Alteration of Metabolic Conditions Impacts the Regulation of IGF-II/H19 Imprinting Status in Prostate Cancer

 ,
,  , , ,
, , , 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
2. Results
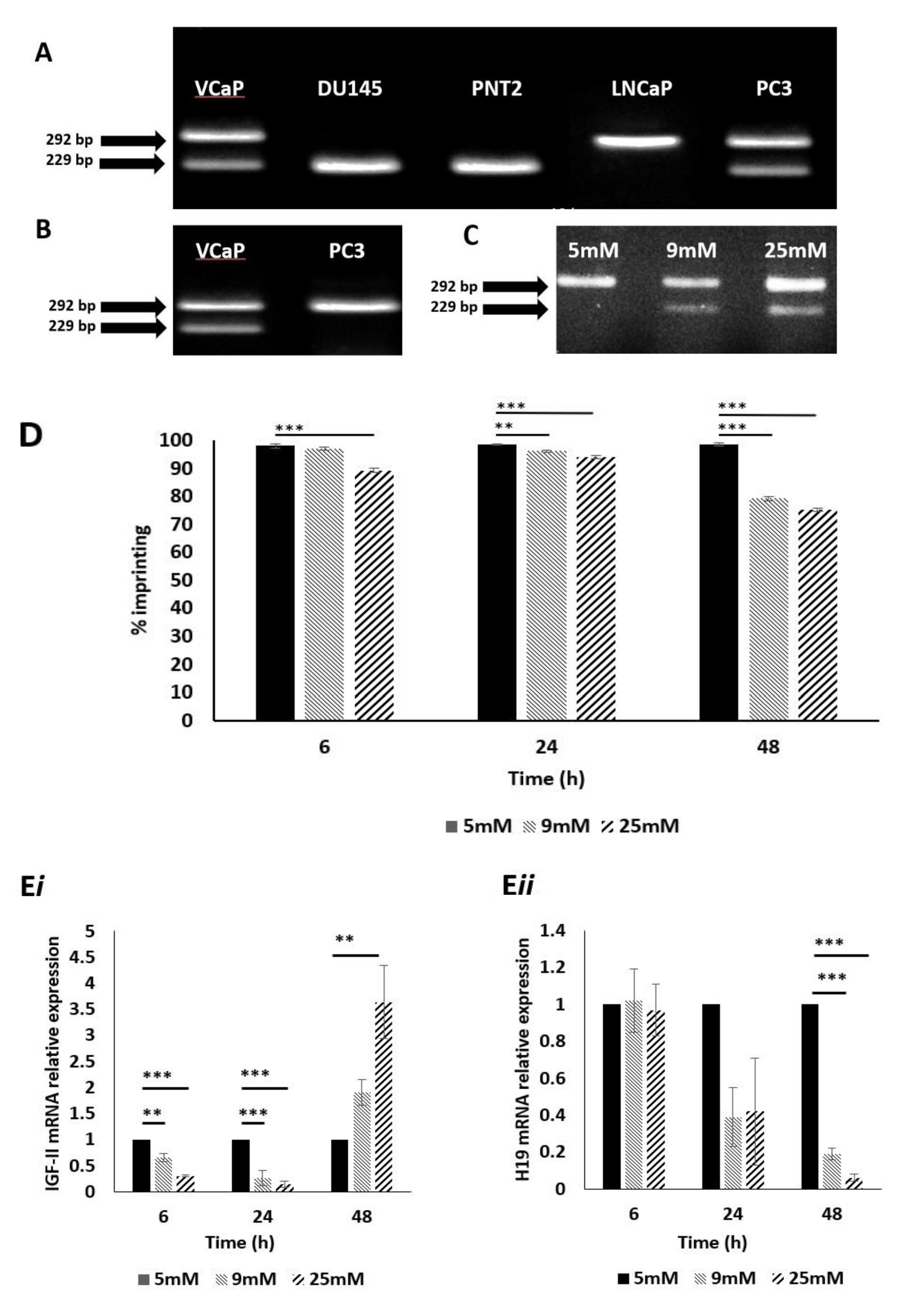
2.1. Identification of Prostate Cell Line Suitability
2.2. Confirmation of IGF-II Imprinting Status in the PC3 and VCaP Cell Lines
2.3. The Impact of Altered Levels of Glucose on IGF-II Imprinting Status
2.4. The Impact of Altered Levels of Glucose on IGF-II Imprinting Percentage
2.5. The Impact of Altered Levels of Glucose on IGF-II mRNA Expression
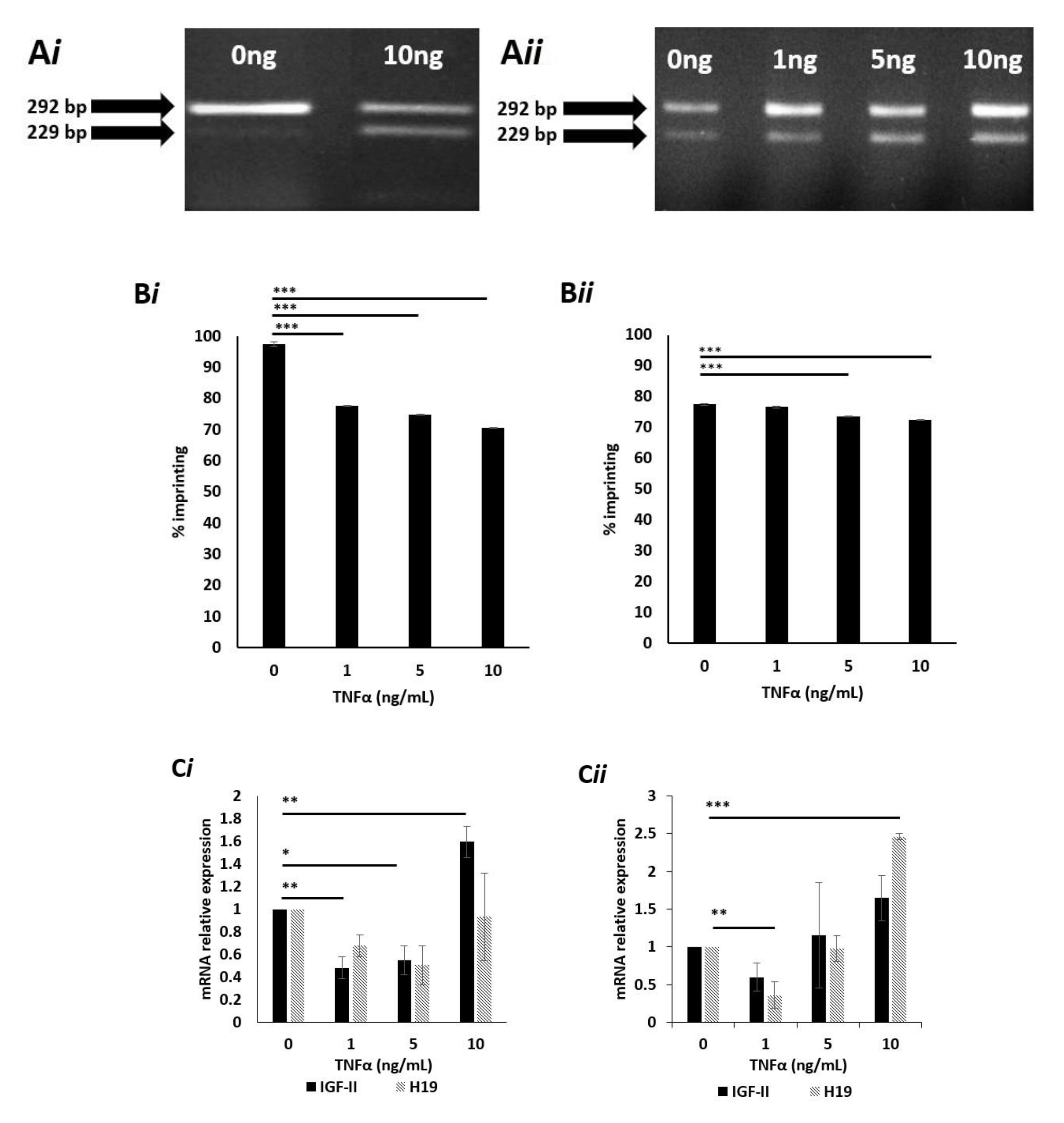
2.6. Effects of TNFα on IGF-II Imprinting Status
2.7. Effects of High Dose TNFα on the Degree of IGF-II Imprinting in Cells Cultured in Normal (5mM) Glucose Media
2.8. Effects of TNFα on IGF-II mRNA Expression in Cells Cultured in Normal (5 mM) Glucose Media
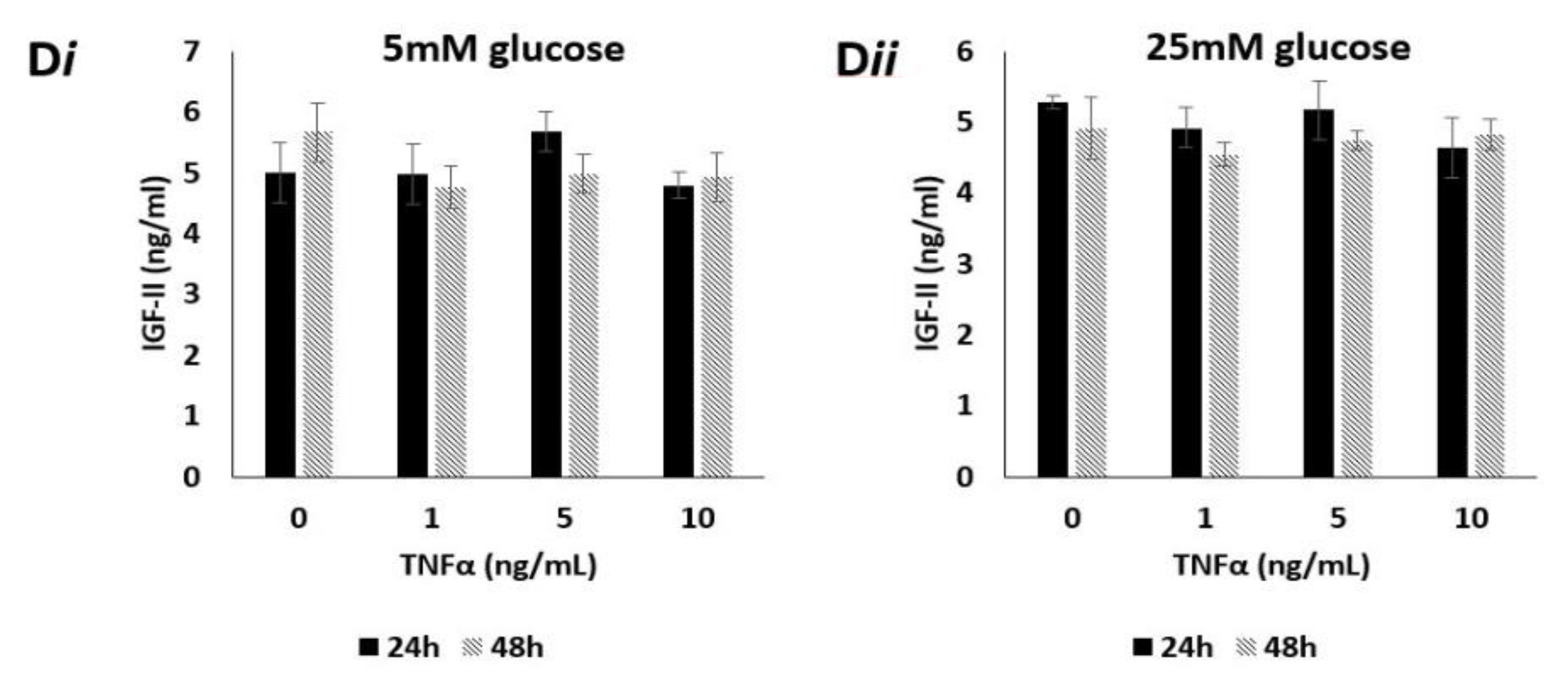
2.9. Effects of TNFα on Levels of Secreted IGF-II Peptide
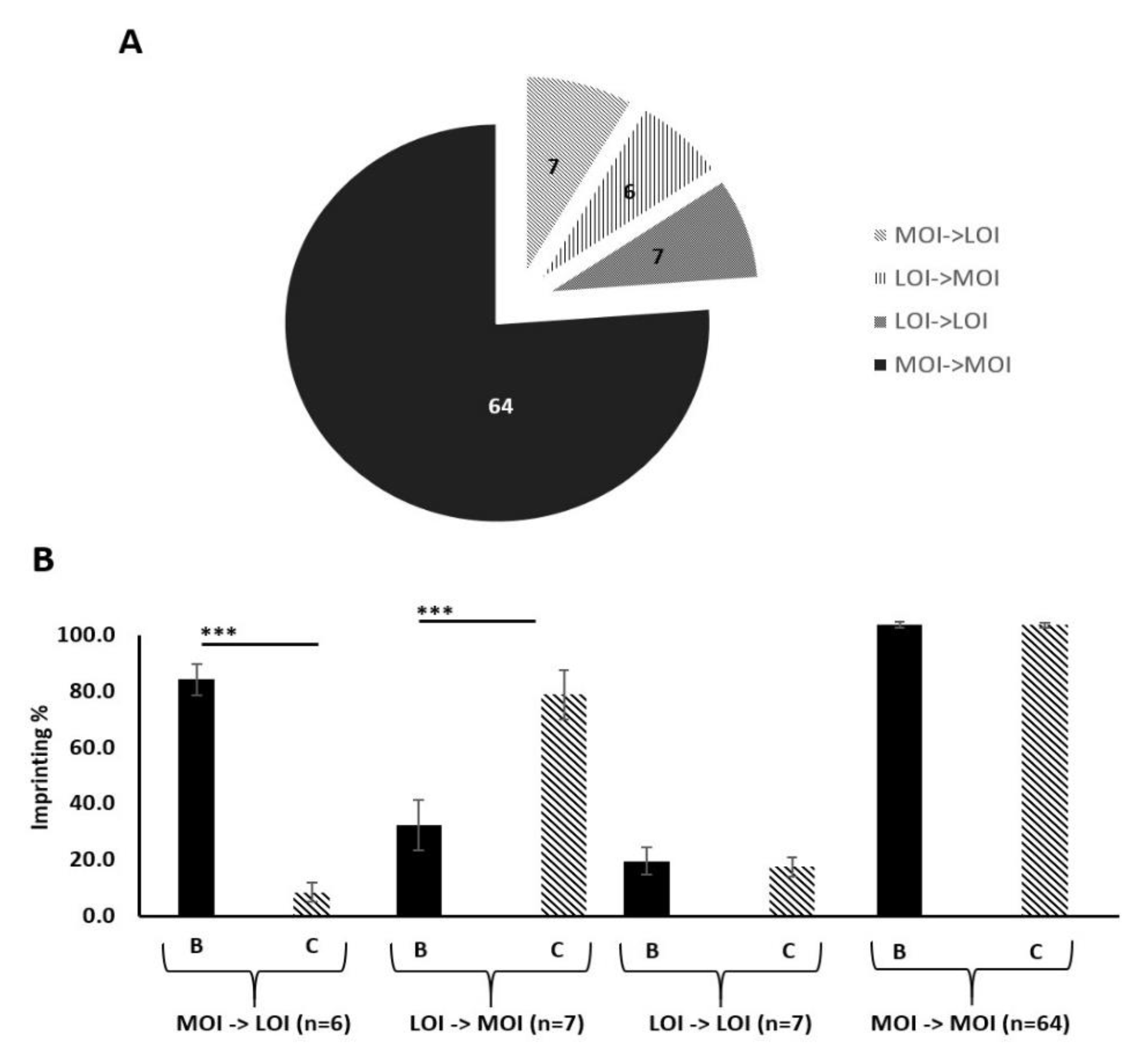
2.10. IGF-II Imprinting Status Does Not Significantly Vary Between Benign and Malignant Paired Prostate Tissue Samples
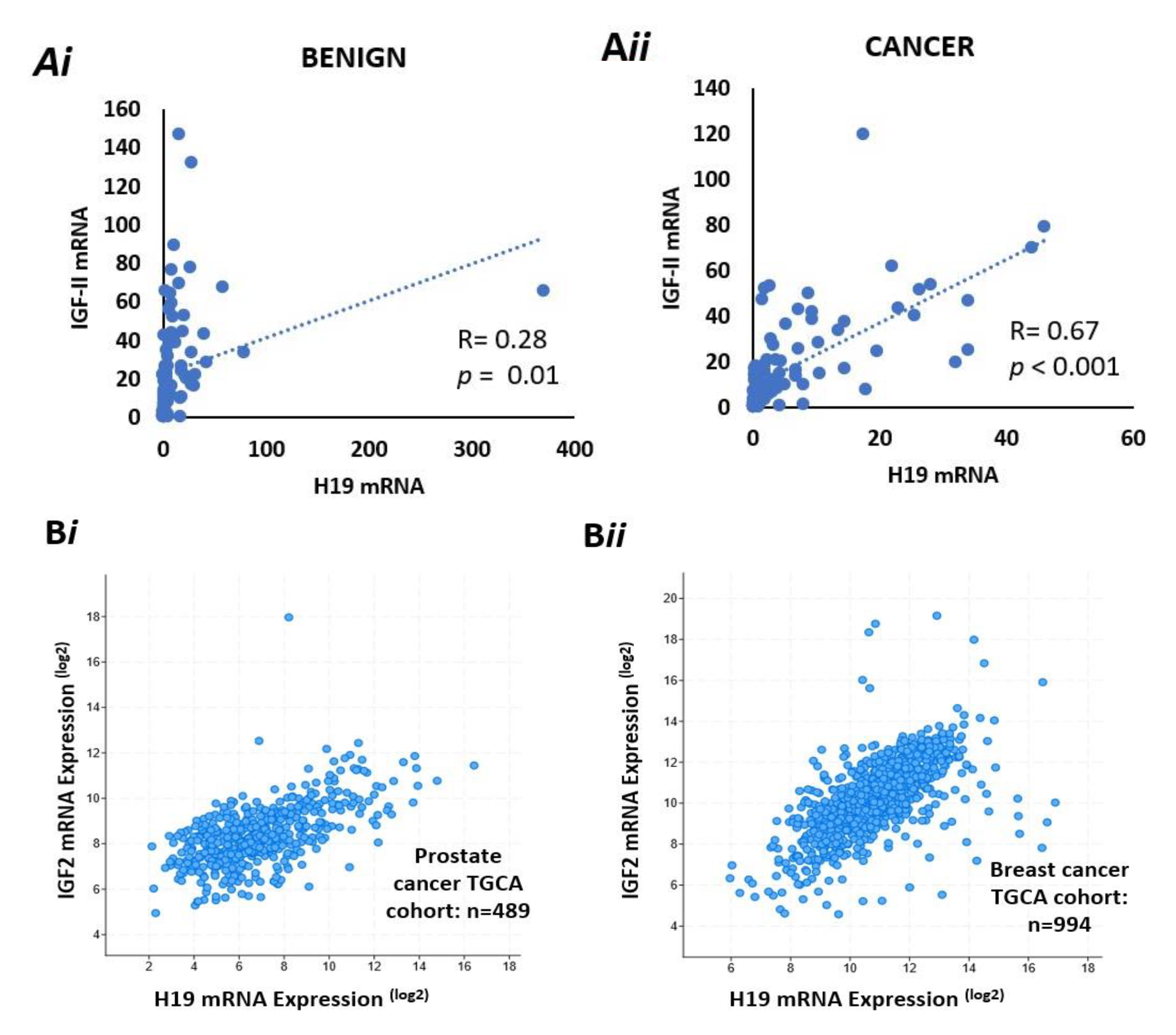
2.11. There Is a Positive Correlation Between IGF-II and H19 mRNA Expression in Benign and Malignant Prostate Tissue
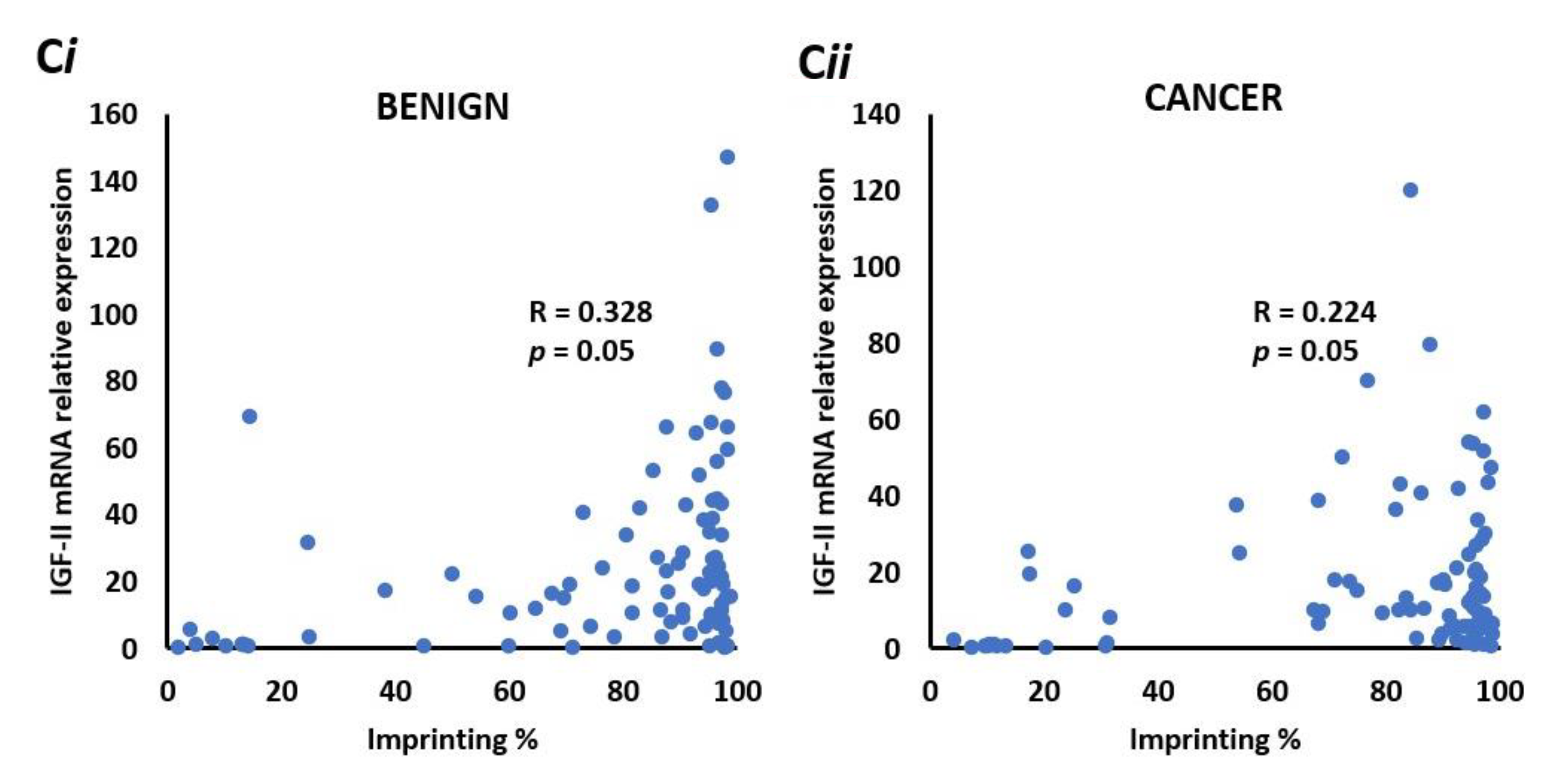
2.12. There is a Positive Correlation between IGF-II mRNA and the Degree of Imprinting in Benign and Malignant Prostate Tissue
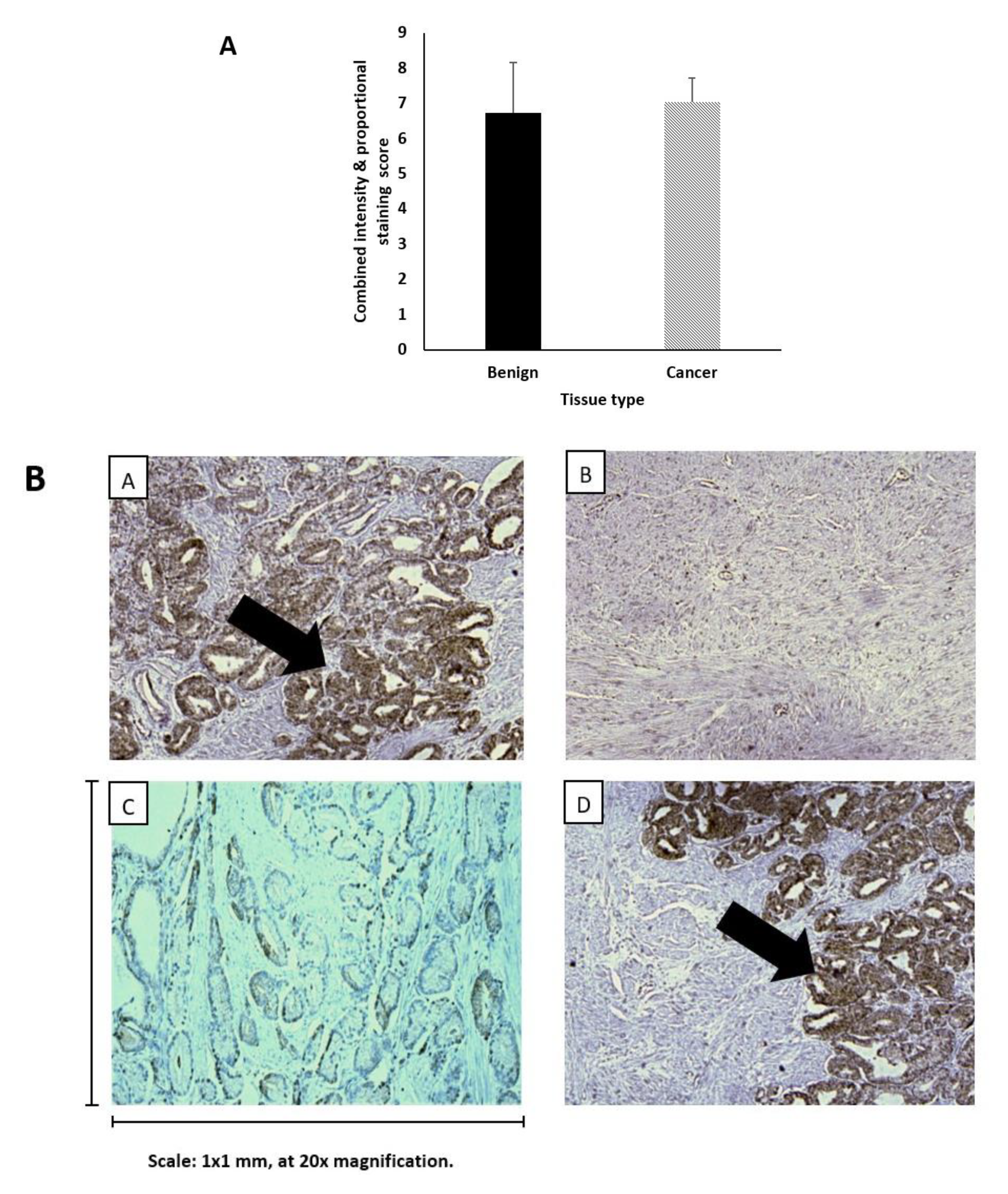
2.13. Levels of IGF-II Peptide Do not Differ between Benign and Malignant Prostate Tissue
3. Discussion
4. Methods
4.1. Prostate Cell Lines
4.2. Isolation of Nucleic Acids
4.3. Preparation of cDNA
4.4. Genotyping of Cell Lines, Using Restriction Fragment Length Polymorphism (RFLP) Analysis
4.5. Dosing Experiments
4.6. Quantitative PCR (qPCR)
4.7. Pyrosequencing
4.8. Radioimmunoassay (RIA)
4.9. Immunohistochemistry (IHC)
4.10. Digital Droplet PCR (ddPCR)
4.11. Use of cBioPortal for Cancer Genomics to Examine co-expression of IHG-II & H19 mRNA
