


On the Therapeutic Potential of ERK4 in Triple-Negative Breast Cancer

 , and
, and 
{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
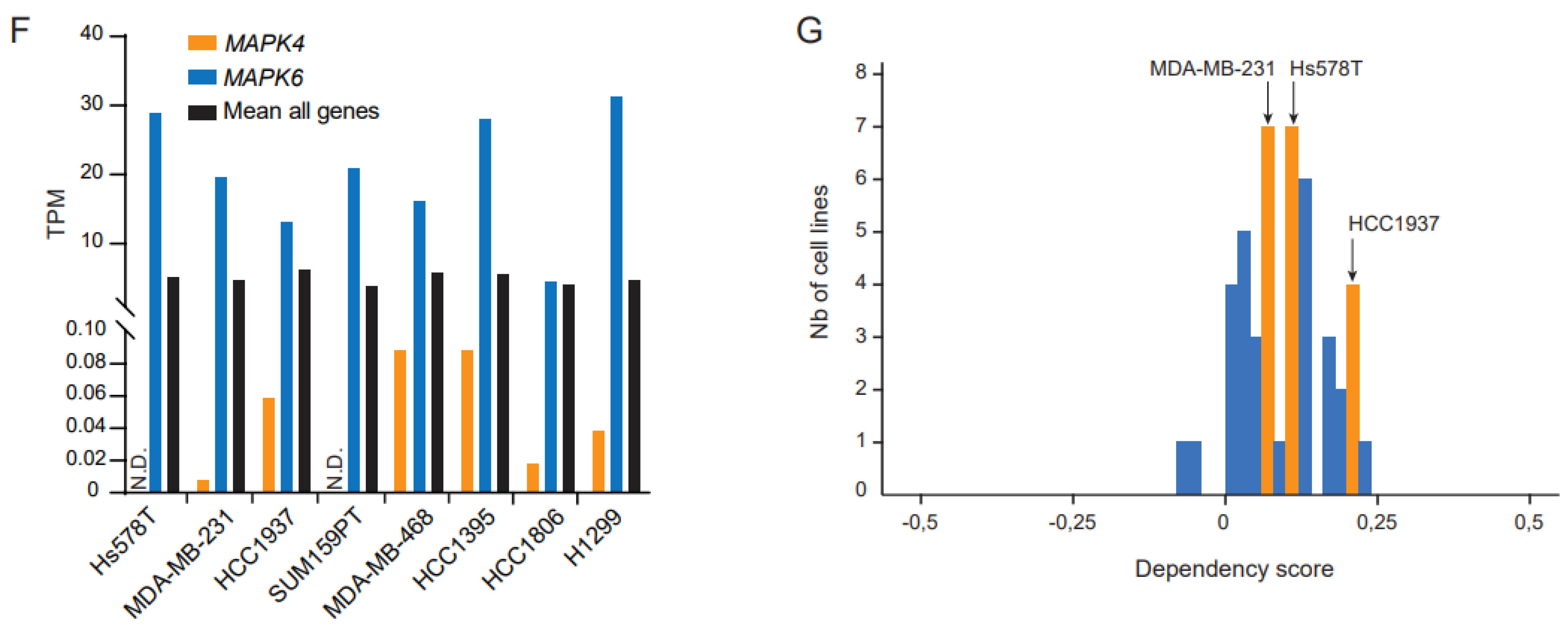
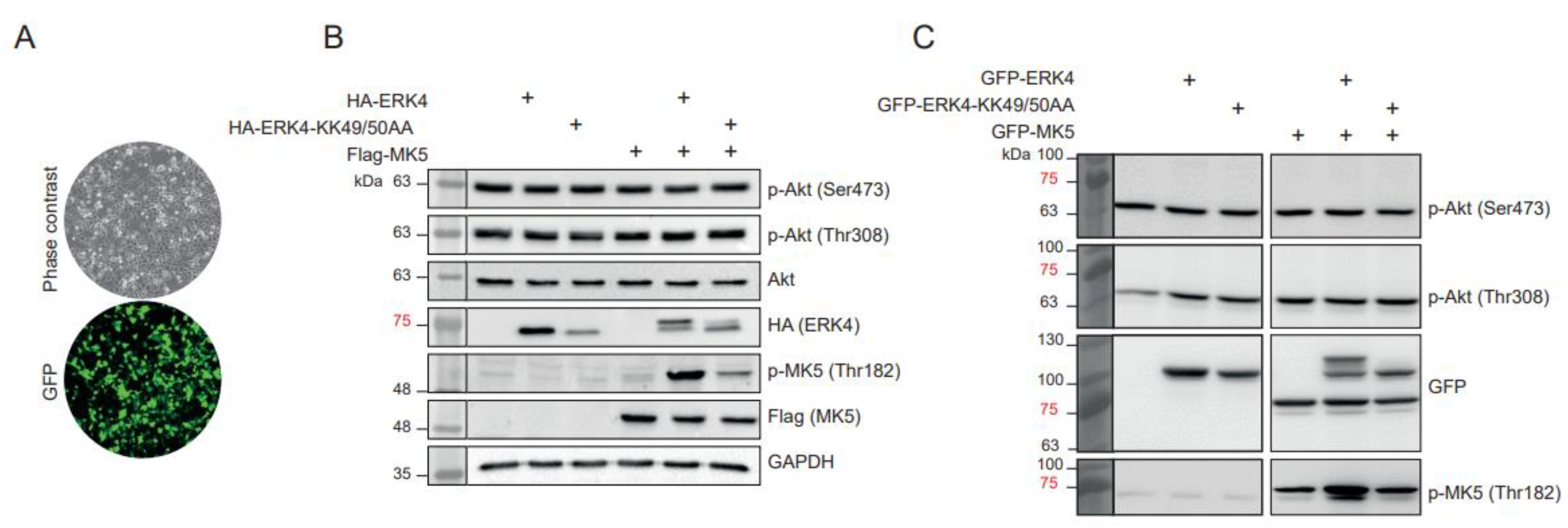
2. Results and Discussion
3. Materials and Methods
3.1. Cell Lines and Cell Culture
3.2. Transfections and RNA Interference
3.3. Western Blot Analysis
3.4. Real-Time Quantitative PCR Analysis
3.5. Gene Expression Profiling
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mathien, S.; Soulez, M.; Klinger, S.; Meloche, S. Erk3 and Erk4. In Encyclopedia of Signaling Molecules, 2nd ed.; Choi, S., Ed.; Springer International Publishing: Cham, Switzerland, 2018; pp. 1632–1638. [Google Scholar]
- Coulombe, P.; Meloche, S. Atypical mitogen-activated protein kinases: Structure, regulation and functions. Biochim. Biophys. Acta 2007, 1773, 1376–1387. [Google Scholar] [CrossRef] [PubMed]
- Boulton, T.G.; Nye, S.H.; Robbins, D.J.; Ip, N.Y.; Radziejewska, E.; Morgenbesser, S.D.; DePinho, R.A.; Panayotatos, N.; Cobb, M.H.; Yancopoulos, G.D. ERKs: A family of protein-serine/threonine kinases that are activated and tyrosine phosphorylated in response to insulin and NGF. Cell 1991, 65, 663–675. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, F.A.; Raden, D.L.; Rigby, M.R.; Davis, R.J. Heterogeneous expression of four MAP kinase isoforms in human tissues. FEBS Lett. 1992, 304, 170–178. [Google Scholar] [CrossRef] [PubMed]
- Rousseau, J.; Klinger, S.; Rachalski, A.; Turgeon, B.; Deleris, P.; Vigneault, E.; Poirier-Heon, J.F.; Davoli, M.A.; Mechawar, N.; El Mestikawy, S.; et al. Targeted inactivation of Mapk4 in mice reveals specific nonredundant functions of Erk3/Erk4 subfamily mitogen-activated protein kinases. Mol. Cell Biol. 2010, 30, 5752–5763. [Google Scholar] [CrossRef]
- Uhlen, M.; Fagerberg, L.; Hallstrom, B.M.; Lindskog, C.; Oksvold, P.; Mardinoglu, A.; Sivertsson, A.; Kampf, C.; Sjostedt, E.; Asplund, A.; et al. Proteomics. Tissue-based map of the human proteome. Science 2015, 347, 1260419. [Google Scholar] [CrossRef]
- Courcelles, M.; Fremin, C.; Voisin, L.; Lemieux, S.; Meloche, S.; Thibault, P. Phosphoproteome dynamics reveal novel ERK1/2 MAP kinase substrates with broad spectrum of functions. Mol. Syst. Biol. 2013, 9, 669. [Google Scholar] [CrossRef]
- Unal, E.B.; Uhlitz, F.; Bluthgen, N. A compendium of ERK targets. FEBS Lett. 2017, 591, 2607–2615. [Google Scholar] [CrossRef]
