
Circulating Exhausted PD-1+CD39+ Helper CD4 T Cells Are Tumor-Antigen-Specific and Predict Response to PD-1/PD-L1 Axis Blockade


 ,
, {kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Patient and Healthy Donor Samples
2.2. CD4 T Cells’ Purification and Phenotyping
2.3. Sorting and Functional Assessment of CD4 T-Cell Subpopulations
2.4. Acute and Chronic In Vitro Stimulation
2.5. Statistical Analyses
3. Results
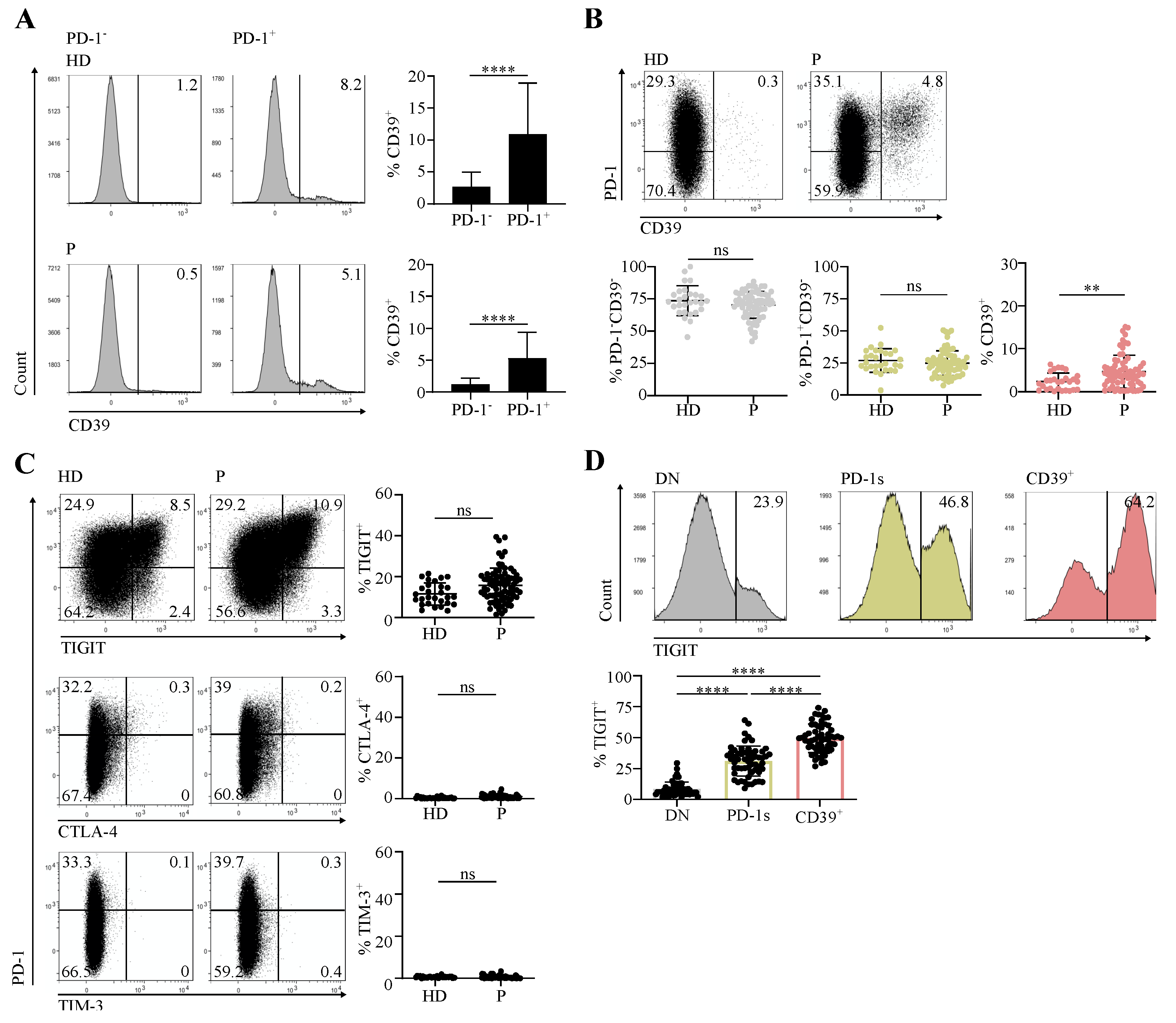
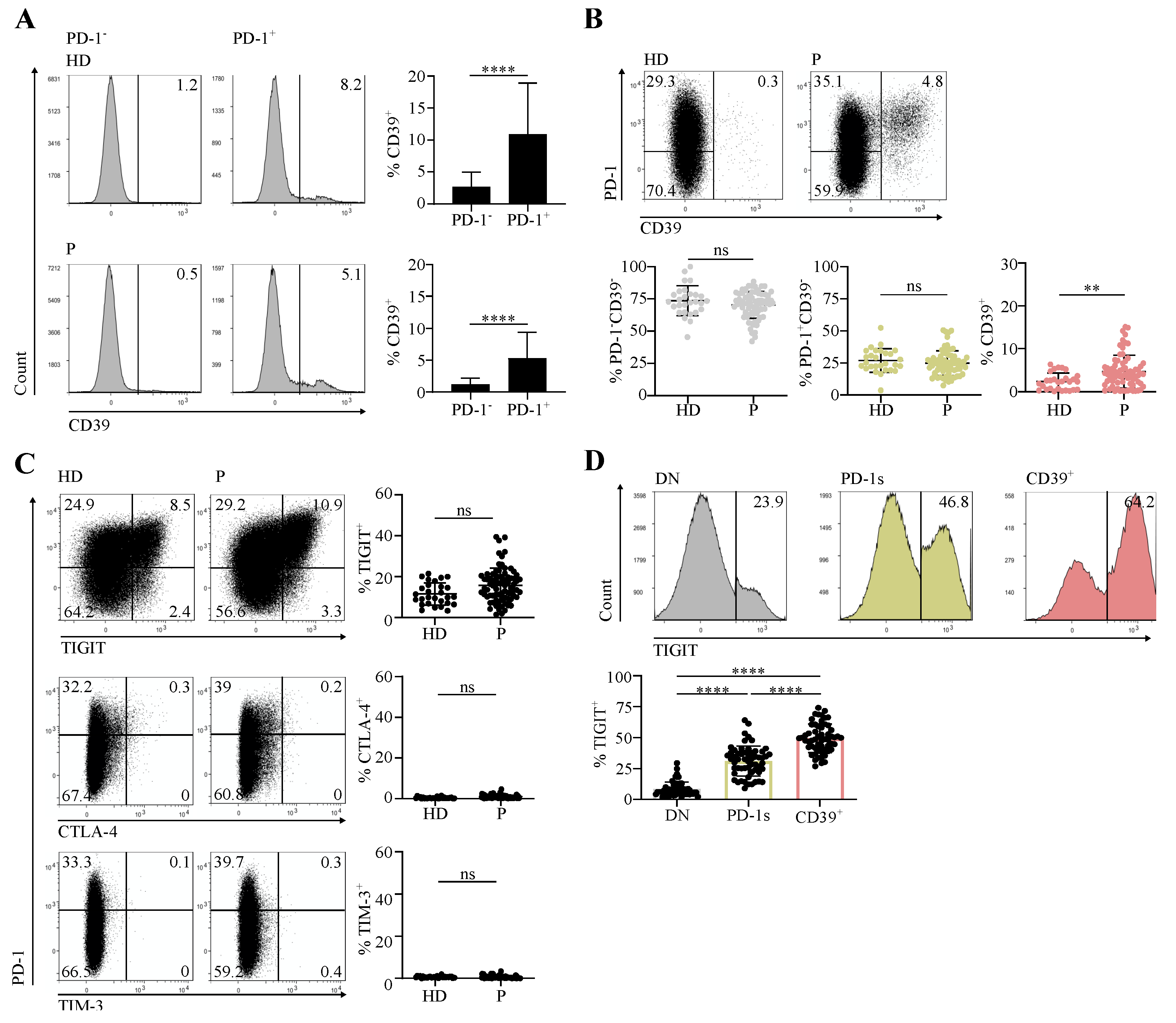
3.1. Circulating CD4 T Cells Contain a PD-1+CD39+ Population Found at Increased Proportions in Cancer Patients in Comparison with Healthy Individuals
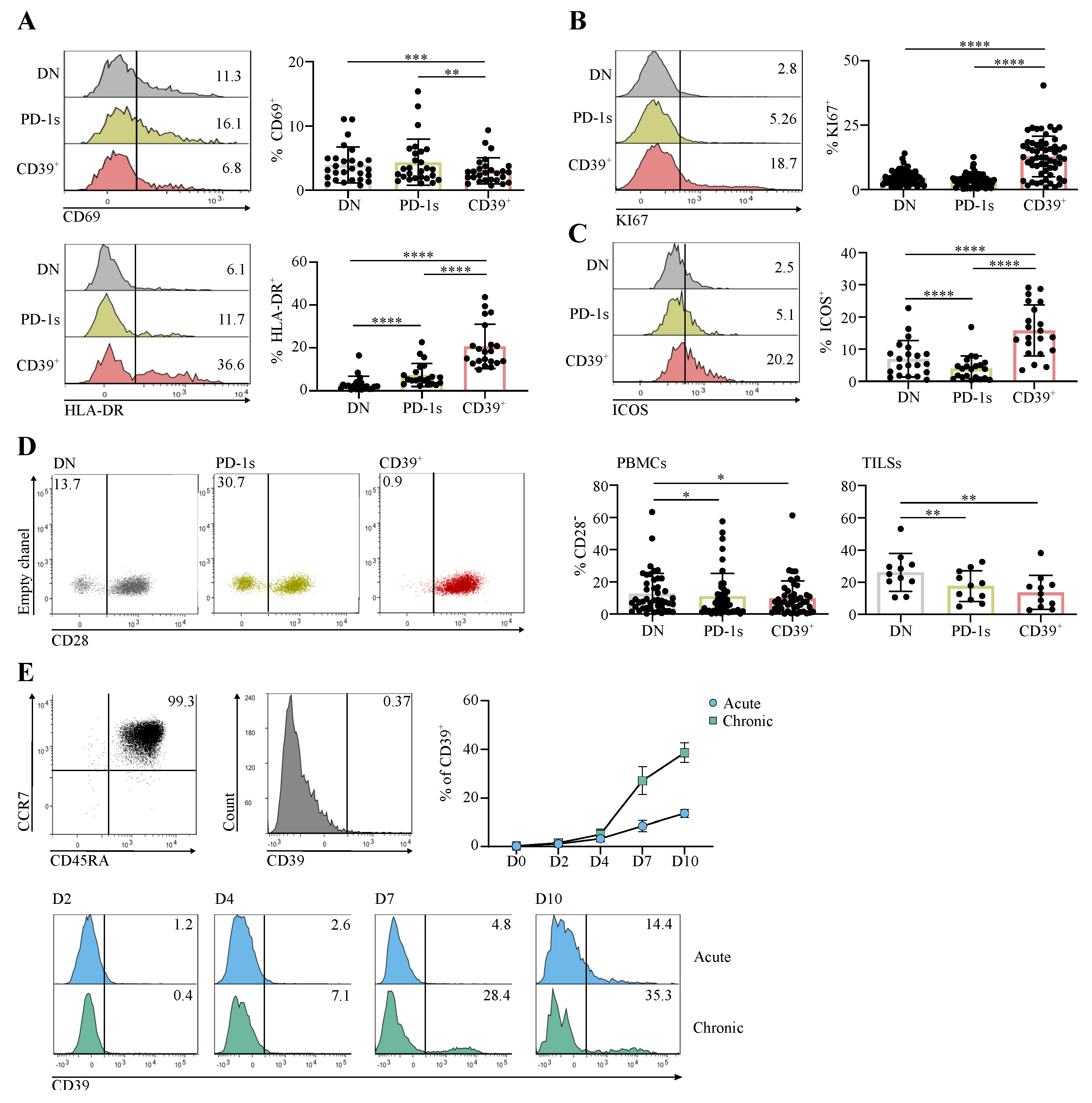
3.2. The Circulating PD-1+CD39+ CD4 T-Cell Subpopulation Is Enriched in Activated and Proliferating Cells That Were at an Early Differentiation Stage
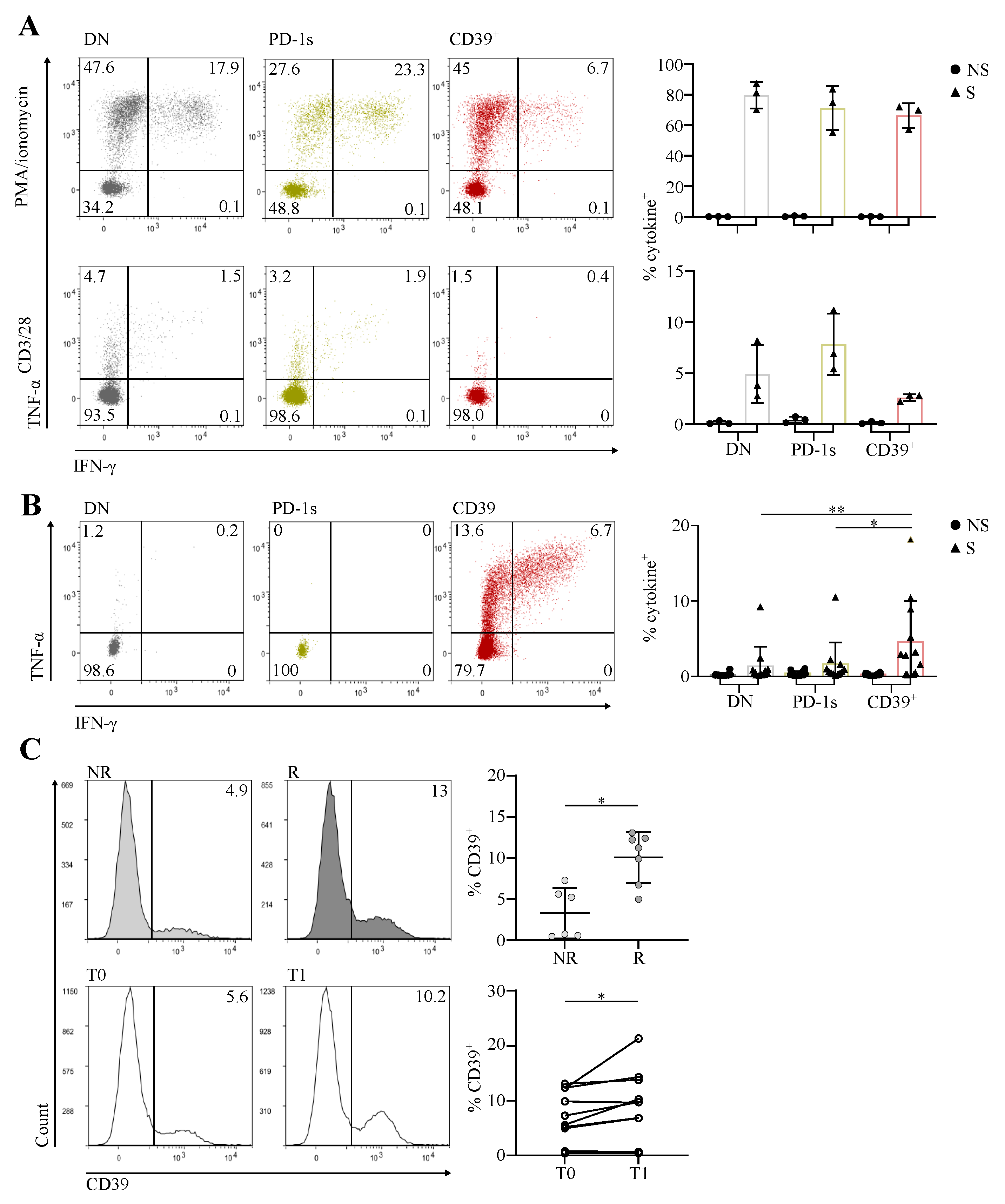
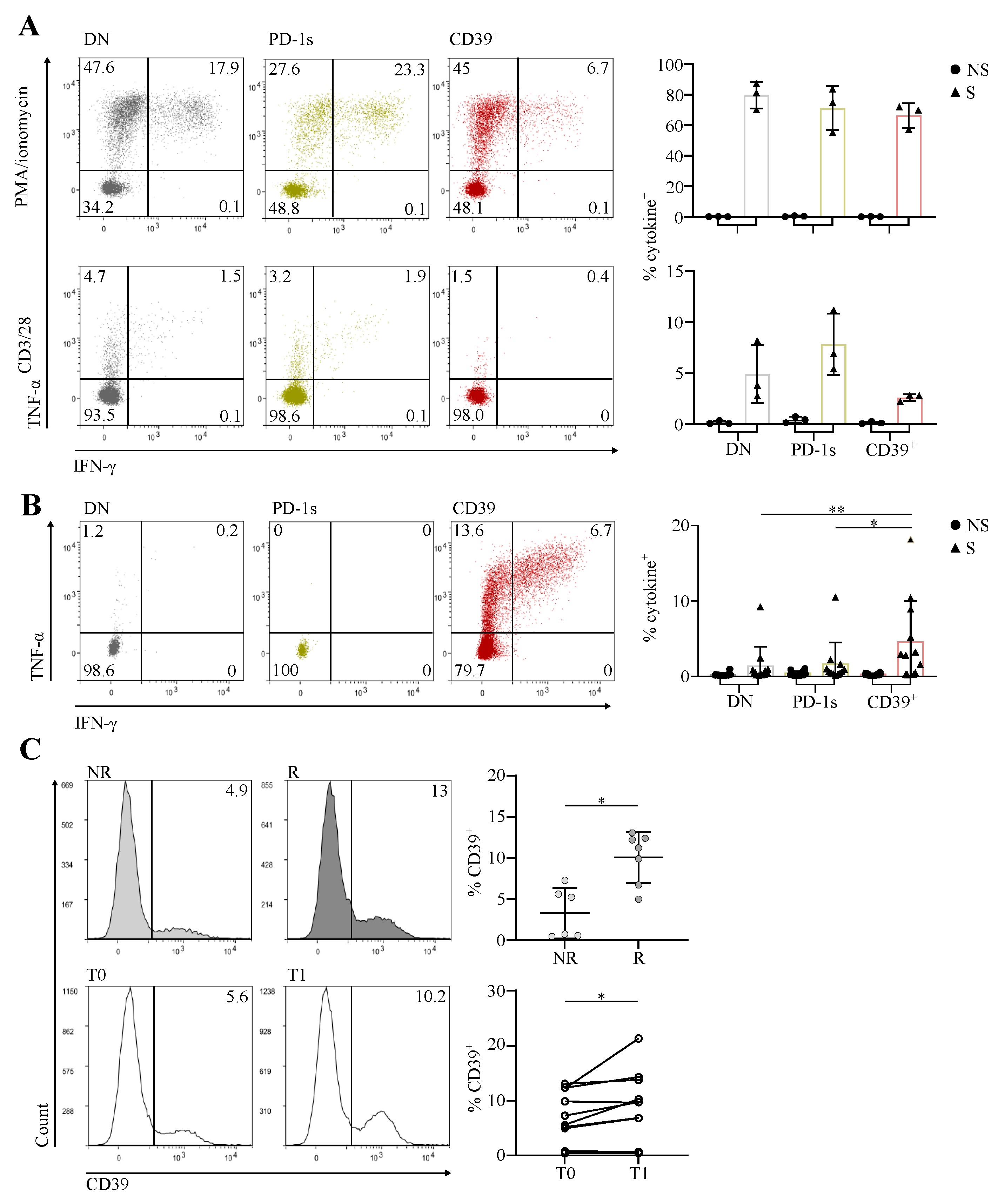
3.3. The Circulating PD-1+CD39+ Memory CD4 T-Cell Subpopulation Contains Tumor-Ag-Specific Exhausted Cells and Predicts the Response to PD-1/PD-L1 Axis Blockade
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sharma, P.; Siddiqui, B.A.; Anandhan, S.; Yadav, S.S.; Subudhi, S.K.; Gao, J.; Goswami, S.; Allison, J.P. The Next Decade of Immune Checkpoint Therapy. Cancer Discov. 2021, 11, 838–857. [Google Scholar] [CrossRef]
- Topalian, S.L.; Taube, J.M.; Anders, R.A.; Pardoll, D.M. Mechanism-Driven Biomarkers to Guide Immune Checkpoint Blockade in Cancer Therapy. Nat. Rev. Cancer 2016, 16, 275–287. [Google Scholar] [CrossRef]
