
Tumour Colonisation of Parvimonas micra Is Associated with Decreased Survival in Colorectal Cancer Patients
 , and
, and 
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Cohort
2.2. Detection of Microbial Markers in Fresh Frozen Tumour Tissue and Faeces Using Quantitative Real-Time PCR (qPCR)
2.3. Molecular Analyses
2.3.1. Microsatellite Instability (MSI) Assessment
2.3.2. Analyses of BRAF and KRAS Mutation
2.4. Statistical Methods
3. Results
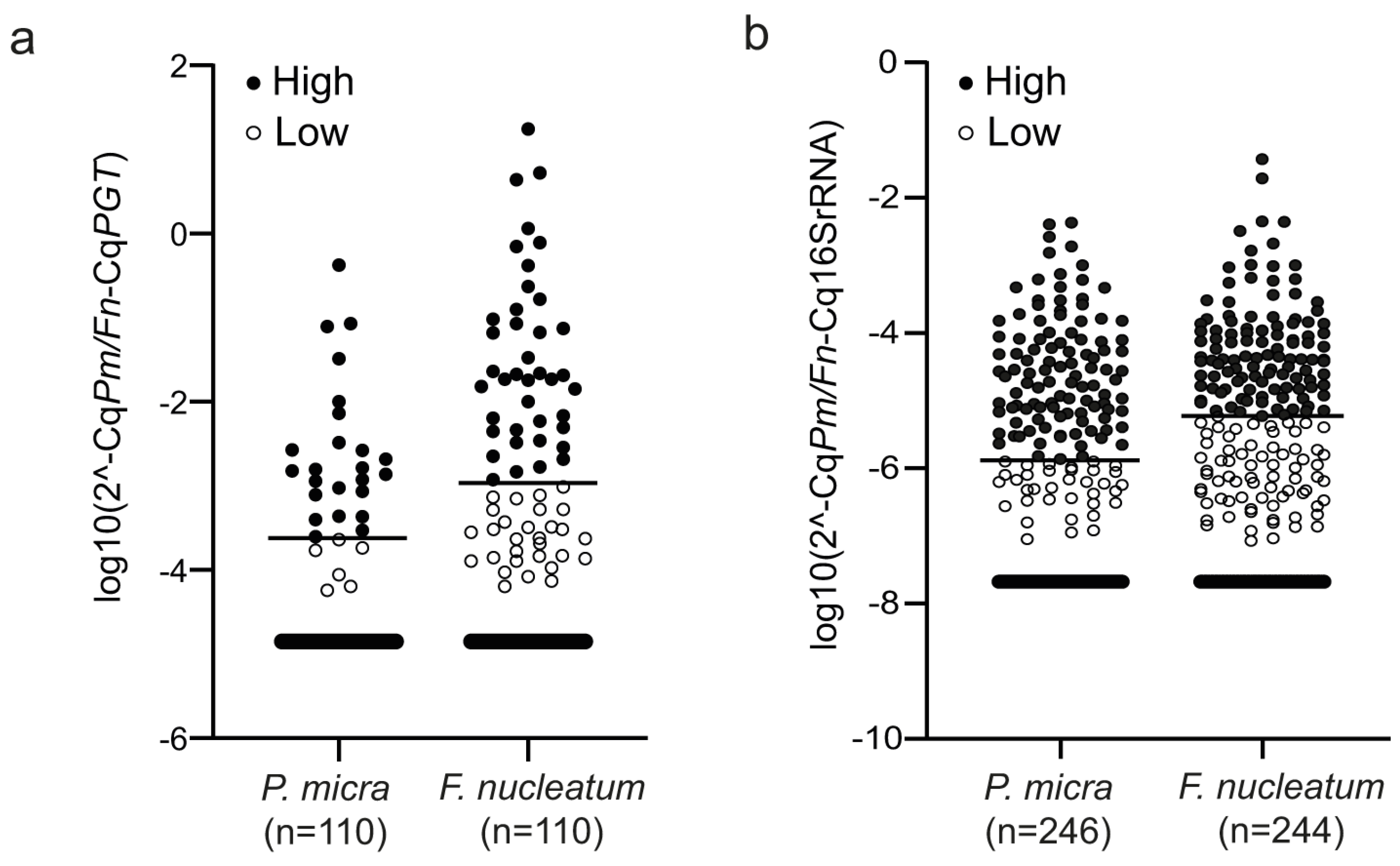
3.1. Distribution of P. micra and F. nucleatum in Tumour Tissue and Faeces of CRC Patients
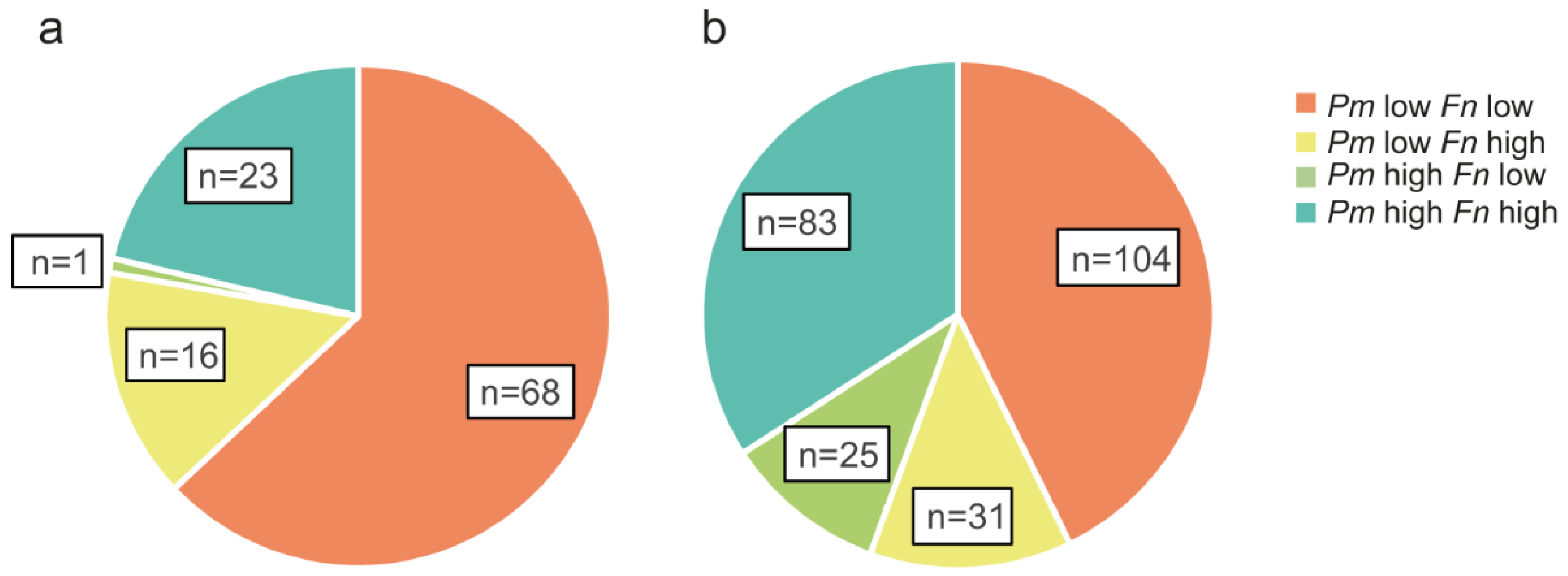
3.2. Association of P. micra and F. nucleatum in Tumour Tissue and Faeces with Clinicopathological and Molecular Parameters
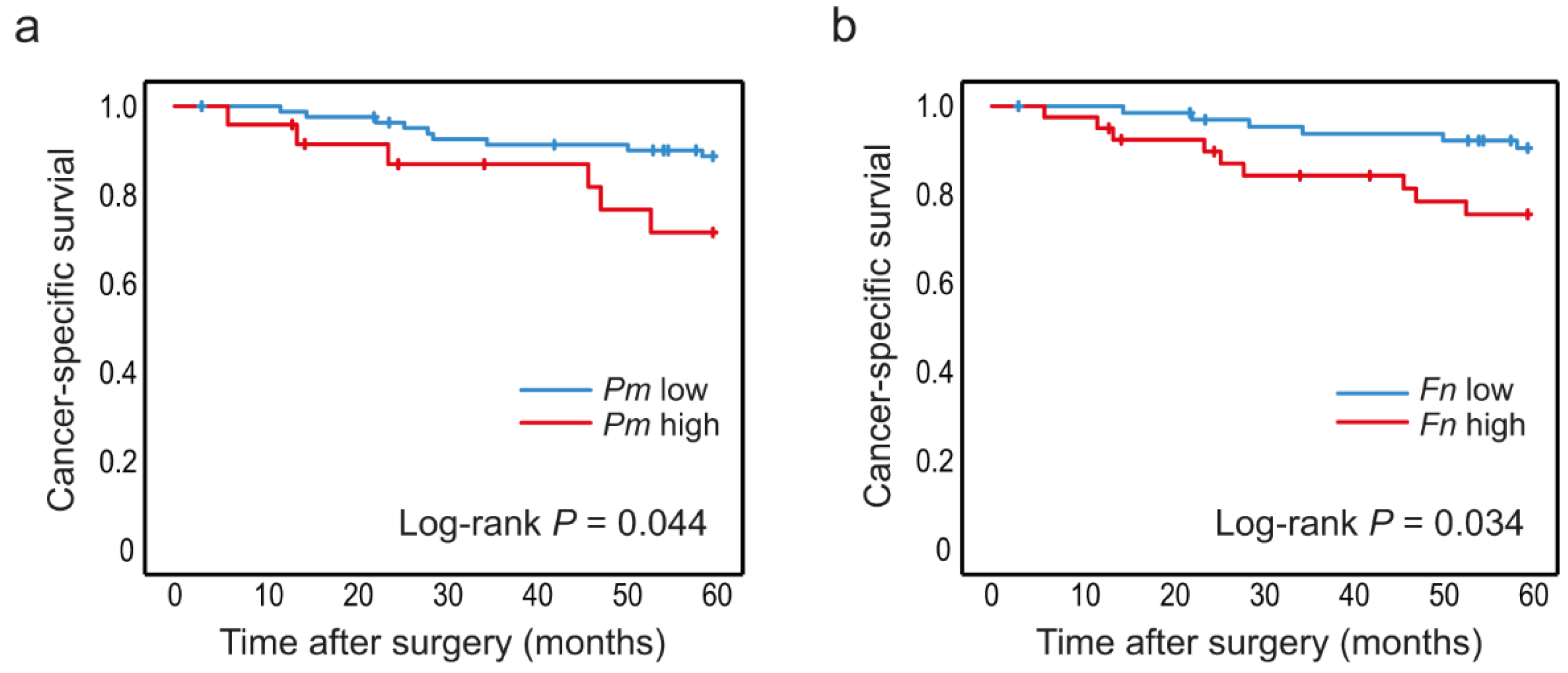
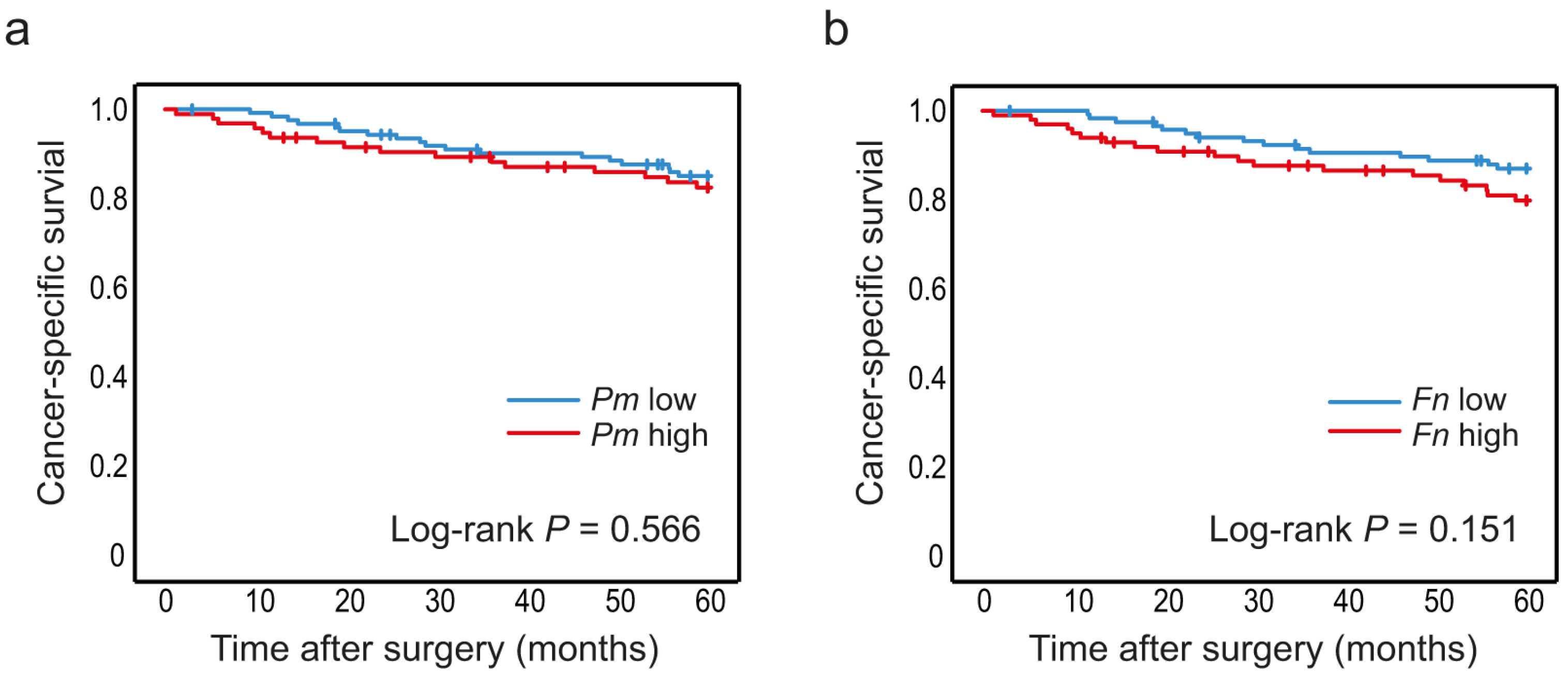
3.3. High Levels of P. micra and F. nucleatum in Tumour Tissue Are Associated with Decreased Patient Survival
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Morgan, E.; Arnold, M.; Gina, A.; Lorenzoni, V.; Cabasag, M.L.; Laversanne, M.; Vignat, J.; Ferlay, J.; Murphy, N.; Bray, F. Global burden of colorectal cancer in 2020 and 2040: Incidence and mortality estimates from GLOBOCAN. Gut 2022. [Google Scholar] [CrossRef]
- Wong, S.H.; Yu, J. Gut microbiota in colorectal cancer: Mechanisms of action and clinical applications. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 690–704. [Google Scholar] [CrossRef] [PubMed]
- Tjalsma, H.; Boleij, A.; Marchesi, J.R.; Dutilh, B.E. A bacterial driver-passenger model for colorectal cancer: Beyond the usual suspects. Nat. Rev. Microbiol. 2012, 10, 575–582. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Pitmon, E.; Wang, K. Microbiome, inflammation and colorectal cancer. Semin. Immunol. 2017, 32, 43–53. [Google Scholar] [CrossRef]
- Long, A.G.; Lundsmith, E.T.; Hamilton, K.E. Inflammation and colorectal cancer. Curr. Colorectal. Cancer Rep. 2017, 13, 341–351. [Google Scholar] [CrossRef] [PubMed]
- Zitvogel, L.; Ma, Y.; Raoult, D.; Kroemer, G.; Gajewski, T.F. The microbiome in cancer immunotherapy: Diagnostic tools and therapeutic strategies. Science 2018, 359, 1366–1370. [Google Scholar] [CrossRef]
