
Research Progress for Targeting Deubiquitinases in Gastric Cancers


Simple Summary
Abstract
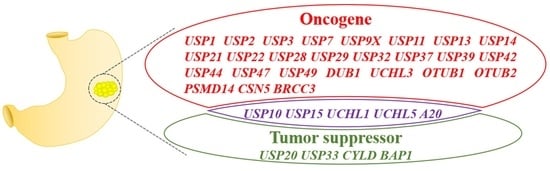
1. Introduction
2. USPs and GC
2.1. USP3
2.2. USP7 and USP11
2.3. USP9X, USP36, and USP49
2.4. USP10
2.5. USP13, USP29, and USP37
2.6. USP14
2.7. USP15
2.8. USP22
2.9. USP28
2.10. USP32
2.11. USP33
2.12. USP39
2.13. USP44
2.14. USP47
2.15. CYLD
2.16. Other USPs Involved in GC
3. UCHs and GC
4. OTUs and GC
5. JAMMs and GC
6. MJDs and GC
7. Conclusions and Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Carrasco-Garcia, E.; Garcia-Puga, M.; Arevalo, S.; Matheu, A. Towards precision medicine: Linking genetic and cellular heterogeneity in gastric cancer. Ther. Adv. Med. Oncol. 2018, 10, 1758835918794628. [Google Scholar] [CrossRef] [PubMed]
- Lei, Z.N.; Teng, Q.X.; Tian, Q.; Chen, W.; Xie, Y.; Wu, K.; Zeng, Q.; Zeng, L.; Pan, Y.; Chen, Z.S.; et al. Signaling pathways and therapeutic interventions in gastric cancer. Signal Transduct. Target. Ther. 2022, 7, 358. [Google Scholar] [CrossRef] [PubMed]
- Correa, P. Gastric cancer: Overview. Gastroenterol. Clin. N. Am. 2013, 42, 211–217. [Google Scholar] [CrossRef]
- Chen, Z.; Li, Z.; Soutto, M.; Wang, W.; Piazuelo, M.B.; Zhu, S.; Guo, Y.; Maturana, M.J.; Corvalan, A.H.; Chen, X.; et al. Integrated Analysis of Mouse and Human Gastric Neoplasms Identifies Conserved microRNA Networks in Gastric Carcinogenesis. Gastroenterology 2019, 156, 1127–1139.e8. [Google Scholar] [CrossRef]
- Zhang, D.; Chu, Y.; Qian, H.; Qian, L.; Shao, J.; Xu, Q.; Yu, L.; Li, R.; Zhang, Q.; Wu, F.; et al. Antitumor Activity of Thermosensitive Hydrogels Packaging Gambogic Acid Nanoparticles and Tumor-Penetrating Peptide iRGD Against Gastric Cancer. Int. J. Nanomed. 2020, 15, 735–747. [Google Scholar] [CrossRef]
- Komander, D.; Clague, M.J.; Urbe, S. Breaking the chains: Structure and function of the deubiquitinases. Nat. Rev. Mol. Cell Biol. 2009, 10, 550–563. [Google Scholar] [CrossRef]
- Kwon, Y.T.; Ciechanover, A. The Ubiquitin Code in the Ubiquitin-Proteasome System and Autophagy. Trends Biochem. Sci. 2017, 42, 873–886. [Google Scholar] [CrossRef]
- Csizmadia, T.; Low, P. The Role of Deubiquitinating Enzymes in the Various Forms of Autophagy. Int. J. Mol. Sci. 2020, 21, 4196. [Google Scholar] [CrossRef]
- Sun, T.; Liu, Z.; Yang, Q. The role of ubiquitination and deubiquitination in cancer metabolism. Mol. Cancer 2020, 19, 146. [Google Scholar] [CrossRef]
- Kaushal, K.; Ramakrishna, S. Deubiquitinating Enzyme-Mediated Signaling Networks in Cancer Stem Cells. Cancers 2020, 12, 3253. [Google Scholar] [CrossRef] [PubMed]
- Han, S.; Wang, R.; Zhang, Y.; Li, X.; Gan, Y.; Gao, F.; Rong, P.; Wang, W.; Li, W. The role of ubiquitination and deubiquitination in tumor invasion and metastasis. Int. J. Biol. Sci. 2022, 18, 2292–2303. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.S.; Kim, S.; Hwang, G.; Song, J. Deubiquitinases: Modulators of Different Types of Regulated Cell Death. Int. J. Mol. Sci. 2021, 22, 4352. [Google Scholar] [CrossRef] [PubMed]
- Jolly, L.A.; Kumar, R.; Penzes, P.; Piper, M.; Gecz, J. The DUB Club: Deubiquitinating Enzymes and Neurodevelopmental Disorders. Biol. Psychiatry 2022, 92, 614–625. [Google Scholar] [CrossRef] [PubMed]
- Harrigan, J.A.; Jacq, X.; Martin, N.M.; Jackson, S.P. Deubiquitylating enzymes and drug discovery: Emerging opportunities. Nat. Rev. Drug Discov. 2018, 17, 57–78. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Ye, C.; Tian, T.; Jiang, Q.; Zhao, P.; Wang, X.; Liu, F.; Shan, J.; Ruan, J. The emerging role of ubiquitin-specific protease 20 in tumorigenesis and cancer therapeutics. Cell Death Dis. 2022, 13, 434. [Google Scholar] [CrossRef]
- Meng, D.; Li, D. Ubiquitin-specific protease 1 overexpression indicates poor prognosis and promotes proliferation, migration, and invasion of gastric cancer cells. Tissue Cell 2022, 74, 101723. [Google Scholar] [CrossRef]
- Li, N.; Wu, L.; Zuo, X.; Luo, H.; Sheng, Y.; Yan, J. USP1 Promotes GC Metastasis via Stabilizing ID2. Dis. Markers 2021, 2021, 3771990. [Google Scholar] [CrossRef]
- Xiao, W.; Wang, J.; Wang, X.; Cai, S.; Guo, Y.; Ye, L.; Li, D.; Hu, A.; Jin, S.; Yuan, B.; et al. Therapeutic targeting of the USP2-E2F4 axis inhibits autophagic machinery essential for zinc homeostasis in cancer progression. Autophagy 2022, 18, 2615–2635. [Google Scholar] [CrossRef]
- Fang, C.L.; Lin, C.C.; Chen, H.K.; Hseu, Y.C.; Hung, S.T.; Sun, D.P.; Uen, Y.H.; Lin, K.Y. Ubiquitin-specific protease 3 overexpression promotes gastric carcinogenesis and is predictive of poor patient prognosis. Cancer Sci. 2018, 109, 3438–3449. [Google Scholar] [CrossRef]
- Wu, X.; Liu, M.; Zhu, H.; Wang, J.; Dai, W.; Li, J.; Zhu, D.; Tang, W.; Xiao, Y.; Lin, J.; et al. Ubiquitin-specific protease 3 promotes cell migration and invasion by interacting with and deubiquitinating SUZ12 in gastric cancer. J. Exp. Clin. Cancer Res. 2019, 38, 277. [Google Scholar] [CrossRef]
- Wu, X.; Wang, H.; Zhu, D.; Chai, Y.; Wang, J.; Dai, W.; Xiao, Y.; Tang, W.; Li, J.; Hong, L.; et al. USP3 promotes gastric cancer progression and metastasis by deubiquitination-dependent COL9A3/COL6A5 stabilisation. Cell Death Dis. 2021, 13, 10. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Jin, M.; Cao, F.; Li, J.; Wu, J.; Xu, L.; Liu, X.; Shi, Y.; Chen, W. Hsa_circ_0017639 expression promotes gastric cancer proliferation and metastasis by sponging miR-224-5p and upregulating USP3. Gene 2020, 750, 144753. [Google Scholar] [CrossRef] [PubMed]
- Jin, G.; Zhang, J.; Cao, T.; Chen, B.; Tian, Y.; Shi, Y. Exosome-mediated lncRNA SND1-IT1 from gastric cancer cells enhances malignant transformation of gastric mucosa cells via up-regulating SNAIL1. J. Transl. Med. 2022, 20, 284. [Google Scholar] [CrossRef] [PubMed]
- Georges, A.; Marcon, E.; Greenblatt, J.; Frappier, L. Identification and Characterization of USP7 Targets in Cancer Cells. Sci. Rep. 2018, 8, 15833. [Google Scholar] [CrossRef]
- Wang, Z.; Kang, W.; Li, O.; Qi, F.; Wang, J.; You, Y.; He, P.; Suo, Z.; Zheng, Y.; Liu, H.M. Abrogation of USP7 is an alternative strategy to downregulate PD-L1 and sensitize gastric cancer cells to T cells killing. Acta Pharm. Sin. B 2021, 11, 694–707. [Google Scholar] [CrossRef]
- Zhang, H.; Deng, T.; Liu, R.; Ning, T.; Yang, H.; Liu, D.; Zhang, Q.; Lin, D.; Ge, S.; Bai, M.; et al. CAF secreted miR-522 suppresses ferroptosis and promotes acquired chemo-resistance in gastric cancer. Mol. Cancer 2020, 19, 43. [Google Scholar] [CrossRef]
- Fu, X.; Xie, W.; Song, X.; Wu, K.; Xiao, L.; Liu, Y.; Zhang, L. Aberrant expression of deubiquitylating enzyme USP9X predicts poor prognosis in gastric cancer. Clin. Res. Hepatol. Gastroenterol. 2017, 41, 687–692. [Google Scholar] [CrossRef]
- Xu, X.; Wang, S.; Wang, H.; Pan, C.; Yang, W.; Yu, J. Hsa_circ_0008434 regulates USP9X expression by sponging miR-6838-5p to promote gastric cancer growth, migration and invasion. BMC Cancer 2021, 21, 1289. [Google Scholar] [CrossRef]
- Zhang, C.; Qian, H.; Liu, K.; Zhao, W.; Wang, L. A Feedback Loop Regulation of LINC01433 And YAP Promotes Malignant Behavior In Gastric Cancer Cells. OncoTargets Ther. 2019, 12, 7949–7962. [Google Scholar] [CrossRef]
- Zeng, Z.; Wu, H.X.; Zhan, N.; Huang, Y.B.; Wang, Z.S.; Yang, G.F.; Wang, P.; Fu, G.H. Prognostic significance of USP10 as a tumor-associated marker in gastric carcinoma. Tumor Biol. 2014, 35, 3845–3853. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.L.; Itahana, Y.; Lei, Z.D.; Chia, N.Y.; Wu, Y.; Yu, Y.; Zhang, S.L.; Thike, A.A.; Pandey, A.; Rozen, S.; et al. TP53 genomic status regulates sensitivity of gastric cancer cells to the histone methylation inhibitor 3-deazaneplanocin A (DZNep). Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2012, 18, 4201–4212. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Chai, L.; Ding, Z.; He, H. CircCOL1A2 Sponges MiR-1286 to Promote Cell Invasion and Migration of Gastric Cancer by Elevating Expression of USP10 to Downregulate RFC2 Ubiquitination Level. J. Microbiol. Biotechnol. 2022, 32, 938–948. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Liu, M.; He, B.; Li, Q. Inhibition of USP11 sensitizes gastric cancer to chemotherapy via suppressing RhoA and Ras-mediated signaling pathways. Clin. Res. Hepatol. Gastroenterol. 2022, 46, 101779. [Google Scholar] [CrossRef] [PubMed]
