
Correlation Between Neurocognitive Outcomes and Neuroaxonal Connectome Alterations After Whole Brain Radiotherapy: A Proof-of-Concept Study †

 ,
,  , , ,
, , ,  and
and
Simple Summary
Abstract
1. Introduction
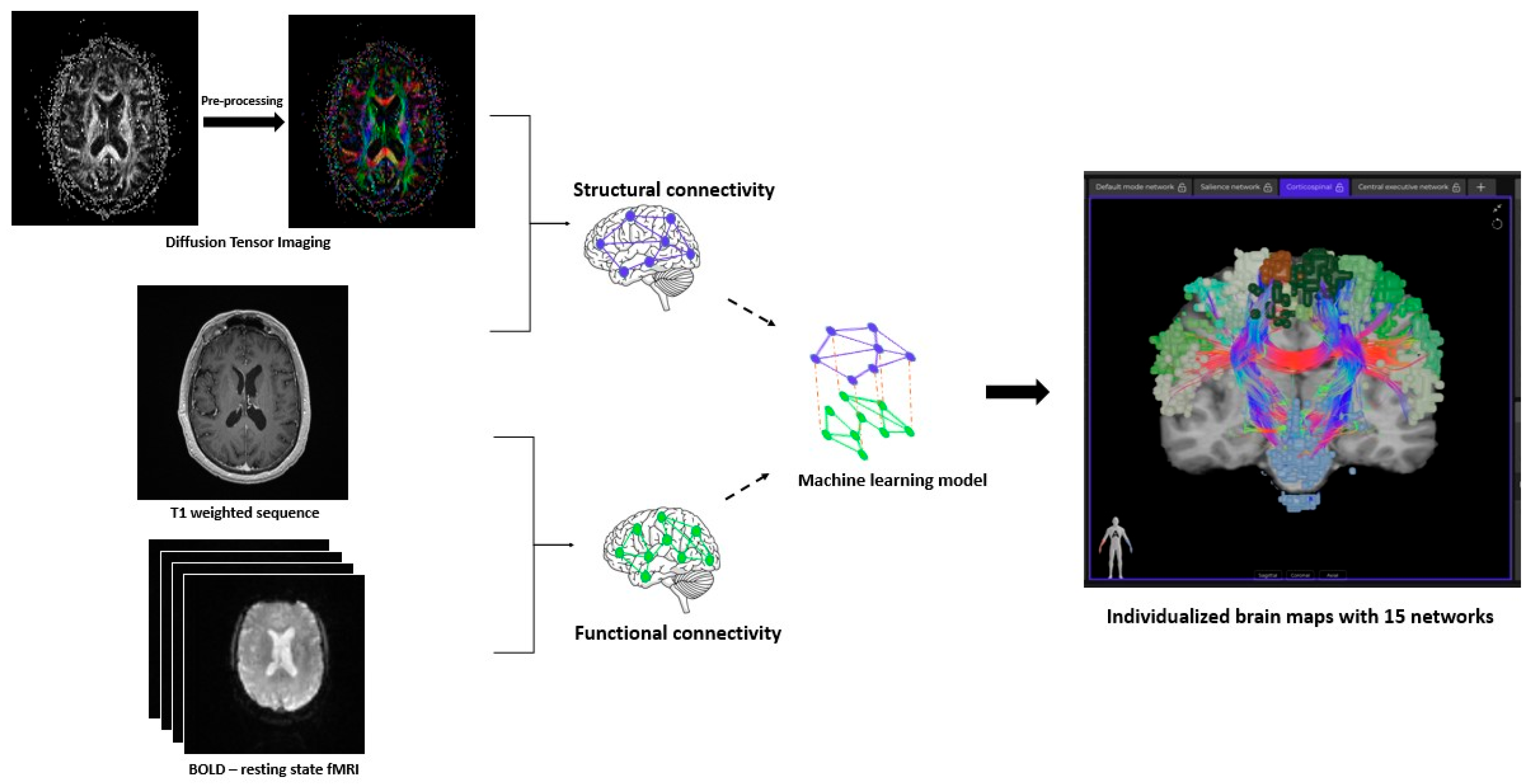
2. Materials and Methods
3. Results
3.1. Baseline Characteristics of Study Population
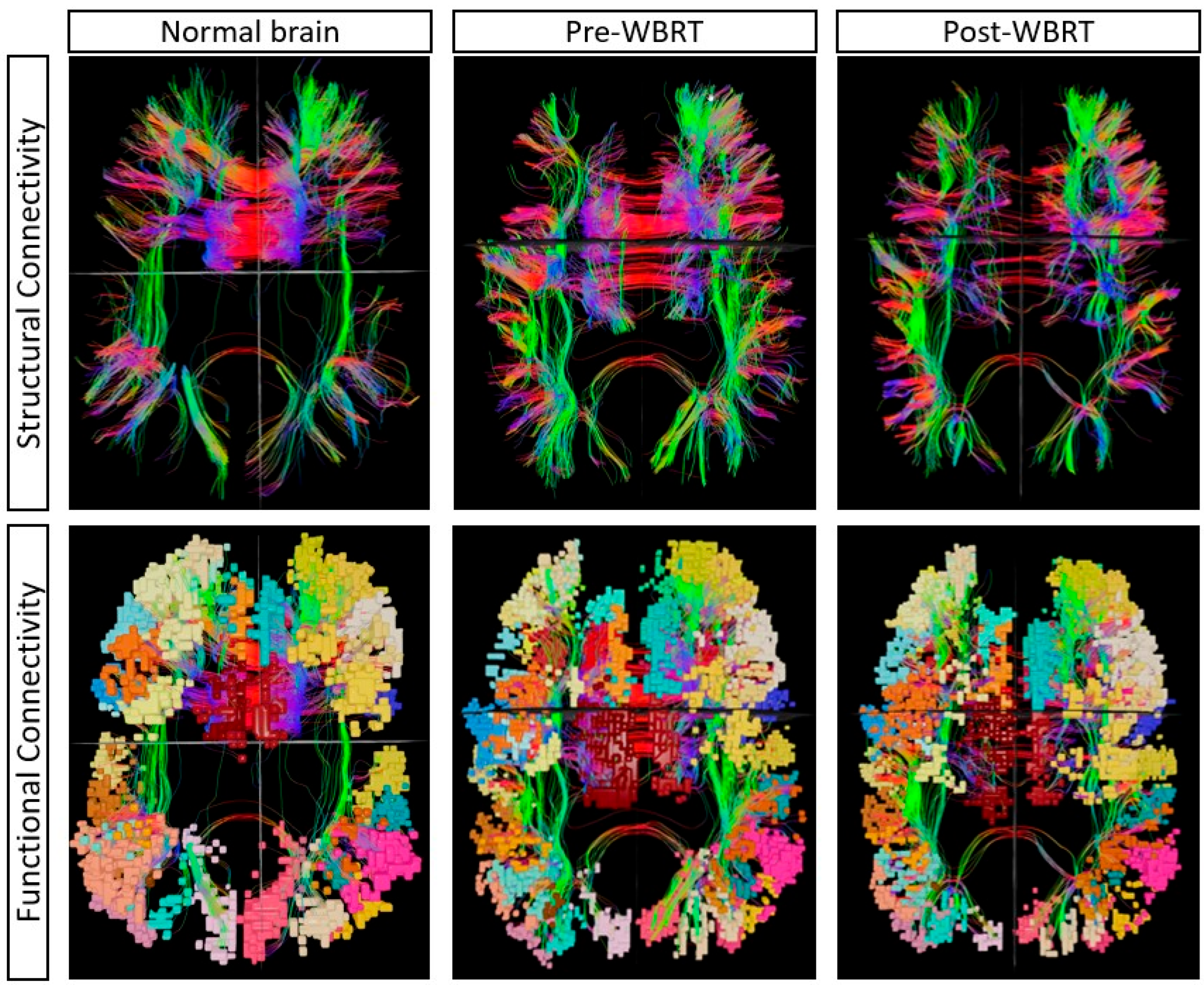
3.2. Baseline (Pre-WBRT) Connectome Evaluation
3.3. Post-WBRT Comparison
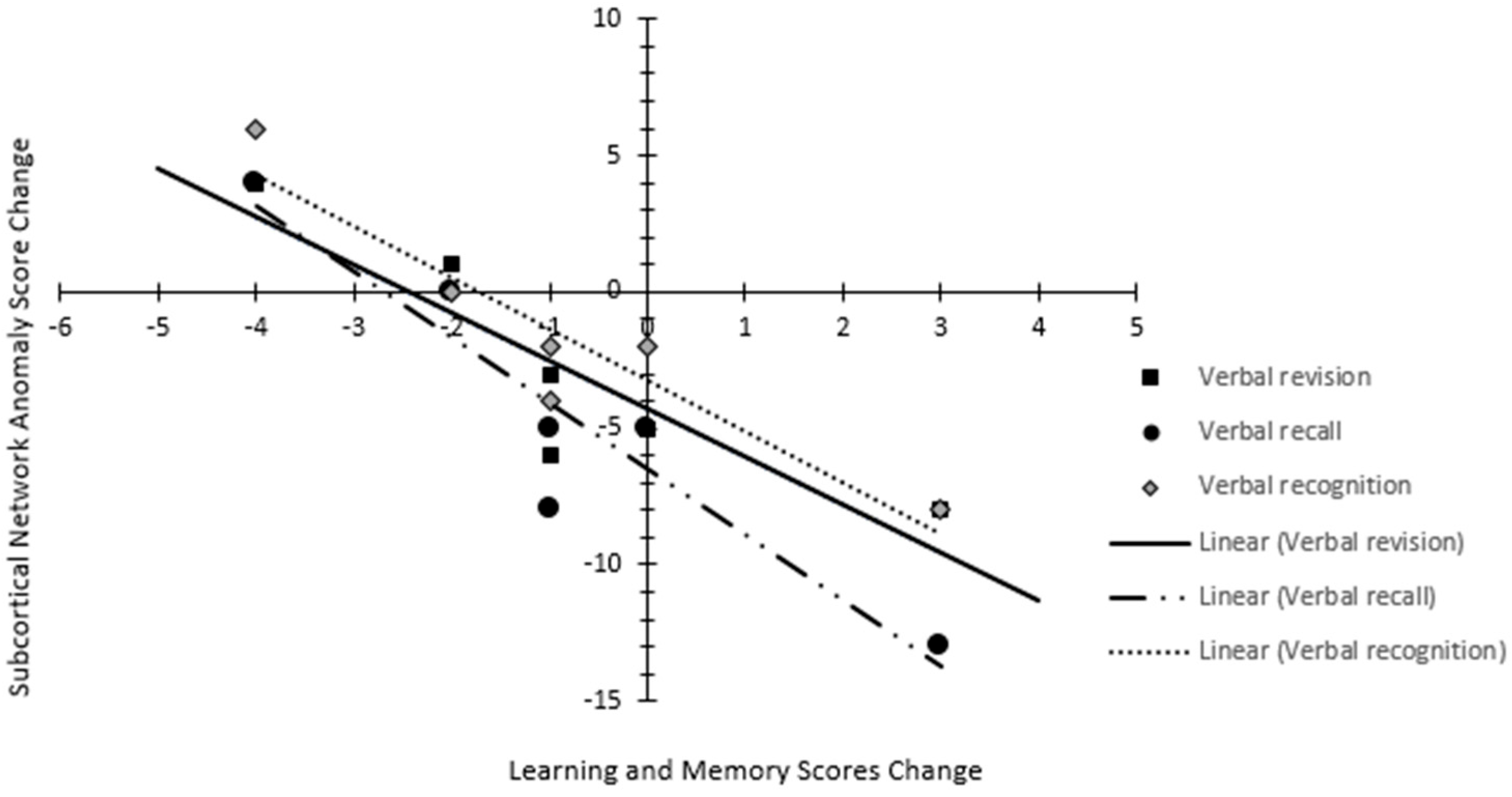
3.4. Neurocognitive Outcomes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| WBRT | Whole brain radiotherapy |
| DTI | Diffusion tensor imaging |
| fMRI | Functional magnetic resonance imaging |
| HCP | Human Connectome Project |
| MRI | Magnetic resonance imaging |
| DICOM | Digital Imaging and Communications in Medicine |
| DWI | Diffusion-weighted images |
| HIPAA | Health Insurance Portability and Accountability Act |
| BOLD | Blood oxygen level dependent |
| MDE | Multiple demand network |
| SEN | Sensorimotor network |
| DMN | Default mode network |
| CEN | Central Executive network |
| SAL | Salience network |
| LIM | Limbic network |
| LIMPA | Para limbic network |
| AUD | Auditory network |
| LAN | Language network |
| VAN | Ventral attention network |
| DAN | Dorsal attention network |
| SCN | Subcortical network |
| MT | Medial temporal network |
| VIS | Visual network |
| ACLAN | Accessory language network |
References
- Sporns, O. The Human Connectome: Origins and Challenges. NeuroImage 2013, 80, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Mori, S.; van Zijl, P.C.M. Fiber Tracking: Principles and Strategies—A Technical Review. NMR Biomed. 2002, 15, 468–480. [Google Scholar] [CrossRef] [PubMed]
- Jbabdi, S.; Johansen-Berg, H. Tractography: Where Do We Go from Here? Brain Connect. 2011, 1, 169–183. [Google Scholar] [CrossRef] [PubMed]
- Fox, M.D.; Raichle, M.E. Spontaneous Fluctuations in Brain Activity Observed with Functional Magnetic Resonance Imaging. Nat. Rev. Neurosci. 2007, 8, 700–711. [Google Scholar] [CrossRef]
- Cohen, A.L.; Fair, D.A.; Dosenbach, N.U.F.; Miezin, F.M.; Dierker, D.; Van Essen, D.C.; Schlaggar, B.L.; Petersen, S.E. Defining Functional Areas in Individual Human Brains Using Resting Functional Connectivity MRI. Neuroimage 2008, 41, 45–57. [Google Scholar] [CrossRef]
- Karbasforoushan, H.; Woodward, N.D. Resting-State Networks in Schizophrenia. Curr. Top. Med. Chem. 2012, 12, 2404–2414. [Google Scholar] [CrossRef]