- Hofman, P.; Heeke, S.; Alix-Panabières, C.; Pantel, K. Liquid Biopsy in the Era of Immuno-Oncology: Is It Ready for Prime-Time Use for Cancer Patients? Ann. Oncol. 2019, 30, 1448–1459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balança, C.-C.; Scarlata, C.-M.; Michelas, M.; Devaud, C.; Sarradin, V.; Franchet, C.; Martinez Gomez, C.; Gomez-Roca, C.; Tosolini, M.; Heaugwane, D.; et al. Dual Relief of T-Lymphocyte Proliferation and Effector Function Underlies Response to PD-1 Blockade in Epithelial Malignancies. Cancer Immunol. Res. 2020, 8, 869–882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thommen, D.S.; Koelzer, V.H.; Herzig, P.; Roller, A.; Trefny, M.; Dimeloe, S.; Kiialainen, A.; Hanhart, J.; Schill, C.; Hess, C.; et al. A Transcriptionally and Functionally Distinct PD-1+ CD8+ T Cell Pool with Predictive Potential in Non-Small-Cell Lung Cancer Treated with PD-1 Blockade. Nat. Med. 2018, 24, 994–1004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, T.D.; Madireddi, S.; de Almeida, P.E.; Banchereau, R.; Chen, Y.-J.J.; Chitre, A.S.; Chiang, E.Y.; Iftikhar, H.; O’Gorman, W.E.; Au-Yeung, A.; et al. Peripheral T Cell Expansion Predicts Tumour Infiltration and Clinical Response. Nature 2020, 579, 274–278. [Google Scholar] [CrossRef] [PubMed]
- Kamphorst, A.O.; Pillai, R.N.; Yang, S.; Nasti, T.H.; Akondy, R.S.; Wieland, A.; Sica, G.L.; Yu, K.; Koenig, L.; Patel, N.T.; et al. Proliferation of PD-1+ CD8 T Cells in Peripheral Blood after PD-1–Targeted Therapy in Lung Cancer Patients. Proc. Natl. Acad. Sci. USA 2017, 114, 4993–4998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balança, C.