- Dai, Z.; Coker, O.O.; Nakatsu, G.; Wu, W.K.K.; Zhao, L.; Chen, Z.; Chan, F.K.L.; Kristiansen, K.; Sung, J.J.Y.; Wong, S.H.; et al. Multi-cohort analysis of colorectal cancer metagenome identified altered bacteria across populations and universal bacterial markers. Microbiome 2018, 6, 70. [Google Scholar] [CrossRef]
- Wirbel, J.; Pyl, P.T.; Kartal, E.; Zych, K.; Kashani, A.; Milanese, A.; Fleck, J.S.; Voigt, A.Y.; Palleja, A.; Ponnudurai, R.; et al. Meta-analysis of fecal metagenomes reveals global microbial signatures that are specific for colorectal cancer. Nat. Med. 2019, 25, 679–689. [Google Scholar] [CrossRef]
- Baxter, N.T.; Ruffin, M.T.T.; Rogers, M.A.; Schloss, P.D. Microbiota-based model improves the sensitivity of fecal immunochemical test for detecting colonic lesions. Genome Med. 2016, 8, 37. [Google Scholar] [CrossRef]
- Drewes, J.L.; White, J.R.; Dejea, C.M.; Fathi, P.; Iyadorai, T.; Vadivelu, J.; Roslani, A.C.; Wick, E.C.; Mongodin, E.F.; Loke, M.F.; et al. High-resolution bacterial 16S rRNA gene profile meta-analysis and biofilm status reveal common colorectal cancer consortia. NPJ Biofilms Microbiomes 2017, 3, 34. [Google Scholar] [CrossRef]
- Shah, M.S.; DeSantis, T.Z.; Weinmaier, T.; McMurdie, P.J.; Cope, J.L.; Altrichter, A.; Yamal, J.M.; Hollister, E.B. Leveraging sequence-based faecal microbial community survey data to identify a composite biomarker for colorectal cancer. Gut 2018, 67, 882–891. [Google Scholar] [CrossRef]
- Yu, J.; Feng, Q.; Wong, S.H.; Zhang, D.; Liang, Q.Y.; Qin, Y.; Tang, L.; Zhao, H.; Stenvang, J.; Li, Y.; et al. Metagenomic analysis of faecal microbiome as a tool towards targeted non-invasive biomarkers for colorectal cancer. Gut 2017, 66, 70–78. [Google Scholar] [CrossRef] [PubMed]
- Rubinstein, M.R.; Baik, J.E.; Lagana, S.M.; Han, R.P.; Raab, W.J.; Sahoo, D.; Dalerba, P.; Wang, T.C.; Han, Y.W. Fusobacterium nucleatum promotes colorectal cancer by inducing Wnt/beta-catenin modulator Annexin A1. EMBO Rep. 2019, 20, e47638. [Google Scholar] [CrossRef] [PubMed]
- Wei, Z.; Cao, S.; Liu, S.; Yao, Z.; Sun, T.; Li, Y.; Li, J.; Zhang, D.; Zhou, Y. Could gut microbiota serve as prognostic biomarker associated with colorectal cancer patients’ survival? A pilot study on relevant mechanism. Oncotarget 2016, 7, 46158–46172. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Cho, N.Y.; Kang, G.H. Prognostic and clinicopathological significance of Fusobacterium nucleatum in colorectal cancer: A systemic review and meta-analysis. J. Pathol. Transl. Med. 2022, 56, 144–151. [Google Scholar] [CrossRef]
