
P720R USP8 Mutation Is Associated with a Better Responsiveness to Pasireotide in ACTH-Secreting PitNETs

 , , , ,
, , , ,  , , ,
, , ,
Abstract
:Simple Summary
Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. ACTH-Secreting Pituitary Cell Culture
2.2. DNA Extraction and Sanger Sequencing
2.3. Cell Transfection and USP8 Silencing
2.4. Measurement of ACTH Levels
2.5. Western Blot Analysis
2.6. Cell Proliferation Assay
2.7. Cell Apoptosis
2.8. Fluorescence Microscopy and In Situ Proximity Ligation Assay
2.9. Statistical Analysis
3. Results
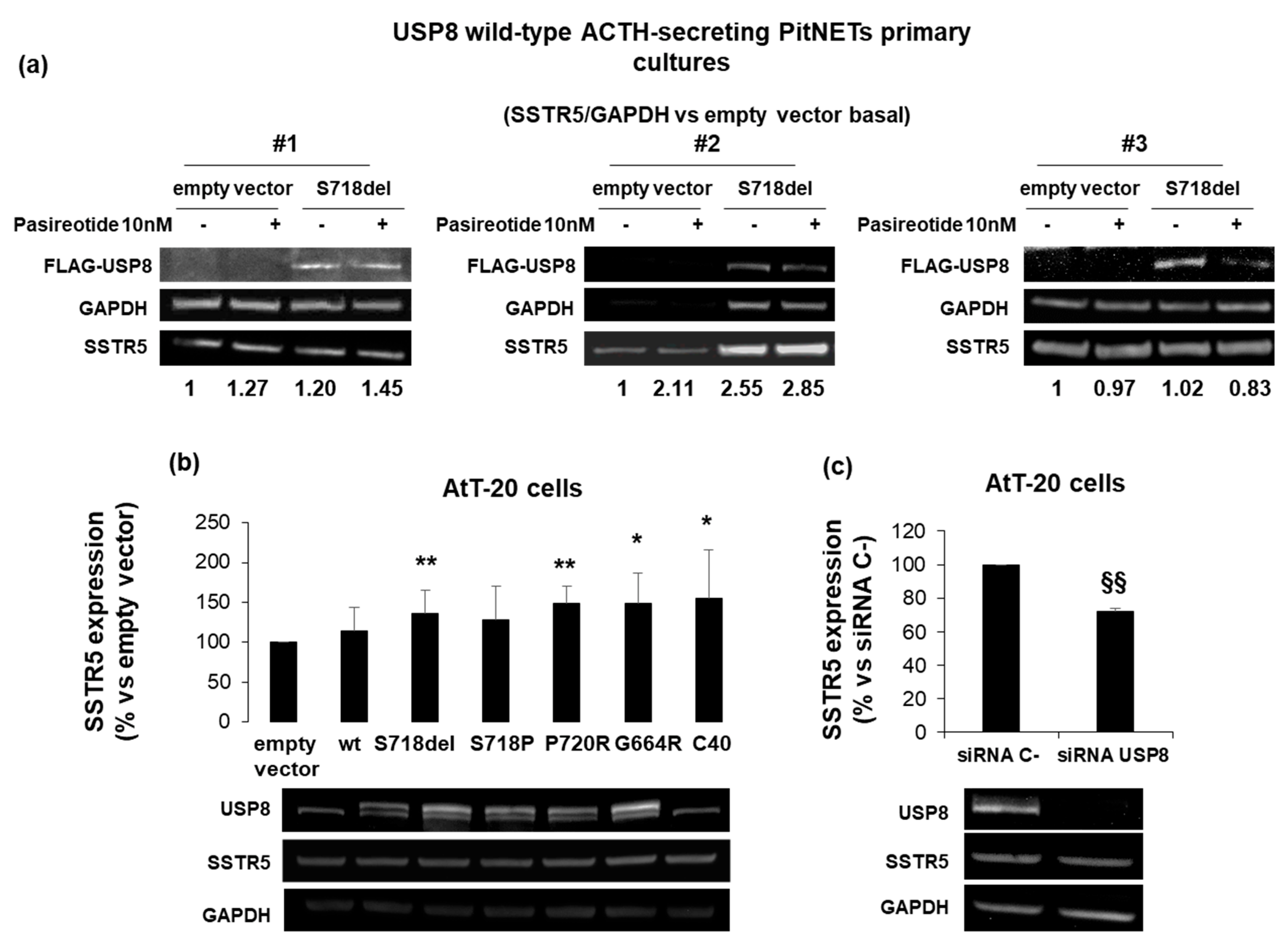
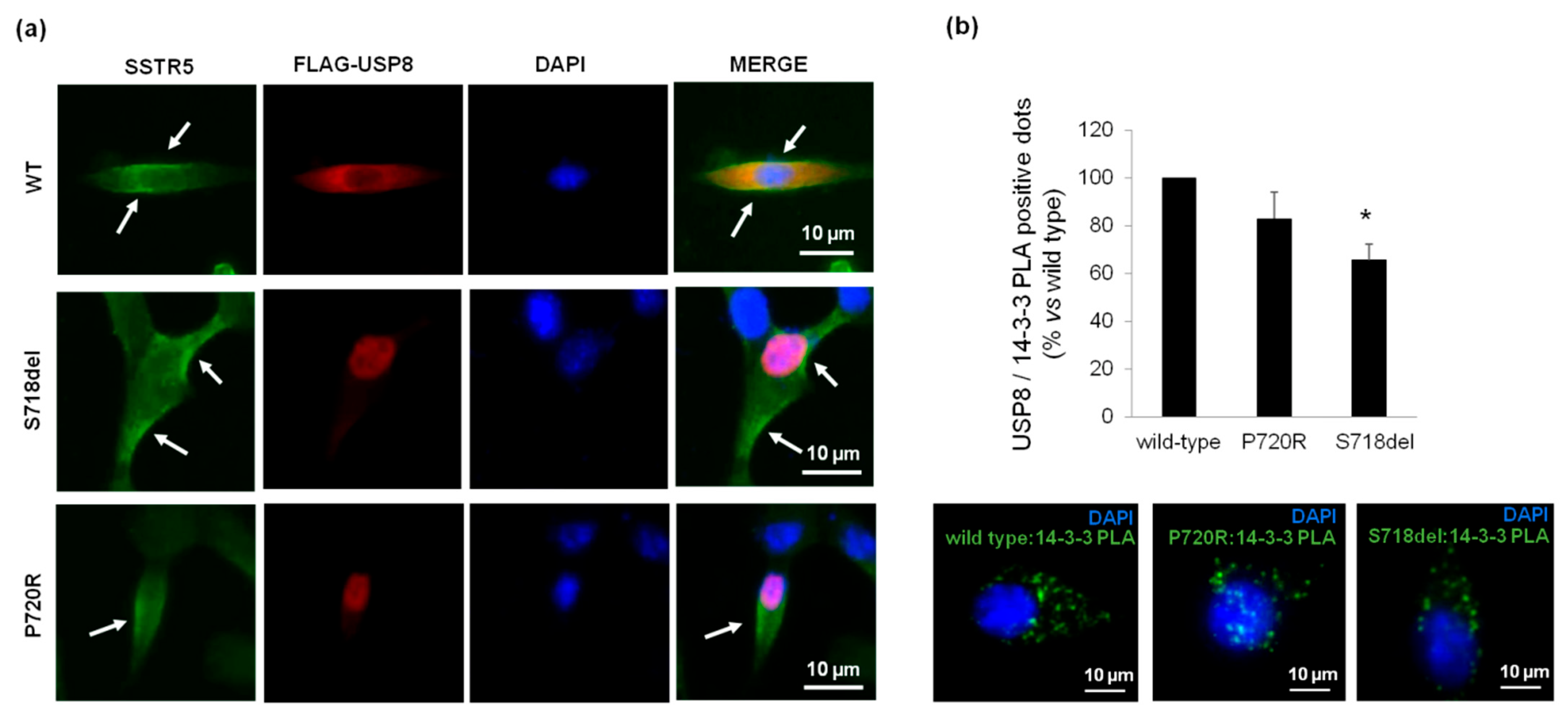
3.1. USP8 Mutants Upregulate SSTR5 Expression Levels in Primary Cultured Cells and AtT20 Cells
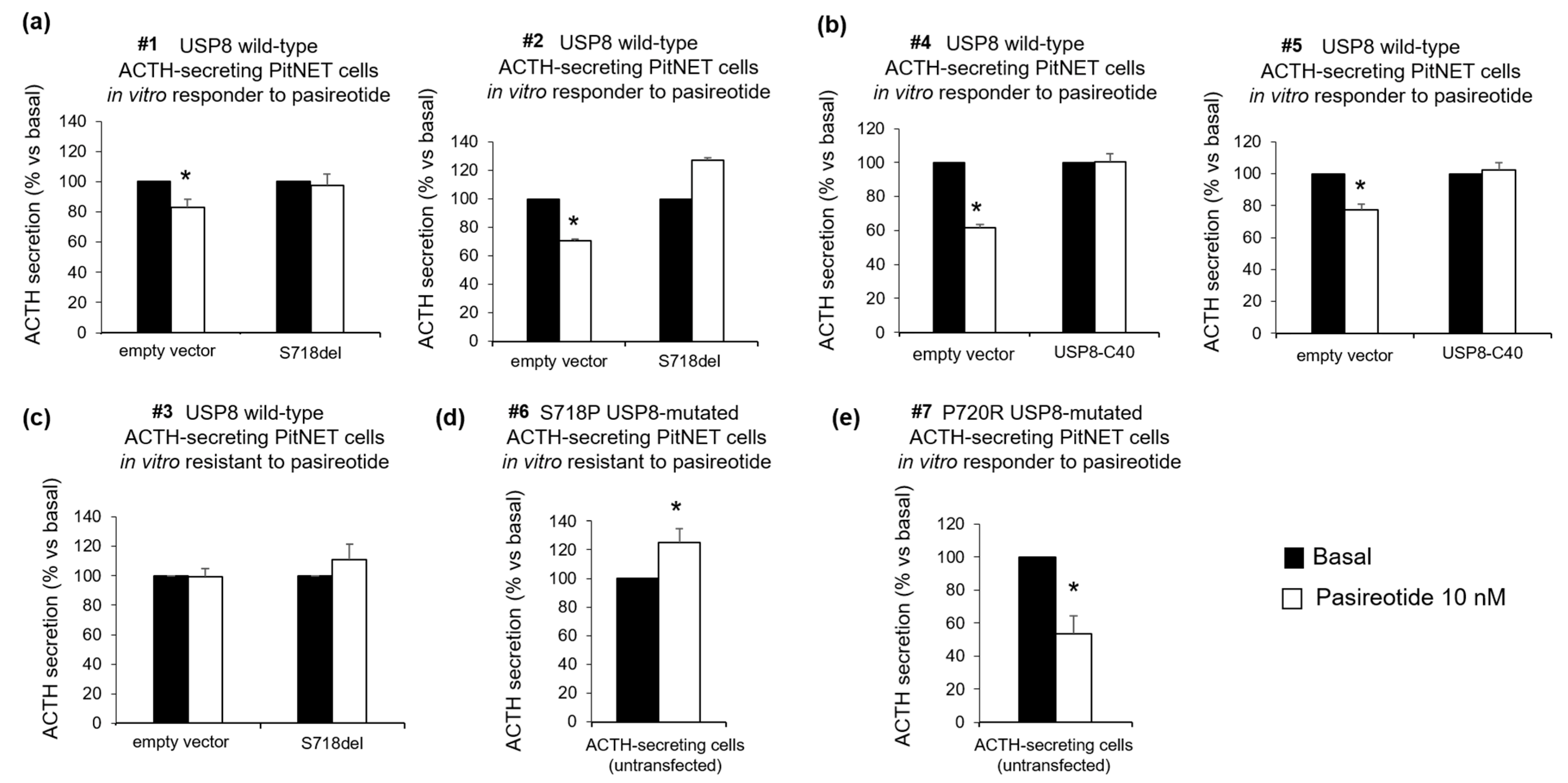
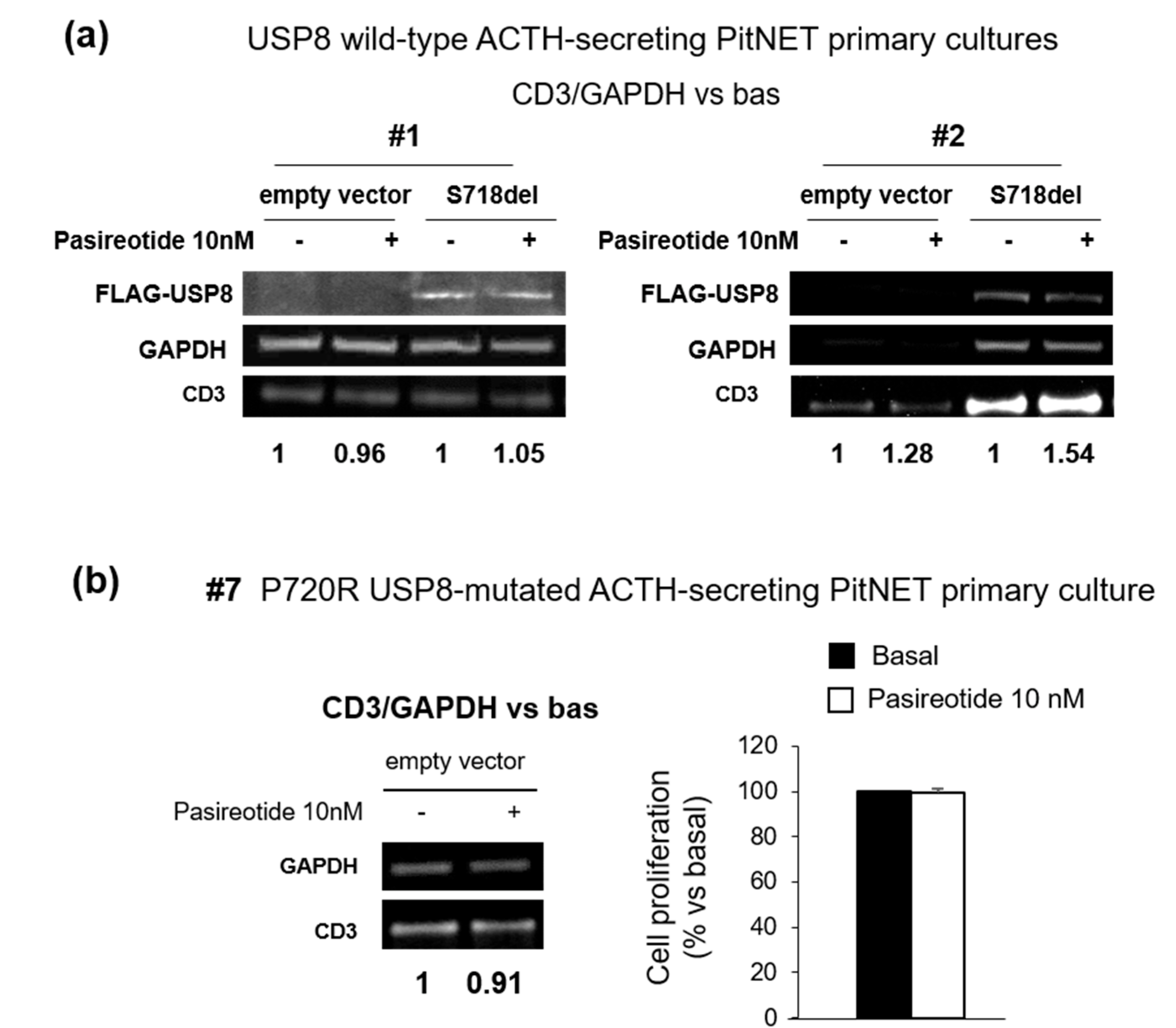
3.2. USP8 Mutants Dissimilarly Modulate the Sensitiveness to Pasireotide-Mediated Antisecretory Action in Primary Cultured Cells
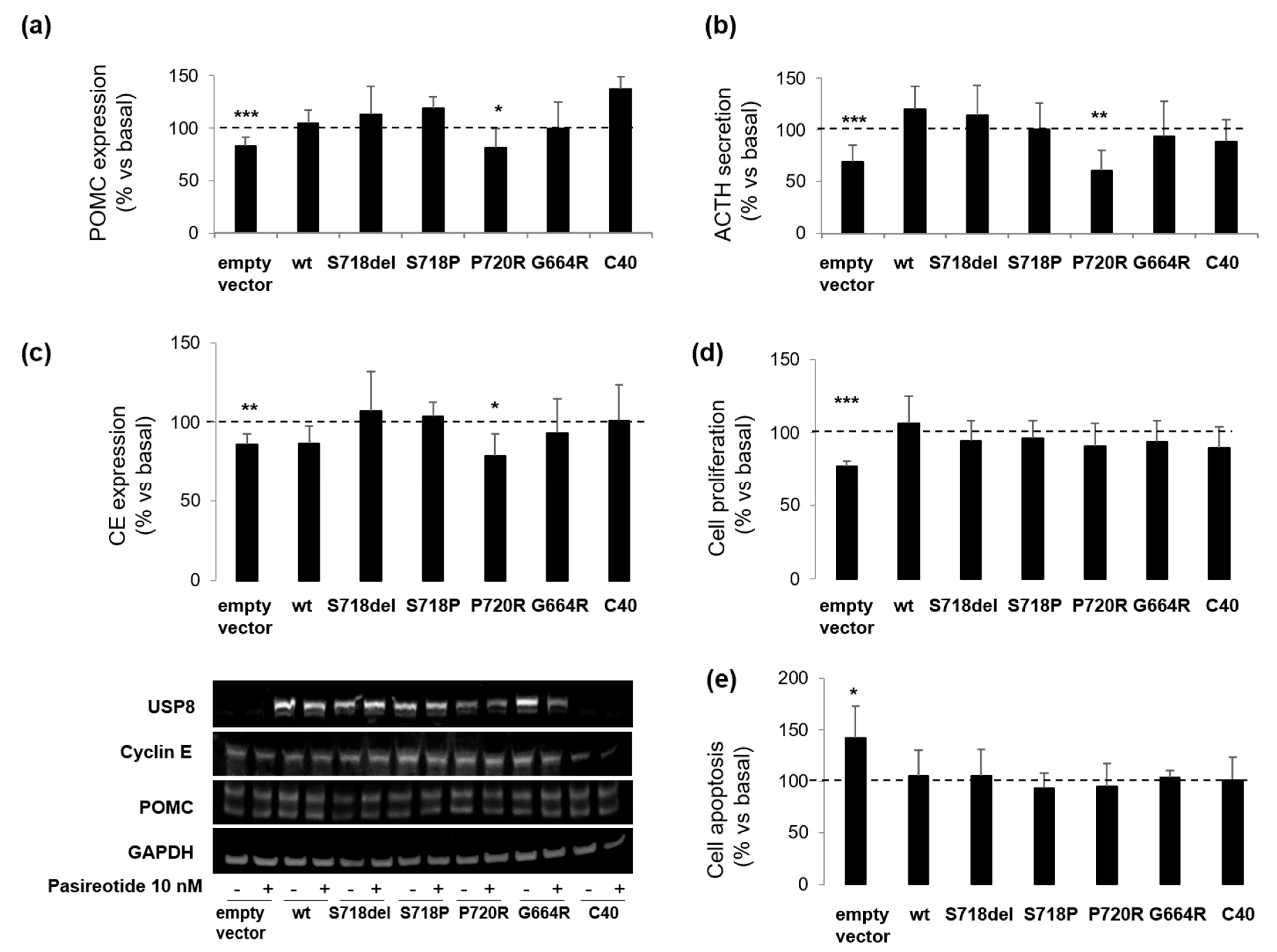
3.3. P720R USP8 Is the Sole Mutant Showing Favourable Responses to Pasireotide in AtT-20 Cells
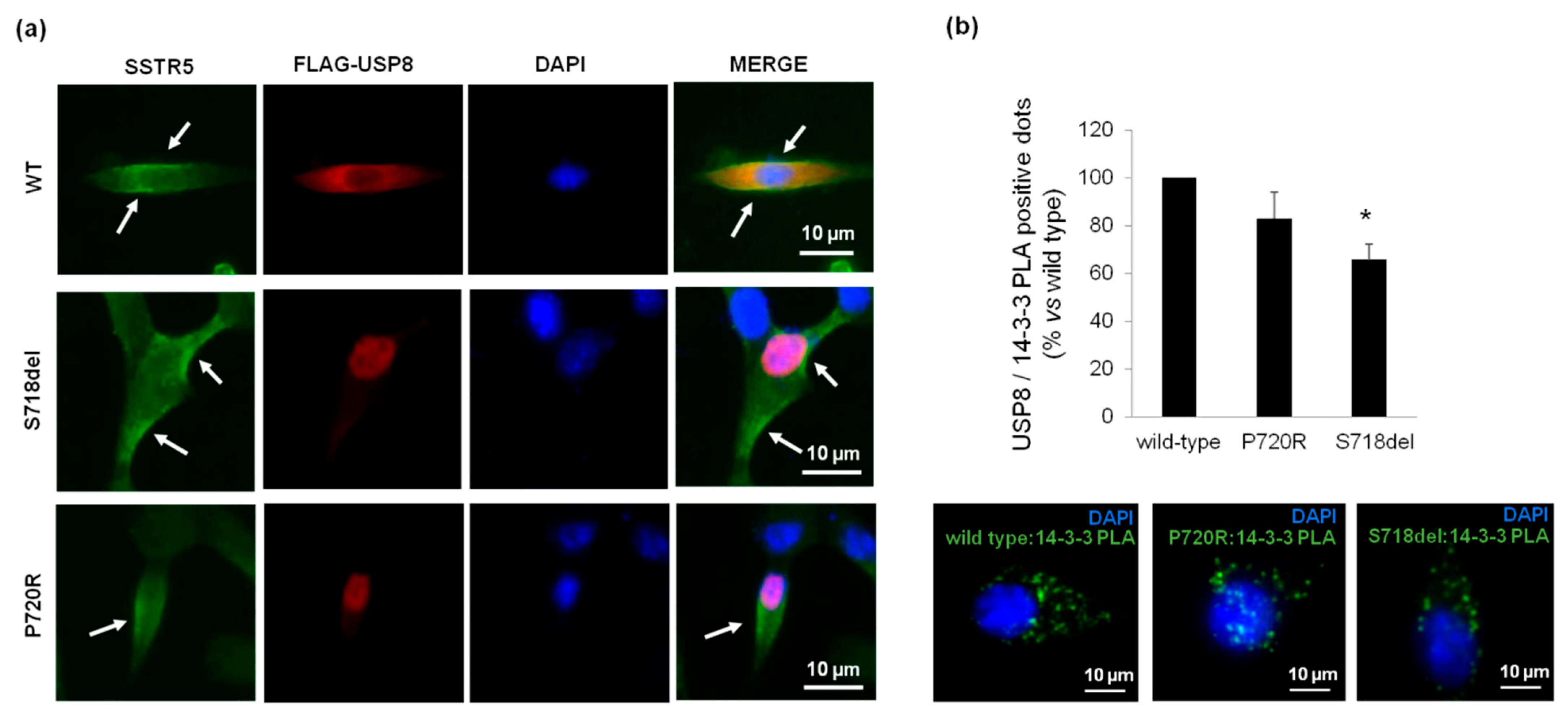
3.4. P720R USP8 Mutant Exhibits 14-3-3 Proteins Binding Ability in AtT-20 Cells
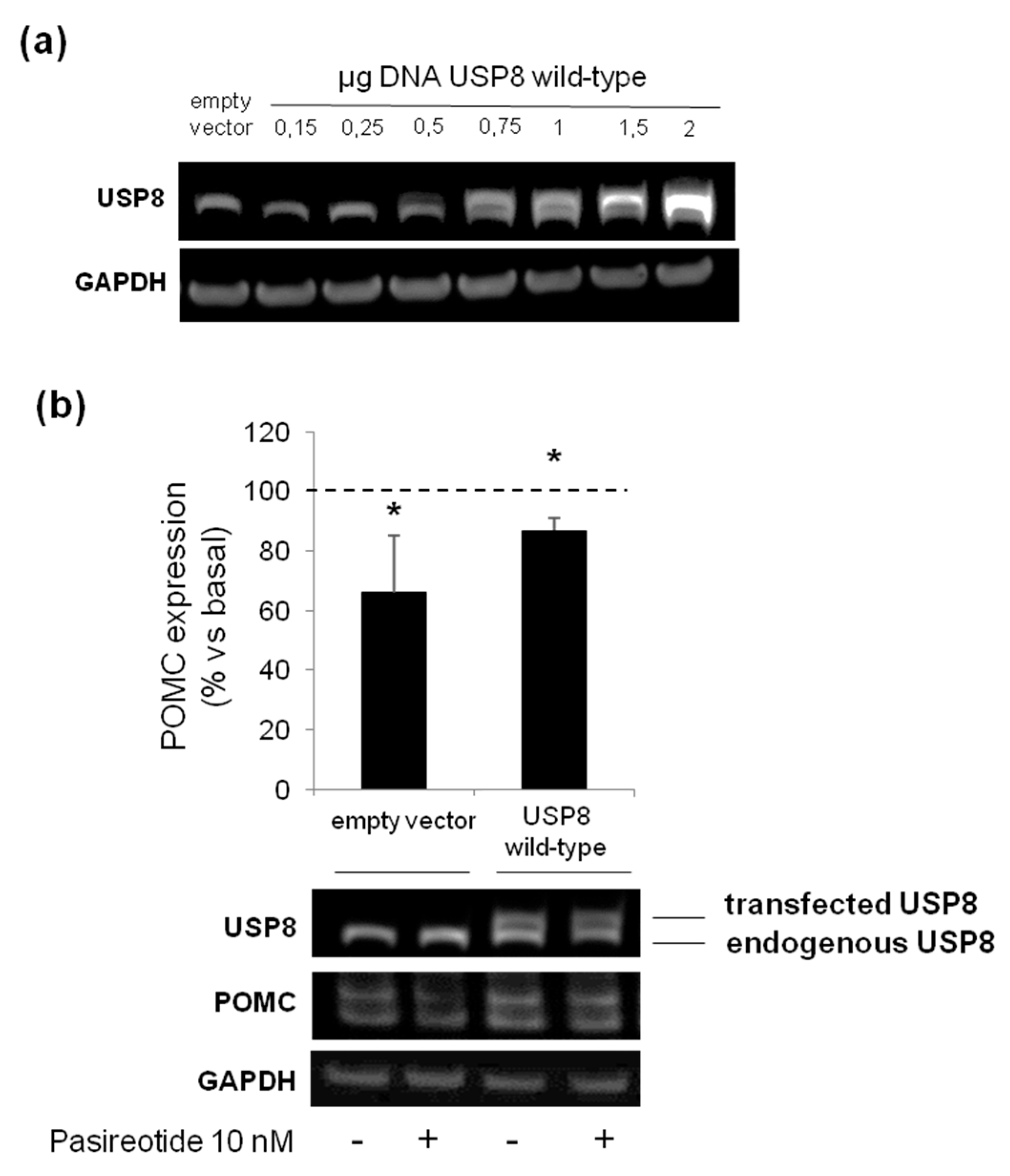
3.5. The Loss of Pasireotide Responsiveness in USP8 Wild Type Overexpressing AtT-20 Cells Is Due to Abundant USP8 Levels
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ma, Z.Y.; Song, Z.J.; Chen, J.H.; Wang, Y.F.; Li, S.Q.; Zhou, L.F.; Mao, Y.; Li, Y.M.; Hu, R.G.; Zhao, Z.Y.; et al. Recurrent gain-of-function USP8 mutations in Cushing’s disease. Cell Res. 2015, 25, 306–317. [Google Scholar] [CrossRef] [PubMed]
