
Thyroid Cancer—The Tumor Immune Microenvironment (TIME) over Time and Space


{kind=link}
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
3. Results
3.1. Single-Cell Profiling
3.2. Immune Checkpoints Analysis
Programmed Cell Death Protein 1/Programmed Cell Death Ligand 1
3.3. Myeloid-Derived Cells
3.4. Macrophages
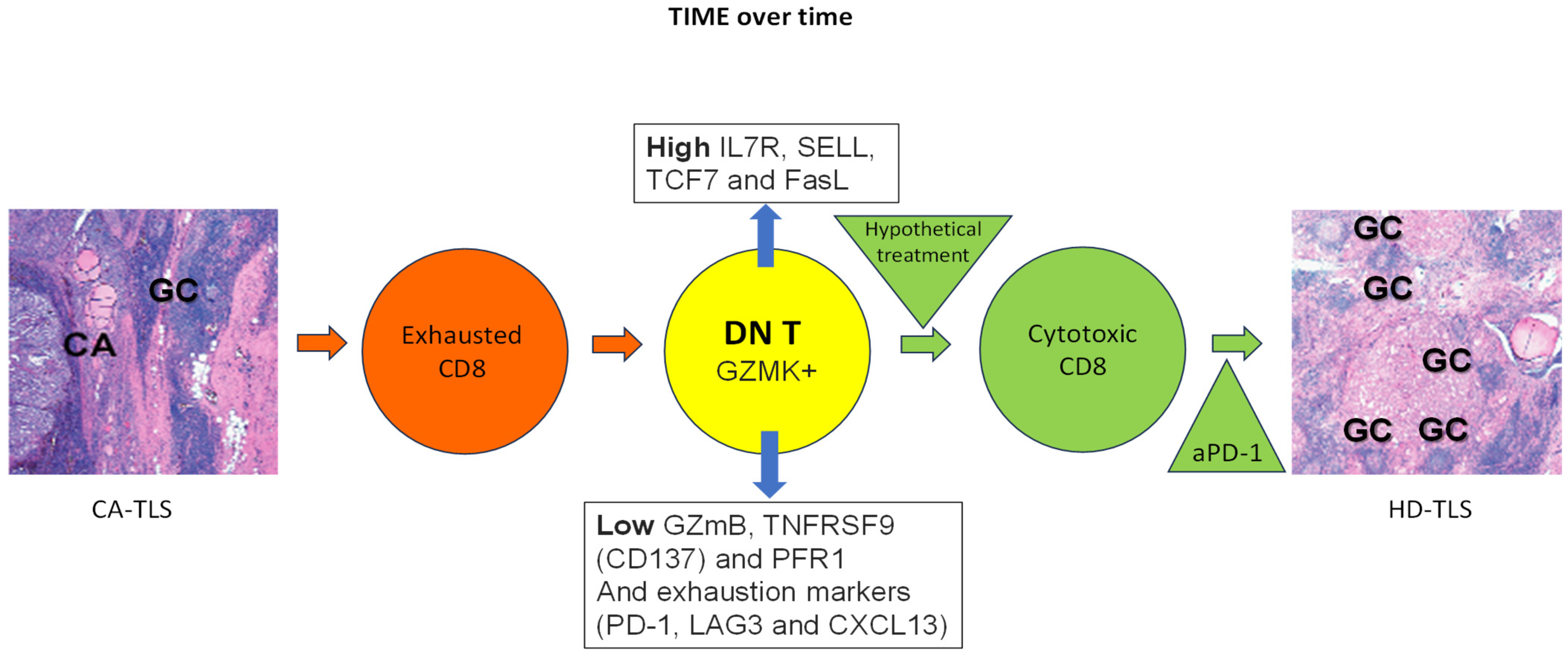
3.5. Double-Negative T Cells
4. Discussion
5. Conclusions
Funding
Data Availability Statement
Conflicts of Interest
References
- de Visser, K.E.; Eichten, A.; Coussens, L.M. Paradoxical roles of the immune system during cancer development. Nat. Rev. Cancer 2006, 6, 24–37. [Google Scholar] [CrossRef] [PubMed]
- Prehn, R.T.; Main, J.M. Immunity to methylcholanthrene-induced sarcomas. J. Natl. Cancer Inst. 1957, 18, 769–778. [Google Scholar]
- Klebanoff, C.A.; Gattinoni, L.; Restifo, N.P. CD8+ T-cell memory in tumor immunology and immunotherapy. Immunol. Rev. 2006, 211, 214–224. [Google Scholar] [CrossRef]
- Turk, M.J.; Guevara-Patino, J.A.; Rizzuto, G.A.; Engelhorn, M.E.; Houghton, A.N. Concomitant tumor immunity to a poorly immunogenic melanoma is prevented by regulatory T cells. J. Exp. Med. 2004, 200, 771–782. [Google Scholar] [CrossRef] [PubMed]
- Romero, P.; Rod Dunbar, P.; Valmori, D.; Pittet, M.; Ogg, G.S.; Rimoldi, D.; Chen, J.-L.; Liénard, D.; Cerottini, J.-C.; Cerundolo, V. Ex vivo staining of metastatic lymph nodes by class I major histocompatibility complex tetramers reveals high numbers of antigen-experienced tumor-specific cytolytic T lymphocytes. J. Exp. Med. 1998, 188, 1641–1650. [Google Scholar] [CrossRef]
- Valmori, D.; Scheibenbogen, C.; Dutoit, V.; Nagorsen, D.; Asemissen, A.M.; Rubio-Godoy, V.; Rimoldi, D.; Guillaume, P.; Romero, P.; Schadendorf, D.; et al. Circulating Tumor-reactive CD8+ T cells in melanoma patients contain a CD45RA+CCR7− effector subset exerting ex vivo tumor-specific cytolytic activity. Cancer Res. 2002, 62, 1743–1750. [Google Scholar] [PubMed]
- Schreiber, R.D.; Old, L.J.; Smyth, M.J. Cancer Immunoediting: Integrating Immunity’s Roles in Cancer Suppression and Promotion. Science 2011, 331, 1565–1570. [Google Scholar] [CrossRef] [PubMed]
- Jaume, J.C. Endocrine autoimmunity. In Greenspan’s Basic & Clinical Endocrinology, 9th ed.; Gardner, D.G., Shoback, D.M., Eds.; McGraw-Hill Medical: New York, NY, USA, 2017; pp. 29–48. [Google Scholar]
- Korman, A.; Yellin, M.; Keler, T. Tumor immunotherapy: Preclinical and clinical activity of anti-CTLA4 antibodies. Curr. Opin. Investig. Drugs 2005, 6, 582–591. [Google Scholar]