- Chen, Y.; Lu, Y.; Ke, Y.; Li, Y. Prognostic impact of the Fusobacterium nucleatum status in colorectal cancers. Medicine 2019, 98, e17221. [Google Scholar] [CrossRef]
- de Carvalho, A.C.; de Mattos Pereira, L.; Datorre, J.G.; Dos Santos, W.; Berardinelli, G.N.; Matsushita, M.M.; Oliveira, M.A.; Durães, R.O.; Guimarães, D.P.; Reis, R.M. Microbiota Profile and Impact of Fusobacterium nucleatum in Colorectal Cancer Patients of Barretos Cancer Hospital. Front. Oncol. 2019, 9, 813. [Google Scholar] [CrossRef]
- Kunzmann, A.T.; Proença, M.A.; Jordao, H.W.; Jiraskova, K.; Schneiderova, M.; Levy, M.; Liska, V.; Buchler, T.; Vodickova, L.; Vymetalkova, V.; et al. Fusobacterium nucleatum tumor DNA levels are associated with survival in colorectal cancer patients. Eur. J. Clin. Microbiol. Infect. Dis. 2019, 38, 1891–1899. [Google Scholar] [CrossRef]
- Mima, K.; Nishihara, R.; Qian, Z.R.; Cao, Y.; Sukawa, Y.; Nowak, J.A.; Yang, J.; Dou, R.; Masugi, Y.; Song, M.; et al. Fusobacterium nucleatum in colorectal carcinoma tissue and patient prognosis. Gut 2016, 65, 1973–1980. [Google Scholar] [CrossRef]
- Lee, D.W.; Han, S.W.; Kang, J.K.; Bae, J.M.; Kim, H.P.; Won, J.K.; Jeong, S.Y.; Park, K.J.; Kang, G.H.; Kim, T.Y. Association Between Fusobacterium nucleatum, Pathway Mutation, and Patient Prognosis in Colorectal Cancer. Ann. Surg. Oncol. 2018, 25, 3389–3395. [Google Scholar] [CrossRef]
- Yamaoka, Y.; Suehiro, Y.; Hashimoto, S.; Hoshida, T.; Fujimoto, M.; Watanabe, M.; Imanaga, D.; Sakai, K.; Matsumoto, T.; Nishioka, M.; et al. Fusobacterium nucleatum as a prognostic marker of colorectal cancer in a Japanese population. J. Gastroenterol. 2018, 53, 517–524. [Google Scholar] [CrossRef] [PubMed]
- Purcell, R.V.; Visnovska, M.; Biggs, P.J.; Schmeier, S.; Frizelle, F.A. Distinct gut microbiome patterns associate with consensus molecular subtypes of colorectal cancer. Sci. Rep. 2017, 7, 11590. [Google Scholar] [CrossRef] [PubMed]