- Reincke, M.; Sbiera, S.; Hayakawa, A.; Theodoropoulou, M.; Osswald, A.; Beuschlein, F.; Meitinger, T.; Mizuno-Yamasaki, E.; Kawaguchi, K.; Saeki, Y.; et al. Mutations in the deubiquitinase gene USP8 cause Cushing’s disease. Nat. Genet. 2015, 47, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Rivas, L.G.; Theodoropoulou, M.; Puar, T.H.; Fazel, J.; Stieg, M.R.; Ferraù, F.; Assié, G.; Gadelha, M.R.; Deutschbein, T.; Fragoso, M.C.; et al. Somatic USP8 mutations are frequent events in corticotroph tumor progression causing Nelson’s tumor. Eur. J. Endocrinol. 2018, 178, 57–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayashi, K.; Inoshita, N.; Kawaguchi, K.; Ibrahim Ardisasmita, A.; Suzuki, H.; Fukuhara, N.; Okada, M.; Nishioka, H.; Takeuchi, Y.; Komada, M.; et al. The USP8 mutational status may predict drug susceptibility in corticotroph adenomas of Cushing’s disease. Eur. J. Endocrinol. 2016, 174, 213–226. [Google Scholar] [CrossRef] [PubMed]
- Faucz, F.R.; Tirosh, A.; Tatsi, C.; Berthon, A.; Hernández-Ramírez, L.C.; Settas, N.; Angelousi, A.; Correa, R.; Papadakis, G.Z.; Chittiboina, P.; et al. Somatic USP8 gene mutations are a common cause of pediatric Cushing disease. J. Clin. Endocrinol. Metab. 2017, 102, 2836–2843. [Google Scholar] [CrossRef]
- Ballmann, C.; Thiel, A.; Korah, H.E.; Reis, A.C.; Saeger, W.; Stepanow, S.; Köhrer, K.; Reifenberger, G.; Knobbe-Thomsen, C.B.; Knappe, U.J.; et al. USP8 mutations in pituitary Cushing adenomas-targeted analysis by next-generation sequencing. J. Endocr. Soc. 2018, 2, 266–278. [Google Scholar] [CrossRef] [Green Version]
