
Sex Differences in Taxane Toxicities



Department of Radiation Oncology, University of California, Irvine, CA 92697, USA
*
Author to whom correspondence should be addressed.
Cancers 2022, 14(14), 3325; https://doi.org/10.3390/cancers14143325
Submission received: 4 June 2022
/
Revised: 28 June 2022
/
Accepted: 29 June 2022
/
Published: 8 July 2022
(This article belongs to the Topic Cancer Biology and Radiation Therapy)
{kind=link}
{kind=link}
Abstract
:Simple Summary
Clinically observed sex differences in acute and long-term taxane chemotherapy-induced normal tissue toxicity are routinely documented but remain poorly understood despite the significant impact such toxicities have on treatment tolerance and quality of life outcomes in cancer survivors. This review draws from pre-clinical and clinical literature to highlight sex-specific mechanisms of action in taxane drug toxicity and proposes hypotheses for sex-specific clinical discrepancies in taxane-induced acute and long-term toxicities. To our knowledge, this is the first review exploring how sex as a biological variable impacts taxane-mediated mechanisms of action and clinical outcomes. In doing so, we have provided a novel framework to investigate and understand common sex differences observed in clinical and pre-clinical research.
Abstract
The taxane family of microtubule poisons and chemotherapeutics have been studied for over 50 years and are among the most frequently used antineoplastic agents today. Still, limited research exists characterizing taxane-induced sex-specific mechanisms of action and toxicities in cancer and non-cancerous tissue. Such research is important to advance cancer treatment outcomes as well as to address clinically observed sex-differences in short- and long-term taxane-induced toxicities that have disproportionate effects on female and male cancer patients. To gain more insight into these underlying differences between the sexes, the following review draws from pre-clinical and clinical paclitaxel and taxane oncology literature, examines sex-discrepancies, and highlights uncharacterized sex-dependent mechanisms of action and clinical outcomes. To our knowledge, this is the first literature review to provide a current overview of the basic and clinical sex dimorphisms of taxane-induced effects. Most importantly, we hope to provide a starting point for improving and advancing sex-specific personalized chemotherapy and cancer treatment strategies as well as to present a novel approach to review sex as a biological variable in basic and clinical biology.
1. Introduction
The taxane family of chemotherapeutics are commonly used therapies for the treatment of hormone-refractory breast, prostate, and ovarian cancers, as well as lung and other cancers in women and men. Despite being the subject of scientific publications spanning over 50 years and among the most widely-used antineoplastic agents today, limited research exists characterizing taxane-induced sex-specific mechanisms of action and toxicities. Such research is important to provide insight into clinically-observed sex disparities in chemotherapeutic efficacies and adverse phenotypes [1,2,3,4,5,6,7,8], as well as providing further context into cancer incidence, susceptibility, and survivorship, where women typically have more favorable diagnoses and outcomes than men [7,9]. In addition, sex is still rarely considered when administering chemotherapeutic doses in the clinic [10]. In women, a chemotherapy dose is more frequently decreased during treatment compared to men due to greater acute normal tissue toxicity, such as nausea, vomiting, or neuropathic pain [11,12]. Increased sensitivity in women to chemotherapy’s adverse reactions is likely due to lower drug clearance rates compared to men [8,12]. These predictable pharmacokinetic sex discrepancies bring into question the validity of current body surface area chemotherapy dosing standards [13].
In order to gain more insight into sex differences from taxane therapy effects, the following review examines existing pre-clinical and clinical literature on taxanes in female and male models and highlights fundamental sex-dependent differences in mechanisms of action and clinical outcomes. To our knowledge, this is the first literature review summarizing sex differences in clinically relevant taxane toxicity phenotypes and provides a starting point, rather than a comprehensive analysis, for understanding, improving, and advancing personalized taxane chemotherapy and cancer treatment strategies for women and men.
This review focuses on identifying sex differences in short- and long-term taxane-induced toxicities that affect quality-of-life (QOL), which is relevant to a growing population of cured and surviving patients [14]. Toxicities impacting QOL are of increasing concern to clinicians as aging cancer survivors are at the highest risk of developing long-term changes in cognitive function from exposure to non-specific therapies, such as cranial radiation and systemic chemotherapy [15,16,17,18]. Additionally, there are no known treatments to prevent or mitigate these effects, which often cause permanent changes in a variety of cognitive domains, spanning learning, memory, social behavior, and pain, and significantly impacting patient QOL. Female cancer survivors are the most susceptible population to long-term cancer treatment-induced cognitive dysfunction. This is in part due to higher survival rates for women compared to men with the same cancer diagnosis and quality of care [9], but also due to increased sensitivity to adverse chemotherapy toxicities [11,12]. Thus, there is a genuine need to further investigate the short- and long-term impacts of systemic anticancer therapies at the normal tissue level.
Drawing from paclitaxel and taxane oncology literature and our own research, we provide a current overview of the basic and clinical sex dimorphisms of taxane-induced effects and highlight promising new directions in the fields of taxane and sex differences research [19].
2. Paclitaxel and Taxane Background
Taxanes, starting with the discovery of paclitaxel in 1962, have an interesting and notable research and translational history that has been well reviewed [20,21,22,23,24]. In brief, paclitaxel, initially given the generic name of taxol upon isolation from the American Northwestern Pacific yew tree, Taxus brevifolia Nutt. (Taxaceae), was the only compound developed into clinical use from a 20-year collaborative effort of a nationwide plant-screening program for anticancer agents by the National Cancer Institute (NCI) and US Department of Agriculture [25]. The antineoplastic effects of paclitaxel were observed in animal cells in the 1970s [25,26,27,28], eventually leading to successful tumor regression in clinical trials by the 1980s [22,29]. Due to the limitations of public commercialization at the time, the NCI and US congress transferred the rights of paclitaxel from public to private property of Bristol-Myers Squibb in 1991, despite concerns over granting proprietary rights for a naturally-derived compound discovered through public funding [30]. The Federal Drug Administration (FDA) approved Bristol-Myers Squibb’s Taxol® (generic term renamed to paclitaxel) as a chemotherapeutic treatment for ovarian cancer in 1992, breast cancer in 1994, and lung cancer in 1999 [31]. Paclitaxel has since become a frontline adjuvant treatment for many cancers and efficacy has led the way to the development of similar antimitotic compounds, known as the taxane family of chemotherapeutics. Of the family, docetaxel and cabazitaxel are also approved for clinical use, most notably for castration-resistant prostate cancers.
Taxanes are classified by their microtubule-stabilizing activity, inhibiting microtubule dynamics by preventing disassembly and cellular mitosis [25,28]. Taxanes are poorly soluble in water and require solvents to prevent crystallization during administration. Docetaxel is sufficiently dissolved in Tween 80 and ethanol, but paclitaxel requires Cremaphor EL, which is a powerful solvent known to induce adverse side-effects in patients through acute cytotoxicity. The novel nanoparticle albumin-bound formulation of paclitaxel (nab-paclitaxel) enables solubility in cells without Cremaphor EL. Semi-synthetic taxanes, docetaxel and cabazitaxel, were developed to address initial concerns over Pacific yew environmental protection as well as adverse reactions and resistance to paclitaxel neoadjuvant treatments [32]. Docetaxel, derived from the fast-growing European yew tree, Taxus baccata, was approved by the FDA in 1999 for breast cancer and in 2004 for prostate cancer. Docetaxel has a longer intracellular drug retention and greater affinity to β-tubulin than paclitaxel and differs in microtubule assembly effects and potency [33]. Cabazitaxel was developed to reduce the affinity to the ATP-binging cassette (ABC) drug efflux pump, MDR1/p-glycoprotein, which is a primary mechanism of resistance for paclitaxel and other chemotherapeutics [32]. Cabazitaxel is also more potent than paclitaxel and has shown effectiveness in docetaxel-resistant tumors. It was approved by the FDA in 2010 as a second-line treatment for docetaxel-resistant prostate cancer [34]. Although docetaxel and cabazitaxel have a higher potency than paclitaxel, paclitaxel is still the most commonly prescribed taxane. As a result, the majority of the literature reviewed studies paclitaxel, but the fundamental mechanisms of microtubule stabilization shared by all taxanes cause qualitatively similar effects and symptoms.
3. Translational and Clinical Research
Although little pre-clinical research exists comparing taxane sex effects, decades of clinical use documents sex-specific pharmacological differences after taxane chemotherapy administration. This is of particular concern for oncologists, as chemotherapy administration is based on body surface area instead of pharmacology and contributes to sex differences in therapy toxicities, which frequently lead to adjusted treatment doses [13]. This section will highlight the major sex-specific clinical observations during taxane treatments and corresponding pre-clinical literature that provide insight into the biological mechanisms behind sex differences. The section also covers basic pharmacokinetics as well as pharmacodynamically-focused adverse drug reactions and neurotoxicity, which account for taxanes’ dose-limiting toxicities and quality of life outcomes.
3.1. Pharmacokinetics
One of the most prominent sex differences in pharmacokinetics is the faster elimination of almost all chemotherapies (and many drugs) in men compared to women [3,8,35]. This is thought to reduce men’s risk of adverse drug reactions, but also brings to question whether each sex regularly receives their maximum tolerated dose, which is critical for optimizing curative intent [12,13,36]. A retrospective population analysis of solid tumor patients receiving paclitaxel infusions (n = 168) found that on average, male patients had a 20% higher maximal elimination capacity compared to female patients [37]. Paclitaxel is known to bind to plasma proteins extensively and non-specifically and, therefore, has negligible red blood cell transport [38], suggesting little impact of sex in hematocrit transport activity [39]. Sex differences in pharmacokinetics may impact drug disposition and important treatment factors, such as intracellular taxane concentrations in normal and metastatic tissue [40].
3.2. Adverse Drug Reactions
Clinical taxane use has been associated with more adverse drug reactions than other chemotherapies, particularly for breast cancer patients. The first group of breast cancer patients to receive paclitaxel (n = 55 studied) upon approval in 1999 demonstrated significantly higher rates of adverse acute toxicities compared to non-taxane-treated patients (n = 83), including arthralgia/myalgia (45% vs. 26%) and ataxia (20% vs. 5%) [41]. Paclitaxel-induced toxicities also seem to be more common in women, as one retrospective analysis of non-small lung cancer patients receiving paclitaxel reported more adverse drug reactions in women (77%) compared to men (66%, p = 0.0004). However, in these lung cancer patients, women demonstrated longer progression-free survival than men (Hazard Ratio (HR) 0.83, p = 0.02) [42]. Such data supports sex-specific trends in overall cancer toxicities as well as cancer outcomes [7]. Women are more likely to suffer from more serious adverse drug toxicities compared to men after most types of chemotherapy exposures [5,11,43], but benefit from better survival outcomes [44,45]. The current understanding of sex dimorphisms in disease suggest sex-specific immune modulation plays a critical role in cancer treatment-induced normal tissue toxicities and survival outcomes [46,47,48], thereby warranting further investigation into the context of taxane treatments.
The most acute taxane-associated adverse drug reactions are hypersensitivity reactions (HSRs), including flushing, hemodynamic alterations, dyspnea, musculoskeletal/neuropathic pain, and gastrointestinal issues [49]. All chemotherapeutics can induce HSRs varying in severity and degree, but taxane-induced HSRs typically manifest during the first or second infusion [50,51]. The immune response involving Immunoglobulin E (IgE) and/or IgG mast cell signaling mediates hypersensitivity reactions, but the impact of sex on mechanisms have yet to be characterized in pre-clinical or clinical literature. Fortunately, patients are successfully treated with a premedication of corticosteroids and antihistamines, but poor measures exist to predict patient risk, severity, and optimal treatment strategy for taxane-induced HSRs. This is in part due to the solvent and taxane moiety-dependent variability in HSR mechanisms, but sex has also been considered as a risk factor due to the pharmacokinetic differences of steroids and taxanes elimination rates and frequent observations of HSRs in breast and ovarian cancer patients [52]. In fact, a recent medical record analysis of 3181 Stanford Cancer Institute patients receiving paclitaxel or docetaxel associated the female sex with an increased risk of overall HSRs (HR 1.26) and gynecology oncology patients had an increased risk of overall (HR 1.34) and high-grade HSRs (HR 2.34) [53]. As mentioned before, such data support other trends of higher adverse toxicities in women. Since HSRs are easily treatable and transient (only associated with infusions), little pre-clinical research has explored sex-specific mechanisms. However, such research could provide critical insight into fundamental sex differences in the immune response that contributes to the development of other toxicities, such as taxane dose-limiting and quality of life-impairing neurotoxicity.