- Burianová, H.; Faizo, N.L.; Gray, M.; Hocking, J.; Galloway, G.; Reutens, D. Altered Functional Connectivity in Mesial Temporal Lobe Epilepsy. Epilepsy Res. 2017, 137, 45–52. [Google Scholar] [CrossRef]
- Holiga, Š.; Hipp, J.F.; Chatham, C.H.; Garces, P.; Spooren, W.; D’Ardhuy, X.L.; Bertolino, A.; Bouquet, C.; Buitelaar, J.K.; Bours, C.; et al. Patients with Autism Spectrum Disorders Display Reproducible Functional Connectivity Alterations. Sci. Transl. Med. 2019, 11, eaat9223. [Google Scholar] [CrossRef]
- Rubinov, M.; Sporns, O. Complex Network Measures of Brain Connectivity: Uses and Interpretations. Neuroimage 2010, 52, 1059–1069. [Google Scholar] [CrossRef]
- Hagmann, P.; Cammoun, L.; Gigandet, X.; Meuli, R.; Honey, C.J.; Wedeen, V.J.; Sporns, O. Mapping the Structural Core of Human Cerebral Cortex. PLoS Biol. 2008, 6, e159. [Google Scholar] [CrossRef]
- Luppi, A.I.; Gellersen, H.M.; Liu, Z.-Q.; Peattie, A.R.D.; Manktelow, A.E.; Adapa, R.; Owen, A.M.; Naci, L.; Menon, D.K.; Dimitriadis, S.I.; et al. Systematic Evaluation of fMRI Data-Processing Pipelines for Consistent Functional Connectomics. Nat. Commun. 2024, 15, 4745. [Google Scholar] [CrossRef] [PubMed]
- Glasser, M.F.; Coalson, T.S.; Robinson, E.C.; Hacker, C.D.; Harwell, J.; Yacoub, E.; Ugurbil, K.; Andersson, J.; Beckmann, C.F.; Jenkinson, M.; et al. A Multi-Modal Parcellation of Human Cerebral Cortex. Nature 2016, 536, 171–178. [Google Scholar] [CrossRef]
- Eickhoff, S.B.; Yeo, B.T.T.; Genon, S. Imaging-Based Parcellations of the Human Brain. Nat. Rev. Neurosci. 2018, 19, 672–686. [Google Scholar] [CrossRef]
- Van Essen, D.C.; Ugurbil, K.; Auerbach, E.; Barch, D.; Behrens, T.E.J.; Bucholz, R.; Chang, A.; Chen, L.; Corbetta, M.; Curtiss, S.W.; et al. The Human Connectome Project: A Data Acquisition Perspective. Neuroimage 2012, 62, 2222–2231. [Google Scholar] [CrossRef] [PubMed]
- Van Essen, D.C.; Smith, S.M.; Barch, D.M.; Behrens, T.E.J.; Yacoub, E.; Ugurbil, K. The WU-Minn Human Connectome Project: An Overview. NeuroImage 2013, 80, 62–79. [Google Scholar] [CrossRef]
- Baker, C.M.; Burks, J.D.; Briggs, R.G.; Conner, A.K.; Glenn, C.A.; Sali, G.; McCoy, T.M.; Battiste, J.D.; O’Donoghue, D.L.; Sughrue, M.E. A Connectomic Atlas of the Human Cerebrum-Chapter 1: Introduction, Methods, and Significance. Oper. Neurosurg. (Hagerstown) 2018, 15, S1–S9. [Google Scholar] [CrossRef]
- Hart, M.G.; Price, S.J.; Suckling, J. Connectome Analysis for Pre-Operative Brain Mapping in Neurosurgery. Br. J. Neurosurg. 2016, 30, 506–517. [Google Scholar] [CrossRef]