4.12. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2016. CA Cancer J. Clin. 2016, 66, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Barber, L.; Gerke, T.; Markt, S.C.; Peisch, S.F.; Wilson, K.M.; Ahearn, T.; Giovannucci, E.; Parmigiani, G.; Mucci, L.A. Family History of Breast or Prostate Cancer and Prostate Cancer Risk. Clin. Cancer Res. 2018, 24, 5910–5917. [Google Scholar] [CrossRef] [PubMed]
- Jones, A.L.; Chinegwundoh, F. Update on prostate cancer in black men within the UK. Ecancermedicalscience 2014, 8, 455. [Google Scholar] [CrossRef]
- Bell, K.J.; Del Mar, C.; Wright, G.; Dickinson, J.; Glasziou, P. Prevalence of incidental prostate cancer: A systematic review of autopsy studies. Int. J. Cancer 2015, 137, 1749–1757. [Google Scholar] [CrossRef] [PubMed]
- Lavalette, C.; Tretarre, B.; Rebillard, X.; Lamy, P.J.; Cenee, S.; Menegaux, F. Abdominal obesity and prostate cancer risk: Epidemiological evidence from the EPICAP study. Oncotarget 2018, 9, 34485–34494. [Google Scholar] [CrossRef]
- Goldgar, D.E.; Easton, D.F.; Cannon-Albright, L.A.; Skolnick, M.H. Systematic population-based assessment of cancer risk in first-degree relatives of cancer probands. J. Natl. Cancer Inst. 1994, 86, 1600–1608. [Google Scholar] [CrossRef]
- Benafif, S.; Eeles, R. Genetic predisposition to prostate cancer. Br. Med. Bull. 2016, 120, 75–89. [Google Scholar] [CrossRef]
- Rebbeck, T.R. Prostate Cancer Disparities by Race and Ethnicity: From Nucleotide to Neighborhood. Cold Spring Harb. Perspect. Med. 2018, 8. [Google Scholar] [CrossRef]
- Taitt, H.E. Global Trends and Prostate Cancer: A Review of Incidence, Detection, and Mortality as Influenced by Race, Ethnicity, and Geographic Location. Am. J. Mens. Health 2018, 12, 1807–1823. [Google Scholar] [CrossRef]
- Allott, E.H.; Hursting, S.D. Obesity and cancer: Mechanistic insights from transdisciplinary studies. Endocr. Relat. Cancer 2015, 22, R365–R386. [Google Scholar] [CrossRef]
- Booth, A.; Magnuson, A.; Fouts, J.; Foster, M. Adipose tissue, obesity and adipokines: Role in cancer promotion. Horm. Mol. Biol. Clin. Investig. 2015, 21, 57–74. [Google Scholar] [CrossRef] [PubMed]