3.3. Neurotoxicity
Most chemotherapeutic agents, including taxanes, have limited blood brain barrier (BBB) penetration and are readily purged from the brain through P-glycoprotein (P-gp) pumps [54,55]. However, certain taxane preparations, like cabazitaxel, have greater BBB permeability [56] or contain solvents associated with additional BBB permeability, such as paclitaxel in Cremophor EL, which inhibits hippocampal cell proliferation in rodents [57]. Dorsal root ganglia and peripheral nerves are subject to chemotherapeutic agent toxicity through blood–nerve barrier permeability, which, due to lymphatic and P-gp absence in nerves, likely causes the commonly observed chemotherapy-induced peripheral neuropathic toxicities [58]. Although such distinguishing features of the central nervous system are thought to exist independent of sex, the majority of such basic research has utilized male rodents and therefore sex differences in the aforementioned neuronal characteristics cannot be entirely ruled out.
Clinical manifestations of taxane-induced neurotoxicity predominantly include peripheral neuropathy and cognitive dysfunction [59]. Limited comparisons of sex differences in the neurotoxicity of taxane recipients exist, but as most clinical data document women (predominantly breast cancer survivors), broader comparisons are confounded and suggest the preponderance of taxane-induced neurotoxic phenotypes that burden women.
3.3.1. Peripheral Neuropathy
Taxane-induced peripheral neuropathy (PN)-associated pain is the most concerning and critical clinical observation affecting patients’ immediate and long-term quality of life [59,60,61,62]. Since the elimination of chemotherapy-induced neutropenia through the implementation of preventative granulocyte colony-stimulating factor treatment [63], PN-associated pain is the most common reason for taxane treatment dose reductions [64]. Although most cancer patients receiving taxanes develop PN [59], no comprehensive analysis of clinically observed sex differences exists. In an analysis of 219 American breast cancer survivors treated with adjuvant paclitaxel, 97% developed PN and 60% developed chronic PN one year following the cessation of treatment cessation [65]. On the other hand, in 82 prostate cancer patients who received docetaxel, only 32% developed PN [66]. Although docetaxel is thought to induce less PN than paclitaxel [59], these data further highlight the distinct clinical sex differences in taxane treatment strategies and PN outcomes that produce a greater burden on female cancer survivors [67,68]. Moreover, only non-pharmacological PN pain management strategies exist for taxane recipients [59,69,70]. One reason for unsuccessful pharmacological approaches may be due to the lack of mechanistic insight regarding fundamental sex differences in taxane-induced PN. However, in recent years, growing pre-clinical evidence suggests that taxane-induced PN manifests through distinct neuroimmune mechanisms in female and male rodents and may provide valuable insight into understanding and treating sex differences in taxane-induced PN as well as other toxicities.
Based on pre-clinical studies primarily using male models [71], glial cells and pro-inflammatory immune responses from innate immune cells are implicated in peripheral neuropathy [72], including taxane-induced pain [73]. Candidate mechanisms, such as toll-like receptor (TLR) 4 activation [74], which is extensively studied in male rodents, have only recently been examined in females. As a result, sex-specific findings in the field of pain have been categorical, illustrating that microglia, and their associated signaling molecules, drive neuropathic pain in only the male sex [75,76,77].
Supporting this, one recent study found that TLR9 inhibition only attenuate paclitaxel-induced mechanical allodynia (touch induced pain) in males and not female mice. However, interestingly, TLR9 antagonism reduced paclitaxel-induced pain in female nude mice lacking T and B cells [78], suggesting that females preferentially utilize humoral/adaptive immunity, but also retain the ability to recruit innate immunity-dependent microglia when adaptive immunity is unavailable [77].
Sex hormones and sex organs drive sex-specific immunity [46]. In one study examining pain thresholds and pro-inflammatory cytokine receptor levels in the dorsal root ganglion of female rats exposed to paclitaxel, ovariectomies significantly increased pain thresholds and decreased receptor expression levels after exposure compared to females with 17β-estradiol and progesterone replacement [79], suggesting that paclitaxel-induced neuropathic pain in females is sex hormone-dependent.
Sex differences in stress phenotypes [80] is a growing field of research as well as a clinical risk factor and comorbidity of taxane-induced PN [68]. In a study examining the neuroendocrine stress axis in rats, paclitaxel-induced hyperalgesia was significantly more attenuated in female rats compared to males following β2-adrenergic receptor reduction through targeted intrathecal antisense oligodeoxynucleotide administration (ODN). By contrast, ODN decreasing glucocorticoid receptors attenuated paclitaxel-induced pain in males, but not females. Additionally, the study found that neonatal handling prevented paclitaxel-induced PN in male but not female rats, providing evidence of distinct sex-specific neuroendocrine mechanisms related to paclitaxel-induced pain [81].
Based on these recent findings and the current understanding of sex differences in immune functions, female-specific pain is likely due to humoral immune responses, such as T and B cell activation, and may require novel treatment approaches and a more sophisticated understanding of mechanisms in both sexes [82,83,84]. These data also bring into question the other possible impacts of sexually dimorphic immune responses in taxane-induced toxicities. Divergent preferential immune function between the sexes, defined as females utilizing more adaptive immunity while males rely more heavily on innate immunity, is becoming a dominant theme in pharmacology and etiology [7,46,85] and, therefore, perhaps it should be the starting point of studying sex as a biological variable in drug and disease research.
3.3.2. Cognitive Dysfunction
Chemotherapy-induced cognitive dysfunction, also known as ‘chemobrain,’ is of growing clinical concern due to detrimental impacts on long-term quality of life in increasing populations of aging cancer survivors [16,86,87,88,89,90,91,92]. As outlined in the above Peripheral Neuropathy section, the existing literature of pre-clinical rodent studies comparing both sexes suggest greater taxane-induced peripheral neuropathic pain in females than males. However, paradoxically, recent (albeit limited) literature comparing paclitaxel-induced cognitive dysfunction in both sexes demonstrates female-specific resistance. Sixteen mg/kg of paclitaxel (dissolved in Cremophor EL and ethanol) in intraperitoneal exposures of C57BL6 mice induced cognitive dysfunction in a prefrontal cortex-associated Novel Object Recognition task and anxiety-like behavior in Elevated Plus Maze in males but not females [93]. We also observed female-specific cognitive and cytotoxicity protection after 150 mg/kg and 300 mg/kg paclitaxel (dissolved in Saline and ethanol) exposures in C57BL6 mice, characterized by significantly less weight loss, higher survival, sustained prefrontal cortex-associated Fear Extinction performance, and less anxiety-like behavior during Light/Dark Box testing in female rodents [94]. In the same study, we hypothesized that (a) reproductive senescence in aged Wild-Type females and (b) RhoB GTPase deficiency would attenuate the female-specific resistance through short-circuiting estrogen-mediated female-specific (a) neuroprotection [95,96] and (b) RhoB-dependent protection [97]. Interestingly, aged (22-month-old) female Wild-Type and female and male RhoB-deficient animals were all resistant from paclitaxel-induced cognitive dysfunction, but as hypothesized, survival and weight decreases occurred in aged Wild-Type and RhoB-deficient females. Such data demonstrating female-specific neuroprotection seem contrary to the cognitive impairment frequently reported with adjuvant taxane treatments in women.
The first group of breast cancer patients to receive the FDA-approved paclitaxel (n = 55 studied) not only demonstrated significantly higher rates of adverse acute toxicities (mentioned in the Adverse Drug Reactions section), but also significantly more mental distress and reduced mental quality of life compared to women who received other chemotherapies (p < 0.023). In addition, this study observed longer emotional recovery for paclitaxel-treated patients, which required an average of 2 years compared to 6–12 months for patients not receiving taxanes [41]. In a recently published study comparing sexes, a small population of nasopharyngeal carcinoma survivors receiving adjuvant docetaxel, cisplatin, and fluorouracil chemotherapy, found that the female sex was associated with cognitive dysfunction (p = 0.039), manifesting in 50% of women (8 of n = 16) and only 20% of men (10 of n = 50) [98]. A 2017 longitudinal study of breast cancer survivors comparing adjuvant chemotherapies with and without taxanes (n = 51) demonstrated short- and long-term cognitive impairment in attention and executive function after all treatments, with a more pronounced impact on short-term verbal learning and speed measures in the taxane group [99]. Another analysis of breast cancer survivors shortly after and 1 year after adjuvant chemotherapy (majority with paclitaxel, up to 8 patients without) demonstrated acute (65%, 24 of n = 37) and long-term (61%, 17 of n = 28) cognitive decline [100]. A recent female-only rodent study supports these clinical data of adjuvant therapy-induced cognitive impairment in women. Brown et al. characterized the diminished hippocampal-dependent cognitive behavior and compromised dendritic architecture and signaling proteins in female hippocampal tissue after adjuvant docetaxel, doxorubicin, and cyclophosphamide therapy [101].
In perhaps the only clinical analysis of cognitive functioning after taxanes in males, 65+ year old castrate-resistant prostate cancer patients treated with adjuvant therapies, including docetaxel, did not experience significant cognitive impairment measured by the Montreal Cognitive Assessment [102]. In contrast, an analysis comparing 65+ year old early-stage breast cancer survivors of all treatments (radiotherapy with or without doxorubicin ± docetaxel) demonstrated significant impacts on cognitive decline in 49% of patients, with the oldest patients (70–81 years) being most sensitive to docetaxel-associated decline (p = 0.05) [103].
Although little research investigates sex-specific burdens of chemotherapy-induced cognitive dysfunction in cancer survivors, it is interesting to point out that almost all clinical studies assessing taxane-induced cognitive outcomes are in breast cancer survivors, while most pre-clinical rodent studies examine outcomes in males [57,104,105,106,107]. The larger amount of female-specific clinical data is in part due to the total of 8.7 million women cancer survivors in the U.S. (over 600,000 more survivors than men), of which 3.8 million are breast cancer survivors [44]. In addition, the larger proportions of women suffering from adverse chemotherapy effects compared to men undoubtably impacts stress and depression and, therefore, cognitive function and quality of life during and after recovery. The diversity in clinical patient data (e.g., cancer type, adjuvant treatments, age, etc.) is not always comparable to highly controlled pre-clinical studies and likely accounts for the discrepancy in taxane-induced cognitive dysfunction phenotypes presenting in women, but less frequently in female rodents.
4. Basic Research
‘Mechanism of action’ in cancer therapy refers to an agent’s primary antineoplastic mechanism, which is often antimitotic in nature. Twentieth century chemotherapy pre-clinical research almost exclusively studied cancer cells, and scientists were predominantly concerned with the ‘mechanisms of action’ directly related to inhibiting cancer growth. In the 21st century, as survival rates and treatment efficacies increased, quality-of-life outcomes became more important and a growing issue for cancer survivors, increasing biomedical research interests in normal tissue effects. Due to the non-specific nature of chemotherapeutics, it is important to consider how mechanisms of action impact normal tissue toxicity and sex.
It is interesting to note that virtually all basic and pre-clinical in vivo cancer drug research done in the 1900s, such as with paclitaxel, utilized female HeLa cervical adenocarcinoma cell lines, the first and most widely used immortalized human cell line derived from 31-year-old Henrietta Lacks’ aggressive cervical cancer in 1951 [25,108]. Although no other cell line has contributed more to foundational biological understanding and biomedical advances, its sex was rarely considered or examined as an experimental variable. Despite little consideration of sex in basic research, clinical data provides justification for such research. As discussed earlier in Translational and Clinical Research, women have better treatment outcomes across most cancer types, and sex differences are readily observed in adverse drug effects, drug disposition, and pharmacokinetics, which undoubtably impacts chemotherapy antineoplastic efficacy and normal tissue toxicity. The following sections draw from fundamental research (not considering sex) of taxane antineoplastic activity and sex-specific cancer data to highlight potential sex-specific mechanisms of toxicity relevant to most, if not all, tissue.
