the strong migration limit with
time steps as the unit of time as
and under weak mutation, so that
remains constant, the above expected values are all equal to the probability that exactly two given individuals out of three, irrespective of the colonies that they are in, use the same strategy (see
Appendix C). Ignoring common positive factors and writing
as
, we get the condition given in Equation (
67) for weak selection to favour
, which completes the proof. □
7. Application to Games in Set-Structured Populations
Tarnita
et al. [
26] consider a population composed of
N individuals and
M sets. They assume that each individual belongs exactly to
K sets, which corresponds to a phenotype. Interactions occur only between individuals within the same sets (individuals interact as many times as the number of sets to which they both belong). With
M sets numbered
, every individual
l is represented by a
M-dimensional vector
. Here,
if individual
l belongs to set
i and zero otherwise, for
, with exactly
K components equal to one and
components equal to zero. The individuals represented by the same vector
belong to the same colony represented by this vector. Here, there are
colonies. We assume that an offspring inherits the
K sets of his parent represented by
with probability
and chooses
K sets represented by
with probability
. These are phenotype mutation probabilities. The number of interactions between two individuals represented by
and
, respectively, is given by:
Therefore,
where
is the proportion of colony
. Two strategies are in use, represented by
and
. The payoff matrix for
i in
against
j in
is denoted by
.
Suppose that phenotype mutation occurs at random, so that
for every couple of phenotypes
. Then, the backward matrix takes the form:
The stationary distribution of this matrix is given by:
Therefore, the condition given in Equation (
67) for
to be favoured by weak selection becomes:
Written in the form:
the condition means that the expected payoff of
exceeds the expected payoff of
when the expected frequencies of
and
are equal among the individuals of the same phenotype for every phenotype.
Suppose now that an
-individual uses strategy
with an opponent only if the two individuals belong to at least
L common sets, where
is a fixed constant, and uses
otherwise. On the other hand, an
-individual always uses strategy
. In this case, the payoff matrix for an individual in colony
against an individual in colony
takes the form:
if
, and:
if
(note that the meaning of
R is different here from the previous section). Moreover, we have:
In this case, the condition given in Equation (
67) for
to be favoured by weak selection reduces to:
which is the same as:
This condition, known as risk dominance in a coordination game (Harsanyi and Selten [
39]), does not depend on the population structure.
8. Discussion
Our main result (Theorem 2) for
to be favoured by weak selection over
in a structured population under weak strategy mutation in the limit of a large population with payoffs in pairwise interactions depending on the locations of the players is given by Equation (67), which can be written in the form:
Here,
is the limiting proportion of time back that a single lineage spends in colony
i in the absence of selection, for
. It represents the expected contribution of colony
i to the whole population in the long run forward in time, called its reproductive value, under the neutral model. With
representing the proportion of colony
i and
the probability for an offspring from colony
i to migrate to colony
j for
, the quantity
represents an expected relative reproductive value of an offspring produced by an individual in colony
i. On the other hand, every individual interacts with an
-individual and with an
-individual with the same probability
in a neutral population at equilibrium, since then, the probability that any given individual uses strategy
or
is equal to the probability that the most recent mutant ancestor of this individual used strategy
or
, which is
in each case from the assumptions on strategy mutation. We have the same approximate probability 1/2 in an equilibrium population under weak selection. With
being the probability that an individual chosen at random in the whole population belongs to colony
j and
, the probability for an individual from colony
j to interact with an individual from colony
k, for
, the left-hand side of Equation (
104) can be interpreted as the expected payoff of
weighted by relative reproductive values of offspring in an equilibrium population near neutrality. The right-hand side of Equation (
104) has a similar interpretation for
, and the inequality guarantees that
is more abundant on average than
at equilibrium near neutrality if individuals are weighted by their relative reproductive values.
This interpretation is very intuitive. It is an alternative to the use of structure coefficients (Nowak
et al. [
32]) for games in structured populations. Moreover, this interpretation suggests an effective payoff matrix (Lessard [
40]):
where
is the payoff matrix for an individual from colony
j in interaction with an individual from colony
k. This means that the game in the structured population is equivalent to a game in a well-mixed population with this matrix as the payoff matrix.
This result has been obtained for a structured population reproducing according to a Moran model with one individual replaced at a time in the strong migration limit as the population size
N tends to infinity. We have shown (Theorem 1) that the genealogical process in the neutral model, which is described by the transition matrix in Equation (
12) from one time step to the previous one, where
and
are given in
Section 3, tends to the standard Kingman coalescent (Kingman [
6]) for the number of ancestors if
time steps are taken as the unit of time where λ is given by Equation (
36), while the ancestors are distributed independently among the colonies according to the stationary distribution
. The proof (
Appendix A) relies on a two-time scale argument and uses a lemma due to Möhle [
31]. A similar result was proven in a different way in Notohara [
2] in the case of a subdivided population that reproduces according to a Wright–Fisher model.