- Garg, S.K.; Maurer, H.; Reed, K.; Selagamsetty, R. Diabetes and cancer: Two diseases with obesity as a common risk factor. Diabetes Obes. Metab. 2014, 16, 97–110. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.C.; Yang, W.V. Hyperglycemia, tumorigenesis, and chronic inflammation. Crit. Rev. Oncol. Hematol. 2016, 108, 146–153. [Google Scholar] [CrossRef]
- Marrone, M.T.; Selvin, E.; Barber, J.R.; Platz, E.A.; Joshu, C.E. Hyperglycemia, Classified with Multiple Biomarkers Simultaneously in Men without Diabetes, and Risk of Fatal Prostate Cancer. Cancer Prev. Res. 2019, 12, 103–112. [Google Scholar] [CrossRef]
- Liu, L.; Greenberg, S.; Russell, S.M.; Nicoll, C.S. Effects of insulin-like growth factors I and II on growth and differentiation of transplanted rat embryos and fetal tissues. Endocrinology 1989, 124, 3077–3082. [Google Scholar] [CrossRef]
- Baxter, R.C.; Holman, S.R.; Corbould, A.; Stranks, S.; Ho, P.J.; Braund, W. Regulation of the insulin-like growth factors and their binding proteins by glucocorticoid and growth hormone in nonislet cell tumor hypoglycemia. J. Clin. Endocrinol. Metab. 1995, 80, 2700–2708. [Google Scholar] [CrossRef]
- Resnicoff, M.; Abraham, D.; Yutanawiboonchai, W.; Rotman, H.L.; Kajstura, J.; Rubin, R.; Zoltick, P.; Baserga, R. The insulin-like growth factor I receptor protects tumor cells from apoptosis in vivo. Cancer Res. 1995, 55, 2463–2469. [Google Scholar] [PubMed]
- Buchanan, C.M.; Phillips, A.R.; Cooper, G.J. Preptin derived from proinsulin-like growth factor II (proIGF-II) is secreted from pancreatic islet beta-cells and enhances insulin secretion. Biochem. J. 2001, 360, 431–439. [Google Scholar] [CrossRef]
- Belobrajdic, D.P.; Frystyk, J.; Jeyaratnaganthan, N.; Espelund, U.; Flyvbjerg, A.; Clifton, P.M.; Noakes, M. Moderate energy restriction-induced weight loss affects circulating IGF levels independent of dietary composition. Eur. J. Endocrinol. 2010, 162, 1075–1082. [Google Scholar] [CrossRef] [PubMed]