4.1. Mitotic Arrest
After initial observations of paclitaxel-induced Kb HeLa-derived cell growth inhibition, published in 1971 [25], tumor drug researcher, Susan Horwitz, was the first to characterize the mechanism of paclitaxel-induced mitotic arrest with her graduate student, Peter Schiff [24]. Their landmark 1979 publication identified paclitaxel as a microtubule assembly promoting poison, unlike previously identified microtubule poisons that prevent microtubule polymerization. In the words of Horwitz, microtubules behave like “paralyzed cytoskeleton[s]” [109] in the presence of β-tubulin-bound paclitaxel. Paclitaxel reduces the critical concentration of tubulin subunits necessary for microtubule polymer formation, increasing the percentage of α- and β-tubulin heterodimers assembled, and inhibits mitosis through the failure of metaphase depolymerization. They observed that paclitaxel-induced microtubule growth was even resistant to cold and calcium depolymerization treatments [28]. The observation of paclitaxel-induced polymerization and the “parlay[sis]” of microtubules causing mitotic arrest was confirmed in a variety of cell and animal models, in both cancerous and non-cancerous tissue [27,110,111,112]. For decades, the dominant perception of paclitaxel’s tumor treatment efficacy was through the inhibition of metaphase bipolar spindle depolymerization and mitotic checkpoint-dependent mitotic arrest [113,114]. However, investigators have also observed taxane-induced apoptosis that is unassociated with mitotic arrest with formidable cancer regression in low-proliferating tumors, suggesting alternative cytotoxic pathways that are critical in taxane antineoplastic activity [111,112,115,116,117]. In fact, drugs designed to exclusively inhibit mitosis have shown limited antineoplastic success and fail to replace microtubule poison chemotherapeutics [111,116,118,119]. Advances in characterizing taxane-induced microtubule dysfunction beyond mitotic arrest suggest alternative cytotoxic mechanisms of action may be the preponderance of taxane anticancer activity (Figure 1).
4.2. Microtubule Dynamic Dysfunction
Although the primary antineoplastic mechanism of taxanes was initially thought to involve mitotic arrest through microtubule dysfunction, current scientific consensus also attributes taxane tumoral regression efficacy to additional, perhaps more potent, microtubule-dependent cytotoxic mechanisms (refer to prior section on Mitotic Arrest). Due to the importance of microtubule function in a range of cellular activity, most notably mitotic spindle assembly, cytoskeletal structure, and cytoplasmic cellular cargo migration [120,121], it is hypothesized that taxanes also elicit antineoplastic activity through mitotic checkpoint-independent cell death [115,117,122] and cellular transport disruption [123,124,125]. In fact, early paclitaxel research demonstrated aberrant microtubule dynamics and toxicity at doses insufficient to induce mitotic arrest [126,127,128,129]. In a notable recent study, adjuvant paclitaxel-treated human breast tumor biopsies revealed that intratumoral drug concentrations were insufficient to induce mitotic arrest and tumor regression efficacy, which was attributed to increased multipolar spindle formations, implicating p53-independent, chromosome missegregation-induced cellular death [122]. Such data suggest that taxanes can produce a range of microtubule-dependent aberrant phenotypes, dependent on the intracellular drug concentration and phase of cell cycle.
Understanding the impact of taxanes on microtubule-dependent nuclear trafficking and signaling is perhaps one of the most challenging phenotypes to characterize due to the extent and complexity of microtubule-mediated mechanisms. However, the influence of taxanes on androgen receptor signaling has been extensively observed and studied in the field of prostate cancer and provides additional understanding about the impact of taxane-induced microtubule dysfunction and sex-specific mechanisms of toxicity.
4.2.1. Androgen Receptor Signaling Dysfunction
In the field of prostate cancer research, taxanes are known to inhibit androgen receptor (AR) signaling through microtubule dysfunction [125,130] (Figure 2). AR signaling mechanisms are the cause and treatment target for prostate cancers [131,132,133]. Androgen deprivation therapy (ADT), which is used to block AR or eliminate AR ligands, initially works to inhibit the growth of prostate cancers as a frontline treatment, but over time castrate-resistant prostate cancer (CRPC) occurs through loss of ADT sensitivity [132]. In 2004, taxanes became the first class of chemotherapeutic drugs that demonstrated improved survival in CRPC and are now used in conjunction with frontline ADT treatments prior to the development of castration resistance [134,135,136,137]. Although research on the effects of taxanes on AR signaling is almost exclusive to the field of prostate oncology, these male-specific observations provide context into fundamental sex differences of systemic taxane exposures for male and female malignant and normal tissue.
Researchers have observed the taxane-dependent inhibition of AR expression and activity in prostate cancer [130,138,139]. Specifically, Darshan et al. demonstrated that paclitaxel inhibits microtubule ligand-induced AR nuclear accumulation and downstream transcriptional activity [125]. The same study indicated a significant correlation between AR cytoplasmic sequestration (instead of nuclear accumulation) and therapeutic efficacy in the circulating tumor cells of CRPC patients receiving taxanes. Interestingly, although not entirely understood, taxane-induced interference with AR signaling produces antineoplastic efficacy in both naïve and castrate-resistant malignancies [132]. One pre-clinical research study observed that chronic AR activation, through testosterone-BSA exposure, enhanced paclitaxel microtubule disrupting dynamics, inhibited cell proliferation, and induced apoptosis in androgen sensitive and insensitive human prostate cancer cell lines [140]. In the same study, xenografted mouse tumors (both androgen sensitive and insensitive) decreased in mass with both testosterone-BSA and paclitaxel separately, an effect that was enhanced in combination. These data support clinical observations of taxane treatment efficacies in prostate cancers that are sensitive and resistant to ADT [134].
Androgen receptor signaling is critical for prostate cancer as well as normal male tissue development, thus providing a potential sex-specific discrepancy in taxane tissue toxicity. AR knockout (KO) mouse studies have not only demonstrated the critical function of AR in male (and female) gonad and gamete development, but also in male body fat, bone, blood, and immune phenotypes [141,142,143,144]. Conditional and constitutive ARKO male mice have severe deficits in both innate and adaptive immunity, resulting in phenotypes such as thymus enlargement, immature B-cell populations, and risk of neutropenia and bacterial infections [141]. Although little research has investigated such ARKO phenotypes in female mice, since ARs dominate signaling function for male sex hormones (while estrogen receptors dominate female sex hormones), taxanes may induce disproportionate toxicity in male tissue. Supporting this idea, a developmentally critical Y-linked gene (found in male mice and men) modulates docetaxel sensitivity through AR signaling [145]. KDM5D (lysine-specific demethylase 5D) physically interacts with nuclear AR and demethylates (deactivates) H3K4me3 transcriptional marks, which normally regulate enhanced AR transcriptional activity (possibly via negative feedback). Accordingly, the attenuation of KDM5D expression led to increases in H3H4me3 marks in promoter regions of AR-regulated genes as well as protection against docetaxel in AR-positive prostate cancer cell lines. In addition, an Oncomine cancer database analysis revealed significantly lower KDM5D expression in CRPC patient samples with a poorer cancer prognosis and treatment outcome. Although KDM5D expression is associated with taxane sensitivity, the fundamental mechanism is still unclear. ERG (E-26 transformation-specific-related gene) is another gene implicated in AR-signaling in prostate cancer [146,147] through microtubule depolymerization. ERG binds to αβ-tubulin, and when overexpressed, reduces binding site availability for taxane-induced microtubule polymerization, leading to taxane resistance in prostate cancer cells and tumors [148]. Such data defines taxane as AR signaling poisons, in part, due to epigenetic regulation. However, it is important to note that AR signaling can also take place through nuclear transport in a microtubule-independent fashion. The capability of tumors to preferentially shuttle AR through nuclear transport is thought to contribute to taxane resistance in prostate cancer. The ability to circumvent microtubule transport machinery has been demonstrated by AR-V7, which is a commonly found AR splice variant that lacks the hinging domain necessary to attach to the tubulin-dynein transporter molecule for minus-end (nuclear) microtubule transport [149]. The identification of AR-V7 splice variants in tumors is associated with advanced CRPC, taxane resistance, and reduced patient survival [150,151,152,153,154,155].
4.2.2. Estrogen-Mediated Microtubule Dynamics
As the field of prostate cancer continues to elucidate the complex relationships between taxane-induced microtubule dysfunction and AR signaling, the role of estrogen and estrogen receptors (ER) on taxane-induced microtubule dynamics is less clear. However, a couple of studies suggest that estrogens and androgens may have opposite roles in microtubule polymerization. One study examining the roles of sex hormone exposures on tubulin polymerization induction observed that prior incubation of tubulin proteins and cells, derived from fetal rat hippocampi, with 17β-estradiol inhibited microtubule assembly, while incubation with testosterone inhibited microtubule disassembly [156]. Other studies have also observed an AR+ dependent relationship between testosterone/androgen and microtubule polymerization [140,157], suggesting the presence of androgens may promote microtubule polymerization/assembly indirectly. On the other hand, recent evidence suggests estrogens may have a direct role in microtubule polymerization and function. Lo et al. conducted a computational protein screen analysis that unexpectedly identified ER as a cognate receptor to the β-tubulin taxane binding site, suggesting tubulin–ER cross-reactivity. The researchers then confirmed that the taxane binding site had affinity with estrogen and selective estrogen receptor modulators (SERMs) in human epithelial cells and modified microtubule dynamics in a similar fashion as paclitaxel [158]. Interestingly, a separate in vitro study observed no synergistic or additive antimetastatic effects with the co-administration of the SERM, tamoxifen, with paclitaxel in ER+ breast cancer cells [159]. Such data suggests that ER ligands may directly interact with tubulin and modulate microtubule dynamics, and in the presence of taxanes, attenuate taxane-induced microtubule polymerization.
In summary, the foregoing sections outline possible sex-hormone-mediated sex differences in taxane-dependent microtubule dysfunction. Taxanes induce AR signaling dysfunction in a microtubule dependent manner that provide a sex-specific toxicity in male tissue due to androgen dominance. In females, the cross-reactivity of ER ligands at the taxane-tubulin binding site may inhibit taxane-mediated microtubule polymerization and attenuate downstream microtubule-dependent toxicity. This suggests sex-hormone-mediated male-specific susceptibility and female-specific protection against taxane-induced microtubule dysfunction. Our recent study, observing significant male-specific normal tissue cytotoxicity after paclitaxel exposure in mice [94], provides further evidence of sex-specific taxane-induced toxicity and highlights the importance of investigating normal tissue effects of systemic cancer treatment.
5. Conclusions
This review provides the first multidisciplinary analysis highlighting how basic sex-specific taxane-induced toxicity mechanisms may impact clinical taxane treatment outcomes. Basic research has established microtubule dynamic dysfunction as the primary antineoplastic mechanism of action through the inhibition of mitosis and other microtubule-dependent cellular functions. Taxane-induced microtubule dependent androgen receptor signaling dysfunction (extensively studied in the field of prostate cancer) and the potential cross-reactivity of estrogen receptor ligands inhibiting taxane-tubulin binding provides the most robust evidence for developing a hypothesis for male-specific toxicity and female-specific protection after paclitaxel exposure. However, clinical observations paradoxically show higher risk in female patients during and after taxane treatment for developing acute and long-term adverse effects. Considering both basic mechanisms and clinical observations, sex hormones (androgens and estrogens), adjuvant exposure strategies, and their cumulative impacts on sex-specific immune modulation likely account for this discrepancy.