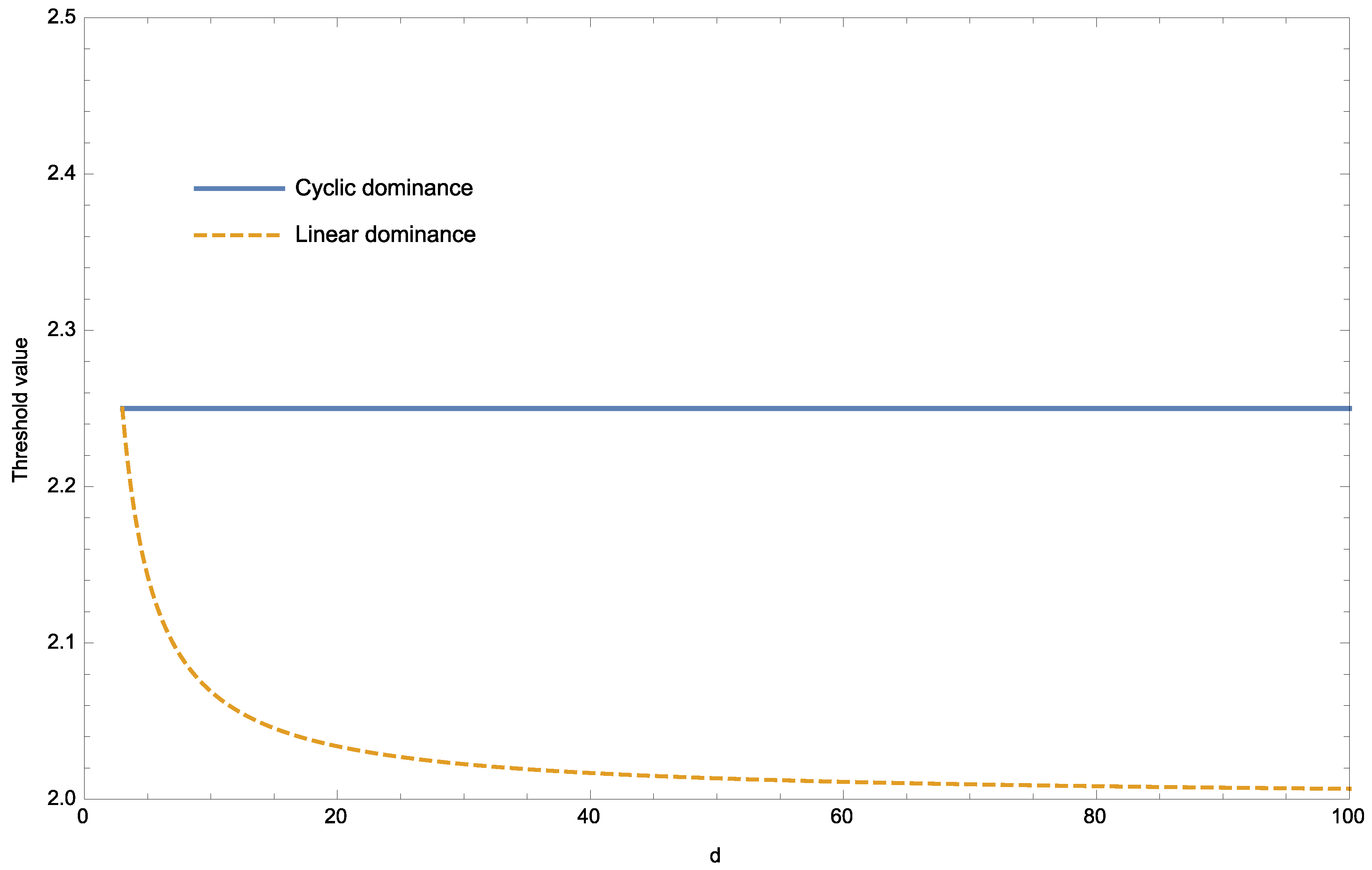
Our main result has been applied to the situation of dominance hierarchy with d colonies in decreasing order of dominance in the case of linear dominance and in counter-clockwise order of dominance in the case of cyclic dominance. Individuals in a given colony can interact with individuals in all other colonies in the case of linear dominance, but only with individuals in the same colony or in the two adjacent colonies in the case of cyclic dominance. Considering the strategies TFT and AllD in a repeated additive prisoner’s dilemma and a cost for defection against a dominant defector, it has been shown that linear dominance is more favourable than cyclic dominance for increasing the expected frequency of TFT at equilibrium as soon as . This has been obtained under the assumptions of colonies of the same size with uniform or symmetric migration and random interactions.
Another application concerns the set-structured population as introduced in Tarnita
et al. [
26], but with colonies of fixed relative sizes and reproduction according to a Moran model instead of a Wright–Fisher model. With uniform mutation from one subset of sets to another of the same size, which defines the phenotype of an individual, the condition for
to be favoured by weak selection is that, for an individual chosen at random in the whole population, the expected payoff of
exceeds the expected payoff of
near neutrality. With strategy
actually used only if the number of common sets to which the two players belong exceeds some threshold, it has been shown that the condition for
to be favoured by weak selection reduces to a condition known as risk dominance (Harsanyi and Selten [
39]) as in a well-mixed population. Note that the same result was obtained in Tarnita
et al. [
26] in the case of a high rate of phenotype mutation, which corresponds to strong migration from one phenotype to another.


{kind=link}
{kind=link}
{kind=link}