- Brissenden, J.E.; Ullrich, A.; Francke, U. Human chromosomal mapping of genes for insulin-like growth factors I and II and epidermal growth factor. Nature 1984, 310, 781–784. [Google Scholar] [CrossRef]
- Rotwein, P. The insulin-like growth factor 2 gene and locus in nonmammalian vertebrates: Organizational simplicity with duplication but limited divergence in fish. J. Biol. Chem. 2018, 293, 15912–15932. [Google Scholar] [CrossRef]
- Ogawa, O.; Eccles, M.R.; Szeto, J.; McNoe, L.A.; Yun, K.; Maw, M.A.; Smith, P.J.; Reeve, A.E. Relaxation of insulin-like growth factor II gene imprinting implicated in Wilms’ tumour. Nature 1993, 362, 749–751. [Google Scholar] [CrossRef] [PubMed]
- Park, K.S.; Mitra, A.; Rahat, B.; Kim, K.; Pfeifer, K. Loss of imprinting mutations define both distinct and overlapping roles for misexpression of IGF2 and of H19 lncRNA. Nucleic Acids Res. 2017, 45, 12766–12779. [Google Scholar] [CrossRef][Green Version]
- Damaschke, N.A.; Yang, B.; Bhusari, S.; Avilla, M.; Zhong, W.; Blute, M.L., Jr.; Huang, W.; Jarrard, D.F. Loss of Igf2 Gene Imprinting in Murine Prostate Promotes Widespread Neoplastic Growth. Cancer Res. 2017, 77, 5236–5247. [Google Scholar] [CrossRef]
- Mishima, C.; Kagara, N.; Tanei, T.; Naoi, Y.; Shimoda, M.; Shimomura, A.; Shimazu, K.; Kim, S.J.; Noguchi, S. Loss of imprinting of IGF2 in fibroadenomas and phyllodes tumors of the breast. Oncol. Rep. 2016, 35, 1511–1518. [Google Scholar] [CrossRef] [PubMed]
- Belharazem, D.; Magdeburg, J.; Berton, A.K.; Beissbarth, L.; Sauer, C.; Sticht, C.; Marx, A.; Hofheinz, R.; Post, S.; Kienle, P.; et al. Carcinoma of the colon and rectum with deregulation of insulin-like growth factor 2 signaling: Clinical and molecular implications. J. Gastroenterol. 2016, 51, 971–984. [Google Scholar] [CrossRef]
- Zhang, M.; Wu, C.H.; Zhu, X.L.; Wang, Y.J. Loss of imprinting of insulin-like growth factor 2 is associated with increased risk of primary lung cancer in the central China region. Asian Pac. J. Cancer Prev. 2014, 15, 7799–7803. [Google Scholar] [CrossRef][Green Version]