- Gogas, H.; Ioannovich, J.; Dafni, U.; Stavropoulou-Giokas, C.; Frangia, K.; Tsoutsos, D.; Panagiotou, P.; Polyzos, A.; Papadopoulos, O.; Stratigos, A.; et al. Prognostic significance of autoimmunity during treatment of melanoma with interferon. N. Engl. J. Med. 2006, 354, 709–718. [Google Scholar] [CrossRef]
- Balkwill, F.; Charles, K.A.; Mantovani, A. Smoldering and polarized inflammation in the initiation and promotion of malignant disease. Cancer Cell 2005, 7, 211–217. [Google Scholar] [CrossRef] [PubMed]
- Imam, S.; Dar, P.; Paparodis, R.; Almotah, K.; Al-Khudhair, A.; Hasan, S.A.; Salim, N.; Jaume, J.C. Nature of coexisting thyroid autoimmune disease determines success or failure of tumor immunity in thyroid cancer. J. Immunother. Cancer 2019, 7, 3. [Google Scholar] [CrossRef]
- Paparodis, R.; Imam, S.; Todorova-Koteva, K.; Staii, A.; Jaume, J.C. Hashimoto’s thyroiditis pathology and risk for thyroid cancer. Thyroid 2014, 24, 1107–1114. [Google Scholar] [CrossRef]
- Chen, Z.; Wang, Y.; Li, D.; Le, Y.; Han, Y.; Jia, L.; Yan, C.; Tian, Z.; Song, W.; Li, F.; et al. Single-Cell RNA Sequencing Revealed a 3-Gene Panel Predicted the Diagnosis and Prognosis of Thyroid Papillary Carcinoma and Associated With Tumor Immune Microenvironment. Front. Oncol. 2022, 12, 862313. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Song, W.; Li, Y.; Liu, Z.; Zhao, K.; Jia, L.; Wang, X.; Jiang, R.; Tian, Y.; He, X. Integrated analysis of tumor microenvironment features to establish a diagnostic model for papillary thyroid cancer using bulk and single-cell RNA sequencing technology. J. Cancer Res. Clin. Oncol. 2023, 149, 16837–16850. [Google Scholar] [CrossRef]
- Chen, W.; Yu, X.; Li, H.; Yuan, S.; Fu, Y.; Hu, H.; Liu, F.; Zhang, Y.; Zhong, S. Single-cell RNA-seq reveals MIF-(CD74 + CXCR4) dependent inhibition of macrophages in metastatic papillary thyroid carcinoma. Oral. Oncol. 2024, 148, 106654. [Google Scholar] [CrossRef]
- Feng, C.; Tao, Y.; Yu, C.; Wang, L.; Liu, X.; Cao, Y. Integrative single-cell transcriptome analysis reveals immune suppressive landscape in the anaplastic thyroid cancer. Cancer Gene Ther. 2023, 30, 1598–1609. [Google Scholar] [CrossRef]
- Ma, H.; Li, G.; Huo, D.; Su, Y.; Jin, Q.; Lu, Y.; Sun, Y.; Zhang, D.; Chen, X. Impact of Hashimoto’s thyroiditis on the tumor microenvironment in papillary thyroid cancer: Insights from single-cell analysis. Front. Endocrinol. 2024, 15, 1339473. [Google Scholar] [CrossRef]