- Löwenmark, T.; Löfgren-Burström, A.; Zingmark, C.; Eklöf, V.; Dahlberg, M.; Wai, S.N.; Larsson, P.; Ljuslinder, I.; Edin, S.; Palmqvist, R. Parvimonas micra as a putative non-invasive faecal biomarker for colorectal cancer. Sci. Rep. 2020, 10, 15250. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Yang, M.; Wang, D.; Zhang, S.; Yan, S.; Zhu, Y.; Chen, W. Alteration of the abundance of Parvimonas micra in the gut along the adenoma-carcinoma sequence. Oncol. Lett. 2020, 20, 106. [Google Scholar] [CrossRef] [PubMed]
- Löwenmark, T.; Li, X.; Löfgren-Burström, A.; Zingmark, C.; Ling, A.; Kellgren, T.G.; Larsson, P.; Ljuslinder, I.; Wai, S.N.; Edin, S.; et al. Parvimonas micra is associated with tumour immune profiles in molecular subtypes of colorectal cancer. Cancer Immunol. Immunother. 2022, 71, 2565–2575. [Google Scholar] [CrossRef]
- Zhao, L.; Zhang, X.; Zhou, Y.; Fu, K.; Lau, H.C.; Chun, T.W.; Cheung, A.H.; Coker, O.O.; Wei, H.; Wu, W.K.; et al. Parvimonas micra promotes colorectal tumorigenesis and is associated with prognosis of colorectal cancer patients. Oncogene 2022, 41, 4200–4210. [Google Scholar] [CrossRef]
- Glimelius, B.; Melin, B.; Enblad, G.; Alafuzoff, I.; Beskow, A.; Ahlström, H.; Bill-Axelson, A.; Birgisson, H.; Björ, O.; Edqvist, P.H.; et al. U-CAN: A prospective longitudinal collection of biomaterials and clinical information from adult cancer patients in Sweden. Acta Oncol. 2018, 57, 187–194. [Google Scholar] [CrossRef]
- Li, X.; Larsson, P.; Ljuslinder, I.; Öhlund, D.; Myte, R.; Löfgren-Burström, A.; Zingmark, C.; Ling, A.; Edin, S.; Palmqvist, R. Ex Vivo Organoid Cultures Reveal the Importance of the Tumor Microenvironment for Maintenance of Colorectal Cancer Stem Cells. Cancers 2020, 12, 923. [Google Scholar] [CrossRef]
- Mima, K.; Cao, Y.; Chan, A.T.; Qian, Z.R.; Nowak, J.A.; Masugi, Y.; Shi, Y.; Song, M.; da Silva, A.; Gu, M.; et al. Fusobacterium nucleatum in Colorectal Carcinoma Tissue According to Tumor Location. Clin. Transl. Gastroenterol. 2016, 7, e200. [Google Scholar] [CrossRef]
- Guinney, J.; Dienstmann, R.; Wang, X.; de Reyniès, A.; Schlicker, A.; Soneson, C.; Marisa, L.; Roepman, P.; Nyamundanda, G.; Angelino, P.; et al. The consensus molecular subtypes of colorectal cancer. Nat. Med. 2015, 21, 1350–1356. [Google Scholar] [CrossRef]