-C.; Salvioni, A.; Scarlata, C.-M.; Michelas, M.; Martinez-Gomez, C.; Gomez-Roca, C.; Sarradin, V.; Tosolini, M.; Valle, C.; Pont, F.; et al. PD-1 Blockade Restores Helper Activity of Tumor-Infiltrating, Exhausted PD-1hiCD39+ CD4 T Cells. JCI Insight 2021, 6, e142513. [Google Scholar] [CrossRef] [PubMed]
- Kortekaas, K.E.; Santegoets, S.J.; Sturm, G.; Ehsan, I.; van Egmond, S.L.; Finotello, F.; Trajanoski, Z.; Welters, M.J.P.; van Poelgeest, M.I.E.; van der Burg, S.H. CD39 Identifies the CD4 + Tumor-Specific T-Cell Population in Human Cancer. Cancer Immunol. Res. 2020, 8, 1311–1321. [Google Scholar] [CrossRef]
- Chaurio, R.A.; Anadon, C.M.; Lee Costich, T.; Payne, K.K.; Biswas, S.; Harro, C.M.; Moran, C.; Ortiz, A.C.; Cortina, C.; Rigolizzo, K.E.; et al. TGF-β-Mediated Silencing of Genomic Organizer SATB1 Promotes Tfh Cell Differentiation and Formation of Intra-Tumoral Tertiary Lymphoid Structures. Immunity 2022, 55, 115–128.e9. [Google Scholar] [CrossRef] [PubMed]
- Cohen, M.; Giladi, A.; Barboy, O.; Hamon, P.; Li, B.; Zada, M.; Gurevich-Shapiro, A.; Beccaria, C.G.; David, E.; Maier, B.B.; et al. The Interaction of CD4+ Helper T Cells with Dendritic Cells Shapes the Tumor Microenvironment and Immune Checkpoint Blockade Response. Nat. Cancer 2022, 3, 303–317. [Google Scholar] [CrossRef]