- Pan, J.; Ye, F.; Yu, C.; Zhu, Q.; Li, J.; Zhang, Y.; Tian, H.; Yao, Y.; Zhu, M.; Shen, Y.; et al. Papillary Thyroid Carcinoma Landscape and Its Immunological Link With Hashimoto Thyroiditis at Single-Cell Resolution. Front. Cell Dev. Biol. 2021, 9, 758339. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Shi, J.; Li, L.; Zhou, X.; Zhang, H.; Zhang, X.; Wang, Y.; Liu, L.; Sheng, L. Single-Cell Transcriptome Analysis Reveals Inter-Tumor Heterogeneity in Bilateral Papillary Thyroid Carcinoma. Front. Immunol. 2022, 13, 840811. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, S.; Nahar, U.; Dahiya, D.; Mukherjee, S.; Dey, P.; Gupta, R.; Radotra, B.; Sachdeva, N.; Sood, A.; Bhadada, S.K.; et al. Role of cytotoxic T cells and PD-1 immune checkpoint pathway in papillary thyroid carcinoma. Front. Endocrinol. 2022, 13, 931647. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Sun, R.; Wei, T. Immune microenvironment in papillary thyroid carcinoma: Roles of immune cells and checkpoints in disease progression and therapeutic implications. Front. Immunol. 2024, 15, 1438235. [Google Scholar] [CrossRef] [PubMed]
- Boruah, M.; Agarwal, S.; Mir, R.A.; Choudhury, S.D.; Sikka, K.; Rastogi, S.; Damle, N.; Sharma, M.C. Unravelling the Reasons Behind Limited Response to Anti-PD Therapy in ATC: A Comprehensive Evaluation of Tumor-Infiltrating Immune Cells and Checkpoints. Endocr. Pathol. 2024, 35, 419–431. [Google Scholar] [CrossRef] [PubMed]
- Pani, F.; Yasuda, Y.; Rousseau, S.T.; Bermea, K.C.; Roshanmehr, S.; Wang, R.; Yegnasubramanian, S.; Caturegli, P.; Adamo, L. Preconditioning of the immune system modulates the response of papillary thyroid cancer to immune checkpoint inhibitors. J. Immunother. Cancer 2022, 10, e005538. [Google Scholar] [CrossRef] [PubMed]
- Rabold, K.; Zoodsma, M.; Grondman, I.; Kuijpers, Y.; Bremmers, M.; Jaeger, M.; Zhang, B.; Hobo, W.; Bonenkamp, H.J.; de Wilt, J.H.W.; et al. Reprogramming of myeloid cells and their progenitors in patients with non-medullary thyroid carcinoma. Nat. Commun. 2022, 13, 6149. [Google Scholar] [CrossRef]
- Zhang, P.; Guan, H.; Yuan, S.; Cheng, H.; Zheng, J.; Zhang, Z.; Liu, Y.; Yu, Y.; Meng, Z.; Zheng, X.; et al. Targeting myeloid derived suppressor cells reverts immune suppression and sensitizes BRAF-mutant papillary thyroid cancer to MAPK inhibitors. Nat. Commun. 2022, 13, 1588. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Li, X.J.; Gangadaran, P.; Jing, X.; Ahn, B.C. Tumor-associated macrophages as a potential therapeutic target in thyroid cancers. Cancer Immunol. Immunother. 2023, 72, 3895–3917. [Google Scholar] [CrossRef]
- Katoh, H.; Okamoto, R.; Yokota, M.; Naito, K.; Kikuchi, M.; Tokito, T.; Sangai, T.; Yamashita, K. CD163+ Tumor-Associated Macrophage Recruitment Predicts Papillary Thyroid Cancer Recurrence. J. Surg. Res. 2024, 303, 532–544. [Google Scholar] [CrossRef] [PubMed]
- Qin, L.; Chen, C.; Gui, Z.; Jiang, Y. IRX5’s influence on macrophage polarization and outcome in papillary thyroid cancer. Front. Oncol. 2024, 14, 1399484. [Google Scholar] [CrossRef]