- Bartolomei, M.S.; Webber, A.L.; Brunkow, M.E.; Tilghman, S.M. Epigenetic mechanisms underlying the imprinting of the mouse H19 gene. Genes Dev. 1993, 7, 1663–1673. [Google Scholar] [CrossRef]
- Reik, W.; Murrell, A. Genomic imprinting. Silence across the border. Nature 2000, 405, 408–409. [Google Scholar] [CrossRef]
- Yan, J.; Zhang, Y.; She, Q.; Li, X.; Peng, L.; Wang, X.; Liu, S.; Shen, X.; Zhang, W.; Dong, Y.; et al. Long Noncoding RNA H19/miR-675 Axis Promotes Gastric Cancer via FADD/Caspase 8/Caspase 3 Signaling Pathway. Cell. Physiol. Biochem. 2017, 42, 2364–2376. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Sheng, B.; Xia, Q.; Guan, X.; Zhang, Y. Association of long non-coding RNA H19 and microRNA-21 expression with the biological features and prognosis of non-small cell lung cancer. Cancer Gene Ther. 2017, 24, 317–324. [Google Scholar] [CrossRef]
- Yoshimizu, T.; Miroglio, A.; Ripoche, M.A.; Gabory, A.; Vernucci, M.; Riccio, A.; Colnot, S.; Godard, C.; Terris, B.; Jammes, H.; et al. The H19 locus acts in vivo as a tumor suppressor. Proc. Natl. Acad. Sci. USA 2008, 105, 12417–12422. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Chen, Q.; Liu, X.; Sun, Q.; Zhao, X.; Deng, R.; Wang, Y.; Huang, J.; Xu, M.; Yan, J.; et al. lncRNA H19/miR-675 axis represses prostate cancer metastasis by targeting TGFBI. FEBS J. 2014, 281, 3766–3775. [Google Scholar] [CrossRef] [PubMed]
- Bhusari, S.; Yang, B.; Kueck, J.; Huang, W.; Jarrard, D.F. Insulin-like growth factor-2 (IGF2) loss of imprinting marks a field defect within human prostates containing cancer. Prostate 2011, 71, 1621–1630. [Google Scholar] [CrossRef]
- Belharazem, D.; Kirchner, M.; Geissler, F.; Bugert, P.; Spahn, M.; Kneitz, B.; Riedmiller, H.; Sauer, C.; Kuffer, S.; Trojan, L.; et al. Relaxed imprinting of IGF2 in peripheral blood cells of patients with a history of prostate cancer. Endocr. Connect. 2012, 1, 87–94. [Google Scholar] [CrossRef]
- Kuffer, S.; Gutting, T.; Belharazem, D.; Sauer, C.; Michel, M.S.; Marx, A.; Trojan, L.; Strobel, P. Insulin-like growth factor 2 expression in prostate cancer is regulated by promoter-specific methylation. Mol. Oncol. 2018, 12, 256–266. [Google Scholar] [CrossRef] [PubMed]