- Boehm, E.T.; Thon, C.; Kupcinskas, J.; Steponaitiene, R.; Skieceviciene, J.; Canbay, A.; Malfertheiner, P.; Link, A. Fusobacterium nucleatum is associated with worse prognosis in Lauren’s diffuse type gastric cancer patients. Sci. Rep. 2020, 10, 16240. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Du, L.; Shi, D.; Kong, C.; Liu, J.; Liu, G.; Li, X.; Ma, Y. Dysbiosis of human gut microbiome in young-onset colorectal cancer. Nat. Commun. 2021, 12, 6757. [Google Scholar] [CrossRef] [PubMed]
- Boland, C.R.; Goel, A. Microsatellite instability in colorectal cancer. Gastroenterology 2010, 138, 2073–2087.e2073. [Google Scholar] [CrossRef] [PubMed]
- Murphy, G.; Devesa, S.S.; Cross, A.J.; Inskip, P.D.; McGlynn, K.A.; Cook, M.B. Sex disparities in colorectal cancer incidence by anatomic subsite, race and age. Int. J. Cancer 2011, 128, 1668–1675. [Google Scholar] [CrossRef] [PubMed]
- Gelsomino, F.; Barbolini, M.; Spallanzani, A.; Pugliese, G.; Cascinu, S. The evolving role of microsatellite instability in colorectal cancer: A review. Cancer Treat. Rev. 2016, 51, 19–26. [Google Scholar] [CrossRef]
- Chen, W.D.; Zhang, X.; Zhang, Y.P.; Yue, C.B.; Wang, Y.L.; Pan, H.W.; Zhang, Y.L.; Liu, H.; Zhang, Y. Fusobacterium Nucleatum is a Risk Factor for Metastatic Colorectal Cancer. Curr. Med. Sci. 2022, 42, 538–547. [Google Scholar] [CrossRef]
- Bullman, S.; Pedamallu, C.S.; Sicinska, E.; Clancy, T.E.; Zhang, X.; Cai, D.; Neuberg, D.; Huang, K.; Guevara, F.; Nelson, T.; et al. Analysis of Fusobacterium persistence and antibiotic response in colorectal cancer. Science 2017, 358, 1443–1448. [Google Scholar] [CrossRef]
- Chen, Y.; Chen, Y.; Zhang, J.; Cao, P.; Su, W.; Deng, Y.; Zhan, N.; Fu, X.; Huang, Y.; Dong, W. Fusobacterium nucleatum Promotes Metastasis in Colorectal Cancer by Activating Autophagy Signaling via the Upregulation of CARD3 Expression. Theranostics 2020, 10, 323–339. [Google Scholar] [CrossRef]
- Chen, S.; Su, T.; Zhang, Y.; Lee, A.; He, J.; Ge, Q.; Wang, L.; Si, J.; Zhuo, W.; Wang, L. Fusobacterium nucleatum promotes colorectal cancer metastasis by modulating KRT7-AS/KRT7. Gut Microbes 2020, 11, 511–525. [Google Scholar] [CrossRef]