- Biterge Sut, B. Molecular profiling of immune cell-enriched Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) interacting protein USP13. Life Sci. 2020, 258, 118170. [Google Scholar] [CrossRef]
- Zhang, T.; Zheng, J.; Qiao, L.; Zhao, W. Deubiquitinase USP13 promotes the epithelial-mesenchymal transition and metastasis in gastric cancer by maintaining Snail protein. Pathol. Res. Pract. 2022, 229, 153705. [Google Scholar] [CrossRef]
- Zhu, Y.; Zhang, Y.; Sui, Z.; Zhang, Y.; Liu, M.; Tang, H. USP14 de-ubiquitinates vimentin and miR-320a modulates USP14 and vimentin to contribute to malignancy in gastric cancer cells. Oncotarget 2017, 8, 48725–48736. [Google Scholar] [CrossRef]
- Fu, Y.; Ma, G.; Liu, G.; Li, B.; Li, H.; Hao, X.; Liu, L. USP14 as a novel prognostic marker promotes cisplatin resistance via Akt/ERK signaling pathways in gastric cancer. Cancer Med. 2018, 7, 5577–5588. [Google Scholar] [CrossRef]
- Chen, X.Y.; Liang, R.; Yi, Y.C.; Fan, H.N.; Chen, M.; Zhang, J.; Zhu, J.S. The m(6)A Reader YTHDF1 Facilitates the Tumorigenesis and Metastasis of Gastric Cancer via USP14 Translation in an m(6)A-Dependent Manner. Front. Cell Dev. Biol. 2021, 9, 647702. [Google Scholar] [CrossRef]
- Zheng, J.; Zhang, H.; Ma, R.; Liu, H.; Gao, P. Long non-coding RNA KRT19P3 suppresses proliferation and metastasis through COPS7A-mediated NF-kappaB pathway in gastric cancer. Oncogene 2019, 38, 7073–7088. [Google Scholar] [CrossRef]
- Zhong, M.; Zhou, L.; Fang, Z.; Yao, Y.Y.; Zou, J.P.; Xiong, J.P.; Xiang, X.J.; Deng, J. Ubiquitin-specific protease 15 contributes to gastric cancer progression by regulating the Wnt/beta-catenin signaling pathway. World J. Gastroenterol. 2021, 27, 4221–4235. [Google Scholar] [CrossRef] [PubMed]
- Huangfu, L.; Fan, B.; Wang, G.; Gan, X.; Tian, S.; He, Q.; Yao, Q.; Shi, J.; Li, X.; Du, H.; et al. Novel prognostic marker LINC00205 promotes tumorigenesis and metastasis by competitively suppressing miRNA-26a in gastric cancer. Cell Death Discov. 2022, 8, 5. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Yang, C.; Ji, J.; Jiang, J.; Shi, M.; Cai, Q.; Yu, Y.; Zhu, Z.; Zhang, J. Deubiquitinating enzyme USP20 is a positive regulator of Claspin and suppresses the malignant characteristics of gastric cancer cells. Int. J. Oncol. 2017, 50, 1136–1146. [Google Scholar] [CrossRef] [PubMed]
- Guo, Q.; Shi, D.; Lin, L.; Li, H.; Wei, Y.; Li, B.; Wu, D. De-Ubiquitinating Enzymes USP21 Regulate MAPK1 Expression by Binding to Transcription Factor GATA3 to Regulate Tumor Growth and Cell Stemness of Gastric Cancer. Front. Cell Dev. Biol. 2021, 9, 641981. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.D.; Cui, B.B.; Sun, L.Y.; Zheng, H.Q.; Huang, Q.; Tong, J.X.; Zhang, Q.F. The co-expression of USP22 and BMI-1 may promote cancer progression and predict therapy failure in gastric carcinoma. Cell Biochem. Biophys. 2011, 61, 703–710. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Jin, Y.J.; Zhang, Y.H.; Meng, H.X.; Zhao, B.S.; Jiang, Y.; Zhu, J.W.; Liang, G.Y.; Kong, D.; Jin, X.M. Ubiquitin-specific peptidase 22 overexpression may promote cancer progression and poor prognosis in human gastric carcinoma. Transl. Res. 2015, 165, 407–416. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Fu, H.L.; Wang, Z.; Huang, H.; Ni, J.; Song, J.; Xia, Y.; Jin, W.L.; Cui, D.X. USP22 maintains gastric cancer stem cell stemness and promotes gastric cancer progression by stabilizing BMI1 protein. Oncotarget 2017, 8, 33329–33342. [Google Scholar] [CrossRef]
- Yang, F.; Zheng, Z.; Xue, X.; Zheng, L.; Qin, J.; Li, H.; Zhou, Y.; Fang, G. Targeted eradication of gastric cancer stem cells by CD44 targeting USP22 small interfering RNA-loaded nanoliposomes. Future Oncol. 2019, 15, 281–295. [Google Scholar] [CrossRef]
- Zheng, H.; Yu, J.; Li, W.; Yang, D.; Gao, C.; Zhang, Q.; Xu, L. Is co-expression of USP22 and HSP90 more effective in predicting prognosis of gastric cancer? Pathol. Res. Pract. 2019, 215, 653–659. [Google Scholar] [CrossRef]
- Liu, H.; Liu, N.; Zhao, Y.; Zhu, X.; Wang, C.; Liu, Q.; Gao, C.; Zhao, X.; Li, J. Oncogenic USP22 supports gastric cancer growth and metastasis by activating c-Myc/NAMPT/SIRT1-dependent FOXO1 and YAP signaling. Aging 2019, 11, 9643–9660. [Google Scholar] [CrossRef]
- Xiao, Y.; Liu, S.; Li, J.; Dai, W.; Tang, W.; Xiang, L.; Zhang, W.; Lin, J.; Wang, J.; Wu, X.; et al. The POU2F1/miR-4490/USP22 axis regulates cell proliferation and metastasis in gastric cancer. Cell. Oncol. (Dordr) 2020, 43, 1017–1033. [Google Scholar] [CrossRef]
- Lim, C.; Xu, J.C.; Chen, T.Y.; Xu, J.X.; Chen, W.F.; Hu, J.W.; Li, Q.L.; Zhang, Y.Q. Ubiquitin-specific peptide 22 acts as an oncogene in gastric cancer in a son of sevenless 1-dependent manner. Cancer Cell Int. 2020, 20, 45. [Google Scholar] [CrossRef]
- Zhao, L.J.; Zhang, T.; Feng, X.J.; Chang, J.; Suo, F.Z.; Ma, J.L.; Liu, Y.J.; Liu, Y.; Zheng, Y.C.; Liu, H.M. USP28 contributes to the proliferation and metastasis of gastric cancer. J. Cell. Biochem. 2018, 120, 7657–7666. [Google Scholar] [CrossRef]
- Qian, W.; Li, Q.; Wu, X.; Li, W.; Li, Q.; Zhang, J.; Li, M.; Zhang, D.; Zhao, H.; Zou, X.; et al. Deubiquitinase USP29 promotes gastric cancer cell migration by cooperating with phosphatase SCP1 to stabilize Snail protein. Oncogene 2020, 39, 6802–6815. [Google Scholar] [CrossRef]
- Dou, N.; Hu, Q.; Li, L.; Wu, Q.; Li, Y.; Gao, Y. USP32 promotes tumorigenesis and chemoresistance in gastric carcinoma via upregulation of SMAD2. Int. J. Biol. Sci. 2020, 16, 1648–1657. [Google Scholar] [CrossRef]
- Chen, Y.; Pang, X.; Ji, L.; Sun, Y.; Ji, Y. Reduced Expression of Deubiquitinase USP33 Is Associated with Tumor Progression and Poor Prognosis of Gastric Adenocarcinoma. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2018, 24, 3496–3505. [Google Scholar] [CrossRef]
- Xia, Y.; Wang, L.; Xu, Z.; Kong, R.; Wang, F.; Yin, K.; Xu, J.; Li, B.; He, Z.; Wang, L.; et al. Reduced USP33 expression in gastric cancer decreases inhibitory effects of Slit2-Robo1 signalling on cell migration and EMT. Cell Prolif. 2019, 52, e12606. [Google Scholar] [CrossRef]
- Wang, D.; Li, Z.; Li, X.; Yan, C.; Yang, H.; Zhuang, T.; Wang, X.; Zang, Y.; Liu, Z.; Wang, T.; et al. DUB1 suppresses Hippo signaling by modulating TAZ protein expression in gastric cancer. J. Exp. Clin. Cancer Res. 2022, 41, 219. [Google Scholar] [CrossRef]
- Wu, L.; Zhao, N.; Zhou, Z.; Chen, J.; Han, S.; Zhang, X.; Bao, H.; Yuan, W.; Shu, X. PLAGL2 promotes the proliferation and migration of gastric cancer cells via USP37-mediated deubiquitination of Snail1. Theranostics 2021, 11, 700–714. [Google Scholar] [CrossRef]
- Wang, X.; Yu, Q.; Huang, L.; Yu, P. Lentivirus-mediated inhibition of USP39 suppresses the growth of gastric cancer cells via PARP activation. Mol. Med. Rep. 2016, 14, 301–306. [Google Scholar] [CrossRef]
- Dong, X.; Su, H.; Jiang, F.; Li, H.; Shi, G.; Fan, L. miR-133a, directly targeted USP39, suppresses cell proliferation and predicts prognosis of gastric cancer. Oncol. Lett. 2018, 15, 8311–8318. [Google Scholar] [CrossRef] [PubMed]