- Bujko, M.; Kober, P.; Boresowicz, J.; Rusetska, N.; Paziewska, A.; Dąbrowska, M.; Piaścik, A.; Pękul, M.; Zieliński, G.; Kunicki, J.; et al. USP8 mutations in corticotroph adenomas determine a distinct gene expression profile irrespective of functional tumour status. Eur. J. Endocrinol. 2019, 181, 615–627. [Google Scholar] [CrossRef]
- Wanichi, I.Q.; de Paula Mariani, B.M.; Frassetto, F.P.; Siqueira, S.A.C.; de Castro Musolino, N.R.; Cunha-Neto, M.B.C.; Ochman, G.; Cescato, V.A.S.; Machado, M.C.; Trarbach, E.B.; et al. Cushing’s disease due to somatic USP8 mutations: A systematic review and meta-analysis. Pituitary 2019, 22, 435–442. [Google Scholar] [CrossRef]
- Weigand, I.; Knobloch, L.; Flitsch, J.; Saeger, W.; Monoranu, C.M.; Höfner, K.; Herterich, S.; Rotermund, R.; Ronchi, C.L.; Buchfelder, M.; et al. Impact of USP8 gene mutations on protein deregulation in Cushing disease. J. Clin. Endocrinol. Metab. 2019, 104, 2535–2546. [Google Scholar] [CrossRef]
- Sbiera, S.; Perez-Rivas, L.G.; Taranets, L.; Weigand, I.; Flitsch, J.; Graf, E.; Monoranu, C.M.; Saeger, W.; Hagel, C.; Honegger, J.; et al. Driver mutations in USP8 wild-type Cushing’s disease. Neuro Oncol. 2019, 21, 1273–1283. [Google Scholar] [CrossRef]