- Li, J.-P.; Wu, C.-Y.; Chen, M.-Y.; Liu, S.-X.; Yan, S.-M.; Kang, Y.-F.; Sun, C.; Grandis, J.R.; Zeng, M.-S.; Zhong, Q. PD-1 + CXCR5 − CD4 + Th-CXCL13 Cell Subset Drives B Cells into Tertiary Lymphoid Structures of Nasopharyngeal Carcinoma. J. Immunother Cancer 2021, 9, e002101. [Google Scholar] [CrossRef]
- Veatch, J.R.; Lee, S.M.; Shasha, C.; Singhi, N.; Szeto, J.L.; Moshiri, A.S.; Kim, T.S.; Smythe, K.; Kong, P.; Fitzgibbon, M.; et al. Neoantigen-Specific CD4+ T Cells in Human Melanoma Have Diverse Differentiation States and Correlate with CD8+ T Cell, Macrophage, and B Cell Function. Cancer Cell 2022, 40, 393–409.e9. [Google Scholar] [CrossRef]
- Zheng, C.; Fass, J.N.; Shih, Y.-P.; Gunderson, A.J.; Sanjuan Silva, N.; Huang, H.; Bernard, B.M.; Rajamanickam, V.; Slagel, J.; Bifulco, C.B.; et al. Transcriptomic Profiles of Neoantigen-Reactive T Cells in Human Gastrointestinal Cancers. Cancer Cell 2022, 40, 410–423.e7. [Google Scholar] [CrossRef] [PubMed]
- Alspach, E.; Lussier, D.M.; Miceli, A.P.; Kizhvatov, I.; DuPage, M.; Luoma, A.M.; Meng, W.; Lichti, C.F.; Esaulova, E.; Vomund, A.N.; et al. MHC-II Neoantigens Shape Tumour Immunity and Response to Immunotherapy. Nature 2019, 574, 696–701. [Google Scholar] [CrossRef] [PubMed]
- Borst, J.; Ahrends, T.; Bąbała, N.; Melief, C.J.M.; Kastenmüller, W. CD4 + T Cell Help in Cancer Immunology and Immunotherapy. Nat. Rev. Immunol. 2018, 18, 635–647. [Google Scholar] [CrossRef]