- Hou, K.; Zhu, Z.; Wang, Y.; Zhang, C.; Yu, S.; Zhu, Q.; Yan, B. Overexpression and Biological Function of Ubiquitin-Specific Protease 42 in Gastric Cancer. PLoS ONE 2016, 11, e0152997. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, S.; Oki, E.; Ando, K.; Iimori, M.; Nakaji, Y.; Nakashima, Y.; Saeki, H.; Oda, Y.; Maehara, Y. High ubiquitin-specific protease 44 expression induces DNA aneuploidy and provides independent prognostic information in gastric cancer. Cancer Med. 2017, 6, 1453–1464. [Google Scholar] [CrossRef] [PubMed]
- Xiang, T.; Jiang, H.S.; Zhang, B.T.; Liu, G. CircFOXO3 functions as a molecular sponge for miR-143-3p to promote the progression of gastric carcinoma via upregulating USP44. Gene 2020, 753, 144798. [Google Scholar] [CrossRef]
- Zhang, B.; Yin, Y.; Hu, Y.; Zhang, J.; Bian, Z.; Song, M.; Hua, D.; Huang, Z. MicroRNA-204-5p inhibits gastric cancer cell proliferation by downregulating USP47 and RAB22A. Med. Oncol. 2015, 32, 331. [Google Scholar] [CrossRef]
- Naghavi, L.; Schwalbe, M.; Ghanem, A.; Naumann, M. Deubiquitinylase USP47 Promotes RelA Phosphorylation and Survival in Gastric Cancer Cells. Biomedicines 2018, 6, 62. [Google Scholar] [CrossRef]
- Liu, Z.; Li, J.; Ding, Y.; Ma, M.; Chen, J.; Lei, W.; Li, L.; Yao, Y.; Yu, X.; Zhong, M.; et al. USP49 mediates tumor progression and poor prognosis through a YAP1-dependent feedback loop in gastric cancer. Oncogene 2022, 41, 2555–2570. [Google Scholar] [CrossRef]
- Xia, J.T.; Chen, L.Z.; Jian, W.H.; Wang, K.B.; Yang, Y.Z.; He, W.L.; He, Y.L.; Chen, D.; Li, W. MicroRNA-362 induces cell proliferation and apoptosis resistance in gastric cancer by activation of NF-kappaB signaling. J. Transl. Med. 2014, 12, 33. [Google Scholar] [CrossRef]
- Zhang, L.; Ding, Y.; Yuan, Z.; Liu, J.; Sun, J.; Lei, F.; Wu, S.; Li, S.; Zhang, D. MicroRNA-500 sustains nuclear factor-kappaB activation and induces gastric cancer cell proliferation and resistance to apoptosis. Oncotarget 2015, 6, 2483–2495. [Google Scholar] [CrossRef]
- Sun, B.; Li, L.; Ma, W.; Wang, S.; Huang, C. MiR-130b inhibits proliferation and induces apoptosis of gastric cancer cells via CYLD. Tumor Biol. 2016, 37, 7981–7987. [Google Scholar] [CrossRef]
- Zhu, M.; Zhou, X.; Du, Y.; Huang, Z.; Zhu, J.; Xu, J.; Cheng, G.; Shu, Y.; Liu, P.; Zhu, W.; et al. miR-20a induces cisplatin resistance of a human gastric cancer cell line via targeting CYLD. Mol. Med. Rep. 2016, 14, 1742–1750. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.F.; Gong, F.M.; Wang, B.S.; Zheng, W. MiR-425-5p promotes tumor progression via modulation of CYLD in gastric cancer. Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 2130–2136. [Google Scholar] [PubMed]
- Ghadami, E.; Nikbakhsh, N.; Fattahi, S.; Kosari-Monfared, M.; Ranaee, M.; Taheri, H.; Amjadi-Moheb, F.; Godazandeh, G.; Shafaei, S.; Nosrati, A.; et al. Epigenetic alterations of CYLD promoter modulate its expression in gastric adenocarcinoma: A footprint of infections. J. Cell. Physiol. 2019, 234, 4115–4124. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, Q.; Xu, G.; Meng, N.; Huang, X.; Jiang, Z.; Chen, C.; Zhang, Y.; Chen, J.; Li, A.; et al. The long noncoding RNA CRAL reverses cisplatin resistance via the miR-505/CYLD/AKT axis in human gastric cancer cells. RNA Biol. 2020, 17, 1576–1589. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Liu, J.; Pan, X.; Peng, C.; Xiong, B.; Feng, M.; Yang, X. miR-454 promotes survival and induces oxaliplatin resistance in gastric carcinoma cells by targeting CYLD. Exp. Ther. Med. 2020, 19, 3604–3610. [Google Scholar] [CrossRef] [PubMed]
- Cui, H.Y.; Rong, J.S.; Chen, J.; Guo, J.; Zhu, J.Q.; Ruan, M.; Zuo, R.R.; Zhang, S.S.; Qi, J.M.; Zhang, B.H. Exosomal microRNA-588 from M2 polarized macrophages contributes to cisplatin resistance of gastric cancer cells. World J. Gastroenterol. 2021, 27, 6079–6092. [Google Scholar] [CrossRef]
- Yue, B.; Cui, R.; Zheng, R.; Jin, W.; Song, C.; Bao, T.; Wang, M.; Yu, F.; Zhao, E. Essential role of ALKBH5-mediated RNA demethylation modification in bile acid-induced gastric intestinal metaplasia. Mol. Ther. Nucleic Acids 2021, 26, 458–472. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, K.; Park, H.L.; Kim, M.S.; Osada, M.; Tokumaru, Y.; Inoue, H.; Mori, M.; Sidransky, D. PGP9.5 methylation in diffuse-type gastric cancer. Cancer Res. 2006, 66, 3921–3927. [Google Scholar] [CrossRef]
- Tokumaru, Y.; Yamashita, K.; Kim, M.S.; Park, H.L.; Osada, M.; Mori, M.; Sidransky, D. The role of PGP9.5 as a tumor suppressor gene in human cancer. Int. J. Cancer 2008, 123, 753–759. [Google Scholar] [CrossRef]
- Yu, J.; Tao, Q.; Cheung, K.F.; Jin, H.; Poon, F.F.; Wang, X.; Li, H.; Cheng, Y.Y.; Rocken, C.; Ebert, M.P.; et al. Epigenetic identification of ubiquitin carboxyl-terminal hydrolase L1 as a functional tumor suppressor and biomarker for hepatocellular carcinoma and other digestive tumors. Hepatology 2008, 48, 508–518. [Google Scholar] [CrossRef]
- Kim, J.H.; Jung, E.J.; Lee, H.S.; Kim, M.A.; Kim, W.H. Comparative analysis of DNA methylation between primary and metastatic gastric carcinoma. Oncol. Rep. 2009, 21, 1251–1259. [Google Scholar] [CrossRef]
- Wang, G.; Zhang, W.; Zhou, B.; Jin, C.; Wang, Z.; Yang, Y.; Wang, Z.; Chen, Y.; Feng, X. The diagnosis value of promoter methylation of UCHL1 in the serum for progression of gastric cancer. Biomed. Res. Int. 2015, 2015, 741030. [Google Scholar] [CrossRef]
- Gu, Y.Y.; Yang, M.; Zhao, M.; Luo, Q.; Yang, L.; Peng, H.; Wang, J.; Huang, S.K.; Zheng, Z.X.; Yuan, X.H.; et al. The de-ubiquitinase UCHL1 promotes gastric cancer metastasis via the Akt and Erk1/2 pathways. Tumor Biol. 2015, 36, 8379–8387. [Google Scholar] [CrossRef]
- Yang, H.; Zhang, C.; Fang, S.; Ou, R.; Li, W.; Xu, Y. UCH-LI acts as a novel prognostic biomarker in gastric cardiac adenocarcinoma. Int. J. Clin. Exp. Pathol. 2015, 8, 13957–13967. [Google Scholar]
- Peng, J.; Meng, Q.; Shu, Y.; Qu, Z. UCHL3 stimulates metastasis of gastric cancer through upregulating IGF2. Minerva Gastroenterol. (Torino) 2022, 68, 343–344. [Google Scholar] [CrossRef]
- Arpalahti, L.; Laitinen, A.; Hagstrom, J.; Mustonen, H.; Kokkola, A.; Bockelman, C.; Haglund, C.; Holmberg, C.I. Positive cytoplasmic UCHL5 tumor expression in gastric cancer is linked to improved prognosis. PLoS ONE 2018, 13, e0193125. [Google Scholar] [CrossRef]
- Zhang, Z.; Hu, X.; Kuang, J.; Liao, J.; Yuan, Q. LncRNA DRAIC inhibits proliferation and metastasis of gastric cancer cells through interfering with NFRKB deubiquitination mediated by UCHL5. Cell. Mol. Biol. Lett. 2020, 25, 29. [Google Scholar] [CrossRef]
- Yan, S.; He, F.; Luo, R.; Wu, H.; Huang, M.; Huang, C.; Li, Y.; Zhou, Z. Decreased expression of BRCA1-associated protein 1 predicts unfavorable survival in gastric adenocarcinoma. Tumor Biol. 2016, 37, 6125–6133. [Google Scholar] [CrossRef]
- Weng, W.; Zhang, Q.; Xu, M.; Wu, Y.; Zhang, M.; Shen, C.; Chen, X.; Wang, Y.; Sheng, W. OTUB1 promotes tumor invasion and predicts a poor prognosis in gastric adenocarcinoma. Am. J. Transl. Res. 2016, 8, 2234–2244. [Google Scholar]
- Ouyang, S.; Zeng, Z.; Liu, Z.; Zhang, Z.; Sun, J.; Wang, X.; Ma, M.; Ye, X.; Yu, J.; Kang, W. OTUB2 regulates KRT80 stability via deubiquitination and promotes tumour proliferation in gastric cancer. Cell Death Discov. 2022, 8, 45. [Google Scholar] [CrossRef]
- Liu, G.; Guo, W.; Qin, J.; Lin, Z. OTUB2 Facilitates Tumorigenesis of Gastric Cancer Through Promoting KDM1A-Mediated Stem Cell-Like Properties. Front. Oncol. 2021, 11, 711735. [Google Scholar] [CrossRef]
- Wisnieski, F.; Santos, L.C.; Calcagno, D.Q.; Geraldis, J.C.; Gigek, C.O.; Anauate, A.C.; Chen, E.S.; Rasmussen, L.T.; Payao, S.L.M.; Artigiani, R.; et al. The impact of DNA demethylation on the upregulation of the NRN1 and TNFAIP3 genes associated with advanced gastric cancer. J. Mol. Med. 2020, 98, 707–717. [Google Scholar] [CrossRef]