- Sbiera, S.; Kunz, M.; Weigand, I.; Deutschbein, T.; Dandekar, T.; Fassnacht, M. The new genetic landscape of Cushing’s disease: Deubiquitinases in the spotlight. Cancers 2019, 11, 1761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martins, C.S.; Camargo, R.C.; Coeli-Lacchini, F.B.; Saggioro, F.P.; Moreira, A.C.; de Castro, M. USP8 mutations and cell cycle regulation in corticotroph adenomas. Horm. Metab. Res. 2020, 52, 117–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castellnou, S.; Vasiljevic, A.; Lapras, V.; Raverot, V.; Alix, E.; Borson-Chazot, F.; Jouanneau, E.; Raverot, G.; Lasolle, H. SST5 expression and USP8 mutation in functioning and silent corticotroph pituitary tumors. Endocr. Connect. 2020, 9, 243–253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sesta, A.; Cassarino, M.F.; Terreni, M.; Ambrogio, A.G.; Libera, L.; Bardelli, D.; Lasio, G.; Losa, M.; Pecori Giraldi, F. Ubiquitin-specific protease 8 mutant corticotrope adenomas present unique secretory and molecular features and shed light on the role of ubiquitylation on ACTH processing. Neuroendocrinology 2020, 110, 119–129. [Google Scholar] [CrossRef]
- Treppiedi, D.; Barbieri, A.M.; Di Muro, G.; Marra, G.; Mangili, F.; Catalano, R.; Esposito, E.; Ferrante, E.; Serban, A.L.; Locatelli, M.; et al. Genetic profiling of a cohort of italian patients with acth-secreting pituitary tumors and characterization of a novel USP8 gene variant. Cancers 2021, 13, 4022. [Google Scholar] [CrossRef]
- Mizuno, E.; Iura, T.; Mukai, A.; Yoshimori, T.; Kitamura, N.; Komada, M. Regulation of epidermal growth factor receptor down-regulation by UBPY-mediated deubiquitination at endosomes. Mol. Biol. Cell. 2005, 16, 5163–5174. [Google Scholar] [CrossRef] [Green Version]
