
Identification of a Candidate Gene Set Signature for the Risk of Progression in IgM MGUS to Smoldering/Symptomatic Waldenström Macroglobulinemia (WM) by a Comparative Transcriptome Analysis of B Cells and Plasma Cells

 , , , , and
, , , , and
Simple Summary
Abstract

1. Introduction
2. Results
2.1. Total RNA Concentration in BM CD19+ Cells and BM CD138+ Cells
2.2. GEP Results
2.3. CD19+ Patterns
2.3.1. Transmembrane Proteins
2.3.2. Hematopoietic Cell Lineage (KEGG Pathway hsa04640)
2.3.3. Cell Adhesion Molecules (CAMs) (KEGG Pathway hsa04514)
2.3.4. Focal Adhesion (KEGG Pathway hsa04510)
2.3.5. Adherents Junctions (KEGG Pathway hsa04520) and WNT/β-Catenin Signaling Pathway (KEGG Pathway hsa04310)
2.3.6. Calcium Signaling Pathway (KEGG Pathway hsa04020)
2.3.7. BCR Signaling Pathway (KEGG Pathway hsa04662)
2.3.8. Complement and Coagulation Cascade (KEGG Pathway hsa04610) and Platelet Activation (KEGG Pathway hsa04611)
2.3.9. Cytokine-Cytokine Receptor Interactions (KEGG Pathway hsa04060) and IL-17 Signaling Pathway (KEGG Pathway hsa04657)
2.3.10. JAK/STAT Signaling Pathway (KEGG Pathway hsa04630)
2.3.11. FOXO Signaling Pathway (KEGG Pathway hsa04068) and TGF-β Signaling Pathway (KEGG Pathway hsa04350)
2.3.12. NFKB Signaling Pathway (KEGG Pathway hsa04064)
2.3.13. p53 Signaling Pathway (KEGG Pathway hsa04115)
2.3.14. Zinc Finger Proteins
2.3.15. Genes Belonging to Pattern 2 with Very High FC in CD19+ Cells
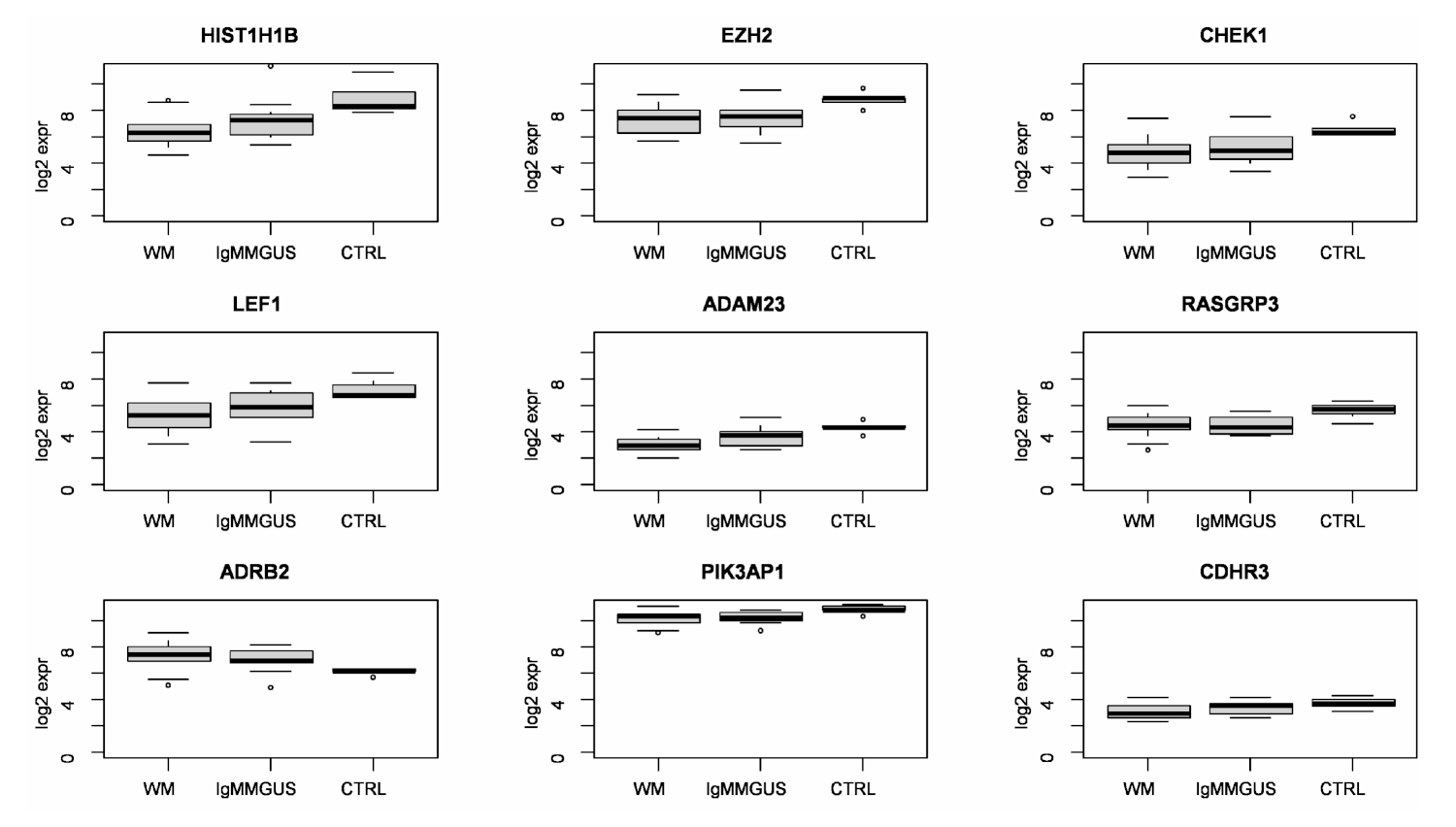
2.3.16. Candidate Genes of the Risk of Progression of IgM MGUS in WM
2.4. CD138+ Cells
2.4.1. Adherens Junction (KEGG Pathway hsa04520), Gap Junction (KEGG Pathway hsa04540), and Tight Junction (KEGG Pathway hsa04530) in Pattern 2 CD138+ Cells
2.4.2. Cell Adhesion Molecules (KEGG Pathway hsa04514) in Pattern 2 CD138+ Cells
2.4.3. p53 Signaling Pathway (KEGG Pathway hsa04115) in Pattern 2 CD138+ Cells
2.4.4. Calcium Signaling Pathway (KEGG Pathway hsa04020) in Pattern 2 CD138+ Cell
2.5. Common Genes Deregulated in CD19+ Cells and CD138+ Cells
3. Discussion
3.1. Different Expressions of Genes Encoding Transmembrane Molecules, Hematopoietic Cell Surface Antigens, Cell and Focal Adhesion Proteins in B Cells
3.2. Adherens Junctions and WNT Signaling Pathway
3.3. Deregulation of Calcium Signaling Pathway
3.4. BCR Signaling Pathway
3.5. Complement and Activation Cascade and Platelet Activation
3.6. Cytokine-Cytokine Receptor Interactions and IL-17 Signaling Pathway
3.7. Gene Expression Alterations of Signaling Pathway Responsible for Cell Cycle, Proliferation and Survival (JAK/STAT, FOXO, TGF-β, NFKB, and p53 Signaling Pathways)
3.8. Deregulation of p53 Signaling Pathway
3.9. Deregulation of Genes in WM vs. IgM MGUS vs. CTRLs in Plasma Cells
4. Materials and Methods
4.1. Patients
4.2. Isolation of BM CD19+ Cells and BM CD138+ Cells Using Immunomagnetic Beads
4.3. Cell Cryopreservation and RNA Extraction
4.4. GEP Experiments
4.5. Gene Expression Data Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bustoros, M.; Sklavenitis-Pistofidis, R.; Kapoor, P.; Liu, C.-J.; Kastritis, E.; Zanwar, S.; Fell, G.; Abeykoon, J.P.; Hornburg, K.; Neuse, C.J.; et al. Progression Risk Stratification of Asymptomatic Waldenström Macroglobulinemia. J. Clin. Oncol. 2019, 37, 1403–1411. [Google Scholar] [CrossRef] [PubMed]
- Gertz, M.A. Waldenström macroglobulinemia: 2019 update on diagnosis, risk stratification, and management. Am. J. Hematol. 2019, 94, 266–276. [Google Scholar] [CrossRef] [PubMed]
- Paludo, J.; Ansell, S.M. Waldenström macroglobulinemia: Biology, genetics, and therapy. Blood Lymphat. Cancer Targets Ther. 2016, 6, 49–58. [Google Scholar] [CrossRef]
- Kyle, R.A.; Benson, J.; Larson, D.; Therneau, T.; Dispenzieri, A.; Melton, L.J.; Rajkumar, S.V. IgM Monoclonal Gammopathy of Undetermined Significance and Smoldering Waldenström’s Macroglobulinemia. Clin. Lymphoma Myeloma 2009, 9, 17–18. [Google Scholar] [CrossRef] [PubMed]