Despite the defined mechanism of action of taxanes on the inhibition of microtubule depolymerization and dynamics, other downstream effects of microtubule dysfunction on cellular activities, whether mitosis, intracellular transport, or cytoskeletal dynamics, may well engage immune response pathways. Accordingly, mounting evidence highlights sexually dimorphic strategies of immune responses, which are mediated by sex hormones and sex organs [46,47,83,85,160]. As mentioned in the Section 3.3.1, the field of pain research, including taxane-induced peripheral neuropathy, has characterized significant sex differences in the last decade [75,76,77]. Females manifest neuropathic pain through humoral response and adaptive immunity, while pain in males is induced through innate immune response [77]. Other fields have characterized similar trends in sex-specific systemic immunity, such as in infectious disease research (e.g., SARS-CoV-2) [161,162,163,164,165] and in chronic autoimmune conditions [46,47,48,85,160]. Therefore, although males are more susceptible to taxane-induced microtubule-dependent dysfunction through reliance on AR signaling, they may be protected from acute and chronic humoral responses initiated by autoimmune dysfunction, which are characteristic of hypersensitivity reactions, nausea, vomiting, and peripheral neuropathy more commonly found in female patients treated with taxanes. Although women are susceptible to greater normal tissue discomfort and toxicity from taxanes, they are also likely to benefit from greater antineoplastic efficacy, potentially accounting for better survival outcomes in female patients.
This review highlights the importance of addressing sex as a biological variable in research and provides a framework to investigate sex differences in other fields of drug and disease research. In the case of taxane research, although the field of prostate cancer clearly identifies androgen receptor signaling as a biological target for taxanes, sex comparisons do not exist, and sex-specific toxicities still remain under-investigated. In the case of peripheral neuropathy research, the recent addition of female models has shifted the fundamental understanding of pain mechanisms and strategies to incorporate sex as an important treatment variable. In sum, pre-clinical studies focused on sex differences remain understudied and are essential to not only decipher clinical observations, but to also compare fundamental biological mechanisms of action for the purpose of advancing personalized treatment strategies and improving the quality of life of male and female cancer survivors alike.
Author Contributions
Writing—Original Draft Preparation N.N.C. and C.L.L.; Conceptualization N.N.C. and C.L.L.; Writing—Review & Editing N.N.C. and C.L.L.; Funding Acquisition N.N.C. and C.L.L. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Institutes of Health (NIH) through the National Institute of Neurological Disorders and Stroke Grant NS089575 (C.L.L.) and by the National Center for Research Resources and the National Center for Advancing Translational Sciences through Grant TL1 TR001415 (N.N.C.). The content is solely the responsibility of the authors and does not necessarily represent the official views of the NIH.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Tran, C.; Knowles, S.R.; Liu, B.A.; Shear, N.H. Gender Differences in Adverse Drug Reactions. J. Clin. Pharmacol. 1998, 38, 1003–1009. [Google Scholar] [CrossRef] [PubMed]
- Rademaker, M. Do Women Have More Adverse Drug Reactions? Am. J. Clin. Dermatol. 2001, 2, 349–351. [Google Scholar] [CrossRef] [PubMed]
- Anderson, G.D. Sex and Racial Differences in Pharmacological Response: Where Is the Evidence? Pharmacogenetics, Pharmacokinetics, and Pharmacodynamics. J. Women’s Health 2005, 14, 19–29. [Google Scholar] [CrossRef] [PubMed]
- Bren, L. FDA Consumer; Federal Drug Administration: Washington, DC, USA, 2005; Volume 39, pp. 10–15. [PubMed]
- Schmetzer, O.; Flörcken, A. Sex Differences in the Drug Therapy for Oncologic Diseases. Handb. Exp. Pharmacol. 2012, 214, 411–442. [Google Scholar] [CrossRef]
- Wang, J.; Huang, Y. Pharmacogenomics of Sex Difference in Chemotherapeutic Toxicity. Curr. Drug Discov. Technol. 2007, 4, 59–68. [Google Scholar] [CrossRef]
- Haupt, S.; Caramia, F.; Klein, S.L.; Rubin, J.B.; Haupt, Y. Sex Disparities Matter in Cancer Development and Therapy. Nat. Rev. Cancer 2021, 21, 393–407. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.-I.; Lim, H.; Moon, A. Sex Differences in Cancer: Epidemiology, Genetics and Therapy. Biomol. Ther. 2018, 26, 335–342. [Google Scholar] [CrossRef]
- Cook, M.B.; McGlynn, K.A.; Devesa, S.S.; Freedman, N.D.; Anderson, W.F. Sex Disparities in Cancer Mortality and Survival. Cancer Epidemiol. Biomark. Prev. 2011, 20, 1629–1637. [Google Scholar] [CrossRef] [Green Version]
- Islam, M.M.; Iqbal, U.; Walther, B.A.; Nguyen, P.A.; Li, Y.C.; Dubey, N.K.; Poly, T.N.; Masud, J.H.B.; Atique, S.; Syed-Abdul, S. Gender-Based Personalized Pharmacotherapy: A Systematic Review. Arch. Gynecol. Obstet. 2017, 295, 1305–1317. [Google Scholar] [CrossRef]
- Davidson, M.; Wagner, A.D.; Kouvelakis, K.; Nanji, H.; Starling, N.; Chau, I.; Watkins, D.; Rao, S.; Peckitt, C.; Cunningham, D. Influence of Sex on Chemotherapy Efficacy and Toxicity in Oesophagogastric Cancer: A Pooled Analysis of Four Randomised Trials. Eur. J. Cancer 2019, 121, 40–47. [Google Scholar] [CrossRef]
- Wagner, A.D.; Oertelt-Prigione, S.; Adjei, A.; Buclin, T.; Cristina, V.; Csajka, C.; Coukos, G.; Dafni, U.; Dotto, G.-P.; Ducreux, M.; et al. Gender Medicine and Oncology: Report and Consensus of an ESMO Workshop. Ann. Oncol. 2019, 30, 1914–1924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wagner, A.D. Sex Differences in Cancer Chemotherapy Effects, and Why We Need to Reconsider BSA-Based Dosing of Chemotherapy. ESMO Open 2020, 5, 770. [Google Scholar] [CrossRef] [PubMed]
- Torre, L.A.; Siegel, R.L.; Ward, E.M.; Jemal, A. Global Cancer Incidence and Mortality Rates and Trends—An Update. Cancer Epidemiol. Biomark. Prev. 2016, 25, 16–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahles, T.A.; Saykin, A.J. Breast Cancer Chemotherapy-Related Cognitive Dysfunction. Clin. Breast Cancer 2002, 3, S84–S90. [Google Scholar] [CrossRef]
- Wefel, J.S.; Schagen, S.B. Chemotherapy-Related Cognitive Dysfunction. Curr. Neurol. Neurosci. Rep. 2012, 12, 267–275. [Google Scholar] [CrossRef]
- Koppelmans, V.; Breteler, M.M.B.; Boogerd, W.; Seynaeve, C.; Schagen, S.B. Late Effects of Adjuvant Chemotherapy for Adult Onset Non-CNS Cancer; Cognitive Impairment, Brain Structure and Risk of Dementia. Crit. Rev. Oncol. Hematol. 2013, 88, 87–101. [Google Scholar] [CrossRef]
- Jansen, C.; Miaskowski, C.; Dodd, M.; Dowling, G.; Kramer, J. Potential Mechanisms for Chemotherapy-Induced Impairments in Cognitive Function. Oncol. Nurs. Forum 2005, 32, 1151–1163. [Google Scholar] [CrossRef] [Green Version]
- Stivers, N.N. Sex Differences in Taxane Cancer Treatment Toxicities. Available online: https://escholarship.org/uc/item/58p5p3pp#author (accessed on 23 June 2022).
- Walsh, V.; Goodman, J. From Taxol to Taxol®: The Changing Identities and Ownership of an Anti-Cancer Drug. Med. Anthropol. 2002, 21, 307–336. [Google Scholar] [CrossRef]
- Renneberg, R. Biotech History: Yew Trees, Paclitaxel Synthesis and Fungi. Biotechnol. J. 2007, 2, 1207–1209. [Google Scholar] [CrossRef]
- Wiernik, P.H.; Schwartz, E.L.; Strauman, J.J.; Dutcher, J.P.; Lipton, R.B.; Paietta, E. Phase I Clinical and Pharmacokinetic Study of Taxol. Cancer Res. 1987, 47, 2486–2493. [Google Scholar]
- Kampan, N.C.; Madondo, M.T.; McNally, O.M.; Quinn, M.; Plebanski, M. Paclitaxel and Its Evolving Role in the Management of Ovarian Cancer. BioMed Res. Int. 2015, 2015, 413076. [Google Scholar] [CrossRef] [PubMed]
- Weaver, B.A. How Taxol/Paclitaxel Kills Cancer Cells. Mol. Biol. Cell 2014, 25, 2677–2681. [Google Scholar] [CrossRef] [PubMed]
- Wani, M.C.; Taylor, H.L.; Wall, M.E.; Coggon, P.; Mcphail, A.T. Plant Antitumor Agents.VI. The Isolation and Structure of Taxol, a Novel Antileukemic and Antitumor Agent from Taxus brevifolia. J. Am. Chem. Soc. 1971, 93, 2325–2327. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, D.A.; Johnson, R.K. Cytologic Evidence That Taxol, an Antineoplastic Agent from Taxus brevifolia, Acts as a Mitotic Spindle Poison. Cancer Treat. Rep. 1978, 62, 1219–1222. [Google Scholar] [PubMed]
- Schiff, P.B.; Horwitz, S.B. Taxol Stabilizes Microtubules in Mouse Fibroblast Cells. Proc. Natl. Acad. Sci. USA 1980, 77, 1561–1565. [Google Scholar] [CrossRef] [Green Version]
- Schiff, P.B.; Fant, J.; Horwitz, S.B. Promotion of Microtubule Assembly in Vitro by Taxol. Nature 1979, 277, 665–667. [Google Scholar] [CrossRef]
- McGuire, W.P.; Rowinsky, E.K.; Rosenhein, N.B.; Grumbine, F.C.; Ettinger, D.S.; Armstrong, D.K.; Donehower, R.C. Taxol: A Unique Antineoplastic Agent with Significant Activity in Advanced Ovarian Epithelial Neoplasms. Ann. Intern. Med. 1989, 111, 273–279. [Google Scholar] [CrossRef]
- United States Congress; House Committee on Small Business; Subcommittee on Regulation; Business Opportunities, and Energy. Exclusive Agreements between Federal Agencies and Bristol-Myers Squibb Co for Drug Development; United States. Congress. House. Committee on Small Business. Subcommittee on Regulation, Business Opportunities, and Energy: Washington, DC, USA, 1992.