- Magnani, M.; Rustici, A.; Zoli, M.; Tuleasca, C.; Chaurasia, B.; Franceschi, E.; Tonon, C.; Lodi, R.; Conti, A. Connectome-Based Neurosurgery in Primary Intra-Axial Neoplasms: Beyond the Traditional Modular Conception of Brain Architecture for the Preservation of Major Neurological Domains and Higher-Order Cognitive Functions. Life 2024, 14, 136. [Google Scholar] [CrossRef] [PubMed]
- Molina, E.S.; Tait, M.J.; Ieva, A.D. Connectomics as a Prognostic Tool of Functional Outcome in Glioma Surgery of the Supplementary Motor Area: Illustrative Case. J. Neurosurg. Case Lessons 2023, 6, CASE23286. [Google Scholar] [CrossRef]
- Bahrami, N.; Seibert, T.M.; Karunamuni, R.; Bartsch, H.; Krishnan, A.; Farid, N.; Hattangadi-Gluth, J.A.; McDonald, C.R. Altered Network Topology in Patients with Primary Brain Tumors After Fractionated Radiotherapy. Brain Connect. 2017, 7, 299–308. [Google Scholar] [CrossRef]
- De Roeck, L.; Blommaert, J.; Dupont, P.; Sunaert, S.; Sleurs, C.; Lambrecht, M. Brain Network Topology and Its Cognitive Impact in Adult Glioma Survivors. Sci. Rep. 2024, 14, 12782. [Google Scholar] [CrossRef]
- Sleurs, C.; Jacobs, S.; Counsell, S.J.; Christiaens, D.; Tournier, J.-D.; Sunaert, S.; Van Beek, K.; Uyttebroeck, A.; Deprez, S.; Batalle, D.; et al. Brain Network Hubs and Cognitive Performance of Survivors of Childhood Infratentorial Tumors. Radiother. Oncol. 2021, 161, 118–125. [Google Scholar] [CrossRef] [PubMed]
- Yarlagadda, S. In Proceedings of the European Society for Radiotherapy and Oncology (ESTRO) 2024 Annual Meeting, Glasgow, UK, 3–7 May 2024.
- Yarlagadda, S.; Belnap, S.C.; Kutuk, T.; Candela, J.; Reyes, T.C.; Menendez, M.A.; Hall, M.D.; Press, R.H.; Mehta, M.P.; McDermott, M.W.; et al. 630: Connectome analysis & cognitive outcomes post Whole-brain RT: Prospective registry’s first analysis. Radiother. Oncol. 2024, 194, S769–S771. [Google Scholar] [CrossRef]
- Kutuk, T.; Abrams, K.J.; Tom, M.C.; Rubens, M.; Appel, H.; Sidani, C.; Hall, M.D.; Tolakanahalli, R.; Wieczorek, D.J.J.; Gutierrez, A.N.; et al. Dedicated Isotropic 3-D T1 SPACE Sequence Imaging for Radiosurgery Planning Improves Brain Metastases Detection and Reduces the Risk of Intracranial Relapse. Radiother. Oncol. 2022, 173, 84–92. [Google Scholar] [CrossRef]
- Tournier, J.-D.; Calamante, F.; Gadian, D.G.; Connelly, A. Direct Estimation of the Fiber Orientation Density Function from Diffusion-Weighted MRI Data Using Spherical Deconvolution. Neuroimage 2004, 23, 1176–1185. [Google Scholar] [CrossRef]