- Jiménez, C.; Prieto-Conde, M.I.; García-Alvarez, M.; Alcoceba, M.; Escalante, F.; Chillon, M.D.C.; De Coca, A.G.; Balanzategui, A.; Cantalapiedra, A.; Aguilar, C.; et al. Unraveling the heterogeneity of IgM monoclonal gammopathies: A gene mutational and gene expression study. Ann. Hematol. 2018, 97, 475–484. [Google Scholar] [CrossRef] [PubMed]
- Kyle, R.A.; Larson, D.R.; Therneau, T.M.; Dispenzieri, A.; Kumar, S.; Cerhan, J.R.; Rajkumar, S.V. Long-Term Follow-up of Monoclonal Gammopathy of Undetermined Significance. N. Engl. J. Med. 2018, 378, 241–249. [Google Scholar] [CrossRef]
- Treon, S.P.; Xu, L.; Guerrera, M.L.; Jimenez, C.; Hunter, Z.R.; Liu, X.; Demos, M.; Gustine, J.; Chan, G.; Munshi, M.; et al. Genomic Landscape of Waldenström Macroglobulinemia and Its Impact on Treatment Strategies. J. Clin. Oncol. 2020, 38, 1198–1208. [Google Scholar] [CrossRef]
- Castillo, J.J.; Xu, L.; Gustine, J.N.; Keezer, A.; Meid, K.; Dubeau, T.E.; Liu, X.; Demos, M.G.; Kofides, A.; Tsakmaklis, N.; et al. CXCR4 mutation subtypes impact response and survival outcomes in patients with Waldenström macroglobulinaemia treated with ibrutinib. Br. J. Haematol. 2019, 187, 356–363. [Google Scholar] [CrossRef]
- Xu, L.; Hunter, Z.R.; Tsakmaklis, N.; Cao, Y.; Yang, G.; Chen, J.; Liu, X.; Kanan, S.; Castillo, J.J.; Tai, Y.-T.; et al. Clonal architecture ofCXCR4WHIM-like mutations in Waldenström Macroglobulinaemia. Br. J. Haematol. 2016, 172, 735–744. [Google Scholar] [CrossRef]
- Hunter, Z.R.; Xu, L.; Tsakmaklis, N.; Demos, M.G.; Kofides, A.; Jimenez, C.; Chan, G.G.; Chen, J.; Liu, X.; Munshi, M.; et al. Insights into the genomic landscape of MYD88 wild-type Waldenström macroglobulinemia. Blood Adv. 2018, 2, 2937–2946. [Google Scholar] [CrossRef]
- Poulain, S.; Roumier, C.; Decambron, A.; Renneville, A.; Herbaux, C.; Bertrand, E.; Tricot, S.; Daudignon, A.; Galiègue-Zouitina, S.; Soenen, V.; et al. MYD88 L265P mutation in Waldenstrom macroglobulinemia. Blood 2013, 121, 4504–4511. [Google Scholar] [CrossRef]
- Varettoni, M.; Zibellini, S.; DeFrancesco, I.; Ferretti, V.V.; Rizzo, E.; Malcovati, L.; Gallì, A.; Della Porta, M.G.; Boveri, E.; Arcaini, L.; et al. Pattern of somatic mutations in patients with Waldenström macroglobulinemia or IgM monoclonal gammopathy of undetermined significance. Haematologica 2017, 102, 2077–2085. [Google Scholar] [CrossRef]
- Xu, L.; Hunter, Z.R.; Yang, G.; Zhou, Y.; Cao, Y.; Liu, X.; Morra, E.; Trojani, A.; Greco, A.; Arcaini, L.; et al. MYD88 L265P in Waldenström macroglobulinemia, immunoglobulin M monoclonal gammopathy, and other B-cell lymphoproliferative disorders using conventional and quantitative allele-specific polymerase chain reaction. Blood 2013, 121, 2051–2058. [Google Scholar] [CrossRef]
- García-Sanz, R.; Dogliotti, I.; Zaccaria, G.M.; Ocio, E.M.; Rubio, A.; Murillo, I.; Escalante, F.; Aguilera, C.; García-Mateo, A.; de Coca, A.G.; et al. 6q deletion in Waldenström macroglobulinaemia negatively affects time to transformation and survival. Br. J. Haematol. 2021, 192, 843–852. [Google Scholar] [CrossRef]
- Gutiérrez, N.C.; Ocio, E.M.; Rivas, J.D.L.; Maiso, P.; Delgado, M.; Fermiñán, E.; Arcos, M.J.; Sanchez, M.L.; Hernández, J.M.; Miguel, J.F.S. Gene expression profiling of B lymphocytes and plasma cells from Waldenström’s macroglobulinemia: Comparison with expression patterns of the same cell counterparts from chronic lymphocytic leukemia, multiple myeloma and normal individuals. Leukemia 2007, 21, 541–549. [Google Scholar] [CrossRef]