- Perrot, I.; Michaud, H.-A.; Giraudon-Paoli, M.; Augier, S.; Docquier, A.; Gros, L.; Courtois, R.; Déjou, C.; Jecko, D.; Becquart, O.; et al. Blocking Antibodies Targeting the CD39/CD73 Immunosuppressive Pathway Unleash Immune Responses in Combination Cancer Therapies. Cell Rep. 2019, 27, 2411–2425.e9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Canale, F.P.; Ramello, M.C.; Núñez, N.; Furlan, C.L.A.; Bossio, S.N.; Serrán, M.G.; Boari, J.T.; del Castillo, A.; Ledesma, M.; Sedlik, C.; et al. CD39 Expression Defines Cell Exhaustion in Tumor-Infiltrating CD8+ T Cells. Cancer Res. 2018, 78, 115–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duhen, T.; Duhen, R.; Montler, R.; Moses, J.; Moudgil, T.; de Miranda, N.F.; Goodall, C.P.; Blair, T.C.; Fox, B.A.; McDermott, J.E.; et al. Co-Expression of CD39 and CD103 Identifies Tumor-Reactive CD8 T Cells in Human Solid Tumors. Nat. Commun. 2018, 9, 2724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eberhardt, C.S.; Kissick, H.T.; Patel, M.R.; Cardenas, M.A.; Prokhnevska, N.; Obeng, R.C.; Nasti, T.H.; Griffith, C.C.; Im, S.J.; Wang, X.; et al. Functional HPV-Specific PD-1+ Stem-like CD8 T Cells in Head and Neck Cancer. Nature 2021, 597, 279–284. [Google Scholar] [CrossRef] [PubMed]
- Reddy, M.; Eirikis, E.; Davis, C.; Davis, H.M.; Prabhakar, U. Comparative Analysis of Lymphocyte Activation Marker Expression and Cytokine Secretion Profile in Stimulated Human Peripheral Blood Mononuclear Cell Cultures: An In Vitro Model to Monitor Cellular Immune Function. J. Immunol. Methods 2004, 293, 127–142. [Google Scholar] [CrossRef] [PubMed]
- Simpson, T.R.; Quezada, S.A.; Allison, J.P. Regulation of CD4 T Cell Activation and Effector Function by Inducible Costimulator (ICOS). Curr. Opin. Immunol. 2010, 22, 326–332. [Google Scholar] [CrossRef]