- Doyen, S.; Nicholas, P.; Poologaindran, A.; Crawford, L.; Young, I.M.; Romero-Garcia, R.; Sughrue, M.E. Connectivity-Based Parcellation of Normal and Anatomically Distorted Human Cerebral Cortex. Hum. Brain Mapp. 2022, 43, 1358–1369. [Google Scholar] [CrossRef]
- Palmer, J.D.; Klamer, B.; Ballman, K.V.; Brown, P.D.; Cerhan, J.H.; Anderson, K.S.; Whitton, A.C.; Greenspoon, J.; Chung, C.; Burri, S.H.; et al. Effect of Stereotactic Radiosurgery Compared to Whole-Brain Radiotherapy for Limited Brain Metastasis on Long Term Cognition and Quality of Life: A Pooled Analysis of NCCTG N107C/CEC.3 and N0574 (Alliance) Randomized Clinical Trials. Int. J. Radiat. Oncol. Biol. Phys. 2020, 108, S175–S176. [Google Scholar] [CrossRef]
- Gondi, V.; Deshmukh, S.; Brown, P.D.; Wefel, J.S.; Armstrong, T.S.; Tome, W.A.; Gilbert, M.R.; Konski, A.; Robinson, C.G.; Bovi, J.A.; et al. Sustained Preservation of Cognition and Prevention of Patient-Reported Symptoms With Hippocampal Avoidance During Whole-Brain Radiation Therapy for Brain Metastases: Final Results of NRG Oncology CC001. Int. J. Radiat. Oncol. Biol. Phys. 2023, 117, 571–580. [Google Scholar] [CrossRef]
- Biswal, B.; Zerrin Yetkin, F.; Haughton, V.M.; Hyde, J.S. Functional Connectivity in the Motor Cortex of Resting Human Brain Using Echo-Planar Mri. Magn. Reson. Med. 1995, 34, 537–541. [Google Scholar] [CrossRef]
- Caspers, J.; Rubbert, C.; Eickhoff, S.B.; Hoffstaedter, F.; Südmeyer, M.; Hartmann, C.J.; Sigl, B.; Teichert, N.; Aissa, J.; Turowski, B.; et al. Within- and across-Network Alterations of the Sensorimotor Network in Parkinson’s Disease. Neuroradiology 2021, 63, 2073–2085. [Google Scholar] [CrossRef]
- Raichle, M.E.; Snyder, A.Z. A Default Mode of Brain Function: A Brief History of an Evolving Idea. NeuroImage 2007, 37, 1083–1090. [Google Scholar] [CrossRef] [PubMed]
- Vanhaudenhuyse, A.; Noirhomme, Q.; Tshibanda, L.J.-F.; Bruno, M.-A.; Boveroux, P.; Schnakers, C.; Soddu, A.; Perlbarg, V.; Ledoux, D.; Brichant, J.-F.; et al. Default Network Connectivity Reflects the Level of Consciousness in Non-Communicative Brain-Damaged Patients. Brain 2010, 133, 161–171. [Google Scholar] [CrossRef] [PubMed]
- Ghumman, S.; Fortin, D.; Noel-Lamy, M.; Cunnane, S.C.; Whittingstall, K. Exploratory Study of the Effect of Brain Tumors on the Default Mode Network. J. Neurooncol. 2016, 128, 437–444. [Google Scholar] [CrossRef] [PubMed]