- Chng, W.J.; Schop, R.F.; Price-Troska, T.; Ghobrial, I.; Kay, N.; Jelinek, D.F.; Gertz, M.A.; Dispenzieri, A.; Lacy, M.; Kyle, R.A.; et al. Gene-expression profiling of Waldenstrom macroglobulinemia reveals a phenotype more similar to chronic lymphocytic leukemia than multiple myeloma. Blood 2006, 108, 2755–2763. [Google Scholar] [CrossRef][Green Version]
- Trojani, A.; Greco, A.; Tedeschi, A.; Lodola, M.; Di Camillo, B.; Ricci, F.; Turrini, M.; Varettoni, M.; Rattotti, S.; Morra, E. Microarray Demonstrates Different Gene Expression Profiling Signatures Between Waldenström Macroglobulinemia and IgM Monoclonal Gammopathy of Undetermined Significance. Clin. Lymphoma Myeloma Leuk. 2013, 13, 208–210. [Google Scholar] [CrossRef]
- García-Sanz, R.; Jiménez, C.; Puig, N.; Paiva, B.; Gutiérrez, N.C.; Rodríguez-Otero, P.; Almeida, J.; Miguel, J.S.; Orfão, A.; González, M.; et al. Origin of Waldenstrom’s macroglobulinaemia. Best Pr. Res. Clin. Haematol. 2016, 29, 136–147. [Google Scholar] [CrossRef]
- Nielsen, J.S.; Chang, A.R.; Wick, D.A.; Sedgwick, C.G.; Zong, Z.; Mungall, A.J.; Martin, S.D.; Kinloch, N.N.; Ott-Langer, S.; Brumme, Z.L.; et al. Mapping the human T cell repertoire to recurrent driver mutations in MYD88 and EZH2 in lymphoma. OncoImmunology 2017, 6, e1321184. [Google Scholar] [CrossRef]
- Gustine, J.N.; Xu, L.; Tsakmaklis, N.; Demos, M.G.; Kofides, A.; Chen, J.G.; Liu, X.; Munshi, M.; Guerrera, M.L.; Chan, G.G.; et al. CXCR4 S338X clonality is an important determinant of ibrutinib outcomes in patients with Waldenström macroglobulinemia. Blood Adv. 2019, 3, 2800–2803. [Google Scholar] [CrossRef]
- Baron, M.; Simon, L.; Poulain, S.; Leblond, V. How Recent Advances in Biology of Waldenstrom’s Macroglobu-linemia May Affect Therapy Strategy. Curr. Oncol. Rep. 2019, 21, 27. [Google Scholar] [CrossRef]
- Wasik, A.M.; Nygren, L.; Almestrand, S.; Zong, F.; Flygare, J.; Wennerholm, S.B.; Saft, L.; Andersson, P.; Kimby, E.; Wahlin, B.E.; et al. Perturbations of the endocannabinoid system in mantle cell lymphoma: Correlations to clinical and pathological features. Oncoscience 2014, 1, 550–557. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Paiva, B.; Montes, M.C.; García-Sanz, R.; Ocio, E.M.; Alonso, J.L.; Heras, N.D.L.; Escalante, F.; Cuello, R.; De Coca, A.G.; Galende, J.; et al. Multiparameter flow cytometry for the identification of the Waldenström’s clone in IgM-MGUS and Waldenström’s Macroglobulinemia: New criteria for differential diagnosis and risk stratification. Leukemia 2014, 28, 166–173. [Google Scholar] [CrossRef] [PubMed]
- Paiva, B.D.L.; Corchete, L.A.; Vidriales, M.-B.; García-Sanz, R.; Perez, J.J.; Aires-Mejia, I.; Sanchez, M.-L.; Barcena, P.; Alignani, D.; Jimenez, C.; et al. The cellular origin and malignant transformation of Waldenström macroglobulinemia. Blood 2015, 125, 2370–2380. [Google Scholar] [CrossRef] [PubMed]
- Kotsianidis, I.; Nakou, E.; Spanoudakis, E.; Bouchliou, I.; Moustakidis, E.; Miltiades, P.; Vadikolia, C.M.; Szydlo, R.; Karadimitris, A.; Tsatalas, C. The Diagnostic Value of CD1d Expression in a Large Cohort of Patients With B-Cell Chronic Lymphoproliferative Disorders. Am. J. Clin. Pathol. 2011, 136, 400–408. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Zhao, S.; Karnad, A.; Freeman, J.W. The biology and role of CD44 in cancer progression: Therapeutic implications. J. Hematol. Oncol. 2018, 11, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Hao, M.; Barlogie, B.; Tricot, G.; Liu, L.; Qiu, L.; Shaughnessy, J.D.; Zhan, F.; Qiu, L. Gene Expression Profiling Reveals Aberrant T-cell Marker Expression on Tumor Cells of Waldenström’s Macroglobulinemia. Clin. Cancer Res. 2019, 25, 201–209. [Google Scholar] [CrossRef] [PubMed]
- Moreau, A.-S.; Sebti, Y.; Duhamel, A.; Roccaro, A.M.; Coiteux, V.; Gastinne, T.; Le Friec, G.; Burwick, N.; Amiot, L.; Ho, A.W.; et al. Clinical relevance of soluble HLA class I molecules in Waldenstrom Macroglobulinemia. Eur. J. Haematol. 2008, 80, 503–509. [Google Scholar] [CrossRef]