- Mizuno, E.; Kitamura, N.; Komada, M. 14-3-3-dependent inhibition of the deubiquitinating activity of UBPY and its cancellation in the M phase. Exp. Cell Res. 2007, 313, 3624–3634. [Google Scholar] [CrossRef]
- Centorrino, F.; Ballone, A.; Wolter, M.; Ottmann, C. Biophysical and structural insight into the USP8/14-3-3 interaction. FEBS Lett. 2018, 592, 1211–1220. [Google Scholar] [CrossRef] [Green Version]
- Dufner, A.; Knobeloch, K.P. Ubiquitin-specific protease 8 (USP8/UBPy): A prototypic multidomain deubiquitinating enzyme with pleiotropic functions. Biochem. Soc. Trans. 2019, 47, 1867–1879. [Google Scholar] [CrossRef]
- Losa, M.; Mortini, P.; Pagnano, A.; Detomas, M.; Cassarino, M.F.; Pecori Giraldi, F. Clinical characteristics and surgical outcome in USP8-mutated human adrenocorticotropic hormone-secreting pituitary adenomas. Endocrine 2019, 63, 240–246. [Google Scholar] [CrossRef]
- Albani, A.; Pérez-Rivas, L.G.; Dimopoulou, C.; Zopp, S.; Colón-Bolea, P.; Roeber, S.; Honegger, J.; Flitsch, J.; Rachinger, W.; Buchfelder, M.; et al. The USP8 mutational status may predict long-term remission in patients with Cushing’s disease. Clin. Endocrinol. 2018, 89, 454–458. [Google Scholar] [CrossRef] [PubMed]
- Colao, A.; Boscaro, M.; Ferone, D.; Casanueva, F.F. Managing Cushing’s disease: The state of the art. Endocrine 2014, 47, 9–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simeoli, C.; Ferrigno, R.; De Martino, M.C.; Iacuaniello, D.; Papa, F.; Angellotti, D.; Pivonello, C.; Patalano, R.; Negri, M.; Colao, A.; et al. The treatment with pasireotide in Cushing’s disease: Effect of long-term treatment on clinical picture and metabolic profile and management of adverse events in the experience of a single center. J. Endocrinol. Investig. 