- Menon, V. Large-Scale Brain Networks and Psychopathology: A Unifying Triple Network Model. Trends Cogn. Sci. 2011, 15, 483–506. [Google Scholar] [CrossRef]
- Chen, A.C.; Oathes, D.J.; Chang, C.; Bradley, T.; Zhou, Z.-W.; Williams, L.M.; Glover, G.H.; Deisseroth, K.; Etkin, A. Causal Interactions between Fronto-Parietal Central Executive and Default-Mode Networks in Humans. Proc. Natl. Acad. Sci. USA 2013, 110, 19944–19949. [Google Scholar] [CrossRef]
- Chand, G.B.; Dhamala, M. Interactions Among the Brain Default-Mode, Salience, and Central-Executive Networks During Perceptual Decision-Making of Moving Dots. Brain Connect. 2016, 6, 249–254. [Google Scholar] [CrossRef]
- Menon, V. Salience Network. Brain Mapp. 2015, 2, 597–611. [Google Scholar]
- Menon, V.; Uddin, L.Q. Saliency, Switching, Attention and Control: A Network Model of Insula Function. Brain Struct. Funct. 2010, 214, 655–667. [Google Scholar] [CrossRef]
- Enatsu, R.; Gonzalez-Martinez, J.; Bulacio, J.; Kubota, Y.; Mosher, J.; Burgess, R.C.; Najm, I.; Nair, D.R. Connections of the Limbic Network: A Corticocortical Evoked Potentials Study. Cortex 2015, 62, 20–33. [Google Scholar] [CrossRef]
- Thompson, M.; Umphred, D. The Limbic Network: Influence over Motor Control, Memory, and Learning; Mosby: Maryland Heights, MO, USA, 2019; ISBN 13: 978-0323721615. [Google Scholar]
- Kringelbach, M.L.; Rolls, E.T. The Functional Neuroanatomy of the Human Orbitofrontal Cortex: Evidence from Neuroimaging and Neuropsychology. Prog. Neurobiol. 2004, 72, 341–372. [Google Scholar] [CrossRef]
- Lou, H.C.; Gross, J.; Biermann-Ruben, K.; Kjaer, T.W.; Schnitzler, A. Coherence in Consciousness: Paralimbic Gamma Synchrony of Self-reference Links Conscious Experiences. Hum. Brain Mapp. 2010, 31, 185–192. [Google Scholar] [CrossRef] [PubMed]
- Camilleri, J.A.; Müller, V.I.; Fox, P.; Laird, A.R.; Hoffstaedter, F.; Kalenscher, T.; Eickhoff, S.B. Definition and Characterization of an Extended Multiple-Demand Network. Neuroimage 2018, 165, 138–147. [Google Scholar] [CrossRef]
- Kuiper, J.J.; Lin, Y.-H.; Young, I.M.; Bai, M.Y.; Briggs, R.G.; Tanglay, O.; Fonseka, R.D.; Hormovas, J.; Dhanaraj, V.; Conner, A.K.; et al. A Parcellation-Based Model of the Auditory Network. Hear. Res. 2020, 396, 108078. [Google Scholar] [CrossRef]
- Hackett, T.A. Information Flow in the Auditory Cortical Network. Hear. Res. 2011, 271, 133–146. [Google Scholar] [CrossRef]