- Du, B.; Liu, M.; Li, C.; Geng, X.; Zhang, X.; Ning, D.; Liu, M. The potential role of TNFAIP3 in malignant transformation of gastric carcinoma. Pathol. Res. Pract. 2019, 215, 152471. [Google Scholar] [CrossRef]
- Guo, T.; Zhang, Y.; Qu, X.; Che, X.; Li, C.; Fan, Y.; Wan, X.; Ma, R.; Hou, K.; Zhou, H.; et al. miR-200a enhances TRAIL-induced apoptosis in gastric cancer cells by targeting A20. Cell Biol. Int. 2018, 42, 506–514. [Google Scholar] [CrossRef]
- Maeda, S.; Otsuka, M.; Hirata, Y.; Mitsuno, Y.; Yoshida, H.; Shiratori, Y.; Masuho, Y.; Muramatsu, M.; Seki, N.; Omata, M. cDNA microarray analysis of Helicobacter pylori-mediated alteration of gene expression in gastric cancer cells. Biochem. Biophy.s Res. Commun. 2001, 284, 443–449. [Google Scholar] [CrossRef]
- Lim, M.C.C.; Maubach, G.; Sokolova, O.; Feige, M.H.; Diezko, R.; Buchbinder, J.; Backert, S.; Schluter, D.; Lavrik, I.N.; Naumann, M. Pathogen-induced ubiquitin-editing enzyme A20 bifunctionally shuts off NF-kappaB and caspase-8-dependent apoptotic cell death. Cell Death Differ. 2017, 24, 1621–1631. [Google Scholar] [CrossRef]
- Jantaree, P.; Chaithongyot, S.; Sokolova, O.; Naumann, M. USP48 and A20 synergistically promote cell survival in Helicobacter pylori infection. Cell. Mol. Life Sci. 2022, 79, 461. [Google Scholar] [CrossRef]
- Jantaree, P.; Yu, Y.; Chaithongyot, S.; Tager, C.; Sarabi, M.A.; Meyer, T.F.; Boccellato, F.; Maubach, G.; Naumann, M. Human gastric fibroblasts ameliorate A20-dependent cell survival in co-cultured gastric epithelial cells infected by Helicobacter pylori. Biochim. Biophys. Acta Mol. Cell Res. 2022, 1869, 119364. [Google Scholar] [CrossRef]
- Sun, F.; Ni, Y.; Zhu, H.; Fang, J.; Wang, H.; Xia, J.; Ding, F.; Shen, H.; Shao, S. microRNA-29a-3p, Up-Regulated in Human Gastric Cells and Tissues with H.Pylori Infection, Promotes the Migration of GES-1 Cells via A20-Mediated EMT Pathway. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2018, 51, 1250–1263. [Google Scholar] [CrossRef]
- Li, J.; Li, Y.; Xu, F.; Sun, B.; Yang, L.; Wang, H. Deubiquitinating enzyme PSMD14 facilitates gastric carcinogenesis through stabilizing PTBP1. Exp. Cell Res. 2022, 415, 113148. [Google Scholar] [CrossRef]
- Kim, J.H.; Choi, J.K.; Cinghu, S.; Jang, J.W.; Lee, Y.S.; Li, Y.H.; Goh, Y.M.; Chi, X.Z.; Lee, K.S.; Wee, H.; et al. Jab1/CSN5 induces the cytoplasmic localization and degradation of RUNX3. J. Cell. Biochem. 2009, 107, 557–565. [Google Scholar] [CrossRef] [PubMed]
- Sang, M.M.; Du, W.Q.; Zhang, R.Y.; Zheng, J.N.; Pei, D.S. Suppression of CSN5 promotes the apoptosis of gastric cancer cells through regulating p53-related apoptotic pathways. Bioorg. Med. Chem. Lett. 2015, 25, 2897–2901. [Google Scholar] [CrossRef]
- Wang, L.; Du, W.Q.; Xie, M.; Liu, M.R.; Huo, F.C.; Yang, J.; Pei, D.S. Jab1 promotes gastric cancer tumorigenesis via non-ubiquitin proteasomal degradation of p14ARF. Gastric Cancer 2020, 23, 1003–1017. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Zhou, C.; Zhou, L.; Wang, Z.; Zhu, X.; Mu, X. Overexpression of DAPK1-mediated inhibition of IKKbeta/CSN5/PD-L1 axis enhances natural killer cell killing ability and inhibits tumor immune evasion in gastric cancer. Cell. Immunol. 2022, 372, 104469. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Zhang, Y.; Ding, M.; Xu, R. Long noncoding RNA TMPO-AS1/miR-126-5p/BRCC3 axis accelerates gastric cancer progression and angiogenesis via activating PI3K/Akt/mTOR pathway. J. Gastroenterol. Hepatol. 2021, 36, 1877–1888. [Google Scholar] [CrossRef] [PubMed]
- Zeng, L.X.; Tang, Y.; Ma, Y. Ataxin-3 expression correlates with the clinicopathologic features of gastric cancer. Int. J. Clin. Exp. Med. 2014, 7, 973–981. [Google Scholar] [PubMed]
- Gavory, G.; O’Dowd, C.R.; Helm, M.D.; Flasz, J.; Arkoudis, E.; Dossang, A.; Hughes, C.; Cassidy, E.; McClelland, K.; Odrzywol, E.; et al. Discovery and characterization of highly potent and selective allosteric USP7 inhibitors. Nat. Chem. Biol. 2018, 14, 118–125. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Liu, Y.; Yang, H.; Liu, H.M. Design, synthesis, biological evaluation and structure-activity relationship study of quinazolin-4(3H)-one derivatives as novel USP7 inhibitors. Eur. J. Med. Chem. 2021, 216, 113291. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.H.; Lee, M.J.; Park, S.; Oh, D.C.; Elsasser, S.; Chen, P.C.; Gartner, C.; Dimova, N.; Hanna, J.; Gygi, S.P.; et al. Enhancement of proteasome activity by a small-molecule inhibitor of USP14. Nature 2010, 467, 179–184. [Google Scholar] [CrossRef]
- Liu, Z.; Zhao, T.; Li, Z.; Sun, K.; Fu, Y.; Cheng, T.; Guo, J.; Yu, B.; Shi, X.; Liu, H. Discovery of [1,2,3]triazolo[4,5-d]pyrimidine derivatives as highly potent, selective, and cellularly active USP28 inhibitors. Acta Pharm. Sin. B 2020, 10, 1476–1491. [Google Scholar] [CrossRef]
- Hu, Y.; Yu, K.; Wang, G.; Zhang, D.; Shi, C.; Ding, Y.; Hong, D.; Zhang, D.; He, H.; Sun, L.; et al. Lanatoside C inhibits cell proliferation and induces apoptosis through attenuating Wnt/beta-catenin/c-Myc signaling pathway in human gastric cancer cell. Biochem. Pharmacol. 2018, 150, 280–292. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.A.; Jeon, Y.K.; Nam, M.J. Galangin induces apoptosis in gastric cancer cells via regulation of ubiquitin carboxy-terminal hydrolase isozyme L1 and glutathione S-transferase P. Food Chem. Toxicol. 2012, 50, 684–688. [Google Scholar] [CrossRef]
- Ye, Y.; Li, X.; Feng, G.; Ma, Y.; Ye, F.; Shen, H.; Sun, K.; Lu, R.; Miao, S. 3,3′-Diindolylmethane induces ferroptosis by BAP1-IP3R axis in BGC-823 gastric cancer cells. Anticancer Drugs 2022, 33, 362–370. [Google Scholar] [CrossRef] [PubMed]
- Comet, I.; Riising, E.M.; Leblanc, B.; Helin, K. Maintaining cell identity: PRC2-mediated regulation of transcription and cancer. Nat. Rev. Cancer 2016, 16, 803–810. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Ping, S.; Xu, Y.; Wang, M.; Jiang, X.; Xiong, L.; Zhang, L.; Yu, H.; Xiong, Z. Non-Coding RNAs in Gastric Cancer: From Malignant Hallmarks to Clinical Applications. Front. Cell Dev. Biol. 2021, 9, 732036. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhang, J.; Cao, G.; Hua, J.; Shan, G.; Lin, W. Emerging roles of circular RNAs in gastric cancer metastasis and drug resistance. J. Exp. Clin. Cancer Res. 2022, 41, 218. [Google Scholar] [CrossRef]
- Lu, Y.; Li, K.; Gao, Y.; Liang, W.; Wang, X.; Chen, L. CircRNAs in gastric cancer: Current research and potential clinical implications. FEBS Lett. 2021, 595, 2644–2654. [Google Scholar] [CrossRef]
- Gong, Y.Q.; Lu, T.L.; Hou, F.T.; Chen, C.W. Antisense long non-coding RNAs in gastric cancer. Clin. Chim. Acta Int. J. Clin. Chem. 2022, 534, 128–137. [Google Scholar] [CrossRef]
- Al-Eidan, A.; Wang, Y.; Skipp, P.; Ewing, R.M. The USP7 protein interaction network and its roles in tumorigenesis. Genes Dis. 2022, 9, 41–50. [Google Scholar] [CrossRef]
- Nininahazwe, L.; Liu, B.; He, C.; Zhang, H.; Chen, Z.S. The emerging nature of Ubiquitin-specific protease 7 (USP7): A new target in cancer therapy. Drug Discov. Today 2021, 26, 490–502. [Google Scholar] [CrossRef]
- Harakandi, C.; Nininahazwe, L.; Xu, H.; Liu, B.; He, C.; Zheng, Y.C.; Zhang, H. Recent advances on the intervention sites targeting USP7-MDM2-p53 in cancer therapy. Bioorg. Chem. 2021, 116, 105273. [Google Scholar] [CrossRef] [PubMed]
- Coombs, N.; Sompallae, R.; Olbermann, P.; Gastaldello, S.; Goppel, D.; Masucci, M.G.; Josenhans, C. Helicobacter pylori affects the cellular deubiquitinase USP7 and ubiquitin-regulated components TRAF6 and the tumour suppressor p53. Int. J. Med. Microbiol. 2011, 301, 213–224. [Google Scholar] [CrossRef] [PubMed]
- Murtaza, M.; Jolly, L.A.; Gecz, J.; Wood, S.A. La FAM fatale: USP9X in development and disease. Cell. Mol. Life Sci. 2015, 72, 2075–2089. [Google Scholar] [CrossRef] [PubMed]