2020, 43, 57–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van der Hoek, J.; Waaijers, M.; van Koetsveld, P.M.; Sprij-Mooij, D.; Feelders, R.A.; Schmid, H.A.; Schoeffter, P.; Hoyer, D.; Cervia, D.; Taylor, J.E.; et al. Distinct functional properties of native somatostatin receptor subtype 5 compared with subtype 2 in the regulation of ACTH release by corticotroph tumor cells. Am. J. Physiol. Endocrinol. Metab. 2005, 289, E278–E287. [Google Scholar] [CrossRef] [Green Version]
- Hofland, L.J.; van der Hoek, J.; Feelders, R.; van Aken, M.O.; van Koetsveld, P.M.; Waaijers, M.; Sprij-Mooij, D.; Bruns, C.; Weckbecker, G.; de Herder, W.W.; et al. The multi-ligand somatostatin analogue SOM230 inhibits ACTH secretion by cultured human corticotroph adenomas via somatostatin receptor type 5. Eur. J. Endocrinol. 2005, 152, 645–654. [Google Scholar] [CrossRef]
- Peverelli, E.; Giardino, E.; Treppiedi, D.; Vitali, E.; Cambiaghi, V.; Locatelli, M.; Lasio, G.B.; Spada, A.; Lania, A.G.; Mantovani, G. Filamin A (FLNA) plays an essential role in somatostatin receptor 2 (SST2) signaling and stabilization after agonist stimulation in human and rat somatotroph tumor cells. Endocrinology 2014, 155, 2932–2941. [Google Scholar] [CrossRef] [Green Version]