- Ferstl, E.C.; Neumann, J.; Bogler, C.; von Cramon, D.Y. The Extended Language Network: A Meta-analysis of Neuroimaging Studies on Text Comprehension. Hum. Brain Mapp. 2007, 29, 581–593. [Google Scholar] [CrossRef]
- Asplund, C.L.; Todd, J.J.; Snyder, A.P.; Marois, R. A Central Role for the Lateral Prefrontal Cortex in Goal-Directed and Stimulus-Driven Attention. Nat. Neurosci. 2010, 13, 507–512. [Google Scholar] [CrossRef] [PubMed]
- Corbetta, M.; Shulman, G.L. Spatial Neglect and Attention Networks. Annu. Rev. Neurosci. 2011, 34, 569–599. [Google Scholar] [CrossRef] [PubMed]
- Behrens, T.E.J.; Johansen-Berg, H.; Woolrich, M.W.; Smith, S.M.; Wheeler-Kingshott, C.A.M.; Boulby, P.A.; Barker, G.J.; Sillery, E.L.; Sheehan, K.; Ciccarelli, O.; et al. Non-Invasive Mapping of Connections between Human Thalamus and Cortex Using Diffusion Imaging. Nat. Neurosci. 2003, 6, 750–757. [Google Scholar] [CrossRef]
- Ji, J.L.; Spronk, M.; Kulkarni, K.; Repovš, G.; Anticevic, A.; Cole, M.W. Mapping the Human Brain’s Cortical-Subcortical Functional Network Organization. Neuroimage 2019, 185, 35–57. [Google Scholar] [CrossRef]
- Bell, P.T.; Shine, J.M. Subcortical Contributions to Large-Scale Network Communication. Neurosci. Biobehav. Rev. 2016, 71, 313–322. [Google Scholar] [CrossRef]
- Seoane, S.; Modroño, C.; González-Mora, J.L.; Janssen, N. Medial Temporal Lobe Contributions to Resting-State Networks. Brain Struct. Funct. 2022, 227, 995–1012. [Google Scholar] [CrossRef] [PubMed]
- Baker, C.M.; Burks, J.D.; Briggs, R.G.; Stafford, J.; Conner, A.K.; Glenn, C.A.; Sali, G.; McCoy, T.M.; Battiste, J.D.; O’Donoghue, D.L.; et al. A Connectomic Atlas of the Human Cerebrum—Chapter 9: The Occipital Lobe. Oper. Neurosurg. 2018, 15, S372. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Wang, J.; Fan, L.; Li, H.; Zhang, W.; Hu, Q.; Jiang, T. Tractography-Based Parcellation of the Human Middle Temporal Gyrus. Sci. Rep. 2015, 5, 18883. [Google Scholar] [CrossRef] [PubMed]
- Kutuk, T.; Balda, A.; Odia, Y.; Hall, M.D.; Appel, H.; Ramos, S.; Menendez, M.R.; Mohler, A.; McDermott, M.; Ahluwalia, M.S.; et al. Implementation of an Interactive, Multi-Language, App-Based Neurocognitive Evaluation Program into Routine Stereotactic Radiosurgery Practice. Cureus J. Med. Sci. 2023, 15. Available online: https://www.cureus.com/abstracts/873-implementation-of-an-interactive-multi-language-app-based-neurocognitive-evaluation-program-into-routine-stereotactic-radiosurgery-practice (accessed on 20 May 2025).