- Brown, R.; Kabani, K.; Favaloro, J.; Yang, S.; Ho, P.J.; Gibson, J.; Fromm, P.; Suen, H.; Woodland, N.; Nassif, N.; et al. CD86+ or HLA-G+ can be transferred via trogocytosis from myeloma cells to T cells and are associated with poor prognosis. Blood 2012, 120, 2055–2063. [Google Scholar] [CrossRef]
- Gavile, C.M.; Barwick, B.G.; Newman, S.; Neri, P.; Nooka, A.K.; Lonial, S.; Lee, K.P.; Boise, L.H. CD86 regulates myeloma cell survival. Blood Adv. 2017, 1, 2307–2319. [Google Scholar] [CrossRef]
- Paulus, A.; Chitta, K.S.; Wallace, P.K.; Advani, P.P.; Akhtar, S.; Kuranz-Blake, M.; Ailawadhi, S.; Chanan-Khan, A.A. Immunophenotyping of Waldenströms Macroglobulinemia Cell Lines Reveals Distinct Patterns of Surface Antigen Expression: Potential Biological and Therapeutic Implications. PLoS ONE 2015, 10, e0122338. [Google Scholar] [CrossRef]
- Josef, K.; Heidi, M.; Robert, P.; Pavel, K.; Marek, T. Expression of CD66 in non-Hodgkin lymphomas and multiple myeloma. Eur. J. Haematol. 2010, 85, 496–501. [Google Scholar] [CrossRef]
- Xiao, F. Neuromyotonia as an unusual neurological complication of primary Sjögren’s syndrome: Case report and literature review. Clin. Rheumatol. 2016, 36, 481–484. [Google Scholar] [CrossRef]
- Bolger, J.C.; Young, L.S. ADAM22 as a prognostic and therapeutic drug target in the treatment of endo-crine-resistant breast cancer. Vitam. Horm. 2013, 93, 307–321. [Google Scholar] [CrossRef]
- Nobile-Orazio, E. IgM paraproteinaemic neuropathies. Curr. Opin. Neurol. 2004, 17, 599–605. [Google Scholar] [CrossRef]
- Klein, C.J.; Moon, J.S.; Mauermann, M.L.; Zeldenrust, S.R.; Wu, Y.; Dispenzieri, A.; Dyck, P.J. The neuropathies of Waldenström’s macroglobulinemia (WM) and IgM-MGUS. Neurol. Sci. 2011, 38, 289–295. [Google Scholar] [CrossRef]
- Sulzmaier, F.J.; Jean, C.; Schlaepfer, D.D. FAK in cancer: Mechanistic findings and clinical applications. Nat. Rev. Cancer 2014, 14, 598–610. [Google Scholar] [CrossRef]
- Yin, B. Focal adhesion kinase as a target in the treatment of hematological malignancies. Leuk. Res. 2011, 35, 1416–1418. [Google Scholar] [CrossRef]
- Ghobrial, I.M.; Maiso, P.; Azab, A.; Liu, Y.; Zhang, Y.; Issa, G.; Azab, F.; Sacco, A.; Quang, P.; Ngo, H.; et al. The bone marrow microenvironment in Waldenstrom macroglobulinemia. Ther. Adv. Hematol. 2011, 2, 267–272. [Google Scholar] [CrossRef]
- Guinn, B.-A.; Bullinger, L.; Thomas, N.S.B.; Mills, K.I.; Greiner, J. SSX2IP expression in acute myeloid leukaemia: An association with mitotic spindle failure in t(8;21), and cell cycle in t(15;17) patients. Br. J. Haematol. 2007, 140, 250–251. [Google Scholar] [CrossRef]
- Wang, W.; Lin, P. Lymphoplasmacytic lymphoma and Waldenström macroglobulinaemia: Clinicopathological features and differential diagnosis. Pathology 2020, 52, 6–14. [Google Scholar] [CrossRef] [PubMed]
- Growková, K.; Kryukova, E.; Kufová, Z.; Filipová, J.; Ševčíková, T.; Říhová, L.; Kaščák, M.; Kryukov, F.; Hájek, R. Waldenström′s macroglobulinemia: Two malignant clones in a monoclonal disease? Molecular background and clinical reflection. Eur. J. Haematol. 2017, 99, 469–478. [Google Scholar] [CrossRef] [PubMed]
- Ek, S.; Högerkorp, C.M.; Dictor, M.; Ehinger, M.; Borrebaeck, C.A. Mantle cell lymphomas express a distinct ge-netic signature affecting lymphocyte trafficking and growth regulation as compared with subpopulations of normal human B cells. Cancer. Res. 2002, 62, 4398–4405. [Google Scholar] [PubMed]
- Nugent, A.; Proia, R.L. The role of G protein-coupled receptors in lymphoid malignancies. Cell. Signal. 2017, 39, 95–107. [Google Scholar] [CrossRef]
- Gerber, J.M.; Gucwa, J.L.; Esopi, D.; Gürel, M.; Haffner, M.C.; Vala, M.; Nelson, W.G.; Jones, R.J.; Yegnasubramanian, S. Genome-wide comparison of the transcriptomes of highly enriched normal and chronic myeloid leukemia stem and progenitor cell populations. Oncotarget 2013, 4, 715–728. [Google Scholar] [CrossRef]