- Wherry, E.J. T Cell Exhaustion. Nat. Immunol. 2011, 12, 492–499. [Google Scholar] [CrossRef] [PubMed]
- Budimir, N.; Thomas, G.D.; Dolina, J.S.; Salek-Ardakani, S. Reversing T-Cell Exhaustion in Cancer: Lessons Learned from PD-1/PD-L1 Immune Checkpoint Blockade. Cancer Immunol. Res. 2022, 10, 146–153. [Google Scholar] [CrossRef]
- Ayyoub, M.; Hesdorffer, C.S.; Montes, M.; Merlo, A.; Speiser, D.; Rimoldi, D.; Cerottini, J.-C.; Ritter, G.; Scanlan, M.; Old, L.J.; et al. An Immunodominant SSX-2-Derived Epitope Recognized by CD4+ T Cells in Association with HLA-DR. J. Clin. Investig. 2004, 113, 1225–1233. [Google Scholar] [CrossRef]
- Ayyoub, M.; Pignon, P.; Classe, J.-M.; Odunsi, K.; Valmori, D. CD4+ T Effectors Specific for the Tumor Antigen NY-ESO-1 Are Highly Enriched at Ovarian Cancer Sites and Coexist with, but Are Distinct from, Tumor-Associated Treg. Cancer Immunol. Res. 2013, 1, 303–308. [Google Scholar] [CrossRef] [Green Version]
- Djenidi, F.; Adam, J.; Goubar, A.; Durgeau, A.; Meurice, G.; de Montpréville, V.; Validire, P.; Besse, B.; Mami-Chouaib, F. CD8+CD103+ Tumor-Infiltrating Lymphocytes Are Tumor-Specific Tissue-Resident Memory T Cells and a Prognostic Factor for Survival in Lung Cancer Patients. J. Immunol. 2015, 194, 3475–3486. [Google Scholar] [CrossRef] [Green Version]
- Mami-Chouaib, F.; Blanc, C.; Corgnac, S.; Hans, S.; Malenica, I.; Granier, C.; Tihy, I.; Tartour, E. Resident Memory T Cells, Critical Components in Tumor Immunology. J. Immunother. Cancer 2018, 6, 87. [Google Scholar] [CrossRef]