- Leemans, K.; De Ridder, M. Cognition: Development of a Cognitive Testing Battery on the iPad for the Evaluation of Patients with Brain Mets. Acta Neurol. Belg. 2022, 122, 145–152. [Google Scholar] [CrossRef]
- Akdemir, E.Y.; Gurdikyan, S.; Reyes, T.C.; Odia, Y.; Menendez, M.A.R.; Yarlagadda, S.; Gal, O.; Hall, M.D.; Press, R.H.; Wieczorek, D.J.; et al. Integrating a Novel Tablet-Based Digital Neurocognitive Assessment Tool in Brain Metastases Patients. J. Neurooncol. 2025. [Google Scholar] [CrossRef]
- Duff, K. Evidence-Based Indicators of Neuropsychological Change in the Individual Patient: Relevant Concepts and Methods. Arch. Clin. Neuropsychol. 2012, 27, 248–261. [Google Scholar] [CrossRef]
- Sridhar, S.; Khamaj, A.; Asthana, M.K. Cognitive Neuroscience Perspective on Memory: Overview and Summary. Front. Hum. Neurosci. 2023, 17, 1217093. [Google Scholar] [CrossRef]
- Haubrich, J.; Vera, L.D.; Manahan-Vaughan, D. Cortico-Subcortical Networks That Determine Behavioral Memory Renewal Are Redefined by Noradrenergic Neuromodulation. Sci. Rep. 2025, 15, 9692. [Google Scholar] [CrossRef]
- Li, Z.; Athwal, D.; Lee, H.-L.; Sah, P.; Opazo, P.; Chuang, K.-H. Locating Causal Hubs of Memory Consolidation in Spontaneous Brain Network in Male Mice. Nat. Commun. 2023, 14, 5399. [Google Scholar] [CrossRef]
- Yoshida, J.; Oñate, M.; Khatami, L.; Vera, J.; Nadim, F.; Khodakhah, K. Cerebellar Contributions to the Basal Ganglia Influence Motor Coordination, Reward Processing, and Movement Vigor. J. Neurosci. 2022, 42, 8406–8415. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Wang, T.; Li, W.; Li, H.; Xu, B.; Zhang, M.; Yue, L.; Wang, P.; Xiao, S. Morphological, Structural, and Functional Networks Highlight the Role of the Cortical-Subcortical Circuit in Individuals With Subjective Cognitive Decline. Front. Aging Neurosci. 2021, 13, 688113. [Google Scholar] [CrossRef] [PubMed]
- Pilly, P.K.; Grossberg, S. How Do Spatial Learning and Memory Occur in the Brain? Coordinated Learning of Entorhinal Grid Cells and Hippocampal Place Cells. J. Cogn. Neurosci. 2012, 24, 1031–1054. [Google Scholar] [CrossRef] [PubMed]
- Ushakov, V.; Sharaev, M.G.; Kartashov, S.I.; Zavyalova, V.V.; Verkhlyutov, V.M.; Velichkovsky, B.M. Dynamic Causal Modeling of Hippocampal Links within the Human Default Mode Network: Lateralization and Computational Stability of Effective Connections. Front. Hum. Neurosci. 2016, 10, 528. [Google Scholar] [CrossRef]
- Sporns, O. Network Attributes for Segregation and Integration in the Human Brain. Curr. Opin. Neurobiol. 2013, 23, 162–171. [Google Scholar] [CrossRef]
- Nenning, K.-H.; Furtner, J.; Kiesel, B.; Schwartz, E.; Roetzer, T.; Fortelny, N.; Bock, C.; Grisold, A.; Marko, M.; Leutmezer, F.; et al. Distributed Changes of the Functional Connectome in Patients with Glioblastoma. Sci. Rep. 2020, 10, 18312. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S. No. | Brain Network | No. of Parcellations | Network Description | Clinical Significance | Ref. |
|---|---|---|---|---|---|
| 1 | Sensorimotor network (SEN) | 30 | Encompasses the primary motor, primary sensory, supplementary motor area, dorsal premotor area, ventral premotor area, and cingulate premotor areas. | Complex motor planning and regulation of muscles in the upper limb and upper trunk muscles. Control of hand movements and in performing specific motor tasks based on visual cues. | [30,31] |
| 2 | Default mode network (DMN) | 27 | Encompasses the medial prefrontal cortex, posterior cingulate cortex, precuneus, and lateral/medial temporal lobes. | Dominant network of the task-negative system. Involved in internal mental states, theory of mind, and numerous other cognitive processes. Frequently abnormal in numerous mental illnesses, including Alzheimer’s. Tumors in the left hemisphere and cerebellum are observed to have a significant effect on DMN. | [32,33,34] |
| 3 | Central Executive network (CEN) | 24 | Encompasses dorsolateral prefrontal cortex, posterior parietal cortex. | Dominant network of the task-positive system (active system). CEN is crucial for actively maintaining and manipulating information in working memory, for rule-based problem-solving, and for decision-making in the context of goal-directed behavior. | [35,36,37] |
| 4 | Salience network (SAL) | 49 | The main components are insula and cingulate cortex with 3 key subcortical structures: amygdala, ventral striatum, and substantia nigra/ventral tegmental area. | Plays a role in transitioning between the DMN and CEN, as well as possibly playing a role in pain sensation. Involved in anticipating outcomes, recognizing reward values, and encoding errors to influence attention allocation and motor preparation. | [38,39] |
| 5 | Limbic network (LIM) | 24 | Includes hippocampus, temporal pole, amygdala, posterior cingulate gyrus, and medial, lateral orbitofrontal cortex. | Responsible for conditioned emotional learning, emotional expression, assessment of motivational content, social and emotional processing, and encoding long-term memories. | [40,41] |
| 6 | Para limbic network (LIMPA) | 34 | Includes orbitofrontal cortex, intermodal frontal lobe, and fusiform gyrus. | Exhibits key integrations with the limbic network and plays a crucial role in regulating brain resource allocation; integrates attention, awareness, and emotion. | [42,43] |
| 7 | Multiple demand extended network (MDE) | 56 | Encompasses lateral prefrontal cortex in the inferior frontal sulcus, frontal operculum, intra-parietal sulcus and bilateral pre-supplementary motor area and adjacent middle cingulate cortex (pre-SMA/MCC), anterior insula (aINS), middle frontal gyrus/posterior inferior frontal sulcus/(MFG/IFS). | Plays a role in higher cognitive functions like novelty, perceptual difficulty, conflict response, and different types of memory. | [44] |