- Tuma, R.S. Taxol’s Journey from Discovery to Use. Oncol. Times 2003, 25, 52–57. [Google Scholar] [CrossRef]
- Yared, J.A.; Tkaczuk, K.H. Update on Taxane Development: New Analogs and New Formulations. Drug Des. Dev. Ther. 2012, 6, 371–384. [Google Scholar] [CrossRef] [Green Version]
- Eisenhauer, E. Docetaxel: Current Status and Future Prospects. J. Clin. Oncol. 1995, 13, 2865–2868. [Google Scholar] [CrossRef]
- Paller, C.J.; Antonarakis, E.S. Cabazitaxel: A Novel Second-Line Treatment for Metastatic Castration-Resistant Prostate Cancer. Drug Des. Dev. Ther. 2011, 5, 117–124. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, E. Gender-Related Differences in Pharmacokinetics and Their Clinical Significance. J. Clin. Pharm. Ther. 1999, 24, 339–346. [Google Scholar] [CrossRef] [PubMed]
- Özdemir, B.C.; Csajka, C.; Dotto, G.P.; Wagner, A.D. Sex Differences in Efficacy and Toxicity of Systemic Treatments: An Undervalued Issue in the Era of Precision Oncology. J. Clin. Oncol. 2018, 36, 2680–2683. [Google Scholar] [CrossRef]
- Joerger, M.; Huitema, A.D.R.; van den Bongard, D.H.J.G.; Schellens, J.H.M.; Beijnen, J.H. Quantitative Effect of Gender, Age, Liver Function, and Body Size on the Population Pharmacokinetics of Paclitaxel in Patients with Solid Tumors. Clin. Cancer Res. 2006, 12, 2150–2157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schrijvers, D. Role of Red Blood Cells in Pharmacokinetics of Chemotherapeutic Agents. Clin. Pharmacokinet. 2003, 42, 779–791. [Google Scholar] [CrossRef]
- Murphy, W.G. The Sex Difference in Haemoglobin Levels in Adults—Mechanisms, Causes, and Consequences. Blood Rev. 2014, 28, 41–47. [Google Scholar] [CrossRef]
- Maloney, S.M.; Hoover, C.A.; Morejon-Lasso, L.V.; Prosperi, J.R. Mechanisms of Taxane Resistance. Cancers 2020, 12, 3323. [Google Scholar] [CrossRef]
- Thornton, L.M.; Carson, W.E.; Shapiro, C.L.; Farrar, W.B.; Andersen, B.L. Delayed Emotional Recovery after Taxane-Based Chemotherapy. Cancer 2008, 113, 638–647. [Google Scholar] [CrossRef] [Green Version]
- Wheatley-Price, P.; Le Maître, A.; Ding, K.; Leighl, N.; Hirsh, V.; Seymour, L.; Bezjak, A.; Shepherd, F.A. The Influence of Sex on Efficacy, Adverse Events, Quality of Life, and Delivery of Treatment in National Cancer Institute of Canada Clinical Trials Group Non-Small Cell Lung Cancer Chemotherapy Trials. J. Thorac. Oncol. 2010, 5, 640–648. [Google Scholar] [CrossRef] [Green Version]
- Cristina, V.; Mahachie, J.; Mauer, M.; Buclin, T.; van Cutsem, E.; Roth, A.; Wagner, A.D. Association of Patient Sex with Chemotherapy-Related Toxic Effects: A Retrospective Analysis of the PETACC-3 Trial Conducted by the EORTC Gastrointestinal Group. JAMA Oncol. 2018, 4, 1003–1006. [Google Scholar] [CrossRef]
- Miller, K.D.; Nogueira, L.; Mariotto, A.B.; Rowland, J.H.; Yabroff, K.R.; Alfano, C.M.; Jemal, A.; Kramer, J.L.; Siegel, R.L. Cancer Treatment and Survivorship Statistics, 2019. CA Cancer J. Clin. 2019, 69, 363–385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer Statistics, 2017. CA Cancer J. Clin. 2017, 67, 7–30. [Google Scholar] [CrossRef] [Green Version]
- Klein, S.L.; Flanagan, K.L. Sex Differences in Immune Responses. Nat. Rev. Immunol. 2016, 16, 626–638. [Google Scholar] [CrossRef] [PubMed]
- Márquez, E.J.; Chung, C.; Marches, R.; Rossi, R.J.; Nehar-Belaid, D.; Eroglu, A.; Mellert, D.J.; Kuchel, G.A.; Banchereau, J.; Ucar, D. Sexual-Dimorphism in Human Immune System Aging. Nat. Commun. 2020, 11, 751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roved, J.; Westerdahl, H.; Hasselquist, D. Sex Differences in Immune Responses: Hormonal Effects, Antagonistic Selection, and Evolutionary Consequences. Horm. Behav. 2017, 88, 95–105. [Google Scholar] [CrossRef]
- Weiss, R.B.; Donehower, R.C.; Wiernik, P.H.; Ohnuma, T.; Gralla, R.J.; Trump, D.L.; Baker, R., Jr.; van Echo, D.A.; von Hoff, D.D.; Leyland-Jones, B. Hypersensitivity Reactions from Taxol. J. Clin. Oncol. 2016, 8, 1263–1268. [Google Scholar] [CrossRef]
- Castells, M.C.; Tennant, N.M.; Sloane, D.E.; Hsu, F.I.; Barrett, N.A.; Hong, D.I.; Laidlaw, T.M.; Legere, H.J.; Nallamshetty, S.N.; Palis, R.I.; et al. Hypersensitivity Reactions to Chemotherapy: Outcomes and Safety of Rapid Desensitization in 413 Cases. J. Allergy Clin. Immunol. 2008, 122, 574–580. [Google Scholar] [CrossRef]
- Limsuwan, T.; Castells, M. Outcomes and Safety of Rapid Desensitization for Chemotherapy Hypersensitivity. Expert Opin. Drug Saf. 2010, 9, 39–53. [Google Scholar] [CrossRef]
- Picard, M.; Castells, M.C. Re-Visiting Hypersensitivity Reactions to Taxanes: A Comprehensive Review. Clin. Rev. Allergy Immunol. 2014, 49, 177–191. [Google Scholar] [CrossRef]
- Lansinger, O.M.; Biedermann, S.; He, Z.; Colevas, A.D. Do Steroids Matter? A Retrospective Review of Premedication for Taxane Chemotherapy and Hypersensitivity Reactions. J. Clin. Oncol. 2021, 39, 3583–3590. [Google Scholar] [CrossRef]
- Kemper, E.M.; van Zandbergen, A.E.; Cleypool, C.; Mos, H.A.; Boogerd, W.; Beijnen, J.H.; van Tellingen, O. Increased Penetration of Paclitaxel into the Brain by Inhibition of P-Glycoprotein. Clin. Cancer Res. 2003, 9, 2849–2855. [Google Scholar]
- Balayssac, D.; Cayre, A.; Authier, N.; Bourdu, S.; Penault-Llorca, F.; Gillet, J.P.; Maublant, J.; Eschalier, A.; Coudore, F. Patterns of P-Glycoprotein Activity in the Nervous System during Vincristine-Induced Neuropathy in Rats. J. Peripher. Nerv. Syst. 2005, 10, 301–310. [Google Scholar] [CrossRef]
- Cisternino, S.; Bourasset, F.; Archimbaud, Y.; Sémiond, D.; Sanderink, G.; Scherrmann, J.-M. Nonlinear Accumulation in the Brain of the New Taxoid TXD258 Following Saturation of P-Glycoprotein at the Blood–Brain Barrier in Mice and Rats. Br. J. Pharmacol. 2003, 138, 1367–1375. [Google Scholar] [CrossRef] [Green Version]
- Huehnchen, P.; Boehmerle, W.; Springer, A.; Freyer, D.; Endres, M. A Novel Preventive Therapy for Paclitaxel-Induced Cognitive Deficits: Preclinical Evidence from C57BL/6 Mice. Transl. Psychiatry 2017, 7, e1185. [Google Scholar] [CrossRef] [Green Version]
- Montague, K.; Malcangio, M. The Therapeutic Potential of Monocyte/Macrophage Manipulation in the Treatment of Chemotherapy-Induced Painful Neuropathy. Front. Mol. Neurosci. 2017, 10, 397. [Google Scholar] [CrossRef]
- da Costa, R.; Passos, G.F.; Quintão, N.L.M.; Fernandes, E.S.; Maia, J.R.L.C.B.; Campos, M.M.; Calixto, J.B. Taxane-Induced Neurotoxicity: Pathophysiology and Therapeutic Perspectives. Br. J. Pharmacol. 2020, 177, 3127–3146. [Google Scholar] [CrossRef] [PubMed]
- Scripture, C.D.; Figg, W.D.; Sparreboom, A. Peripheral Neuropathy Induced by Paclitaxel: Recent Insights and Future Perspectives. Curr. Neuropharmacol. 2006, 4, 165–172. [Google Scholar] [CrossRef] [Green Version]
- Tofthagen, C.; McAllister, R.D.; Visovsky, C. Peripheral Neuropathy Caused by Paclitaxel and Docetaxel: An Evaluation and Comparison of Symptoms. J. Adv. Pract. Oncol. 2013, 4, 204–215. [Google Scholar] [CrossRef]
- Augusto, C.; Pietro, M.; Cinzia, M.; Sergio, C.; Sara, C.; Luca, G.; Scaioli, V. Peripheral Neuropathy Due to Paclitaxel: Study of the Temporal Relationships between the Therapeutic Schedule and the Clinical Quantitative Score (QST) and Comparison with Neurophysiological Findings. J. Neuro-Oncol. 2008, 86, 89–99. [Google Scholar] [CrossRef]
- Sarosy, G.; Kohn, E.; Stone, D.A.; Rothenberg, M.; Jacob, J.; Adamo, D.O.; Ognibene, F.P.; Cunnion, R.E.; Reed, E. Phase I Study of Taxol and Granulocyte Colony-Stimulating Factor in Patients with Refractory Ovarian Cancer. J. Clin. Oncol. 1992, 10, 1165–1170. [Google Scholar] [CrossRef]
- Seretny, M.; Currie, G.L.; Sena, E.S.; Ramnarine, S.; Grant, R.; Macleod, M.R.; Colvin, L.A.; Fallon, M. Incidence, Prevalence, and Predictors of Chemotherapy-Induced Peripheral Neuropathy: A Systematic Review and Meta-Analysis. Pain® 2014, 155, 2461–2470. [Google Scholar] [CrossRef] [Green Version]
- Tanabe, Y.; Hashimoto, K.; Shimizu, C.; Hirakawa, A.; Harano, K.; Yunokawa, M.; Yonemori, K.; Katsumata, N.; Tamura, K.; Ando, M.; et al. Paclitaxel-Induced Peripheral Neuropathy in Patients Receiving Adjuvant Chemotherapy for Breast Cancer. Int. J. Clin. Oncol. 2011, 18, 132–138. [Google Scholar] [CrossRef] [PubMed]
- Nakai, Y.; Tanaka, N.; Ichikawa, K.; Miyake, M.; Anai, S.; Fujimoto, K. Appropriate Number of Docetaxel Cycles in Castration-Resistant Prostate Cancer Patients Considering Peripheral Neuropathy and Oncological Control. Chemotherapy 2020, 65, 119–124. [Google Scholar] [CrossRef] [PubMed]
- Winters-Stone, K.M.; Horak, F.; Jacobs, P.G.; Trubowitz, P.; Dieckmann, N.F.; Stoyles, S.; Faithfull, S. Falls, Functioning, and Disability Among Women with Persistent Symptoms of Chemotherapy-Induced Peripheral Neuropathy. J. Clin. Oncol. 2017, 35, 2604–2612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kober, K.M.; Mazor, M.; Abrams, G.; Olshen, A.; Conley, Y.P.; Hammer, M.; Schumacher, M.; Chesney, M.; Smoot, B.; Mastick, J.; et al. Phenotypic Characterization of Paclitaxel-Induced Peripheral Neuropathy in Cancer Survivors. J. Pain Symptom Manag. 2018, 56, 908–919.e3. [Google Scholar] [CrossRef]
- Windebank, A.J.; Grisold, W. Chemotherapy-Induced Neuropathy. J. Peripher. Nerv. Syst. 2008, 13, 27–46. [Google Scholar] [CrossRef]
- Weimer, L.H. Medication-Induced Peripheral Neuropathy. Curr. Neurol. Neurosci. Rep. 2003, 3, 86–92. [Google Scholar] [CrossRef]
- Mogil, J.S. Sex Differences in Pain and Pain Inhibition: Multiple Explanations of a Controversial Phenomenon. Nat. Rev. Neurosci. 2012, 13, 859–866. [Google Scholar] [CrossRef]