- Kang, W.; Cheng, A.S.; Yu, J.; To, K.F. Emerging role of Hippo pathway in gastric and other gastrointestinal cancers. World J. Gastroenterol. 2016, 22, 1279–1288. [Google Scholar] [CrossRef] [PubMed]
- Hergovich, A. Mammalian Hippo signalling: A kinase network regulated by protein-protein interactions. Biochem. Soc. Trans. 2012, 40, 124–128. [Google Scholar] [CrossRef]
- Zhao, B.; Li, L.; Tumaneng, K.; Wang, C.Y.; Guan, K.L. A coordinated phosphorylation by Lats and CK1 regulates YAP stability through SCF(beta-TRCP). Genes Dev. 2010, 24, 72–85. [Google Scholar] [CrossRef]
- Mussell, A.; Frangou, C.; Zhang, J. Regulation of the Hippo signaling pathway by deubiquitinating enzymes in cancer. Genes Dis. 2019, 6, 335–341. [Google Scholar] [CrossRef]
- Li, L.; Liu, T.; Li, Y.; Wu, C.; Luo, K.; Yin, Y.; Chen, Y.; Nowsheen, S.; Wu, J.; Lou, Z.; et al. The deubiquitinase USP9X promotes tumor cell survival and confers chemoresistance through YAP1 stabilization. Oncogene 2018, 37, 2422–2431. [Google Scholar] [CrossRef]
- Kim, M.S.; Yoo, K.J.; Kang, I.; Chung, H.M.; Baek, K.H. A novel cysteine protease HeLa DUB-1 responsible for cleaving the ubiquitin in human ovarian cancer cells. Int. J. Oncol. 2004, 25, 373–379. [Google Scholar] [CrossRef]
- Kim, M.S.; Kim, Y.K.; Kim, Y.S.; Seong, M.; Choi, J.K.; Baek, K.H. Deubiquitinating enzyme USP36 contains the PEST motif and is polyubiquitinated. Biochem. Biophys. Res. Commun. 2005, 330, 797–804. [Google Scholar] [CrossRef]
- Tao, L.; Liu, X.; Jiang, X.; Zhang, K.; Wang, Y.; Li, X.; Jiang, S.; Han, T. USP10 as a Potential Therapeutic Target in Human Cancers. Genes 2022, 13, 831. [Google Scholar] [CrossRef]
- Li, D.; Zeng, Z.; Yu, T.; Qin, J.; Wu, J.; Song, J.C.; Zhou, Z.Y.; Yuan, J.P. Expression and clinical implication of S100A12 in gastric carcinoma. Tumor Biol. 2016, 37, 6551–6559. [Google Scholar] [CrossRef]
- Peng, X.C.; Zeng, Z.; Huang, Y.N.; Deng, Y.C.; Fu, G.H. Clinical significance of TM4SF1 as a tumor suppressor gene in gastric cancer. Cancer Med. 2018, 7, 2592–2600. [Google Scholar] [CrossRef]
- Kwon, S.K.; Saindane, M.; Baek, K.H. p53 stability is regulated by diverse deubiquitinating enzymes. Biochim. Biophys. Acta Rev. Cancer 2017, 1868, 404–411. [Google Scholar] [CrossRef]
- Dong, B.; Wu, Y. Epigenetic Regulation and Post-Translational Modifications of SNAI1 in Cancer Metastasis. Int. J. Mol. Sci. 2021, 22, 11062. [Google Scholar] [CrossRef]
- Zhou, B.P.; Deng, J.; Xia, W.; Xu, J.; Li, Y.M.; Gunduz, M.; Hung, M.C. Dual regulation of Snail by GSK-3beta-mediated phosphorylation in control of epithelial-mesenchymal transition. Nat. Cell Biol. 2004, 6, 931–940. [Google Scholar] [CrossRef]
- de Poot, S.A.H.; Tian, G.; Finley, D. Meddling with Fate: The Proteasomal Deubiquitinating Enzymes. J. Mol. Biol. 2017, 429, 3525–3545. [Google Scholar] [CrossRef]
- Wang, F.; Ning, S.; Yu, B.; Wang, Y. USP14: Structure, Function, and Target Inhibition. Front. Pharmacol. 2021, 12, 801328. [Google Scholar] [CrossRef]
- Zhu, Y.; Zhang, C.; Gu, C.; Li, Q.; Wu, N. Function of Deubiquitinating Enzyme USP14 as Oncogene in Different Types of Cancer. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2016, 38, 993–1002. [Google Scholar] [CrossRef]
- Anita, R.; Paramasivam, A.; Priyadharsini, J.V.; Chitra, S. The m6A readers YTHDF1 and YTHDF3 aberrations associated with metastasis and predict poor prognosis in breast cancer patients. Am. J. Cancer Res. 2020, 10, 2546–2554. [Google Scholar]
- Pi, J.; Wang, W.; Ji, M.; Wang, X.; Wei, X.; Jin, J.; Liu, T.; Qiang, J.; Qi, Z.; Li, F.; et al. YTHDF1 Promotes Gastric Carcinogenesis by Controlling Translation of FZD7. Cancer Res. 2021, 81, 2651–2665. [Google Scholar] [CrossRef]
- Schweitzer, K.; Bozko, P.M.; Dubiel, W.; Naumann, M. CSN controls NF-kappaB by deubiquitinylation of IkappaBalpha. EMBO J. 2007, 26, 1532–1541. [Google Scholar] [CrossRef]
- Das, T.; Song, E.J.; Kim, E.E. The Multifaceted Roles of USP15 in Signal Transduction. Int. J. Mol. Sci. 2021, 22, 4728. [Google Scholar] [CrossRef]
- Glinsky, G.V. Genomic models of metastatic cancer: Functional analysis of death-from-cancer signature genes reveals aneuploid, anoikis-resistant, metastasis-enabling phenotype with altered cell cycle control and activated Polycomb Group (PcG) protein chromatin silencing pathway. Cell Cycle 2006, 5, 1208–1216. [Google Scholar] [CrossRef]
- Feng, T.; Ling, S.; Xu, C.; Ying, L.; Su, D.; Xu, X. Ubiquitin-specific peptidase 22 in cancer. Cancer Lett. 2021, 514, 30–37. [Google Scholar] [CrossRef]
- Atanassov, B.S.; Mohan, R.D.; Lan, X.; Kuang, X.; Lu, Y.; Lin, K.; McIvor, E.; Li, W.; Zhang, Y.; Florens, L.; et al. ATXN7L3 and ENY2 Coordinate Activity of Multiple H2B Deubiquitinases Important for Cellular Proliferation and Tumor Growth. Mol. Cell 2016, 62, 558–571. [Google Scholar] [CrossRef]
- Qiu, G.Z.; Mao, X.Y.; Ma, Y.; Gao, X.C.; Wang, Z.; Jin, M.Z.; Sun, W.; Zou, Y.X.; Lin, J.; Fu, H.L.; et al. Ubiquitin-specific protease 22 acts as an oncoprotein to maintain glioma malignancy through deubiquitinating B cell-specific Moloney murine leukemia virus integration site 1 for stabilization. Cancer Sci. 2018, 109, 2199–2210. [Google Scholar] [CrossRef]
- Kim, D.; Hong, A.; Park, H.I.; Shin, W.H.; Yoo, L.; Jeon, S.J.; Chung, K.C. Deubiquitinating enzyme USP22 positively regulates c-Myc stability and tumorigenic activity in mammalian and breast cancer cells. J. Cell. Physiol. 2017, 232, 3664–3676. [Google Scholar] [CrossRef]
- Vigil, D.; Cherfils, J.; Rossman, K.L.; Der, C.J. Ras superfamily GEFs and GAPs: Validated and tractable targets for cancer therapy? Nat. Rev. Cancer 2010, 10, 842–857. [Google Scholar] [CrossRef]
- Wang, Y.; Jia, Z.; Gao, J.; Zhou, T.; Zhang, X.; Zu, G. Clinicopathological and Prognostic Value of USP22 Expression in Gastric Cancer: A Systematic Review and Meta-Analysis and Database Validation. Front. Surg. 2022, 9, 920595. [Google Scholar] [CrossRef]
- Wang, X.; Liu, Z.; Zhang, L.; Yang, Z.; Chen, X.; Luo, J.; Zhou, Z.; Mei, X.; Yu, X.; Shao, Z.; et al. Targeting deubiquitinase USP28 for cancer therapy. Cell Death Dis. 2018, 9, 186. [Google Scholar] [CrossRef] [PubMed]
- Prieto-Garcia, C.; Tomaskovic, I.; Shah, V.J.; Dikic, I.; Diefenbacher, M. USP28: Oncogene or Tumor Suppressor? A Unifying Paradigm for Squamous Cell Carcinoma. Cells 2021, 10, 2652. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Wang, Y.; Yang, X.H.; Kang, T.; Zhao, Y.; Wang, C.; Evers, B.M.; Zhou, B.P. The deubiquitinase USP28 stabilizes LSD1 and confers stem-cell-like traits to breast cancer cells. Cell Rep. 2013, 5, 224–236. [Google Scholar] [CrossRef] [PubMed]
- Paulding, C.A.; Ruvolo, M.; Haber, D.A. The Tre2 (USP6) oncogene is a hominoid-specific gene. Proc. Natl. Acad. Sci. USA 2003, 100, 2507–2511. [Google Scholar] [CrossRef]
- Akhavantabasi, S.; Akman, H.B.; Sapmaz, A.; Keller, J.; Petty, E.M.; Erson, A.E. USP32 is an active, membrane-bound ubiquitin protease overexpressed in breast cancers. Mamm. Genome 2010, 21, 388–397. [Google Scholar] [CrossRef]
- Chen, S.; Chen, X.; Li, Z.; Mao, J.; Jiang, W.; Zhu, Z.; Li, Y.; Jiang, Z.; Zhao, W.; Tan, G.; et al. Identification of ubiquitin-specific protease 32 as an oncogene in glioblastoma and the underlying mechanisms. Sci. Rep. 2022, 12, 6445. [Google Scholar] [CrossRef]
- Van Leuken, R.J.; Luna-Vargas, M.P.; Sixma, T.K.; Wolthuis, R.M.; Medema, R.H. Usp39 is essential for mitotic spindle checkpoint integrity and controls mRNA-levels of aurora B. Cell Cycle 2008, 7, 2710–2719. [Google Scholar] [CrossRef]