| 8 | Auditory network (AUD) | 30 | Encompasses superior temporal gyrus, inferior frontal gyrus, medial frontal gyrus, adjacent insula. | Responsible for discriminating auditory information. Involved in interpreting incoming speech and determining volume, frequency, and onset time of auditory information. | [45,46] |
| 9 | Language network (LAN) | 15 | Includes posterior temporal areas (Wernicke’s area), parietal areas (Geschwind’s area), inferior frontal areas (Broca’s area 44 and 45), supplementary motor area (SMA), area 55b, and area 8C. | Responsible for text comprehension and articulating thoughts into verbal and written words. | [47] |
| 10 | Ventral attention network (VAN) | 11 | Encompasses the lateral and inferior frontal/prefrontal cortex and the temporo-parietal junction. | Involved in attention switching, visual–spatial perception, episodic memory retrieval, and consciousness. | [48,49] |
| 11 | Dorsal attention network (DAN) | 37 | Encompasses the dorsolateral prefrontal cortex, left/right posterior intraparietal sulci, and the frontal eye fields. | Mainly involved in goal-directed attention. Also plays a role in working memory, motor, and executive functions. | [48,49] |
| 12 | Subcortical network (SCN) | 14 | Includes basal ganglia, diencephalon, cerebellum, brain stem, and thalamus. | Plays a regulatory role in cognitive processing, sensory gating, and learning. Implicated in the development of Parkinson’s disease, psychosis, and bipolar disorder. | [50,51,52] |
| 13 | Medial temporal network (MT) | 16 | Encompasses perirhinal ectorhinal cortex (part of temporal lobe), para hippocampal regions PHA1, 2, and 3. | Critically involved in visuospatial processing and episodic memory by processing contextual information. MT-related abnormalities have been extensively demonstrated in patients with temporal lobe epilepsy. | [53] |
| 14 | Visual network (VIS) | 52 | Includes primary visual areas (V1-V4), dorsal visual stream, ventral visual stream, and the lateral visual stream. | Primarily involved in visual processing, and plays role in hand–eye coordination. | [54] |
| 15 | Accessory language network (ACLAN) | 4 | Pathway in the middle temporal gyrus, superior temporal sulcus. | Involved in verbal memory, speech recognition, and the representation of lexical concepts. | [55] |
| Neurocognitive Domain | Test | Description |
|---|---|---|
| Learning and memory | Verbal recall | Total number of words correctly recalled over 3 trials |
| Verbal revision | Total number of words correctly recalled | |
| Verbal recognition | Total number of list words correctly identified minus total number of non-list words incorrectly identified | |
| Attention and speed of processing | Symbol match | Total number of correct answers |
| Executive functions | Ordering numbers and letters | Total time in whole seconds to complete the array |
| Verbal fluency | Words that start with | Total number of valid words |
| Fine motor speed | Ordering numbers | Total time in whole seconds to complete the array |
| S. No. | Brain Network | Baseline Anomaly Frequency | Post-WBRT Anomaly Frequency | Proportional Change (%) |
|---|---|---|---|---|
| 1 | Sensorimotor network (SEN) | 57 | 40 | −29.82% |
| 2 | Default mode network (DMN) | 18 | 8 | −55.56% |
| 3 | Central Executive network (CEN) | 74 | 82 | 10.81% |
| 4 | Salience network (SAL) | 105 | 101 | −3.81% |
| 5 | Limbic network (LIM) | 30 | 52 | 73.33% |
| 6 | Para limbic network (LIMPA) | 107 | 161 | 50.47% |
| 7 | Multiple demand extended network (MDE) | 258 | 242 | −6.20% |
| 8 | Auditory network (AUD) | 18 | 13 | −27.78% |
| 9 | Language network (LAN) | 21 | 23 | 9.52% |
| 10 | Ventral attention network (VAN) | 6 | 6 | 0.00 |
| 11 | Dorsal attention network (DAN) | 78 | 81 | 3.85% |
| 12 | Subcortical network (SCN) | 12 | 15 | 25.00% |
| 13 | Medial temporal network (MT) | 7 | 9 | 28.57% |
| 14 | Visual network (VIS) | 126 | 79 | −37.30% |
| 15 | Accessory language network (ACLAN) | 0 | 2 | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yarlagadda, S.; Belnap, S.; Candela, J.; Kutuk, T.; Reyes, T.C.; Ramirez Menendez, M.; Hall, M.; Press, R.; Odia, Y.; Mehta, M.; et al. Correlation Between Neurocognitive Outcomes and Neuroaxonal Connectome Alterations After Whole Brain Radiotherapy: A Proof-of-Concept Study. Cancers 2025, 17, 1752. https://doi.org/10.3390/cancers17111752
Yarlagadda S, Belnap S, Candela J, Kutuk T, Reyes TC, Ramirez Menendez M, Hall M, Press R, Odia Y, Mehta M, et al. Correlation Between Neurocognitive Outcomes and Neuroaxonal Connectome Alterations After Whole Brain Radiotherapy: A Proof-of-Concept Study. Cancers. 2025; 17(11):1752. https://doi.org/10.3390/cancers17111752
Chicago/Turabian StyleYarlagadda, Sreenija, Starlie Belnap, John Candela, Tugce Kutuk, Thailin Companioni Reyes, Miguel Ramirez Menendez, Matthew Hall, Robert Press, Yazmin Odia, Minesh Mehta, and et al. 2025. "Correlation Between Neurocognitive Outcomes and Neuroaxonal Connectome Alterations After Whole Brain Radiotherapy: A Proof-of-Concept Study" Cancers 17, no. 11: 1752. https://doi.org/10.3390/cancers17111752
APA StyleYarlagadda, S., Belnap, S., Candela, J., Kutuk, T., Reyes, T. C., Ramirez Menendez, M., Hall, M., Press, R., Odia, Y., Mehta, M., McDermott, M., & Kotecha, R. (2025). Correlation Between Neurocognitive Outcomes and Neuroaxonal Connectome Alterations After Whole Brain Radiotherapy: A Proof-of-Concept Study. Cancers, 17(11), 1752. https://doi.org/10.3390/cancers17111752