Effects of Ellagic Acid on Angiogenic Factors in Prostate Cancer Cells




 ,
,  ,
, 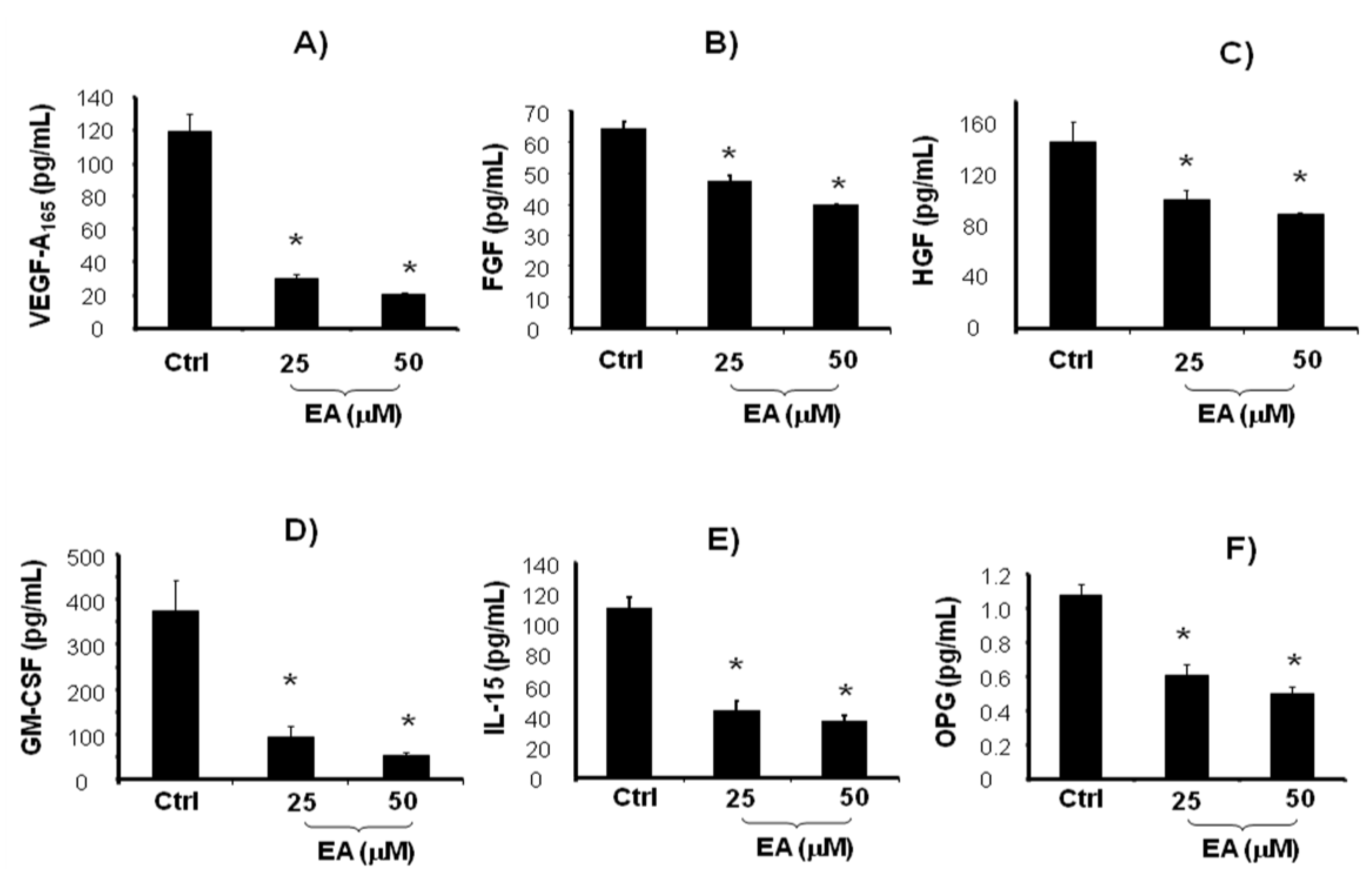{kind=link}
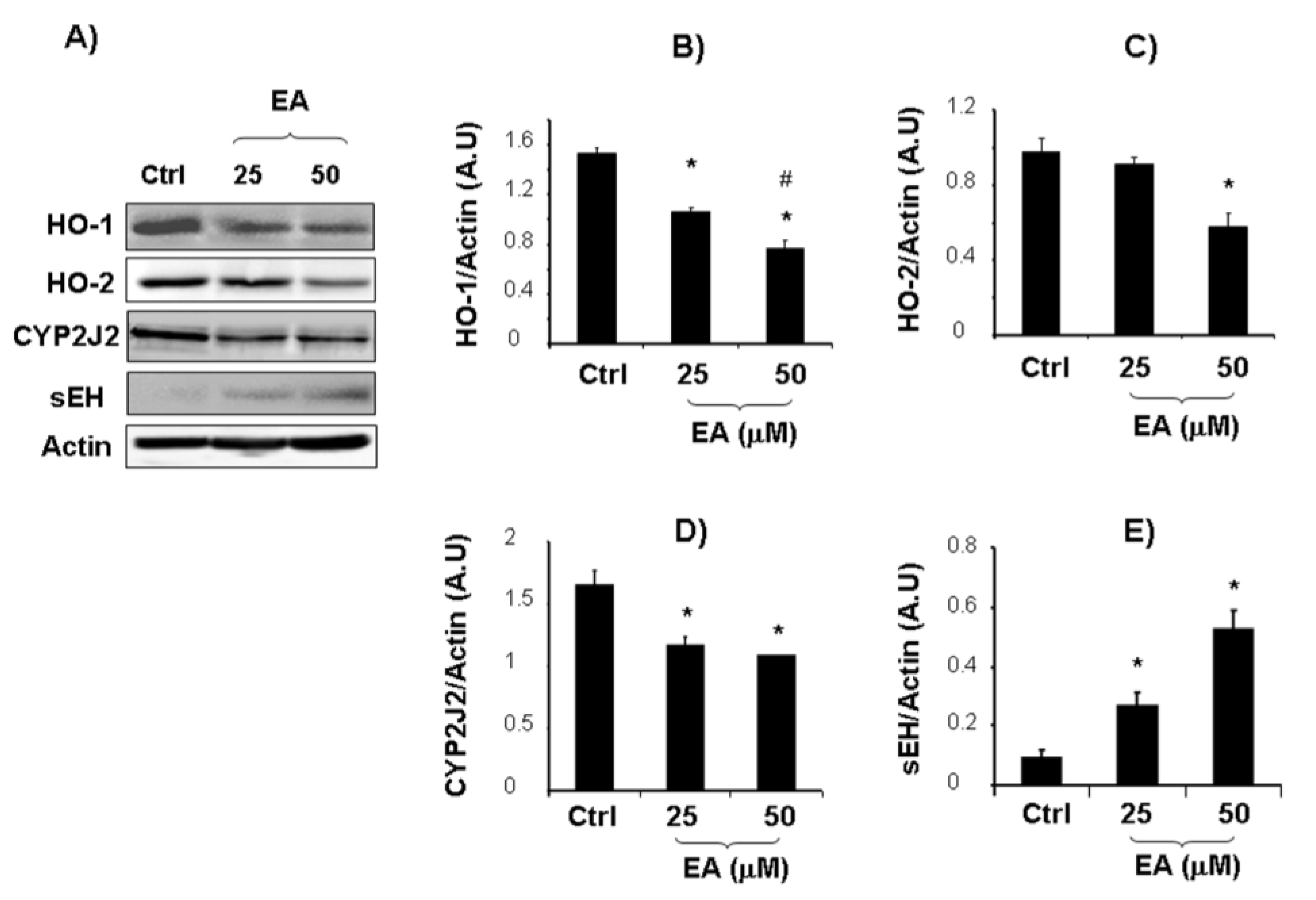{kind=link}
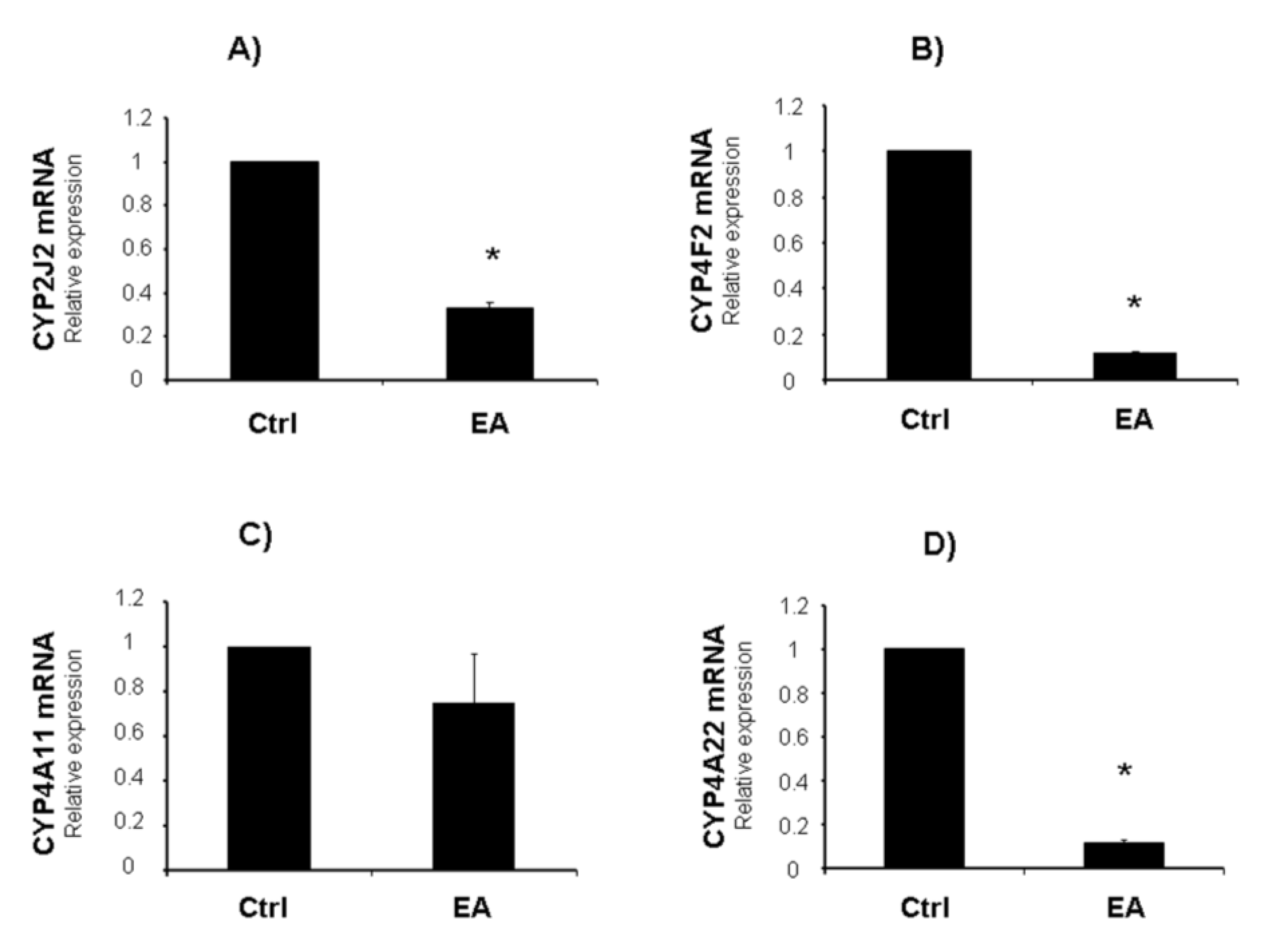{kind=link}
Abstract
Share and Cite
Vanella, L.; Di Giacomo, C.; Acquaviva, R.; Barbagallo, I.; Li Volti, G.; Cardile, V.; Abraham, N.G.; Sorrenti, V. Effects of Ellagic Acid on Angiogenic Factors in Prostate Cancer Cells. Cancers 2013, 5, 726-738. https://doi.org/10.3390/cancers5020726
Vanella L, Di Giacomo C, Acquaviva R, Barbagallo I, Li Volti G, Cardile V, Abraham NG, Sorrenti V. Effects of Ellagic Acid on Angiogenic Factors in Prostate Cancer Cells. Cancers. 2013; 5(2):726-738. https://doi.org/10.3390/cancers5020726
Chicago/Turabian StyleVanella, Luca, Claudia Di Giacomo, Rosaria Acquaviva, Ignazio Barbagallo, Giovanni Li Volti, Venera Cardile, Nader G. Abraham, and Valeria Sorrenti. 2013. "Effects of Ellagic Acid on Angiogenic Factors in Prostate Cancer Cells" Cancers 5, no. 2: 726-738. https://doi.org/10.3390/cancers5020726
APA StyleVanella, L., Di Giacomo, C., Acquaviva, R., Barbagallo, I., Li Volti, G., Cardile, V., Abraham, N. G., & Sorrenti, V. (2013). Effects of Ellagic Acid on Angiogenic Factors in Prostate Cancer Cells. Cancers, 5(2), 726-738. https://doi.org/10.3390/cancers5020726