- Ding, K.; Ji, J.; Zhang, X.; Huang, B.; Chen, A.; Zhang, D.; Li, X.; Wang, X.; Wang, J. RNA splicing factor USP39 promotes glioma progression by inducing TAZ mRNA maturation. Oncogene 2019, 38, 6414–6428. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Liu, Z.; Zhang, E.; Zhang, P.; Wang, Y.; Hang, J.; Li, Q. USP39 promotes tumorigenesis by stabilizing and deubiquitinating SP1 protein in hepatocellular carcinoma. Cell Signal. 2021, 85, 110068. [Google Scholar] [CrossRef]
- Li, X.; Yuan, J.; Song, C.; Lei, Y.; Xu, J.; Zhang, G.; Wang, W.; Song, G. Deubiquitinase USP39 and E3 ligase TRIM26 balance the level of ZEB1 ubiquitination and thereby determine the progression of hepatocellular carcinoma. Cell Death Differ. 2021, 28, 2315–2332. [Google Scholar] [CrossRef]
- Zhang, Z.; Liu, W.; Bao, X.; Sun, T.; Wang, J.; Li, M.; Liu, C. USP39 facilitates breast cancer cell proliferation through stabilization of FOXM1. Am. J. Cancer Res. 2022, 12, 3644–3661. [Google Scholar]
- Giam, M.; Rancati, G. Aneuploidy and chromosomal instability in cancer: A jackpot to chaos. Cell Div. 2015, 10, 3. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Foreman, O.; Wigle, D.A.; Kosari, F.; Vasmatzis, G.; Salisbury, J.L.; van Deursen, J.; Galardy, P.J. USP44 regulates centrosome positioning to prevent aneuploidy and suppress tumorigenesis. J. Clin. Investig. 2012, 122, 4362–4374. [Google Scholar] [CrossRef] [PubMed]
- Peschiaroli, A.; Skaar, J.R.; Pagano, M.; Melino, G. The ubiquitin-specific protease USP47 is a novel beta-TRCP interactor regulating cell survival. Oncogene 2010, 29, 1384–1393. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Liu, Y.; Xu, X.; Zhang, W.; Yu, T.; Jia, J.; Liu, C. Deubiquitinase USP47/UBP64E Regulates beta-Catenin Ubiquitination and Degradation and Plays a Positive Role in Wnt Signaling. Mol. Cell. Biol. 2015, 35, 3301–3311. [Google Scholar] [CrossRef]
- Bufalieri, F.; Infante, P.; Bernardi, F.; Caimano, M.; Romania, P.; Moretti, M.; Lospinoso Severini, L.; Talbot, J.; Melaiu, O.; Tanori, M.; et al. ERAP1 promotes Hedgehog-dependent tumorigenesis by controlling USP47-mediated degradation of betaTrCP. Nat. Commun. 2019, 10, 3304. [Google Scholar] [CrossRef]
- Bignell, G.R.; Warren, W.; Seal, S.; Takahashi, M.; Rapley, E.; Barfoot, R.; Green, H.; Brown, C.; Biggs, P.J.; Lakhani, S.R.; et al. Identification of the familial cylindromatosis tumour-suppressor gene. Nat. Genet. 2000, 25, 160–165. [Google Scholar] [CrossRef]
- Mathis, B.J.; Lai, Y.; Qu, C.; Janicki, J.S.; Cui, T. CYLD-mediated signaling and diseases. Curr. Drug Targets 2015, 16, 284–294. [Google Scholar] [CrossRef]
- Yang, W.L.; Jin, G.; Li, C.F.; Jeong, Y.S.; Moten, A.; Xu, D.; Feng, Z.; Chen, W.; Cai, Z.; Darnay, B.; et al. Cycles of ubiquitination and deubiquitination critically regulate growth factor-mediated activation of Akt signaling. Sci. Signal 2013, 6, ra3. [Google Scholar] [CrossRef]
- Zhu, M.; Zhao, H.; Liao, J.; Xu, X. HERC2/USP20 coordinates CHK1 activation by modulating CLASPIN stability. Nucleic Acids Res. 2014, 42, 13074–13081. [Google Scholar] [CrossRef]
- Pfoh, R.; Lacdao, I.K.; Saridakis, V. Deubiquitinases and the new therapeutic opportunities offered to cancer. Endocr. Relat. Cancer 2015, 22, T35–T54. [Google Scholar] [CrossRef] [PubMed]
- Fang, R.; Xu, J.; Lin, H.; Xu, X.; Tian, F. The histone demethylase lysine-specific demethylase-1-mediated epigenetic silence of KLF2 contributes to gastric cancer cell proliferation, migration, and invasion. Tumor Biol. 2017, 39, 1010428317698356. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Xu, Y.; Xu, T.; Fan, R.; Jiang, T.; Cao, M.; Shi, L.; Song, J. CircPIP5K1A activates KRT80 and PI3K/AKT pathway to promote gastric cancer development through sponging miR-671-5p. Biomed. Pharmacother. 2020, 126, 109941. [Google Scholar] [CrossRef]
- Malynn, B.A.; Ma, A. A20: A multifunctional tool for regulating immunity and preventing disease. Cell. Immunol. 2019, 340, 103914. [Google Scholar] [CrossRef] [PubMed]
- Bellail, A.C.; Olson, J.J.; Yang, X.; Chen, Z.J.; Hao, C. A20 ubiquitin ligase-mediated polyubiquitination of RIP1 inhibits caspase-8 cleavage and TRAIL-induced apoptosis in glioblastoma. Cancer Discov. 2012, 2, 140–155. [Google Scholar] [CrossRef]
- Zhu, G.; Ye, J.; Huang, Y.; Zheng, W.; Hua, J.; Yang, S.; Zhuang, J.; Wang, J. Receptor-interacting protein-1 promotes the growth and invasion in gastric cancer. Int. J. Oncol. 2016, 48, 2387–2398. [Google Scholar] [CrossRef][Green Version]
- Spataro, V.; Buetti-Dinh, A. POH1/Rpn11/PSMD14: A journey from basic research in fission yeast to a prognostic marker and a druggable target in cancer cells. Br. J. Cancer 2022, 127, 788–799. [Google Scholar] [CrossRef]
- Sun, T.; Liu, Z.; Bi, F.; Yang, Q. Deubiquitinase PSMD14 promotes ovarian cancer progression by decreasing enzymatic activity of PKM2. Mol. Oncol. 2021, 15, 3639–3658. [Google Scholar] [CrossRef]
- Lv, J.; Zhang, S.; Wu, H.; Lu, J.; Lu, Y.; Wang, F.; Zhao, W.; Zhan, P.; Lu, J.; Fang, Q.; et al. Deubiquitinase PSMD14 enhances hepatocellular carcinoma growth and metastasis by stabilizing GRB2. Cancer Lett. 2020, 469, 22–34. [Google Scholar] [CrossRef]
- Zhi, T.; Jiang, K.; Xu, X.; Yu, T.; Zhou, F.; Wang, Y.; Liu, N.; Zhang, J. ECT2/PSMD14/PTTG1 axis promotes the proliferation of glioma through stabilizing E2F1. Neuro-oncology 2019, 21, 462–473. [Google Scholar] [CrossRef]
- Lee, H.J.; Lee, D.M.; Seo, M.J.; Kang, H.C.; Kwon, S.K.; Choi, K.S. PSMD14 Targeting Triggers Paraptosis in Breast Cancer Cells by Inducing Proteasome Inhibition and Ca(2+) Imbalance. Int. J. Mol. Sci. 2022, 23, 2648. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Liu, X.; Zhou, N.; Wu, Q.; Zhou, L.; Li, Q. Gene expression profiling and bioinformatics analysis of gastric carcinoma. Exp. Mol. Pathol. 2014, 96, 361–366. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.O.; Li, C.W.; Xia, W.; Cha, J.H.; Chan, L.C.; Wu, Y.; Chang, S.S.; Lin, W.C.; Hsu, J.M.; Hsu, Y.H.; et al. Deubiquitination and Stabilization of PD-L1 by CSN5. Cancer Cell 2016, 30, 925–939. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Yao, Z.; Wang, J.; Zhang, W.; Yang, Y.; Zhang, Y.; Qu, X.; Zhu, Y.; Zou, J.; Peng, S.; et al. Macrophage-derived CCL5 facilitates immune escape of colorectal cancer cells via the p65/STAT3-CSN5-PD-L1 pathway. Cell Death Differ. 2020, 27, 1765–1781. [Google Scholar] [CrossRef]
- Chen, S.; Liu, Y.; Zhou, H. Advances in the Development Ubiquitin-Specific Peptidase (USP) Inhibitors. Int. J. Mol. Sci. 2021, 22, 4546. [Google Scholar] [CrossRef]
- Prieto-Garcia, C.; Hartmann, O.; Reissland, M.; Braun, F.; Fischer, T.; Walz, S.; Schulein-Volk, C.; Eilers, U.; Ade, C.P.; Calzado, M.A.; et al. Maintaining protein stability of Np63 via USP28 is required by squamous cancer cells. EMBO Mol. Med. 2020, 12, e11101. [Google Scholar] [CrossRef]
- Ma, A.; Tang, M.; Zhang, L.; Wang, B.; Yang, Z.; Liu, Y.; Xu, G.; Wu, L.; Jing, T.; Xu, X.; et al. USP1 inhibition destabilizes KPNA2 and suppresses breast cancer metastasis. Oncogene 2019, 38, 2405–2419. [Google Scholar] [CrossRef]
- Fu, C.; Zhu, X.; Xu, P.; Li, Y. Pharmacological inhibition of USP7 promotes antitumor immunity and contributes to colon cancer therapy. Onco. Targets Ther. 2019, 12, 609–617. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| DUBs | Upstream Regulatory Events | Substrate(s) | Biological Effects | Ref. |
|---|---|---|---|---|
| USPs | ||||
| USP1 | None reported | None reported | USP1 silencing hindered the proliferation, migration, and invasion of GC cells. | [17] |
| None reported | ID2 | USP1 promoted GC metastasis by stabilizing ID2. | [18] | |
| USP2 | None reported | E2F4 | USP2 promoted GC progression by facilitating E2F4/autophagy/zinc homeostasis axis. | [19] |
| USP3 | None reported | None reported | USP3 promoted GC cell proliferation and spreading by regulating cell-cycle-control- and EMT-related molecules. | [20] |