- Hackshaw-McGeagh, L.; Lane, J.A.; Persad, R.; Gillatt, D.; Holly, J.M.; Koupparis, A.; Rowe, E.; Johnston, L.; Cloete, J.; Shiridzinomwa, C.; et al. Prostate cancer—Evidence of exercise and nutrition trial (PrEvENT): Study protocol for a randomised controlled feasibility trial. Trials 2016, 17, 123. [Google Scholar] [CrossRef]
- Bowers, L.W.; Rossi, E.L.; O’Flanagan, C.H.; de Graffenried, L.A.; Hursting, S.D. The Role of the Insulin/IGF System in Cancer: Lessons Learned from Clinical Trials and the Energy Balance-Cancer Link. Front. Endocrinol. 2015, 6, 77. [Google Scholar] [CrossRef]
- Shin, J.H.; Li, R.W.; Gao, Y.; Bickhart, D.M.; Liu, G.E.; Li, W.; Wu, S.; Li, C.J. Butyrate Induced IGF2 Activation Correlated with Distinct Chromatin Signatures Due to Histone Modification. Gene Regul. Syst. Biol. 2013, 7, 57–70. [Google Scholar] [CrossRef]
- Yang, B.; Wagner, J.; Damaschke, N.; Yao, T.; Wuerzberger-Davis, S.M.; Lee, M.H.; Svaren, J.; Miyamoto, S.; Jarrard, D.F. A novel pathway links oxidative stress to loss of insulin growth factor-2 (IGF2) imprinting through NF-kappaB activation. PLoS ONE 2014, 9, e88052. [Google Scholar] [CrossRef]
- Fu, V.X.; Dobosy, J.R.; Desotelle, J.A.; Almassi, N.; Ewald, J.A.; Srinivasan, R.; Berres, M.; Svaren, J.; Weindruch, R.; Jarrard, D.F. Aging and cancer-related loss of insulin-like growth factor 2 imprinting in the mouse and human prostate. Cancer Res. 2008, 68, 6797–6802. [Google Scholar] [CrossRef] [PubMed]
- Rancourt, R.C.; Harris, H.R.; Barault, L.; Michels, K.B. The prevalence of loss of imprinting of H19 and IGF2 at birth. FASEB J. 2013, 27, 3335–3343. [Google Scholar] [CrossRef] [PubMed]
- Verona, R.I.; Bartolomei, M.S. Role of H19 3’ sequences in controlling H19 and Igf2 imprinting and expression. Genomics 2004, 84, 59–68. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, J.; Christiansen, J.; Lykke-Andersen, J.; Johnsen, A.H.; Wewer, U.M.; Nielsen, F.C. A family of insulin-like growth factor II mRNA-binding proteins represses translation in late development. Mol. Cell. Biol. 1999, 19, 1262–1270. [Google Scholar] [CrossRef] [PubMed]