- Klicznik, M.M.; Morawski, P.A.; Höllbacher, B.; Varkhande, S.R.; Motley, S.J.; Kuri-Cervantes, L.; Goodwin, E.; Rosenblum, M.D.; Long, S.A.; Brachtl, G.; et al. Human CD4 + CD103 + Cutaneous Resident Memory T Cells Are Found in the Circulation of Healthy Individuals. Sci. Immunol. 2019, 4, eaav8995. [Google Scholar] [CrossRef] [PubMed]
- Ugolini, A.; Nuti, M. CD137+ T-Cells: Protagonists of the Immunotherapy Revolution. Cancers 2021, 13, 456. [Google Scholar] [CrossRef] [PubMed]
- Jacquelot, N.; Roberti, M.P.; Enot, D.P.; Rusakiewicz, S.; Ternès, N.; Jegou, S.; Woods, D.M.; Sodré, A.L.; Hansen, M.; Meirow, Y.; et al. Predictors of Responses to Immune Checkpoint Blockade in Advanced Melanoma. Nat. Commun. 2017, 8, 592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zizzari, I.G.; Di Filippo, A.; Botticelli, A.; Strigari, L.; Pernazza, A.; Rullo, E.; Pignataro, M.G.; Ugolini, A.; Scirocchi, F.; Di Pietro, F.R.; et al. Circulating CD137+ T Cells Correlate with Improved Response to Anti-PD1 Immunotherapy in Patients with Cancer. Clin. Cancer Res. 2022, 28, 1027–1037. [Google Scholar] [CrossRef]
- Gupta, P.K.; Godec, J.; Wolski, D.; Adland, E.; Yates, K.; Pauken, K.E.; Cosgrove, C.; Ledderose, C.; Junger, W.G.; Robson, S.C.; et al. CD39 Expression Identifies Terminally Exhausted CD8+ T Cells. PLoS Pathog. 2015, 11, e1005177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martinez-Gomez, C.; Michelas, M.; Scarlata, C.