| TGF-β upregulates USP3 expression | SUZ12 | USP3 promoted GC metastasis by stabilizing SUZ12. | [21] | |
| None reported | COL9A3/COL6A5 | USP3 promoted GC cell migration, invasion, and EMT via binding to and deubiquitinating COL9A3 and COL6A5. | [22] | |
| Hsa_circ_0017639 upregulates USP3 expression by sponging miR-224-5p | None reported | Hsa_circ_0017639 participates in GC progression by regulating the miR-224-5p/USP3 axis. | [23] | |
| LncRNA SND1-IT1 functions as a ceRNA to upregulate USP3 expression via absorbing miR-1245b-5p and simultaneously recruiting DDX54 to enhance USP3 mRNA stability | SNAIL1 | Exosomal SND1-IT1 from GC cells upregulated USP3 expression, thus mediating SNAIL1 stabilization and accelerating the migration and invasion of gastric mucosa cells. | [24] | |
| USP7 | None reported | USP11, PPM1G, DHX40, DDX24 and TRIP12 | Unclear | [25] |
| None reported | PD-L1 | USP7 served as an upstream DUB of PD-L1; USP7 abrogation hindered GC cell growth by downregulating PD-L1-mediated immunosuppression and enhanced cell cycle arrest simultaneously by stabilizing p53. | [26] | |
| Cisplatin and paclitaxel promoted USP7 expression in CAFs | hnRNPA1 | Cisplatin and paclitaxel promoted the secretion of miR-522 from CAFs by activating USP7/hnRNPA1 axis, resulting in ferroptosis suppression, and acquired chemoresistance by inhibiting ALOX15 expression and lipid-ROS accumulation in GC cells. | [27] | |
| USP9X | None reported | None reported | USP9X was overexpressed and predicted poorer survival in GC. | [28] |
| Hsa_circ_0008434 enhances USP9X expression by sponging miR-6838-5p | None reported | Hsa_circ_0008434 promoted GC proliferation, invasion, and migration by regulating miR-6838-5p/USP9X axis. | [29] | |
| None reported | YAP | The LINC01433-YAP feedback loop promoted GC cell proliferation, migration, invasion, and chemotherapy resistance. LINC01433 increased the stability but decreased the phosphorylation of YAP by enhancing its interaction with USP9X and attenuating its interaction with LATS1, respectively. | [30] | |
| USP10 | None reported | None reported | USP10 was an independent prognostic marker for patients with GC. | [31] |
| 3-Deazaneplanocin A treatment upregulates USP10 expression by reducing EZH2 binding on its promoter | p53 | Stabilization of p53 by USP10 seemed to be correlated with the sensitivity of GC cells to 3-Deazaneplanocin A. | [32] | |
| CircCOL1A2 upregulates USP10 expression by sponging miR-1286 | RFC2 | CircCOL1A2 sponges miR-1286 to promote GC cell migration and invasion by increasing USP10 level to stabilize RFC2. | [33] | |
| USP11 | None reported | None reported | USP11 overexpression promoted proliferation and migration and alleviated paclitaxel’s toxicity in GC cells by inhibiting RhoA and Ras signaling. | [34] |
| USP13 | None reported | None reported | High expression of USP13 predicted poor prognosis in GC. | [35] |
| None reported | Snail | USP13 promoted the EMT and metastasis of GC cells by stabilizing Snail. | [36] | |
| USP14 | miR-320a inhibits USP14 expression by targeting its 3′-UTR | Vimentin | USP14-mediated deubiquitination of vimentin enhanced the aggressiveness of GC cells, and miR-320a served as a tumor suppressor by inhibiting both USP14 and vimentin. | [37] |
| None reported | None reported | USP14 silencing sensitized GC cells to cisplatin by impeding Akt/ERK signaling pathways. | [38] | |
| YTHDF1 enhanced USP14 protein translation in a m6A-dependent manner | None reported | YTHDF1 promoted GC progression and metastasis by promoting USP14 protein translation in an m6A-dependent manner. | [39] | |
| USP15 | None reported | None reported | USP15 overexpression inhibited GC cell proliferation, migration, and invasion. | [40] |
| None reported | None reported | USP15 promoted GC progression through the Wnt/β-catenin signaling pathway. | [41] | |
| USP15 is potential regulated by LINC00205/miR-26a axis | None reported | Unclear | [42] | |
| USP20 | None reported | None reported | USP20 inhibited GC cell growth and G1/S cell cycle transition by regulating Claspin. | [43] |
| USP21 | None reported | GATA3 | USP21 upregulated MAPK1 expression by stabilizing GATA3 to promote GC cell growth and stemness. | [44] |
| USP22 | None reported | None reported | Coordinate expression of USP22 and BMI1 correlated with GC progression and treatment failure. | [45] |
| None reported | None reported | High expression of USP22 correlated with GC progression and has synergistic effects with c-Myc. | [46] | |
| None reported | BMI1 | USP22 contributed to gastric CSC stemness maintenance and GC progression by stabilizing BMI1. | [47] | |
| None reported | None reported | USP22 siRNA-loaded nanoliposomes decorated with CD44 antibodies selectively target and eliminate CD44+ GC stem cells. | [48] | |
| None reported | None reported | Positive co-expression of USP22 and HSP90 might be more effective in predicting prognosis of GC. | [49] | |
| None reported | None reported | USP22 promotes GC progression and metastasis through c-Myc/NAMPT/SIRT1-dependent FOXO1 and YAP signaling. | [50] | |
| POU2F1 upregulates USP22 expression by suppressing the expression of miR-4490 | None reported | POU2F1-miR-4490-USP22 signaling axis plays a significant role in GC development and progression. | [51] | |
| None reported | None reported | USP22 overexpression in GC induces the upregulation of SOS1 and activation of the RAS/ERK and PI3K/AKT pathways. | [52] | |
| USP28 | None reported | None reported | USP28 promoted cell proliferation and metastasis of GC cells by regulating LSD1. | [53] |
| USP29 | TGFβ, TNFα, and hypoxia induced the transcription of USP29 | Snail | USP29 cooperated with SCP1 to prevent Snail degradation and further promoted GC cell metastasis. | [54] |
| USP32 | None reported | SMAD2 | USP32 is involved in GC development and chemoresistance through the upregulation of SMAD2. | [55] |
| USP33 | None reported | None reported | USP33 overexpression inhibited GC cell proliferation, migration, and invasion. | [56] |
| None reported | ROBO1 | USP33 contributed to SLIT2-ROBO1 axis in inhibiting GC cell migration and EMT process. | [57] | |
| DUB1 | None reported | TAZ | DUB1 stabilized TAZ protein and promoted GC progression via the Hippo/TAZ axis. | [58] |
| USP37 | PLAGL2 activated USP37 transcription | Snail | USP37, which is transcriptionally activated by PLAGL2, deubiquitinates and stabilizes Snail1 to promote the proliferation, EMT, and metastasis of GC cells. | [59] |
| USP39 | None reported | None reported | USP39 silencing inhibited the growth of GC cells via PARP activation. | [60] |
| miR-133a inhibited USP39 expression by targeting its 3′-UTR | None reported | miR-133a suppressed GC proliferation by regulating USP39. | [61] | |
| USP42 | None reported | None reported | USP42 depression inhibited GC cell proliferation and invasion. | [62] |
| USP44 | None reported | None reported | USP44 overexpression resulted in DNA aneuploidy. | [63] |
| CircFOXO3 upregulates USP44 expression by sponging miR-143-3p | None reported | CircFOXO3 promoted GC cells proliferation and migration by increasing USP44 expression through targeting miR-143-3p. | [64] | |
| USP47 | miR-204-5p inhibited USP47 expression via binding its 3′-UTR | None reported | MiR-204-5p functioned as a tumor suppressor in GC by suppressing USP47 and RAB22A. | [65] |
| None reported | None reported | USP47 regulates NF-κB activity by stabilizing β-TrCP and contributes to chemoresistance of GC cells. | [66] | |
| USP49 | USP49 was transcriptionally activated by the YAP1/TEAD4 complex | YAP1 | USP49 and YAP1 form a positive feedback loop to promote malignant progression of GC. | [67] |