- Aberg, E.; Perander, M.; Johansen, B.; Julien, C.; Meloche, S.; Keyse, S.M.; Seternes, O.M. Regulation of MAPK-activated protein kinase 5 activity and subcellular localization by the atypical MAPK ERK4/MAPK4. J. Biol. Chem. 2006, 281, 35499–35510. [Google Scholar] [CrossRef]
- Kant, S.; Schumacher, S.; Singh, M.K.; Kispert, A.; Kotlyarov, A.; Gaestel, M. Characterization of the atypical MAPK ERK4 and its activation of the MAPK-activated protein kinase MK5. J. Biol. Chem. 2006, 281, 35511–35519. [Google Scholar] [CrossRef]
- Schumacher, S.; Laass, K.; Kant, S.; Shi, Y.; Visel, A.; Gruber, A.D.; Kotlyarov, A.; Gaestel, M. Scaffolding by ERK3 regulates MK5 in development. EMBO J. 2004, 23, 4770–4779. [Google Scholar] [CrossRef]
- Seternes, O.M.; Mikalsen, T.; Johansen, B.; Michaelsen, E.; Armstrong, C.G.; Morrice, N.A.; Turgeon, B.; Meloche, S.; Moens, U.; Keyse, S.M. Activation of MK5/PRAK by the atypical MAP kinase ERK3 defines a novel signal transduction pathway. EMBO J. 2004, 23, 4780–4791. [Google Scholar] [CrossRef] [PubMed]
- Ronkina, N.; Gaestel, M. MAPK-Activated Protein Kinases: Servant or Partner? Annu. Rev. Biochem. 2022, 91, 505–540. [Google Scholar] [CrossRef] [PubMed]
- Deleris, P.; Rousseau, J.; Coulombe, P.; Rodier, G.; Tanguay, P.L.; Meloche, S. Activation loop phosphorylation of the atypical MAP kinases ERK3 and ERK4 is required for binding, activation and cytoplasmic relocalization of MK5. J. Cell Physiol. 2008, 217, 778–788. [Google Scholar] [CrossRef]
- Ronkina, N.; Schuster-Gossler, K.; Hansmann, F.; Kunze-Schumacher, H.; Sandrock, I.; Yakovleva, T.; Lafera, J.; Baumgartner, W.; Krueger, A.; Prinz, I.; et al. Germ Line Deletion Reveals a Nonessential Role of Atypical Mitogen-Activated Protein Kinase 6/Extracellular Signal-Regulated Kinase 3. Mol. Cell. Biol. 2019, 39, e00516-18. [Google Scholar] [CrossRef] [PubMed]
- Soulez, M.; Saba-El-Leil, M.K.; Turgeon, B.; Mathien, S.; Coulombe, P.; Klinger, S.; Rousseau, J.; Levesque, K.; Meloche, S. Reevaluation of the Role of Extracellular Signal-Regulated Kinase 3 in Perinatal Survival and Postnatal Growth Using New Genetically Engineered Mouse Models. Mol. Cell. Biol. 2019, 39, e00527-18. [Google Scholar] [CrossRef] [PubMed]
- Shen, T.; Wang, W.; Zhou, W.; Coleman, I.; Cai, Q.; Dong, B.; Ittmann, M.M.; Creighton, C.J.; Bian, Y.; Meng, Y.; et al. MAPK4 promotes prostate cancer by concerted activation of androgen receptor and AKT. J. Clin. Investig. 2021, 131. [Google Scholar] [CrossRef]