- Formoso, K.; Susperreguy, S.; Freichel, M.; Birnbaumer, L. RNA-seq analysis reveals TRPC genes to impact an unexpected number of metabolic and regulatory pathways. Sci. Rep. 2020, 10, 1–20. [Google Scholar] [CrossRef]
- Argyropoulos, K.V.; Vogel, R.; Ziegler, C.; Altan-Bonnet, G.; Velardi, E.; Calafiore, M.; Dogan, A.; Arcila, M.; Patel, M.; Knapp, K.; et al. Clonal B cells in Waldenström’s macroglobulinemia exhibit functional features of chronic active B-cell receptor signaling. Leukemia 2016, 30, 1116–1125. [Google Scholar] [CrossRef][Green Version]
- Sekiguchi, N.; Nomoto, J.; Nagata, A.; Kiyota, M.; Fukuda, I.; Yamada, K.; Takezako, N.; Kobayashi, Y. Gene Expression Profile Signature of Aggressive Waldenström Macroglobulinemia with Chromosome 6q Deletion. BioMed Res. Int. 2018, 2018, 6728128. [Google Scholar] [CrossRef]
- Efremov, D.G.; Turkalj, S.; Laurenti, L. Mechanisms of B Cell Receptor Activation and Responses to B Cell Receptor Inhibitors in B Cell Malignancies. Cancers 2020, 12, 1396. [Google Scholar] [CrossRef]
- Jiménez, C.; Alonso-Álvarez, S.; Alcoceba, M.; Ordóñez, G.R.; García-Álvarez, M.; Prieto-Conde, M.I.; Chillón, M.C.; Balanzategui, A.; Corral, R.; Marín, L.A.; et al. From Waldenström’s macroglobulinemia to aggressive diffuse large B-cell lymphoma: A whole-exome analysis of abnormalities leading to transformation. Blood Cancer J. 2017, 7, e591. [Google Scholar] [CrossRef]
- Munshi, M.; Liu, X.; Chen, J.G.; Xu, L.; Tsakmaklis, N.; Demos, M.G.; Kofides, A.; Guerrera, M.L.; Jimenez, C.; Chan, G.G.; et al. SYK is activated by mutated MYD88 and drives pro-survival signaling in MYD88 driven B-cell lymphomas. Blood Cancer J. 2020, 10, 12. [Google Scholar] [CrossRef]
- Safavi, S.; Hansson, M.; Karlsson, K.; Biloglav, A.; Johansson, B.; Paulsson, K. Novel gene targets detected by genomic profiling in a consecutive series of 126 adults with acute lymphoblastic leukemia. Haematologica 2015, 100, 55–61. [Google Scholar] [CrossRef]
- Castillo, J.J.; Gustine, J.N.; Meid, K.; Dubeau, T.; Severns, P.; Xu, L.; Yang, G.; Hunter, Z.R.; Treon, S.P. Low levels of von Willebrand markers associate with high serum IgM levels and improve with response to therapy, in patients with Waldenström macroglobulinaemia. Br. J. Haematol. 2018, 184, 1011–1014. [Google Scholar] [CrossRef]
- Jalali, S.; Ansell, S.M. The Bone Marrow Microenvironment in Waldenström Macroglobulinemia. Hematol. Clin. N. Am. 2018, 32, 777–786. [Google Scholar] [CrossRef]
- Hunter, Z.R.; Xu, L.; Yang, G.; Tsakmaklis, N.; Vos, J.M.; Liu, X.; Chen, J.; Manning, R.J.; Chen, J.G.; Brodsky, P.; et al. Transcriptome sequencing reveals a profile that corresponds to genomic variants in Waldenström macroglobulinemia. Blood 2016, 128, 827–838. [Google Scholar] [CrossRef]
- Zanwar, S.; Abeykoon, J.P.; Kapoor, P. Novel Treatment Strategies in the Management of Waldenström Macroglobulinemia. Curr. Hematol. Malig. Rep. 2020, 15, 31–43. [Google Scholar] [CrossRef]
- De Brachène, A.C.; Demoulin, J.-B. FOXO transcription factors in cancer development and therapy. Cell. Mol. Life Sci. 2016, 73, 1159–1172. [Google Scholar] [CrossRef]
- Han, W.; Matissek, S.J.; Jackson, D.A.; Sklavanitis, B.; Elsawa, S.F. Targeting IL-6 receptor reduces IgM levels and tumor growth in Waldenström macroglobulinemia. Oncotarget 2019, 10, 3400–3407. [Google Scholar] [CrossRef][Green Version]
- Wada, N.; Zhan, M.; Hori, Y.; Honma, K.; Ikeda, J.-I.; Morii, E. Characterization of subpopulation lacking both B-cell and plasma cell markers in Waldenstrom macroglobulinemia cell line. Lab. Investig. 2013, 94, 79–88. [Google Scholar] [CrossRef]