- Yu, T.; Guo, F.; Yu, Y.; Sun, T.; Ma, D.; Han, J.; Qian, Y.; Kryczek, I.; Sun, D.; Nagarsheth, N.; et al. Fusobacterium nucleatum Promotes Chemoresistance to Colorectal Cancer by Modulating Autophagy. Cell 2017, 170, 548–563.e516. [Google Scholar] [CrossRef]
- Horiuchi, A.; Kokubu, E.; Warita, T.; Ishihara, K. Synergistic biofilm formation by Parvimonas micra and Fusobacterium nucleatum. Anaerobe 2020, 62, 102100. [Google Scholar] [CrossRef] [PubMed]
- Dejea, C.M.; Wick, E.C.; Hechenbleikner, E.M.; White, J.R.; Mark Welch, J.L.; Rossetti, B.J.; Peterson, S.N.; Snesrud, E.C.; Borisy, G.G.; Lazarev, M.; et al. Microbiota organization is a distinct feature of proximal colorectal cancers. Proc. Natl. Acad. Sci. USA 2014, 111, 18321–18326. [Google Scholar] [CrossRef] [PubMed]
- Johnson, C.H.; Dejea, C.M.; Edler, D.; Hoang, L.T.; Santidrian, A.F.; Felding, B.H.; Ivanisevic, J.; Cho, K.; Wick, E.C.; Hechenbleikner, E.M.; et al. Metabolism links bacterial biofilms and colon carcinogenesis. Cell Metab. 2015, 21, 891–897. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Faeces | ||||||
|---|---|---|---|---|---|---|
| P. micra | p Value | F. nucleatum | p Value | |||
| Fresh frozen tissue | High | Low | High | Low | ||
| P. micra, n (%) | ||||||
| High | 20 (83.3) | 4 (16.7) | <0.001 | 18 (75.0) | 6 (25.0) | <0.001 |
| Low | 22 (26.2) | 62 (73.8) | 28 (34.1) | 54 (65.9) | ||
| F. nucleatum, n (%) | ||||||
| High | 22 (55.0) | 18 (45.0) | 0.014 | 24 (61.5) | 15 (38.5) | 0.009 |
| Low | 20 (29.4) | 48 (70.6) | 23 (34.3) | 44 (65.7) | ||
| P. micra | p Value | F. nucleatum | p Value | |||
|---|---|---|---|---|---|---|
| Low | High | Low | High | |||
| Age, n (%) | ||||||
| ≤59 | 11 (61.1) | 7 (38.9) | 0.023 | 7 (38.9) | 11 (61.1) | 0.017 |
| 60–69 | 32 (91.4) | 3 (8.6) | 28 (80.0) | 7 (20.0) | ||
| 70–79 | 33 (80.5) | 8 (19.5) | 27 (65.9) | 14 (34.1) | ||
| ≥80 | 10 (62.5) | 6 (37.5) | 8 (50.0) | 8 (50.0) | ||
| Gender, n (%) | ||||||
| Male | 53 (89.8) | 6 (10.2) | 0.002 | 43 (71.7) | 17 (28.3) | 0.073 |
| Female | 33 (64.7) | 18 (35.3) | 27 (54.0) | 23 (46.0) | ||
| Location, n (%) | ||||||
| Right colon | 23 (69.7) | 10 (30.3) | 0.269 | 12 (37.5) | 20 (62.5) | 0.002 |
| Left colon | 16 (76.2) | 5 (23.8) | 16 (76.2) | 5 (23.8) | ||
| Rectum | 47 (83.9) | 9 (16.1) | 42 (73.7) | 15 (26.3) | ||