- Vallejo, R.; Tilley, D.M.; Vogel, L.; Benyamin, R. The Role of Glia and the Immune System in the Development and Maintenance of Neuropathic Pain. Pain Pract. 2010, 10, 167–184. [Google Scholar] [CrossRef]
- Boyette-Davis, J.; Xin, W.; Zhang, H.; Dougherty, P.M. Intraepidermal Nerve Fiber Loss Corresponds to the Development of Taxol-Induced Hyperalgesia and Can Be Prevented by Treatment with Minocycline. Pain® 2011, 152, 308–313. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, H.; Zhang, H.; Kosturakis, A.K.; Jawad, A.B.; Dougherty, P.M. Toll-Like Receptor 4 Signaling Contributes to Paclitaxel-Induced Peripheral Neuropathy. J. Pain 2014, 15, 712–725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosen, S.; Ham, B.; Mogil, J.S. Sex Differences in Neuroimmunity and Pain. J. Neurosci. Res. 2017, 95, 500–508. [Google Scholar] [CrossRef] [PubMed]
- Gregus, A.M.; Levine, I.S.; Eddinger, K.A.; Yaksh, T.L.; Buczynski, M.W. Sex Differences in Neuroimmune and Glial Mechanisms of Pain. Pain® 2021, 162, 2186–2200. [Google Scholar] [CrossRef]
- Sorge, R.E.; Mapplebeck, J.C.S.; Rosen, S.; Beggs, S.; Taves, S.; Alexander, J.K.; Martin, L.J.; Austin, J.-S.; Sotocinal, S.G.; Chen, D.; et al. Different Immune Cells Mediate Mechanical Pain Hypersensitivity in Male and Female Mice. Nat. Neurosci. 2015, 18, 1081–1083. [Google Scholar] [CrossRef] [Green Version]
- Luo, X.; Huh, Y.; Bang, S.; He, Q.; Zhang, L.; Matsuda, M.; Ji, R.-R. Macrophage Toll-like Receptor 9 Contributes to Chemotherapy-Induced Neuropathic Pain in Male Mice. J. Neurosci. 2019, 39, 6848–6864. [Google Scholar] [CrossRef]
- Wang, Y.; Li, N.; Zhao, Y.; Zhang, L.J. Effects of Female Sex Hormones on Chemotherapeutic Paclitaxel-Induced Neuropathic Pain and Involvement of Inflammatory Signal. J. Biol. Regul. Homeost. Agents 2018, 32, 1157–1163. [Google Scholar]
- Bale, T.L.; Epperson, C.N. Sex Differences and Stress across the Lifespan. Nat. Neurosci. 2015, 18, 1413–1420. [Google Scholar] [CrossRef]
- Ferrari, L.F.; Araldi, D.; Green, P.; Levine, J.D. Marked Sexual Dimorphism in Neuroendocrine Mechanisms for the Exacerbation of Paclitaxel-Induced Painful Peripheral Neuropathy by Stress. Pain 2020, 161, 865–874. [Google Scholar] [CrossRef]
- Luo, X.; Gu, Y.; Tao, X.; Serhan, C.N.; Ji, R.-R. Resolvin D5 Inhibits Neuropathic and Inflammatory Pain in Male but Not Female Mice: Distinct Actions of D-Series Resolvins in Chemotherapy-Induced Peripheral Neuropathy. Front. Pharmacol. 2019, 10, 745. [Google Scholar] [CrossRef]
- Lopes, D.M.; Malek, N.; Edye, M.; Jager, S.B.; McMurray, S.; McMahon, S.B.; Denk, F. Sex Differences in Peripheral Not Central Immune Responses to Pain-Inducing Injury. Sci. Rep. 2017, 7, 16460. [Google Scholar] [CrossRef] [Green Version]
- Parvathy, S.S.; Masocha, W. Coadministration of Indomethacin and Minocycline Attenuates Established Paclitaxel-Induced Neuropathic Thermal Hyperalgesia: Involvement of Cannabinoid CB1 Receptors. Sci. Rep. 2015, 5, 10541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Natri, H.; Garcia, A.R.; Buetow, K.H.; Trumble, B.C.; Wilson, M.A. The Pregnancy Pickle: Evolved Immune Compensation Due to Pregnancy Underlies Sex Differences in Human Diseases. Trends Genet. 2019, 35, 478–488. [Google Scholar] [CrossRef] [PubMed]
- Dietrich, J.; Kaiser, J. Cancer, Chemotherapy and Cognitive Dysfunction. US Neurol. 2016, 12, 43–45. [Google Scholar] [CrossRef] [Green Version]
- Vitali, M.; Ripamonti, C.I.; Roila, F.; Proto, C.; Signorelli, D.; Imbimbo, M.; Corrao, G.; Brissa, A.; Rosaria, G.; de Braud, F.; et al. Cognitive Impairment and Chemotherapy: A Brief Overview. Crit. Rev. Oncol. Hematol. 2017, 118, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Myers, J.S. Chemotherapy-Related Cognitive Impairment: The Breast Cancer Experience. Oncol. Nurs. Forum 2012, 39, E31–E40. [Google Scholar] [CrossRef] [Green Version]
- Loh, K.P.; Janelsins, M.C.; Mohile, S.G.; Holmes, H.M.; Hsu, T.; Inouye, S.K.; Karuturi, M.S.; Kimmick, G.G.; Lichtman, S.M.; Magnuson, A.; et al. Chemotherapy-Related Cognitive Impairment in Older Patients with Cancer. J. Geriatr. Oncol. 2016, 7, 270–280. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Caeyenberghs, K. Longitudinal Assessment of Chemotherapy-Induced Changes in Brain and Cognitive Functioning: A Systematic Review. Neurosci. Biobehav. Rev. 2018, 92, 304–317. [Google Scholar] [CrossRef]
- Argyriou, A.A.; Assimakopoulos, K.; Iconomou, G.; Giannakopoulou, F.; Kalofonos, H.P. Either Called “Chemobrain” or “Chemofog,” the Long-Term Chemotherapy-Induced Cognitive Decline in Cancer Survivors Is Real. J. Pain Symptom Manag. 2011, 41, 126–139. [Google Scholar] [CrossRef]
- Janelsins, M.C.; Heckler, C.E.; Peppone, L.J.; Kamen, C.; Mustian, K.M.; Mohile, S.G.; Magnuson, A.; Kleckner, I.R.; Guido, J.J.; Young, K.L.; et al. Cognitive Complaints in Survivors of Breast Cancer after Chemotherapy Compared with Age-Matched Controls: An Analysis from a Nationwide, Multicenter, Prospective Longitudinal Study. J. Clin. Oncol. Am. Soc. Clin. Oncol. 2017, 35, 506–514. [Google Scholar] [CrossRef]
- Liang, L.; Wei, J.; Tian, L.; Nagendra, B.V.P.; Gao, F.; Zhang, J.; Xu, L.; Wang, H.; Huo, F.Q. Paclitaxel Induces Sex-Biased Behavioral Deficits and Changes in Gene Expression in Mouse Prefrontal Cortex. Neuroscience 2020, 426, 168–178. [Google Scholar] [CrossRef]
- Chmielewski-Stivers, N.; Petit, B.; Ollivier, J.; Monceau, V.; Tsoutsou, P.; Pousa, A.Q.; Lin, X.; Limoli, C.; Vozenin, M.-C. Sex-Specific Differences in Toxicity Following Systemic Paclitaxel Treatment and Localized Cardiac Radiotherapy. Cancers 2021, 13, 3973. [Google Scholar] [CrossRef] [PubMed]
- Hausmann, M. Why Sex Hormones Matter for Neuroscience: A Very Short Review on Sex, Sex Hormones, and Functional Brain Asymmetries. J. Neurosci. Res. 2017, 95, 40–49. [Google Scholar] [CrossRef] [Green Version]
- Gaignard, P.; Liere, P.; Thérond, P.; Schumacher, M.; Slama, A.; Guennoun, R. Role of Sex Hormones on Brain Mitochondrial Function, with Special Reference to Aging and Neurodegenerative Diseases. Front. Aging Neurosci. 2017, 9, 406. [Google Scholar] [CrossRef]
- Médale-Giamarchi, C.; Lajoie-Mazenc, I.; Malissein, E.; Meunier, E.; Couderc, B.; Bergé, Y.; Filleron, T.; Keller, L.; Marty, C.; Lacroix-Triki, M.; et al. RhoB Modifies Estrogen Responses in Breast Cancer Cells by Influencing Expression of the Estrogen Receptor. Breast Cancer Res. 2013, 15, R6. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Lian, C.L.; Zheng, H.; Lin, L.E.; Yu, Y.F.; Lin, Q.; Wu, S.G. Cognitive Dysfunction in Patients with Nasopharyngeal Carcinoma after Induction Chemotherapy. Oral Oncol. 2020, 111, 104921. [Google Scholar] [CrossRef] [PubMed]
- Cerulla, N.; Arcusa, À.; Navarro, J.-B.; Garolera, M.; Enero, C.; Chico, G.; Fernández-Morales, L. Role of Taxanes in Chemotherapy-Related Cognitive Impairment: A Prospective Longitudinal Study. Breast Cancer Res. Treat. 2017, 164, 179–187. [Google Scholar] [CrossRef] [PubMed]
- Wefel, J.S.; Saleeba, A.K.; Buzdar, A.U.; Meyers, C.A. Acute and Late Onset Cognitive Dysfunction Associated with Chemotherapy in Women with Breast Cancer. Cancer 2010, 116, 3348–3356. [Google Scholar] [CrossRef] [Green Version]
- Brown, T.; McElroy, T.; Simmons, P.; Walters, H.; Ntagwabira, F.; Wang, J.; Byrum, S.D.; Allen, A.R. Cognitive Impairment Resulting from Treatment with Docetaxel, Doxorubicin, and Cyclophosphamide. Brain Res. 2021, 1760, 147397. [Google Scholar] [CrossRef]
- Alibhai, S.M.H.; Breunis, H.; Timilshina, N.; Hansen, A.R.; Joshua, A.M.; Warde, P.R.; Gregg, R.W.; Fleshner, N.E.; Tomlinson, G.; Hotte, S.J.; et al. The Effect of Docetaxel, Enzalutamide, Abiraterone, and Radium-223 on Cognitive Function in Older Men with Metastatic Castrate-Resistant Prostate Cancer (MCRPC). J. Clin. Oncol. 2020, 38, 73. [Google Scholar] [CrossRef]
- Lange, M.; Heutte, N.; Rigal, O.; Noal, S.; Kurtz, J.-E.; Lévy, C.; Allouache, D.; Rieux, C.; Lefel, J.; Clarisse, B.; et al. Decline in Cognitive Function in Older Adults with Early-Stage Breast Cancer After Adjuvant Treatment. Oncologist 2016, 21, 1337–1348. [Google Scholar] [CrossRef] [Green Version]
- Fardell, J.E.; Zhang, J.; de Souza, R.; Vardy, J.; Johnston, I.; Allen, C.; Henderson, J.; Piquette-Miller, M. The Impact of Sustained and Intermittent Docetaxel Chemotherapy Regimens on Cognition and Neural Morphology in Healthy Mice. Psychopharmacology 2013, 231, 841–852. [Google Scholar] [CrossRef] [PubMed]
- Seigers, R.; Loos, M.; van Tellingen, O.; Boogerd, W.; Smit, A.B.; Schagen, S.B. Cognitive Impact of Cytotoxic Agents in Mice. Psychopharmacology 2014, 232, 17–37. [Google Scholar] [CrossRef] [PubMed]
- Callaghan, C.K.; O’Mara, S.M. Long-Term Cognitive Dysfunction in the Rat Following Docetaxel Treatment Is Ameliorated by the Phosphodiesterase-4 Inhibitor, Rolipram. Behav. Brain Res. 2015, 290, 84–89. [Google Scholar] [CrossRef]
- Panoz-Brown, D.; Carey, L.M.; Smith, A.E.; Gentry, M.; Sluka, C.M.; Corbin, H.E.; Wu, J.E.; Hohmann, A.G.; Crystal, J.D. The Chemotherapeutic Agent Paclitaxel Selectively Impairs Reversal Learning While Sparing Prior Learning, New Learning and Episodic Memory. Neurobiol. Learn. Mem. 2017, 144, 259–270. [Google Scholar] [CrossRef] [Green Version]
- Office of Science Policy. Significant Research Advances Enabled by HeLa Cells. Available online: https://osp.od.nih.gov/scientific-sharing/hela-cells-timeline/ (accessed on 16 August 2021).