- Samba-Mondonga, M.; Calvé, A.; Mallette, F.A.; Santos, M.M. MyD88 Regulates the Expression of SMAD4 and the Iron Regulatory Hormone Hepcidin. Front. Cell Dev. Biol. 2018, 6. [Google Scholar] [CrossRef]
- Akyurek, N.; Ren, Y.; Rassidakis, G.Z.; Schlette, E.J.; Medeiros, L.J. Expression of inhibitor of apoptosis proteins in B-cell non-Hodgkin and Hodgkin lymphomas. Cancer 2006, 107, 1844–1851. [Google Scholar] [CrossRef]
- Arribas, A.J.; Campos-Martín, Y.; Gómez-Abad, C.; Algara, P.; Sanchez-Beato, M.; Rodriguez-Pinilla, M.S.; Montes-Moreno, S.; Martinez, N.; Alves-Ferreira, J.; Piris, M.A.; et al. Nodal marginal zone lymphoma: Gene expression and miRNA profiling identify diagnostic markers and potential therapeutic targets. Blood 2012, 119, e9–e21. [Google Scholar] [CrossRef]
- Di Rorà, A.G.L.; Bocconcelli, M.; Ferrari, A.; Terragna, C.; Bruno, S.; Imbrogno, E.; Beeharry, N.; Robustelli, V.; Ghetti, M.; Napolitano, R.; et al. Synergism Through WEE1 and CHK1 Inhibition in Acute Lympho-blastic Leukemia. Cancers 2019, 11, 1654. [Google Scholar] [CrossRef]
- Di Tullio, A.; Rouault-Pierre, K.; Abarrategi, A.; Mian, S.; Grey, W.; Gribben, J.; Stewart, A.; Blackwood, E.; Bonnet, D. The combination of CHK1 inhibitor with G-CSF overrides cytarabine resistance in human acute myeloid leukemia. Nat. Commun. 2017, 8, 1679. [Google Scholar] [CrossRef]
- Pals, S.T.; Kersten, M.J.; Spaargaren, M. Targeting cell adhesion and homing as strategy to cure Waldenström’s macroglobulinemia. Best Pr. Res. Clin. Haematol. 2016, 29, 161–168. [Google Scholar] [CrossRef]
- Zhang, X.-D.; Baladandayuthapani, V.; Lin, H.; Mulligan, G.; Li, B.; Esseltine, D.-L.W.; Qi, L.; Xu, J.; Hunziker, W.; Barlogie, B.; et al. Tight Junction Protein 1 Modulates Proteasome Capacity and Proteasome Inhibitor Sensitivity in Multiple Myeloma via EGFR/JAK1/STAT3 Signaling. Cancer Cell 2016, 29, 639–652. [Google Scholar] [CrossRef]
- Owen, R.G.; Treon, S.P.; Al-Katib, A.; Fonseca, R.; Greipp, P.R.; McMaster, M.L.; Morra, E.; Pangalis, G.A.; Miguel, J.F.S.; Branagan, A.R.; et al. Clinicopathological definition of Waldenstrom’s macroglobulinemia: Consensus Panel Recommendations from the Second International Workshop on Waldenstrom’s Macroglobulinemia. Semin. Oncol. 2003, 30, 110–115. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| ZNF Differentially Expressed in CD19+ Cells | ||||||||
|---|---|---|---|---|---|---|---|---|
| Gene Symbol | Pattern | q Value | Mean WM | Mean MGUS | Mean CTRL | FC (p Value) WM vs. MGUS | FC (p Value) MGUS vs. CTRL | FC (p Value) WM vs. CTRL |
| ZNF804A | p1 | 0.0021 | 8.36 | 6.01 | 5.12 | 5.12 (0.00036) | 1.85 (0.184) | 9.48 (0.000003) |
| ZNF215 | p1 | 0.0273 | 3.44 | 2.85 | 2.60 | 1.51 (0.000202) | 1.19 (0.14) | 1.79 (0.000784) |
| IKZF2 | p2 | 0.0453 | 6.49 | 7.72 | 8.83 | −2.33 (0.025399) | −2.16 (0.026) | −5.05 (0.000026) |
| Genes Differentially Expressed in Pattern 2 CD19+ with Very High FC | ||||||||
|---|---|---|---|---|---|---|---|---|
| Gene Symbol | Pattern | q Value | Mean WM | Mean MGUS | Mean CTRL | FC (p Value) WM vs. MGUS | FC (p Value) MGUS vs. CTRL | FC (p Value) WM vs. CTRL |
| HIST1H1B | p2 | 0.0093 | 6.41 | 7.18 | 8.85 | −1.71 (0.118) | −3.18 (0.022) | −5.45 (0.000477) |
| ADARB1 | p2 | 0.0127 | 5.98 | 7.11 | 8.21 | −2.19 (0.011) | −2.16 (0.015) | −4.72 (0.000004) |
| ADAM28 | p2 | 0.0171 | 5.71 | 6.76 | 7.44 | −2.08 (0.008) | −1.6 (0.154) | −3.33 (0.000568) |
| APBB2 | p2 | 0.0248 | 4.61 | 5.88 | 7.07 | −2.42 (0.024) | −2.27 (0.034) | −5.5 (0.000009) |
| EZH2 | p2 | 0.0374 | 7.25 | 7.37 | 8.79 | −1.09 (0.733) | −2.67 (0.002) | −2.91 (0.000034) |
| DEGs Indicating a Possible Risk of Progression of IgM MGUS to WM in B Cells | ||||||||
|---|---|---|---|---|---|---|---|---|
| Gene Symbol | Pattern | q Value | Mean WM | Mean MGUS | Mean CTRL | FC (p Value) WM vs. MGUS | FC (p Value) MGUS vs. CTRL | FC (p Value) WM vs. CTRL |