| Stage, n (%) | ||||||
| I | 29 (90.6) | 3 (9.4) | 0.171 | 23 (74.2) | 8 (25.8) | 0.425 |
| II | 28 (73.7) | 10 (26.3) | 24 (63.2) | 14 (36.8) | ||
| III | 21 (70.0) | 9 (30.0) | 17 (54.8) | 14 (45.2) | ||
| IV | 7 (77.8) | 2 (22.2) | 5 (55.6) | 4 (44.4) | ||
| Tumour grade, n (%) | ||||||
| Low grade | 71 (80.7) | 17 (19.3) | 0.143 | 58 (65.9) | 30 (34.1) | 0.207 |
| High grade | 13 (65.0) | 7 (35.0) | 10 (50.0) | 10 (50.0) | ||
| Tumour type, n (%) | ||||||
| Non-mucinous | 72 (79.1) | 19 (20.9) | 0.525 | 59 (64.8) | 32(35.2) | 0.416 |
| Mucinous | 12 (70.6) | 5 (29.4) | 9 (52.9) | 8 (47.1) | ||
| P. micra | p Value | F. nucleatum | p Value | |||
|---|---|---|---|---|---|---|
| Low | High | Low | High | |||
| KRASstatus, n (%) | ||||||
| Wild type | 51 (76.1) | 16 (23.9) | 0.638 | 43 (65.2) | 23 (34.8) | 0.691 |
| Mutant | 35 (81.4) | 8 (18.6) | 27 (61.4) | 17 (38.6) | ||
| BRAFstatus, n (%) | ||||||
| Wild type | 75 (82.4) | 16 (17.6) | 0.030 | 62 (68.1) | 29 (31.9) | 0.039 |
| Mutant | 11 (57.9) | 8 (42.1) | 8 (42.1) | 11 (57.9) | ||
| MSI status, n (%) | ||||||
| MSS | 80 (83.3) | 16 (16.7) | 0.002 | 66 (68.8) | 30 (31.3) | 0.006 |
| MSI | 6 (42.9) | 8 (57.1) | 4 (28.6) | 10 (71.4) | ||
| Univariable | Multivariable P. micra | Multivariable F. nucleatum | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| HR | 95.0% CI | p Value | HR | 95.0% CI | p Value | HR | 95.0% CI | p Value | ||
| Stage | ||||||||||
| I+II | 1 | 1 | 1 | |||||||
| III | 6.26 | (2.04–19.2) | 0.001 | 10.2 | (1.23–84.62) | 0.032 | 10.3 | (1.23–86.2) | 0.032 | |
| IV | 34.07 | (11.26–103.0) | <0.001 | 119 | (12.8–1106) | <0.001 | 91.2 | (10.0–828) | <0.001 | |
| Gender | ||||||||||
| Female | 1 | |||||||||
| Male | 1.21 | (0.61–2.42) | 0.584 | NA | - | NA | - | |||
| Age | ||||||||||
| ≤59 | 1 | |||||||||
| 60–69 | 0.42 | (0.18–1.02) | 0.055 | NA | - | NA | - | |||
| 70–79 | 0.51 | (0.21–1.22) | 0.129 | NA | - | NA | - | |||
| ≥80 | 0.56 | (0.18–1.80) | 0.333 | NA | - | NA | - | |||
| Location | ||||||||||
| Right colon | 1 | |||||||||
| Left colon | 0.81 | (0.24–2.75) | 0.731 | NA | - | NA | - | |||
| Rectum | 0.98 | (0.42–2.27) | 0.954 | NA | - | NA | - | |||
| Tumour grade | ||||||||||
| Low grade | 1 | 1 | 1 | |||||||