- Horwitz, S.B.; Goldman, I.D. A Conversation with Susan Band Horwitz. Annu. Rev. Pharmacol. Toxicol. 2015, 55, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Brabander, M.; Geuens, G.; Nuydens, R.; Willebrords, R.; de Mey, J. Taxol Induces the Assembly of Free Microtubules in Living Cells and Blocks the Organizing Capcity of the Centrosomes and Kinetochores. Proc. Natl. Acad. Sci. USA 1981, 78, 5608–5612. [Google Scholar] [CrossRef] [Green Version]
- Shi, J.; Orth, J.D.; Mitchison, T. Cell Type Variation in Responses to Antimitotic Drugs That Target Microtubules and Kinesin-5. Cancer Res. 2008, 68, 3269–3276. [Google Scholar] [CrossRef] [Green Version]
- Milas, L.; Hunter, N.R.; Kurdoglu, B.; Mason, K.A.; Meyn, R.E.; Stephens, L.C.; Peters, L.J. Kinetics of Mitotic Arrest and Apoptosis in Murine Mammary and Ovarian Tumors Treated with Taxol. Cancer Chemother. Pharmacol. 1995, 35, 297–303. [Google Scholar] [CrossRef]
- Waters, J.C.; Chen, R.H.; Murray, A.W.; Salmon, E.D. Localization of Mad2 to Kinetochores Depends on Microtubule Attachment, Not Tension. J. Cell Biol. 1998, 141, 1181–1191. [Google Scholar] [CrossRef] [Green Version]
- Sudo, T.; Nitta, M.; Saya, H.; Ueno, N.T. Dependence of Paclitaxel Sensitivity on a Functional Spindle Assembly Checkpoint. Cancer Res. 2004, 64, 2502–2508. [Google Scholar] [CrossRef] [Green Version]
- Komlodi-Pasztor, E.; Sackett, D.; Wilkerson, J.; Fojo, T. Mitosis Is Not a Key Target of Microtubule Agents in Patient Tumors. Nat. Rev. Clin. Oncol. 2011, 8, 244–250. [Google Scholar] [CrossRef]
- Mitchison, T.J. The Proliferation Rate Paradox in Antimitotic Chemotherapy. Mol. Biol. Cell 2012, 23, 1–6. [Google Scholar] [CrossRef]
- Milross, C.G.; Mason, K.A.; Hunter, N.R.; Chung, W.K.; Peters, L.J.; Milas, L. Relationship of Mitotic Arrest and Apoptosis to Antitumor Effect of Paclitaxel. J. Natl. Cancer Inst. 1996, 88, 1308–1314. [Google Scholar] [CrossRef]
- Chakravarty, A.; Shinde, V.; Tabernero, J.; Cervantes, A.; Cohen, R.B.; Dees, E.C.; Burris, H.; Infante, J.R.; Macarulla, T.; Elez, E.; et al. Phase I Assessment of New Mechanism-Based Pharmacodynamic Biomarkers for MLN8054, a Small-Molecule Inhibitor of Aurora A Kinase. Cancer Res. 2011, 71, 675–685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Florian, S.; Mitchison, T.J. Anti-Microtubule Drugs. In The Mitotic Spindle. Methods in Molecular Biology; Chang, P., Ohi, R., Eds.; Humana Press Inc.: New York, NY, USA, 2016. [Google Scholar] [CrossRef]
- Gundersen, G.G.; Cook, T.A. Microtubules and Signal Transduction. Curr. Opin. Cell Biol. 1999, 11, 81–94. [Google Scholar] [CrossRef]
- Etienne-Manneville, S. From Signaling Pathways to Microtubule Dynamics: The Key Players. Curr. Opin. Cell Biol. 2010, 22, 104–111. [Google Scholar] [CrossRef] [PubMed]
- Zasadil, L.M.; Andersen, K.A.; Yeum, D.; Rocque, G.B.; Wilke, L.G.; Tevaarwerk, A.J.; Raines, R.T.; Burkard, M.E.; Weaver, B.A. Cytotoxicity of Paclitaxel in Breast Cancer Is Due to Chromosome Missegregation on Multipolar Spindles. Sci. Transl. Med. 2014, 6, 229ra43. [Google Scholar] [CrossRef] [Green Version]
- Thadani-Mulero, M.; Nanus, D.M.; Giannakakou, P. Androgen Receptor on the Move: Boarding the Microtubule Expressway to the Nucleus. Cancer Res. 2012, 72, 4611–4615. [Google Scholar] [CrossRef] [Green Version]
- Carbonaro, M.; Escuin, D.; O’Brate, A.; Thadani-Mulero, M.; Giannakakou, P. Microtubules Regulate Hypoxia-Inducible Factor-1α Protein Trafficking and Activity: Implications for Taxane Therapy. J. Biol. Chem. 2012, 287, 11859–11869. [Google Scholar] [CrossRef] [Green Version]
- Darshan, M.S.; Loftus, M.S.; Thadani-Mulero, M.; Levy, B.P.; Escuin, D.; Zhou, X.K.; Gjyrezi, A.; Chanel-Vos, C.; Shen, R.; Tagawa, S.T.; et al. Taxane-Induced Blockade to Nuclear Accumulation of the Androgen Receptor Predicts Clinical Responses in Metastatic Prostate Cancer. Cancer Res. 2011, 71, 6019–6029. [Google Scholar] [CrossRef] [Green Version]
- Jordan, M.A.; Wendell, K.; Gardiner, S.; Brent Derry, W.; Copp, H.; Wilson, L. Mitotic Block Induced in HeLa Cells by Low Concentrations of Paclitaxel (Taxol) Results in Abnormal Mitotic Exit and Apoptotic Cell Death. Cancer Res. 1996, 56, 816–825. [Google Scholar] [PubMed]
- Torres, K.; Horwitz, S.B. Mechanisms of Taxol-Induced Cell Death Are Concentration Dependent. Cancer Res. 1998, 58, 3620–3626. [Google Scholar] [PubMed]
- Jordan, M.A.; Toso, R.J.; Thrower, D.; Wilson, L. Mechanism of Mitotic Block and Inhibition of Cell Proliferation by Taxol at Low Concentrations. Proc. Natl. Acad. Sci. USA 1993, 90, 9552–9556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ikui, A.E.; Chia-Ping, H.Y.; Matsumoto, T.; Horwitz, S.B. Low Concentrations of Taxol Cause Mitotic Delay Followed by Premature Dissociation of P55CDC from Mad2 and BubR1 and Abrogation of the Spindle Checkpoint, Leading to Aneuploidy. Cell Cycle 2005, 4, 1385–1388. [Google Scholar] [CrossRef] [Green Version]
- Zhu, M.-L.; Horbinski, C.M.; Garzotto, M.; Qian, D.Z.; Beer, T.M.; Kyprianou, N. Tubulin-Targeting Chemotherapy Impairs Androgen Receptor Activity in Prostate Cancer. Cancer Res. 2010, 70, 7992–8002. [Google Scholar] [CrossRef] [Green Version]
- Feldman, B.J.; Feldman, D. The Development of Androgen-Independent Prostate Cancer. Nat. Rev. Cancer 2001, 1, 34–45. [Google Scholar] [CrossRef] [PubMed]
- Watson, P.A.; Arora, V.K.; Sawyers, C.L. Emerging Mechanisms of Resistance to Androgen Receptor Inhibitors in Prostate Cancer. Nat. Rev. Cancer 2015, 15, 701–711. [Google Scholar] [CrossRef] [Green Version]
- Attard, G.; Richards, J.; de Bono, J.S. New Strategies in Metastatic Prostate Cancer: Targeting the Androgen Receptor Signaling Pathway. Clin. Cancer Res. 2011, 17, 1649–1657. [Google Scholar] [CrossRef] [Green Version]
- Antonarakis, E.S.; Tagawa, S.T.; Galletti, G.; Worroll, D.; Ballman, K.; Vanhuyse, M.; Sonpavde, G.; North, S.; Albany, C.; Tsao, C.-K.; et al. Randomized, Noncomparative, Phase II Trial of Early Switch from Docetaxel to Cabazitaxel or Vice Versa, with Integrated Biomarker Analysis, in Men with Chemotherapy-Naïve, Metastatic, Castration-Resistant Prostate Cancer. J. Clin. Oncol. 2017, 35, 3181–3188. [Google Scholar] [CrossRef] [Green Version]
- de Bono, J.S.; Oudard, S.; Ozguroglu, M.; Hansen, S.; Machiels, J.-P.; Kocak, I.; Gravis, G.; Bodrogi, I.; Mackenzie, M.J.; Shen, L.; et al. Prednisone plus Cabazitaxel or Mitoxantrone for Metastatic Castration-Resistant Prostate Cancer Progressing after Docetaxel Treatment: A Randomised Open-Label Trial. Lancet 2010, 376, 1147–1154. [Google Scholar] [CrossRef]
- Petrylak, D.P.; Tangen, C.M.; Hussain, M.H.A.; Lara, P.N.; Jones, J.A.; Taplin, M.E.; Burch, P.A.; Berry, D.; Moinpour, C.; Kohli, M.; et al. Docetaxel and Estramustine Compared with Mitoxantrone and Prednisone for Advanced Refractory Prostate Cancer. N. Engl. J. Med. 2004, 351, 1513–1520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tannock, I.F.; de Wit, R.; Berry, W.R.; Horti, J.; Pluzanska, A.; Chi, K.N.; Oudard, S.; Théodore, C.; James, N.D.; Turesson, I.; et al. Docetaxel plus Prednisone or Mitoxantrone plus Prednisone for Advanced Prostate Cancer. N. Engl. J. Med. 2004, 351, 1502–1512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuroda, K.; Liu, H.; Kim, S.; Guo, M.; Navarro, V.; Bander, N.H. Docetaxel Down-Regulates the Expression of Androgen Receptor and Prostate-Specific Antigen but Not Prostate-Specific Membrane Antigen in Prostate Cancer Cell Lines: Implications for PSA Surrogacy. Prostate 2009, 69, 1579–1585. [Google Scholar] [CrossRef]
- Gan, L.; Chen, S.; Wang, Y.; Watahiki, A.; Bohrer, L.; Sun, Z.; Wang, Y.; Huang, H. Inhibition of the Androgen Receptor as a Novel Mechanism of Taxol Chemotherapy in Prostate Cancer. Cancer Res. 2009, 69, 8386–8394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kampa, M.; Kogia, C.; Theodoropoulos, P.A.; Anezinis, P.; Charalampopoulos, I.; Papakonstanti, E.A.; Stathopoulos, E.N.; Hatzoglou, A.; Stournaras, C.; Gravanis, A.; et al. Activation of Membrane Androgen Receptors Potentiates the Antiproliferative Effects of Paclitaxel on Human Prostate Cancer Cells. Mol. Cancer Ther. 2006, 5, 1342–1351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lai, J.-J.; Lai, K.-P.; Zeng, W.; Chuang, K.-H.; Altuwaijri, S.; Chang, C. Androgen Receptor Influences on Body Defense System via Modulation of Innate and Adaptive Immune Systems: Lessons from Conditional AR Knockout Mice. Am. J. Pathol. 2012, 181, 1504–1512. [Google Scholar] [CrossRef] [Green Version]