| HIST1H1B | p2 | 0.009 | 6.41 | 7.18 | 8.85 | −1.71 (0.118) | −3.18 (0.022) | −5.45 (0.000477) |
| EZH2 | p2 | 0.037 | 7.25 | 7.37 | 8.79 | −1.09 (0.733) | −2.67 (0.002) | −2.91 (0.000034) |
| CHEK1 | p2 | 0.040 | 4.81 | 5.14 | 6.49 | −1.26 (0.411) | −2.55 (0.004) | −3.22 (0.000024) |
| LEF1 | p2 | 0.041 | 5.24 | 5.82 | 7.19 | −1.5 (0.19) | −2.58 (0.01) | −3.86 (0.000136) |
| ADAM23 | p2 | 0.045 | 4.12 | 4.84 | 6.15 | −1.64 (0.137) | −2.48 (0.011) | −4.07 (0.000032) |
| RASGRP3 | p3 | 0.045 | 4.57 | 4.47 | 5.60 | 1.07 (0.687) | −2.19 (0.001) | −2.05 (0.002118) |
| ADRB2 | p1 | 0.047 | 7.33 | 6.98 | 6.13 | 1.28 (0.226) | 1.8 (0.005) | 2.31 (0.000003) |
| PIK3AP1 | p2 | 0.047 | 10.15 | 10.24 | 10.84 | −1.06 (0.565) | −1.51 (0.002) | −1.61 (0.000206) |
| CDHR3 | p2 | 0.048 | 4.82 | 5.31 | 6.29 | −1.4 (0.142) | −1.98 (0.028) | −2.77 (0.001467) |
| Genes Differentially Expressed in CD138+ | ||||||||
|---|---|---|---|---|---|---|---|---|
| Gene Symbol | Pattern | q Value | Mean WM | Mean MGUS | Mean CTRL | FC (p Value) WM vs. MGUS | FC (p Value) MGUS vs. CTRL | FC (p Value) WM vs. CTRL |
| TJP1 | p2 | 0.0005 | 3.62 | 5.02 | 6.24 | −2.65 (0.002) | −2.33 (0.013) | −6.18 (0.0000003) |
| ESRP1 | p2 | 0.002 | 4.44 | 5.4 | 6.44 | −1.94 (0.002) | −2.07 (0.008) | −4.02 (0.000005) |
| FGF2 | p2 | 0.0021 | 2.25 | 2.45 | 3.26 | −1.15 (0.072) | −1.75 (0.005) | −2.01 (0.000822) |
| SEPT10 | p2 | 0.0082 | 3.93 | 5.02 | 6.02 | −2.14 (0.004) | −1.99 (0.025) | −4.26 (0.000007) |
| GREB1 | p3 | 0.0092 | 1.92 | 1.75 | 3.46 | 1.12 (0.005) | −3.28 (0.005) | −2.92 (0.013437) |
| PERP | p2 | 0.0097 | 3.39 | 4.28 | 5.51 | −1.85 (0.03) | −2.35 (0.002) | −4.4 (0.000000002) |
| NFIB | p2 | 0.0134 | 2.46 | 3.38 | 4.06 | −1.9 (0.022) | −1.59 (0.138) | −3.04 (0.000078) |
| CALCRL | p2 | 0.0164 | 3.05 | 3.87 | 4.36 | −1.77 (0.008) | −1.41 (0.171) | −2.48 (0.000142) |
| PPARGC1A | p2 | 0.0191 | 3.32 | 4.49 | 4.49 | −2.24 (0.004) | −1 (0.993) | −2.25 (0.000901) |
| TUBB2B | p2 | 0.0225 | 3.26 | 3.31 | 4.85 | −1.04 (0.729) | −2.92 (0.006) | −3.03 (0.0065) |
| PLA2G2D | p2 | 0.0276 | 4.85 | 5.6 | 6.62 | −1.69 (0.019) | −2.02 (0.003) | −3.41 (0.0000004) |
| PARD3 | p2 | 0.0279 | 5.23 | 5.95 | 6.56 | −1.65 (0.005) | −1.52 (0.071) | −2.52 (0.000226) |
| OVOL1 | p2 | 0.0293 | 5.49 | 6.03 | 6.54 | −1.46 (0.016) | −1.42 (0.059) | −2.07 (0.000058) |
| TTLL7 | p2 | 0.0341 | 2.92 | 3.18 | 4.07 | −1.2 (0.175) | −1.86 (0.011) | −2.22 (0.001204) |
| GBA3 | p4 | 0.041 | 3.43 | 5.1 | 4.58 | −3.2 (0.004) | 1.44 (0.316) | −2.22 (0.001208) |
| MYO5B | p2 | 0.0422 | 5.03 | 5.66 | 6.37 | −1.55 (0.025) | −1.63 (0.043) | −2.52 (0.000136) |
| SYNM | p2 | 0.0426 | 4.84 | 5.81 | 5.91 | −1.97 (0.003) | −1.07 (0.75) | −2.11 (0.000054) |
| KIAA1804 | p2 | 0.0428 | 3.92 | 4.28 | 5.28 | −1.29 (0.216) | −1.99 (0.003) | −2.57 (0.0000005) |
| NRG3 | p4 | 0.043 | 3.01 | 4.05 | 4.03 | −2.06 (0.002) | 1.01 (0.959) | −2.03 (0.007143) |
| KCTD1 | p2 | 0.0434 | 5.27 | 5.77 | 6.35 | −1.41 (0.024) | −1.5 (0.018) | −2.11 (0.000012) |
| CNTN1 | p2 | 0.0435 | 2.56 | 3.03 | 3.39 | −1.39 (0.011) | −1.29 (0.163) | −1.79 (0.00149) |
| LAPTM4B | p2 | 0.0437 | 4.71 | 5.27 | 6.49 | −1.48 (0.086) | −2.32 (0.001) | −3.42 (0.000001) |
| CADPS2 | p2 | 0.0438 | 5.47 | 6.87 | 8.12 | −2.63 (0.011) | −2.38 (0.014) | −6.26 (0.0000005) |
| ARHGAP32 | p2 | 0.0451 | 4.02 | 4.85 | 5.4 | −1.77 (0.002) | −1.47 (0.061) | −2.61 (0.000052) |
| GPR125 | p2 | 0.0453 | 3.58 | 4.05 | 4.93 | −1.38 (0.031) | −1.84 (0.012) | −2.55 (0.000346) |
| UCHL1 | p2 | 0.0454 | 5.24 | 5.86 | 6.52 | −1.54 (0.065) | −1.58 (0.051) | −2.42 (0.000002) |
| MARC2 | p2 | 0.0461 | 4.8 | 6.18 | 6.46 | −2.59 (0.001) | −1.22 (0.433) | −3.17 (0.000017) |
| TTLL7 | p2 | 0.047 | 4.61 | 5.45 | 6.27 | −1.79 (0.03) | −1.76 (0.055) | −3.15 (0.00004) |