- Ning, J.; Hou, X.; Hao, J.; Zhang, W.; Shi, Y.; Huang, Y.; Ruan, X.; Zheng, X.; Gao, M. METTL3 inhibition induced by M2 macrophage-derived extracellular vesicles drives anti-PD-1 therapy resistance via M6A-CD70-mediated immune suppression in thyroid cancer. Cell Death Differ. 2023, 30, 2265–2279. [Google Scholar] [CrossRef] [PubMed]
- Can, N.Y.; Ayturk, S.; Celik, M.; Sezer, Y.A.; Ozyilmaz, F.; Tastekin, E.; Sut, N.; Ustun, F.; Bulbul, B.Y.; Puyan, F.O.; et al. Histological perspective on the effects of tumor-associated macrophages in the tumor microenvironment surrounding papillary thyroid carcinoma. Pol. J. Pathol. 2016, 67, 332–344. [Google Scholar] [CrossRef] [PubMed]
- Stempin, C.C.; Geysels, R.C.; Park, S.; Palacios, L.M.; Volpini, X.; Motran, C.C.; Acosta Rodríguez, E.V.; Nicola, J.P.; Cheng, S.Y.; Pellizas, C.G.; et al. Secreted Factors by Anaplastic Thyroid Cancer Cells Induce Tumor-Promoting M2-like Macrophage Polarization through a TIM3-Dependent Mechanism. Cancers 2021, 13, 4821. [Google Scholar] [CrossRef]
- Fischer, K.; Voelkl, S.; Heymann, J.; Przybylski, G.K.; Mondal, K.; Laumer, M.; Kunz-Schughart, L.; Schmidt, C.A.; Andreesen, R.; Mackensen, A. Isolation and characterization of human antigen-specific TCRαβ+ CD4−CD8− double-negative regulatory T cells. Blood 2005, 105, 2828–2835. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.X.; Yang, L.; Young, K.J.; DuTemple, B.; Zhang, L. Identification of a previously unknown antigen-specific regulatory T cell and its mechanism of suppression. Nat. Med. 2006, 6, 782–789. [Google Scholar] [CrossRef]
- Merims, S.; Li, X.; Joe, B.; Dokouhaki, P.; Han, M.; Childs, R.W.; Wang, Z.Y.; Gupta, V.; Minden, M.D.; Zhang, L. Anti-leukemia effect of ex vivo expanded DNT cells from AML patients: A potential novel autologous T-cell adoptive immunotherapy. Leukemia 2011, 25, 1415–1422. [Google Scholar] [CrossRef]
- Imam, S.; Paparodis, R.; Sharma, D.; Jaume, J.C. Lymphocytic Profiling in Thyroid Cancer Provides Clues for Failure of Tumor Immunity. Endocr. Relat. Cancer 2014, 21, 505–516. [Google Scholar] [CrossRef]
- Imam, S.; Paparodis, R.D.; Rafiqi, S.I.; Ali, S.; Niaz, A.; Kanzy, A.; Tovar, Y.E.; Madkhali, M.A.; Elsherif, A.; Khogeer, F.; et al. Thyroid Cancer Screening Using Tumor-Associated DN T Cells as Immunogenomic Markers. Front. Oncol. 2022, 12, 891002. [Google Scholar] [CrossRef]
- Okamura, K.; Wang, L.; Nagayama, S.; Yamashita, M.; Tate, T.; Matsumoto, S.; Takamatsu, M.; Kitano, S.; Kiyotani, K.; Nakamura, Y. Characterization of double-negative T cells in colorectal cancers and their corresponding lymph nodes. Oncoimmunology 2024, 13, 2373530. [Google Scholar] [CrossRef]