- Liao, B.; Hu, Y.; Herrick, D.J.; Brewer, G. The RNA-binding protein IMP-3 is a translational activator of insulin-like growth factor II leader-3 mRNA during proliferation of human K562 leukemia cells. J. Biol. Chem. 2005, 280, 18517–18524. [Google Scholar] [CrossRef] [PubMed]
- Bergman, D.; Halje, M.; Nordin, M.; Engstrom, W. Insulin-like growth factor 2 in development and disease: A mini-review. Gerontology 2013, 59, 240–249. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Rohan, T. Role of the insulin-like growth factor family in cancer development and progression. J. Natl. Cancer Inst. 2000, 92, 1472–1489. [Google Scholar] [CrossRef]
- Yamanaka, Y.; Wilson, E.M.; Rosenfeld, R.G.; Oh, Y. Inhibition of insulin receptor activation by insulin-like growth factor binding proteins. J. Biol. Chem. 1997, 272, 30729–30734. [Google Scholar] [CrossRef]
- Radhakrishnan, V.K.; Hernandez, L.C.; Anderson, K.; Tan, Q.; De Leon, M.; De Leon, D.D. Expression of Intratumoral IGF-II Is Regulated by the Gene Imprinting Status in Triple Negative Breast Cancer from Vietnamese Patients. Int. J. Endocrinol. 2015, 2015, 401851. [Google Scholar] [CrossRef]
- Zuo, Q.S.; Yan, R.; Feng, D.X.; Zhao, R.; Chen, C.; Jiang, Y.M.; Cruz-Correa, M.; Casson, A.G.; Kang, X.D.; Han, F.; et al. Loss of imprinting and abnormal expression of the insulin-like growth factor 2 gene in gastric cancer. Mol. Carcinog. 2011, 50, 390–396. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Damaschke, N.; Yao, T.; McCormick, J.; Wagner, J.; Jarrard, D. Pyrosequencing for accurate imprinted allele expression analysis. J. Cell. Biochem. 2015, 116, 1165–1170. [Google Scholar] [CrossRef] [PubMed]
- Davies, S.C.; Wass, J.A.; Ross, R.J.; Cotterill, A.M.; Buchanan, C.R.; Coulson, V.J.; Holly, J.M. The induction of a specific protease for insulin-like growth factor binding protein-3 in the circulation during severe illness. J. Endocrinol. 1991, 130, 469–473. [Google Scholar] [CrossRef] [PubMed]