- Wang, W.; Shen, T.; Dong, B.; Creighton, C.J.; Meng, Y.; Zhou, W.; Shi, Q.; Zhou, H.; Zhang, Y.; Moore, D.D.; et al. MAPK4 overexpression promotes tumor progression via noncanonical activation of AKT/mTOR signaling. J. Clin. Investig. 2019, 129, 1015–1029. [Google Scholar] [CrossRef]
- Tian, S.; Lou, L.; Tian, M.; Lu, G.; Tian, J.; Chen, X. MAPK4 deletion enhances radiation effects and triggers synergistic lethality with simultaneous PARP1 inhibition in cervical cancer. J. Exp. Clin. Cancer Res. CR 2020, 39, 143. [Google Scholar] [CrossRef]
- Zeng, X.; Jiang, S.; Ruan, S.; Guo, Z.; Guo, J.; Liu, M.; Ye, C.; Dong, J. MAPK4 silencing together with a PARP1 inhibitor as a combination therapy in triplenegative breast cancer cells. Mol. Med. Rep. 2021, 24, 548. [Google Scholar] [CrossRef]
- Wang, W.; Han, D.; Cai, Q.; Shen, T.; Dong, B.; Lewis, M.T.; Wang, R.; Meng, Y.; Zhou, W.; Yi, P.; et al. MAPK4 promotes triple negative breast cancer growth and reduces tumor sensitivity to PI3K blockade. Nat. Commun. 2022, 13, 245. [Google Scholar] [CrossRef]
- Barretina, J.; Caponigro, G.; Stransky, N.; Venkatesan, K.; Margolin, A.A.; Kim, S.; Wilson, C.J.; Lehar, J.; Kryukov, G.V.; Sonkin, D.; et al. The Cancer Cell Line Encyclopedia enables predictive modelling of anticancer drug sensitivity. Nature 2012, 483, 603–607. [Google Scholar] [CrossRef] [PubMed]
- Tsherniak, A.; Vazquez, F.; Montgomery, P.G.; Weir, B.A.; Kryukov, G.; Cowley, G.S.; Gill, S.; Harrington, W.F.; Pantel, S.; Krill-Burger, J.M.; et al. Defining a Cancer Dependency Map. Cell 2017, 170, 564–576.e16. [Google Scholar] [CrossRef] [PubMed]
- Pearson, G.; Robinson, F.; Beers Gibson, T.; Xu, B.E.; Karandikar, M.; Berman, K.; Cobb, M.H. Mitogen-activated protein (MAP) kinase pathways: Regulation and physiological functions. Endocr. Rev. 2001, 22, 153–183. [Google Scholar]
- Gonzalez, F.A.; Raden, D.L.; Davis, R.J. Identification of substrate recognition determinants for human ERK1 and ERK2 protein kinases. J. Biol. Chem. 1991, 266, 22159–22163. [Google Scholar] [CrossRef] [PubMed]
- Songyang, Z.; Lu, K.P.; Kwon, Y.T.; Tsai, L.H.; Filhol, O.; Cochet, C.; Brickey, D.A.; Soderling, T.R.; Bartleson, C.; Graves, D.J.; et al. A structural basis for substrate specificities of protein Ser/Thr kinases: Primary sequence preference of casein kinases I and II, NIMA, phosphorylase kinase, calmodulin-dependent kinase II, CDK5, and Erk1. Mol. Cell. Biol. 1996, 16, 6486–6493. [Google Scholar] [CrossRef] [PubMed]