- Yeh, S.; Tsai, M.-Y.; Xu, Q.; Mu, X.-M.; Lardy, H.; Huang, K.-E.; Lin, H.; Yeh, S.-D.; Altuwaijri, S.; Zhou, X.; et al. Generation and Characterization of Androgen Receptor Knockout (ARKO) Mice: An in Vivo Model for the Study of Androgen Functions in Selective Tissues. Proc. Natl. Acad. Sci. USA 2002, 99, 13498–13503. [Google Scholar] [CrossRef] [Green Version]
- Kerkhofs, S.; Denayer, S.; Haelens, A.; Claessens, F. Androgen Receptor Knockout and Knock-in Mouse Models. J. Mol. Endocrinol. 2009, 42, 11–17. [Google Scholar] [CrossRef] [Green Version]
- Venken, K.; de Gendt, K.; Boonen, S.; Ophoff, J.; Bouillon, R.; Swinnen, J.V.; Verhoeven, G.; Vanderschueren, D. Relative Impact of Androgen and Estrogen Receptor Activation in the Effects of Androgens on Trabecular and Cortical Bone in Growing Male Mice: A Study in the Androgen Receptor Knockout Mouse Model. J. Bone Miner. Res. 2006, 21, 576–585. [Google Scholar] [CrossRef]
- Komura, K.; Jeong, S.H.; Hinohara, K.; Qu, F.; Wang, X.; Hiraki, M.; Azuma, H.; Lee, G.S.M.; Kantoff, P.W.; Sweeney, C.J. Resistance to Docetaxel in Prostate Cancer Is Associated with Androgen Receptor Activation and Loss of KDM5D Expression. Proc. Natl. Acad. Sci. USA 2016, 113, 6259–6264. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Chi, P.; Rockowitz, S.; Iaquinta, P.J.; Shamu, T.; Shukla, S.; Gao, D.; Sirota, I.; Carver, B.S.; Wongvipat, J.; et al. ETS Factors Reprogram the Androgen Receptor Cistrome and Prime Prostate Tumorigenesis in Response to PTEN Loss. Nat. Med. 2013, 19, 1023–1029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, J.; Yu, J.; Mani, R.-S.; Cao, Q.; Brenner, C.J.; Cao, X.; Wang, X.; Wu, L.; Li, J.; Hu, M.; et al. An Integrated Network of Androgen Receptor, Polycomb, and TMPRSS2-ERG Gene Fusions in Prostate Cancer Progression. Cancer Cell 2010, 17, 443–454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galletti, G.; Matov, A.; Beltran, H.; Fontugne, J.; Miguel Mosquera, J.; Cheung, C.; MacDonald, T.Y.; Sung, M.; O’Toole, S.; Kench, J.G.; et al. ERG Induces Taxane Resistance in Castration-Resistant Prostate Cancer. Nat. Commun. 2014, 5, 5548. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.; Jamalruddin, M.A.B.; Au, C.C.; Mukhtar, E.; Portella, L.; Berger, A.; Worroll, D.; Rickman, D.S.; Nanus, D.M.; Giannakakou, P. AR-V7 Exhibits Non-Canonical Mechanisms of Nuclear Import and Chromatin Engagement in Castrate-Resistant Prostate Cancer. bioRxiv 2021. [Google Scholar] [CrossRef]
- Hu, R.; Dunn, T.A.; Wei, S.; Isharwal, S.; Veltri, R.W.; Humphreys, E.; Han, M.; Partin, A.W.; Vessella, R.L.; Isaacs, W.B.; et al. Ligand-Independent Androgen Receptor Variants Derived from Splicing of Cryptic Exons Signify Hormone-Refractory Prostate Cancer. Cancer Res. 2009, 69, 16–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hörnberg, E.; Ylitalo, E.B.; Crnalic, S.; Antti, H.; Stattin, P.; Widmark, A.; Bergh, A.; Wikström, P. Expression of Androgen Receptor Splice Variants in Prostate Cancer Bone Metastases Is Associated with Castration-Resistance and Short Survival. PLoS ONE 2011, 6, e19059. [Google Scholar] [CrossRef] [Green Version]
- Maughan, B.L.; Antonarakis, E.S. Clinical Relevance of Androgen Receptor Splice Variants in Castration-Resistant Prostate Cancer. Curr. Treat. Options Oncol. 2015, 16, 57. [Google Scholar] [CrossRef]
- Tagawa, S.T.; Antonarakis, E.S.; Gjyrezi, A.; Galletti, G.; Kim, S.; Worroll, D.; Stewart, J.; Zaher, A.; Szatrowski, T.P.; Ballman, K.V.; et al. Expression of AR-V7 and ARv567es in Circulating Tumor Cells Correlates with Outcomes to Taxane Therapy in Men with Metastatic Prostate Cancer Treated in TAXYNERGY. Clin. Cancer Res. 2019, 25, 1880–1888. [Google Scholar] [CrossRef] [Green Version]
- Rizzo, A.; Mollica, V.; Rosellini, M.; Marchetti, A.; Ricci, A.D.; Fiorentino, M.; Battelli, N.; Santoni, M.; Massari, F. Exploring the Association between Metastatic Sites and Androgen Receptor Splice Variant 7 (AR-V7) in Castration-Resistant Prostate Cancer Patients: A Meta-Analysis of Prospective Clinical Trials. Pathol. Res. Pract. 2021, 222, 153440. [Google Scholar] [CrossRef]
- Antonarakis, E.S.; Lu, C.; Wang, H.; Luber, B.; Nakazawa, M.; Roeser, J.C.; Chen, Y.; Mohammad, T.A.; Chen, Y.; Fedor, H.L.; et al. AR-V7 and Resistance to Enzalutamide and Abiraterone in Prostate Cancer. N. Engl. J. Med. 2014, 371, 1028–1038. [Google Scholar] [CrossRef] [Green Version]
- Kipp, J.L.; Ramirez, V.D. Estradiol and Testosterone Have Opposite Effects on Microtubule Polymerization. Neuroendocrinology 2003, 77, 258–272. [Google Scholar] [CrossRef] [PubMed]
- Butler, R.; Leigh, P.N.; Gallo, J.-M. Androgen-Induced up-Regulation of Tubulin Isoforms in Neuroblastoma Cells. J. Neurochem. 2001, 78, 854–861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lo, Y.C.; Cormier, O.; Liu, T.; Nettles, K.W.; Katzenellenbogen, J.A.; Stearns, T.; Altman, R.B. Pocket Similarity Identifies Selective Estrogen Receptor Modulators as Microtubule Modulators at the Taxane Site. Nat. Commun. 2019, 10, 1033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dougherty, M.K.; Schumaker, L.M.; Jordan, V.C.; Welshons, W.V.; Curran, E.M.; Ellis, M.J.; El-Ashry, D. Estrogen Receptor Expression and Sensitivity to Paclitaxel in Breast Cancer. Cancer Biol. Ther. 2004, 3, 460–467. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, T.; Iwasaki, A. Sex Differences in Immune Responses. Science 2021, 371, 347–348. [Google Scholar] [CrossRef]
- Takahashi, T.; Ellingson, M.K.; Wong, P.; Israelow, B.; Lucas, C.; Klein, J.; Silva, J.; Mao, T.; Oh, J.E.; Tokuyama, M.; et al. Sex Differences in Immune Responses That Underlie COVID-19 Disease Outcomes. Nature 2020, 588, 315–320. [Google Scholar] [CrossRef]
- Scully, E.P.; Haverfield, J.; Ursin, R.L.; Tannenbaum, C.; Klein, S.L. Considering How Biological Sex Impacts Immune Responses and COVID-19 Outcomes. Nat. Rev. Immunol. 2020, 20, 442–447. [Google Scholar] [CrossRef]
- Brodin, P. Immune Determinants of COVID-19 Disease Presentation and Severity. Nat. Med. 2021, 27, 28–33. [Google Scholar] [CrossRef]
- Jin, J.-M.; Bai, P.; He, W.; Wu, F.; Liu, X.-F.; Han, D.-M.; Liu, S.; Yang, J.-K. Gender Differences in Patients with COVID-19: Focus on Severity and Mortality. Front. Public Health 2020, 8, 152. [Google Scholar] [CrossRef]
- Peckham, H.; de Gruijter, N.M.; Raine, C.; Radziszewska, A.; Ciurtin, C.; Wedderburn, L.R.; Rosser, E.C.; Webb, K.; Deakin, C.T. Male Sex Identified by Global COVID-19 Meta-Analysis as a Risk Factor for Death and ITU Admission. Nat. Commun. 2020, 11, 6317. [Google Scholar] [CrossRef]
Figure 1.
Known taxane mechanism of action. Taxanes bind to the inner surface of microtubule filaments, on the β-tubulin globular protein of the heterodimer, and the protofilament tubule arrangement. Taxanes prevent microtubule depolymerization, preventing normal microtubule dynamics for cellular activities and impacting cytoskeletal structure and microtubule function, such as mitosis and microtubule-dependent cellular transport. Created with BioRender.
Figure 1.
Known taxane mechanism of action. Taxanes bind to the inner surface of microtubule filaments, on the β-tubulin globular protein of the heterodimer, and the protofilament tubule arrangement. Taxanes prevent microtubule depolymerization, preventing normal microtubule dynamics for cellular activities and impacting cytoskeletal structure and microtubule function, such as mitosis and microtubule-dependent cellular transport. Created with BioRender.
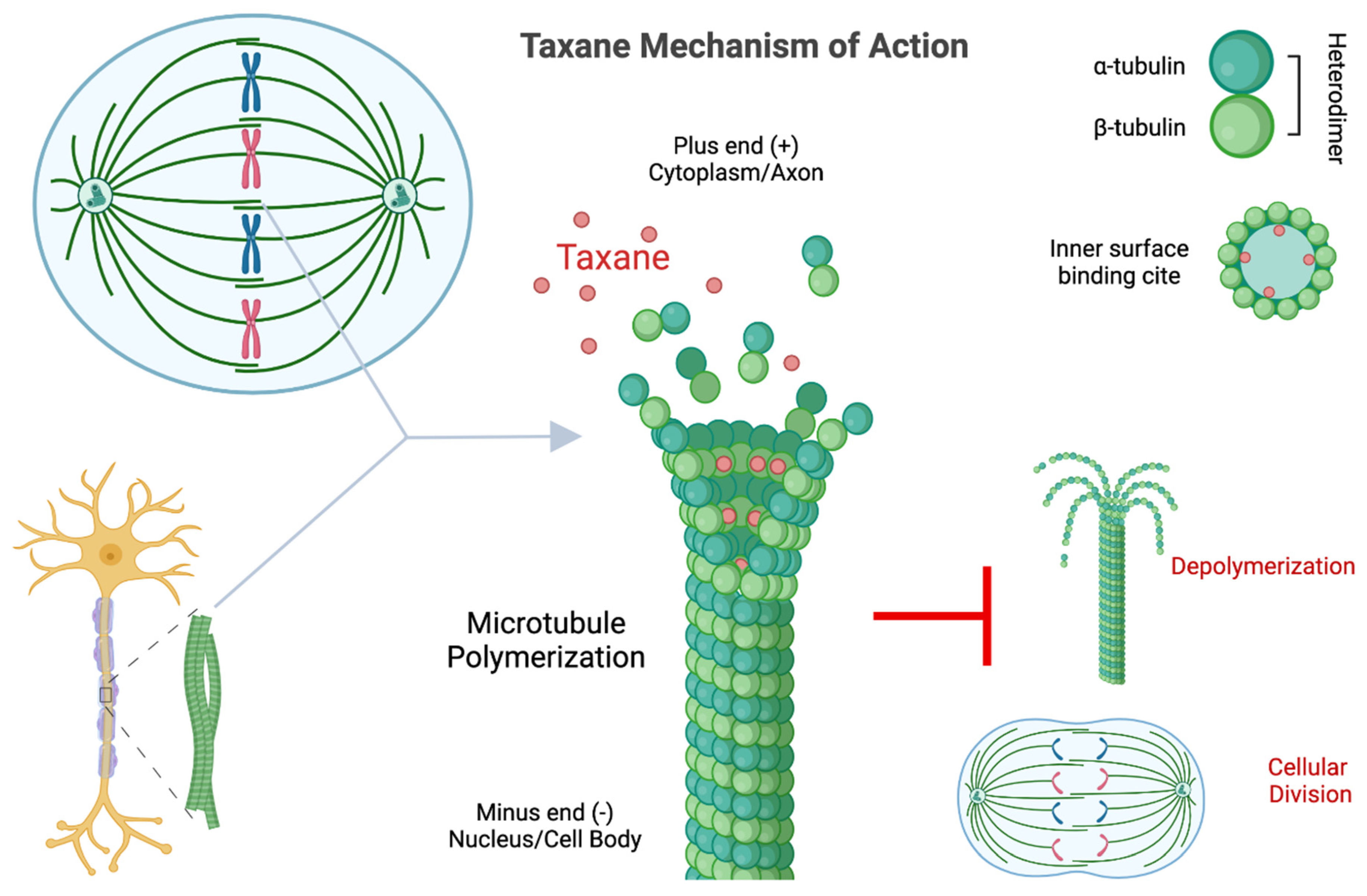
Figure 2.
Taxane-induced androgen receptor (AR) microtubule-dependent signaling dysfunction. In the field of prostate cancer research, taxanes are observed to interfere with ligand-mediated AR signaling by inhibiting AR binding to dynein motor proteins, preventing nuclear trafficking of AR. Created with BioRender.
Figure 2.
Taxane-induced androgen receptor (AR) microtubule-dependent signaling dysfunction. In the field of prostate cancer research, taxanes are observed to interfere with ligand-mediated AR signaling by inhibiting AR binding to dynein motor proteins, preventing nuclear trafficking of AR. Created with BioRender.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Chmielewski, N.N.; Limoli, C.L. Sex Differences in Taxane Toxicities. Cancers 2022, 14, 3325. https://doi.org/10.3390/cancers14143325
AMA Style
Chmielewski NN, Limoli CL. Sex Differences in Taxane Toxicities. Cancers. 2022; 14(14):3325. https://doi.org/10.3390/cancers14143325
Chicago/Turabian StyleChmielewski, Nicole N., and Charles L. Limoli. 2022. "Sex Differences in Taxane Toxicities" Cancers 14, no. 14: 3325. https://doi.org/10.3390/cancers14143325
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.