| MYLK | p2 | 0.0486 | 6.13 | 6.96 | 7.53 | −1.78 (0.007) | −1.49 (0.072) | −2.64 (0.000028) |
| Gene Symbol | Pattern CD19+ | Pattern CD138+ | CD19+ | CD138+ | ||||
|---|---|---|---|---|---|---|---|---|
| FC WM vs. MGUS | FC MGUS vs. CTRL | FC WM vs. CTRL | FC WM vs. MGUS | FC MGUS vs. CTRL | FC WM vs. CTRL | |||
| ARHGAP32 | p2 | p2 | −2.52 | −1.55 | −3.9 | −1.77 | −1.47 | −2.61 |
| MARC2 | p2 | p2 | −2.47 | −1.51 | −3.72 | −2.59 | −1.22 | −3.17 |
| LAPTM4B | p2 | p2 | −1.76 | −1.57 | −2.76 | −1.48 | −2.32 | −3.42 |
| PLA2G2D | p2 | p2 | −1.5 | −1.1 | −1.65 | −1.69 | −2.02 | −3.41 |
| KIAA1804 | p2 | p2 | −1.36 | −2.02 | −2.76 | −1.29 | −1.99 | −2.57 |
| MYLK | p2 | p2 | −2.04 | −1.28 | −2.61 | −1.78 | −1.49 | −2.64 |
| SYNM | p2 | p2 | −1.59 | −1.15 | −1.84 | −1.97 | −1.07 | −2.11 |
| TJP1 | p4 | p2 | −1.33 | 1.09 | −1.22 | −2.65 | −2.33 | −6.18 |
| GBA3 | p4 | p2 | −1.87 | 1.51 | −1.24 | −3.54 | 1.44 | −2.46 |
| NFIB | p4 | p2 | −1.77 | 1.12 | −1.59 | −1.9 | −1.59 | −3.04 |
| PPARGC1A | p4 | p2 | −1.27 | 1.15 | −1.1 | −2.24 | −1 | −2.25 |
| Parameter | Total n = 49 | WM n = 36 | IgM MGUS n = 13 |
|---|---|---|---|
| Median age (years) | 68 | 69 | 67 |
| Sex n (%) | |||
| Male | 20 (42) | 14 (39) | 6 (50) |
| Female | 29 (58) | 22 (61) | 7 (50) |
| % of bone marrow involvement (median) | 20 | 30 | 2 |
| M-protein level (g/dL) median | 0.8 | 1.1 | 0.3 |
| Light chain | |||
| κ n (%) | 35 (71) | 25 (69) | 10 (77) |
| λ n (%) | 9 (18) | 7 (19) | 2 (15) |
| Both n (%) | 3 (6) | 3 (8) | 0 (0) |
| Missing data n | 2 | 1 | 1 |
| Haemoglobin (g/dL) median | 12.2 | 11.3 | 12.9 |
| Beta2 microglobulin (μg/mL) median (interval) | 2.9 (2–8.24) | 3.1 (1.87–8.24) | 2.3 (2–7.5) |
| Lactate dehydrogenase (U/L, UL 1 480) median (interval) | 200 (85–670) | 200 (85–670) | 200 (156–325) |
| Presence of BJ 2 proteinuria n (%) | 19 (39) | 18 (50) | 1 (8) |
| Presence of immunoparesis n (%) | 10 (20) | 10 (28) | 0 (0) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Trojani, A.; Di Camillo, B.; Bossi, L.E.; Leuzzi, L.; Greco, A.; Tedeschi, A.; Frustaci, A.M.; Deodato, M.; Zamprogna, G.; Beghini, A.; et al. Identification of a Candidate Gene Set Signature for the Risk of Progression in IgM MGUS to Smoldering/Symptomatic Waldenström Macroglobulinemia (WM) by a Comparative Transcriptome Analysis of B Cells and Plasma Cells. Cancers 2021, 13, 1837. https://doi.org/10.3390/cancers13081837
Trojani A, Di Camillo B, Bossi LE, Leuzzi L, Greco A, Tedeschi A, Frustaci AM, Deodato M, Zamprogna G, Beghini A, et al. Identification of a Candidate Gene Set Signature for the Risk of Progression in IgM MGUS to Smoldering/Symptomatic Waldenström Macroglobulinemia (WM) by a Comparative Transcriptome Analysis of B Cells and Plasma Cells. Cancers. 2021; 13(8):1837. https://doi.org/10.3390/cancers13081837
Chicago/Turabian StyleTrojani, Alessandra, Barbara Di Camillo, Luca Emanuele Bossi, Livia Leuzzi, Antonino Greco, Alessandra Tedeschi, Anna Maria Frustaci, Marina Deodato, Giulia Zamprogna, Alessandro Beghini, and et al. 2021. "Identification of a Candidate Gene Set Signature for the Risk of Progression in IgM MGUS to Smoldering/Symptomatic Waldenström Macroglobulinemia (WM) by a Comparative Transcriptome Analysis of B Cells and Plasma Cells" Cancers 13, no. 8: 1837. https://doi.org/10.3390/cancers13081837
APA StyleTrojani, A., Di Camillo, B., Bossi, L. E., Leuzzi, L., Greco, A., Tedeschi, A., Frustaci, A. M., Deodato, M., Zamprogna, G., Beghini, A., & Cairoli, R. (2021). Identification of a Candidate Gene Set Signature for the Risk of Progression in IgM MGUS to Smoldering/Symptomatic Waldenström Macroglobulinemia (WM) by a Comparative Transcriptome Analysis of B Cells and Plasma Cells. Cancers, 13(8), 1837. https://doi.org/10.3390/cancers13081837