| High grade | 4.80 | (2.24–10.28) | <0.001 | 3.16 | (1.01–9.93) | 0.048 | 2.73 | (0.86–8.69) | 0.089 | |
| Tumour type | ||||||||||
| Non-mucinous | 1 | |||||||||
| Mucinous | 2.10 | (0.86–5.15) | 0.103 | NA | - | NA | - | |||
| KRAS | ||||||||||
| Wild type | 1 | |||||||||
| Mutant | 1.36 | (0.58–3.17) | 0.483 | NA | - | NA | - | |||
| BRAF | ||||||||||
| Wild type | 1 | |||||||||
| Mutant | 1.35 | (0.46–3.99) | 0.589 | NA | - | NA | - | |||
| MSI | ||||||||||
| MSS | 1 | |||||||||
| MSI | 0.78 | (0.18–3.34) | 0.739 | NA | - | NA | - | |||
| Tumour tissue | ||||||||||
| P. micra low | 1 | 1 | ||||||||
| P. micra high | 2.77 | (0.98–7.78) | 0.054 | 3.82 | (1.12–13.0) | 0.032 | NA | - | ||
| F. nucleatum low | 1 | 1 | ||||||||
| F. nucleatum high | 2.91 | (1.04–8.19) | 0.043 | NA | - | 2.70 | (0.83–8.80) | 0.099 | ||
| Faeces | ||||||||||
| P. micra low | 1 | |||||||||
| P. micra high | 1.22 | (0.62–2.39) | 0.567 | NA | - | NA | - | |||
| F. nucleatum low | 1 | |||||||||
| F. nucleatum high | 1.64 | (0.83–3.22) | 0.155 | NA | - | NA | - | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Löwenmark, T.; Löfgren-Burström, A.; Zingmark, C.; Ljuslinder, I.; Dahlberg, M.; Edin, S.; Palmqvist, R. Tumour Colonisation of Parvimonas micra Is Associated with Decreased Survival in Colorectal Cancer Patients. Cancers 2022, 14, 5937. https://doi.org/10.3390/cancers14235937
Löwenmark T, Löfgren-Burström A, Zingmark C, Ljuslinder I, Dahlberg M, Edin S, Palmqvist R. Tumour Colonisation of Parvimonas micra Is Associated with Decreased Survival in Colorectal Cancer Patients. Cancers. 2022; 14(23):5937. https://doi.org/10.3390/cancers14235937
Chicago/Turabian StyleLöwenmark, Thyra, Anna Löfgren-Burström, Carl Zingmark, Ingrid Ljuslinder, Michael Dahlberg, Sofia Edin, and Richard Palmqvist. 2022. "Tumour Colonisation of Parvimonas micra Is Associated with Decreased Survival in Colorectal Cancer Patients" Cancers 14, no. 23: 5937. https://doi.org/10.3390/cancers14235937
APA StyleLöwenmark, T., Löfgren-Burström, A., Zingmark, C., Ljuslinder, I., Dahlberg, M., Edin, S., & Palmqvist, R. (2022). Tumour Colonisation of Parvimonas micra Is Associated with Decreased Survival in Colorectal Cancer Patients. Cancers, 14(23), 5937. https://doi.org/10.3390/cancers14235937