| CYLD | miR-362 inhibited CYLD expression via targeting its 3′-UTR | None reported | Upregulated miR-362 promoted GC cell proliferation and cisplatin resistance by repressing CYLD and activating NF-κB signaling. | [68] |
| miR-500 inhibited CYLD expression via targeting its 3′-UTR | None reported | Upregulated miR-500 promoted GC cell proliferation and cisplatin resistance by repressing CYLD, OTUD7B, and TAX1BP1 and activating NF-κB signaling. | [69] | |
| miR-130b inhibited CYLD expression via targeting its 3′-UTR | None reported | Upregulated miR-130b promoted GC cell proliferation and inhibited apoptosis by repressing CYLD. | [70] | |
| miR-20a negatively regulated CYLD expression by targeting its 3′ UTR | None reported | Upregulated miR-20a augmented the resistance of GC cells to cisplatin by inhibiting CYLD and activating NF-κB signaling and its downstream targets. | [71] | |
| miR-425-5p negatively regulated CYLD expression by targeting its 3′ UTR | None reported | Upregulated miR-425-5p may promote GC cell migration and invasion by repressing CYLD. | [72] | |
| CYLD expression was inversely correlated with hypermethylation of its promoter, which could be induced by some infectious agents | None reported | Decreased CYLD level may be associated with gender, patient’s age, high grade, and no lymph-node metastasis in GC patients. | [73] | |
| LncRNA CRAL functions as a ceRNA to upregulate CYLD expression via absorbing miR-505 | None reported | LncRNA CRAL improved cisplatin resistance via the miR-505/CYLD/AKT axis in GC cells. | [74] | |
| miR-454 inhibited CYLD expression via targeting its 3′-UTR | None reported | miR-454 supported survival and induced oxaliplatin resistance in GC cells by repressing CYLD. | [75] | |
| Exosomal miR-588 from M2 polarized macrophages inhibited CYLD expression via targeting its 3′-UTR | None reported | Exosomal miR-588 contributed to cisplatin GC cell resistance by repressing CYLD. | [76] | |
| ZNF333 decreased CYLD level by binding to its promoter | None reported | ALKBH5 promoted the process of bile-acid-induced gastric intestinal metaplasia by ZNF333/CYLD/CDX2 pathway. | [77] | |
| UCHs | ||||
| UCHL1 | UCHL1 may be epigenetically inactivated via promoter methylation | None reported | UCHL1 methylation was correlated with poor clinical outcome in GC patients. | [78,79,80,81,82] |
| None reported | None reported | UCHL1 promoted GC cell proliferation and metastasis by activating the Akt and ERK1/2 pathways. | [83] | |
| None reported | None reported | UCHLI silencing inhibited GC cell proliferation and metastasis. | [84] | |
| UCHL3 | None reported | None reported | UCHL3 stimulated GC metastasis by upregulating IGF2. | [85] |
| UCHL5 | None reported | None reported | A certain subgroup of GC patients with high expression of UCHL5 had better prognosis. | [86] |
| None reported | NFRKB | LncRNA DRAIC inhibited GC proliferation and metastasis by mediating ubiquitination degradation of NFRKB by interfering with its combination with UCHL5. | [87] | |
| BAP1 | None reported | None reported | BAP1 downregulation predicts unfavorable survival in GC. | [88] |
| OTUs | ||||
| OTUB1 | None reported | None reported | OTUB1 enhanced tumor invasiveness and predicted a poor prognosis in GC. | [89] |
| OTUB2 | None reported | KRT80 | OTUB2 enhanced KRT80 stability via deubiquitination and promoted GC proliferation. | [90] |
| None reported | KDM1A | OTUB2 promoted GC tumorigenesis by enhancing KDM1A-mediated stem cell-like properties. | [91] | |
| A20 | A20 expression was inversely correlated with methylation at specific CpG sites in its intronic region | None reported | Increased A20 levels were associated with poor clinical outcomes. | [92] |
| None reported | None reported | A20 depletion inhibited the capacity of proliferation, migration, and invasion of GC cells. | [93] | |
| miR-200a inhibited A20 expression by targeting its 3′-UTR | RIP1 | miR-200a prevented RIP1 polyubiquitination and enhanced TRAIL sensitivity by targeting A20 in GC cells. | [94] | |
| H. pylori-induced NF-κB activation elevated A20 expression | Procaspase-8 | Increased A20 expression by USP48 inhibited K63-linked ubiquitinylation of procaspase-8, restricting caspase-8 activity and apoptosis in GC cells. | [95,96,97,98] | |
| H. pylori infection decreased A20 expression by upregulating miR-29a-3p | None reported | A20 silencing promoted gastric epithelial cell migration. | [99] | |
| JAMMs | ||||
| PSMD14 | None reported | PTBP1 | PSMD14 stabilized PTBP1 to promote GC cell proliferation and invasion. | [100] |
| CSN5 | None reported | RUNX3 | CSN5 facilitates nuclear export and degradation of RUNX3. | [101] |
| None reported | None reported | CSN5 knockdown inhibited proliferation and promoted apoptosis of GC cells by regulating p53-related apoptotic pathways. | [102] | |
| None reported | p14ARF | CSN5 potentiated GC progression by decreasing p14ARF expression through non-ubiquitin pathway. | [103] | |
| The activity of CSN5 was enhanced by IKKβ, and IKKβ expression was inhibited by DAPK1 | PD-L1 | Overexpression of DAPK1 enhanced NK cell killing and suppressed tumor immune evasion in GC cells by inhibiting the IKKβ/CSN5/PD-L1 axis. | [104] | |
| BRCC3 | LncRNA TMPO-AS1 functions as a ceRNA to upregulate BRCC3 expression via absorbing miR-126-5p | None reported | TMPO-AS1 accelerated GC cell proliferation, migration, and angiogenesis by regulating miR-126-5p/BRCC3 axis and activating PI3K/Akt/mTOR signaling. | [105] |
| MJDs | ||||
| Ataxin-3 | None reported | None reported | Decreased Ataxin-3 expression correlated with clinicopathologic features of GC. | [106] |
| Compound | Structure | Affected DUB | IC50 (μM) | Effects | Ref. |
|---|---|---|---|---|---|
| Emetine |  | USP2 | Not applicable | Emetine inhibited autophagy and GC progression by blocking USP2-E2F4 interaction | [19] |
| Almac4 |  | USP7 | 0.0015 ± 0.001 | Almac4 treatment enhanced the sensitivity of GC cells to T-cell killing and inhibited GC cell proliferation by elevating p53 | [26,107] |
| C9 |  | USP7 | 4.86 | C9 suppressed proliferation of MGC-803 GC cells by decreasing MDM2 expression and thus increasing p53 and p21 levels | [108] |
| IU1 |  | USP14 | 4.7 ± 0.7 | IU1 restrained cell growth and tumor-promoting effects induced by YTHDF1 in GC cells | [39,109] |
| Compound 19 |  | USP28 | 1.10 ± 0.02 | Compound 19 bound to USP28 and inhibited malignant behaviors of GC cells by downregulating LSD1 and c-Myc | [110] |
| Lanatoside C |  | USP28 | Not applicable | Lanatoside C suppressed cell proliferation and induced apoptosis by partially attenuating the binding between USP28 and c-Myc in GC cells | [111] |
| Galangin |  | UCHL1 | Not applicable | Galangin-induced growth inhibitory effect in SNU-484 GC cells was accompanied by UCHL1 upregulation | [112] |
| 3,3′-Diindolylmethane |  | BAP1 | Not applicable | 3,3′-Diindolylmethane induced ferroptosis in BGC-823 GC cells by BAP1 upregulation | [113] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
An, T.; Lu, Y.; Gong, Z.; Wang, Y.; Su, C.; Tang, G.; Hou, J. Research Progress for Targeting Deubiquitinases in Gastric Cancers. Cancers 2022, 14, 5831. https://doi.org/10.3390/cancers14235831
An T, Lu Y, Gong Z, Wang Y, Su C, Tang G, Hou J. Research Progress for Targeting Deubiquitinases in Gastric Cancers. Cancers. 2022; 14(23):5831. https://doi.org/10.3390/cancers14235831
Chicago/Turabian StyleAn, Tao, Yanting Lu, Zhaoqi Gong, Yongtao Wang, Chen Su, Guimei Tang, and Jingjing Hou. 2022. "Research Progress for Targeting Deubiquitinases in Gastric Cancers" Cancers 14, no. 23: 5831. https://doi.org/10.3390/cancers14235831
APA StyleAn, T., Lu, Y., Gong, Z., Wang, Y., Su, C., Tang, G., & Hou, J. (2022). Research Progress for Targeting Deubiquitinases in Gastric Cancers. Cancers, 14(23), 5831. https://doi.org/10.3390/cancers14235831