- Dean, S.J.; Perks, C.M.; Holly, J.M.; Bhoo-Pathy, N.; Looi, L.M.; Mohammed, N.A.; Mun, K.S.; Teo, S.H.; Koobotse, M.O.; Yip, C.H.; et al. Loss of PTEN expression is associated with IGFBP2 expression, younger age, and late stage in triple-negative breast cancer. Am. J. Clin. Pathol. 2014, 141, 323–333. [Google Scholar] [CrossRef]
- NICE. Type 2 Diabetes: Prevention in People at High Risk. NICE Public Health Guideline 38. 2012. Available online: https://www.nice.org.uk/guidance/ph38/resources/type-2-diabetes-prevention-in-people-at-high-risk-pdf-1996304192197 (accessed on 21 September 2020).







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kingshott, G.; Biernacka, K.; Sewell, A.; Gwiti, P.; Barker, R.; Zielinska, H.; Gilkes, A.; McCarthy, K.; Martin, R.M.; Lane, J.A.; et al. Alteration of Metabolic Conditions Impacts the Regulation of IGF-II/H19 Imprinting Status in Prostate Cancer. Cancers 2021, 13, 825. https://doi.org/10.3390/cancers13040825
Kingshott G, Biernacka K, Sewell A, Gwiti P, Barker R, Zielinska H, Gilkes A, McCarthy K, Martin RM, Lane JA, et al. Alteration of Metabolic Conditions Impacts the Regulation of IGF-II/H19 Imprinting Status in Prostate Cancer. Cancers. 2021; 13(4):825. https://doi.org/10.3390/cancers13040825
Chicago/Turabian StyleKingshott, Georgina, Kalina Biernacka, Alex Sewell, Paida Gwiti, Rachel Barker, Hanna Zielinska, Amanda Gilkes, Kathryn McCarthy, Richard M. Martin, J. Athene Lane, and et al. 2021. "Alteration of Metabolic Conditions Impacts the Regulation of IGF-II/H19 Imprinting Status in Prostate Cancer" Cancers 13, no. 4: 825. https://doi.org/10.3390/cancers13040825
APA StyleKingshott, G., Biernacka, K., Sewell, A., Gwiti, P., Barker, R., Zielinska, H., Gilkes, A., McCarthy, K., Martin, R. M., Lane, J. A., McGeagh, L., Koupparis, A., Rowe, E., Oxley, J., Holly, J. M. P., & Perks, C. M. (2021). Alteration of Metabolic Conditions Impacts the Regulation of IGF-II/H19 Imprinting Status in Prostate Cancer. Cancers, 13(4), 825. https://doi.org/10.3390/cancers13040825