- Treppiedi, D.; Di Muro, G.; Mangili, F.; Catalano, R.; Giardino, E.; Barbieri, A.M.; Locatelli, M.; Arosio, M.; Spada, A.; Peverelli, E.; et al. Filamin A is required for somatostatin receptor type 5 expression and pasireotide-mediated signaling in pituitary corticotroph tumor cells. Mol. Cell. Endocrinol. 2021, 524, 111159. [Google Scholar] [CrossRef]
- Kakihara, K.; Asamizu, K.; Moritsugu, K.; Kubo, M.; Kitaguchi, T.; Endo, A.; Kidera, A.; Ikeguchi, M.; Kato, A.; Komada, M.; et al. Molecular basis of ubiquitin-specific protease 8 autoinhibition by the WW-like domain. Commun. Biol. 2021, 41272. [Google Scholar] [CrossRef]
- Colao, A.; Petersenn, S.; Newell-Price, J.; Findling, J.W.; Gu, F.; Maldonado, M.; Schoenherr, U.; Mills, D.; Salgado, L.R.; Biller, B.M. Pasireotide B2305 Study Group. A 12-month phase 3 study of pasireotide in Cushing’s disease. N. Engl. J. Med. 2012, 366, 914–924. [Google Scholar] [CrossRef] [Green Version]
- Tulipano, G.; Stumm, R.; Pfeiffer, M.; Kreienkamp, H.; Höllt, V.; Schulz, S. Differential beta-arrestin trafficking and endosomal sorting of somatostatin receptor subtypes. J. Biol. Chem. 2004, 279, 21374–21382. [Google Scholar] [CrossRef] [Green Version]
- Vidal, M.; Cusick, M.E.; Barabasi, A.L. Interactome networks and human disease. Cell 2011, 144, 986–998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ottmann, C. Small-molecule modulators of 14-3-3 protein–protein interactions. Bioorg. Med. Chem. 2013, 21, 4058–4062. [Google Scholar] [CrossRef] [PubMed]