-M.; Salvioni, A.; Gomez-Roca, C.; Sarradin, V.; Lauzéral-Vizcaino, F.; Féliu, V.; Dupret-Bories, A.; Ferron, G.; et al. Circulating Exhausted PD-1+CD39+ Helper CD4 T Cells Are Tumor-Antigen-Specific and Predict Response to PD-1/PD-L1 Axis Blockade. Cancers 2022, 14, 3679. https://doi.org/10.3390/cancers14153679
Martinez-Gomez C, Michelas M, Scarlata C-M, Salvioni A, Gomez-Roca C, Sarradin V, Lauzéral-Vizcaino F, Féliu V, Dupret-Bories A, Ferron G, et al. Circulating Exhausted PD-1+CD39+ Helper CD4 T Cells Are Tumor-Antigen-Specific and Predict Response to PD-1/PD-L1 Axis Blockade. Cancers. 2022; 14(15):3679. https://doi.org/10.3390/cancers14153679
Chicago/Turabian StyleMartinez-Gomez, Carlos, Marie Michelas, Clara-Maria Scarlata, Anna Salvioni, Carlos Gomez-Roca, Victor Sarradin, Françoise Lauzéral-Vizcaino, Virginie Féliu, Agnès Dupret-Bories, Gwénaël Ferron, and et al. 2022. "Circulating Exhausted PD-1+CD39+ Helper CD4 T Cells Are Tumor-Antigen-Specific and Predict Response to PD-1/PD-L1 Axis Blockade" Cancers 14, no. 15: 3679. https://doi.org/10.3390/cancers14153679
APA StyleMartinez-Gomez, C., Michelas, M., Scarlata, C.-M., Salvioni, A., Gomez-Roca, C., Sarradin, V., Lauzéral-Vizcaino, F., Féliu, V., Dupret-Bories, A., Ferron, G., Sarini, J., Devaud, C., Delord, J.-P., Balança, C.-C., Martinez, A., & Ayyoub, M. (2022). Circulating Exhausted PD-1+CD39+ Helper CD4 T Cells Are Tumor-Antigen-Specific and Predict Response to PD-1/PD-L1 Axis Blockade. Cancers, 14(15), 3679. https://doi.org/10.3390/cancers14153679