- Zhu, C.; Dai, Y.; Zhang, H.; Ruan, Y.; Zhou, Y.; Dai, Y.; Fan, L.; Jia, T.; Lu, H.; Chen, Q. T cell exhaustion is associated with the risk of papillary thyroid carcinoma and can be a predictive and sensitive biomarker for diagnosis. Diagn. Pathol. 2021, 16, 84. [Google Scholar] [CrossRef]
- Xie, Z.; Li, X.; He, Y.; Wu, S.; Wang, S.; Sun, J.; He, Y.; Lun, Y.; Zhang, J. Immune Cell Confrontation in the Papillary Thyroid Carcinoma Microenvironment. Front. Endocrinol. 2020, 11, 570604. [Google Scholar] [CrossRef]
- Wu, Z.; Zheng, Y.; Sheng, J.; Han, Y.; Yang, Y.; Pan, H.; Yao, J. CD3+CD4−CD8− (Double-Negative) T Cells in Inflammation, Immune Disorders and Cancer. Front. Immunol. 2022, 13, 816005. [Google Scholar] [CrossRef] [PubMed]
- Prins, R.M.; Incardona, F.; Lau, R.; Lee, P.; Claus, S.; Zhang, W.; Black, K.L.; Wheeler, C.J. Characterization of defective CD4−CD8− T cells in murine tumors generated independent of antigen specificity. J. Immunol. 2004, 172, 1602–1611. [Google Scholar] [CrossRef] [PubMed]
- Tiberti, S.; Catozzi, C.; Croci, O.; Ballerini, M.; Cagnina, D.; Soriani, C.; Scirgolea, C.; Gong, Z.; He, J.; Macandog, A.D.; et al. GZMKhigh CD8+ T effector memory cells are associated with CD15high neutrophil abundance in non-metastatic colorectal tumors and predict poor clinical outcome. Nat. Commun. 2022, 13, 6752. [Google Scholar] [CrossRef] [PubMed]
- Santiago, L.; Castro, M.; Sanz-Pamplona, R.; Garzon, M.; Ramirez-Labrada, A.; Tapia, E.; Moreno, V.; Layunta, E.; Gil-Gómez, G.; Garrido, M.; et al. Extracellular granzyme a promotes colorectal cancer development by enhancing gut inflammation. Cell Rep. 2020, 32, 107847. [Google Scholar] [CrossRef]
- Paparodis, R.D.; Karvounis, E.; Bantouna, D.; Chourpiliadis, C.; Chourpiliadi, H.; Livadas, S.; Imam, S.; Jaume, J.C. Incidentally Discovered Papillary Thyroid Microcarcinomas Are More Frequently Found in Patients with Chronic Lymphocytic Thyroiditis Than with Multinodular Goiter or Graves’ Disease. Thyroid. 2020, 30, 531–535. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jaume, J.C. Thyroid Cancer—The Tumor Immune Microenvironment (TIME) over Time and Space. Cancers 2025, 17, 794. https://doi.org/10.3390/cancers17050794
Jaume JC. Thyroid Cancer—The Tumor Immune Microenvironment (TIME) over Time and Space. Cancers. 2025; 17(5):794. https://doi.org/10.3390/cancers17050794
Chicago/Turabian StyleJaume, Juan Carlos. 2025. "Thyroid Cancer—The Tumor Immune Microenvironment (TIME) over Time and Space" Cancers 17, no. 5: 794. https://doi.org/10.3390/cancers17050794
APA StyleJaume, J. C. (2025). Thyroid Cancer—The Tumor Immune Microenvironment (TIME) over Time and Space. Cancers, 17(5), 794. https://doi.org/10.3390/cancers17050794