- Sluchanko, N.N. Association of multiple phosphorylated proteins with the 14–3-3 regulatory hubs: Problems and perspectives. J. Mol. Biol. 2018, 430, 20–26. [Google Scholar] [CrossRef] [PubMed]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Treppiedi, D.; Marra, G.; Di Muro, G.; Esposito, E.; Barbieri, A.M.; Catalano, R.; Mangili, F.; Bravi, F.; Locatelli, M.; Lania, A.G.; et al. P720R USP8 Mutation Is Associated with a Better Responsiveness to Pasireotide in ACTH-Secreting PitNETs. Cancers 2022, 14, 2455. https://doi.org/10.3390/cancers14102455
Treppiedi D, Marra G, Di Muro G, Esposito E, Barbieri AM, Catalano R, Mangili F, Bravi F, Locatelli M, Lania AG, et al. P720R USP8 Mutation Is Associated with a Better Responsiveness to Pasireotide in ACTH-Secreting PitNETs. Cancers. 2022; 14(10):2455. https://doi.org/10.3390/cancers14102455
Chicago/Turabian StyleTreppiedi, Donatella, Giusy Marra, Genesio Di Muro, Emanuela Esposito, Anna Maria Barbieri, Rosa Catalano, Federica Mangili, Francesca Bravi, Marco Locatelli, Andrea Gerardo Lania, and et al. 2022. "P720R USP8 Mutation Is Associated with a Better Responsiveness to Pasireotide in ACTH-Secreting PitNETs" Cancers 14, no. 10: 2455. https://doi.org/10.3390/cancers14102455
APA StyleTreppiedi, D., Marra, G., Di Muro, G., Esposito, E., Barbieri, A. M., Catalano, R., Mangili, F., Bravi, F., Locatelli, M., Lania, A. G., Ferrante, E., Indirli, R., Nozza, E., Arlati, F., Spada, A., Arosio, M., Mantovani, G., & Peverelli, E. (2022). P720R USP8 Mutation Is Associated with a Better Responsiveness to Pasireotide in ACTH-Secreting PitNETs. Cancers, 14(10), 2455. https://doi.org/10.3390/cancers14102455