- Ronkina, N.; Johansen, C.; Bohlmann, L.; Lafera, J.; Menon, M.B.; Tiedje, C.; Laass, K.; Turk, B.E.; Iversen, L.; Kotlyarov, A.; et al. Comparative Analysis of Two Gene-Targeting Approaches Challenges the Tumor-Suppressive Role of the Protein Kinase MK5/PRAK. PLoS ONE 2015, 10, e0136138. [Google Scholar] [CrossRef] [PubMed]
- Coulombe, P.; Rodier, G.; Pelletier, S.; Pellerin, J.; Meloche, S. Rapid turnover of extracellular signal-regulated kinase 3 by the ubiquitin-proteasome pathway defines a novel paradigm of mitogen-activated protein kinase regulation during cellular differentiation. Mol. Cell. Biol. 2003, 23, 4542–4558. [Google Scholar] [CrossRef]
- Perander, M.; Aberg, E.; Johansen, B.; Dreyer, B.; Guldvik, I.J.; Outzen, H.; Keyse, S.M.; Seternes, O.M. The Ser(186) phospho-acceptor site within ERK4 is essential for its ability to interact with and activate PRAK/MK5. Biochem. J. 2008, 411, 613–622. [Google Scholar] [CrossRef]
- Girondel, C.; Levesque, K.; Langlois, M.J.; Pasquin, S.; Saba-El-Leil, M.K.; Rivard, N.; Friesel, R.; Servant, M.J.; Gauchat, J.F.; Lesage, S.; et al. Loss of interleukin-17 receptor D promotes chronic inflammation-associated tumorigenesis. Oncogene 2021, 40, 452–464. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boudghene-Stambouli, F.; Soulez, M.; Ronkina, N.; Dörrie, A.; Kotlyarov, A.; Seternes, O.-M.; Gaestel, M.; Meloche, S. On the Therapeutic Potential of ERK4 in Triple-Negative Breast Cancer. Cancers 2023, 15, 25. https://doi.org/10.3390/cancers15010025
Boudghene-Stambouli F, Soulez M, Ronkina N, Dörrie A, Kotlyarov A, Seternes O-M, Gaestel M, Meloche S. On the Therapeutic Potential of ERK4 in Triple-Negative Breast Cancer. Cancers. 2023; 15(1):25. https://doi.org/10.3390/cancers15010025
Chicago/Turabian StyleBoudghene-Stambouli, Fadia, Mathilde Soulez, Natalia Ronkina, Anneke Dörrie, Alexey Kotlyarov, Ole-Morten Seternes, Matthias Gaestel, and Sylvain Meloche. 2023. "On the Therapeutic Potential of ERK4 in Triple-Negative Breast Cancer" Cancers 15, no. 1: 25. https://doi.org/10.3390/cancers15010025
APA StyleBoudghene-Stambouli, F., Soulez, M., Ronkina, N., Dörrie, A., Kotlyarov, A., Seternes, O.-M., Gaestel, M., & Meloche, S. (2023). On the Therapeutic Potential of ERK4 in Triple-Negative Breast Cancer. Cancers, 15(1), 25. https://doi.org/10.3390/cancers15010025