
Expression and Impact of C1GalT1 in Cancer Development and Progression



Abstract
:Simple Summary
Abstract
1. Introduction
2. C1galt1 Is Overexpressed in Many Epithelial Cancers and Is Associated with Poorer Prognosis and Poorer Patient Survival
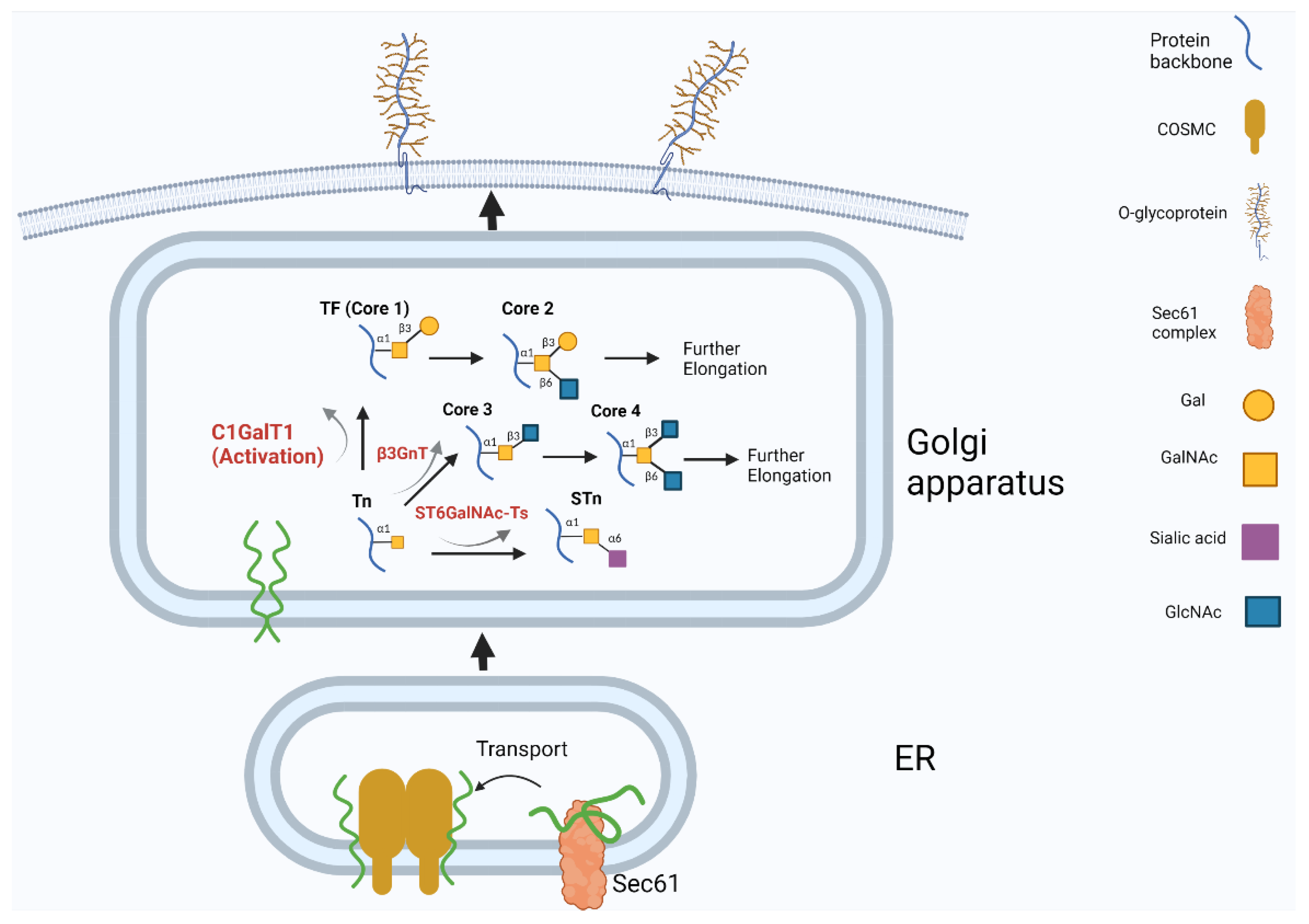
2.1. Possible Mechanisms of C1GalT1 Overexpression in Cancer
2.2. C1GalT Overexpression Increases the Occurrence of TF Antigen on Cancer Cell Surface
2.3. Change of C1GalT1 Expression Alters Glycosylation of Cell Membrane Mucin Proteins and Their Interaction with Partner Molecules
2.4. Change of C1GalT1 Expression Alters the Glycosylation and Function of Receptor Tyrosine Kinases
2.5. Change of C1GalT1 Expression Alters the Glycosylation and Function of Cell Surface Integrins
2.6. C1GalT1 Activity Regulates the Glycosylation and Function of Cell Surface Death Receptors
3. Conclusions Remarks
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| C1GalT1 | Core-1 β1:3galactosyltransferase |
| CBR | COSMC-binding region |
| ECM | extracellular matrix |
| EGFR | Epidermal growth factor receptor |
| ER | Endoplasmic reticulum |
| FGFR2 | Fibroblast growth factor receptor-2 |
| Gal | galactose |
| GalNAc | N-acetyl-galactosamine |
| GALNT | N-acetyl-galactosaminyltransferase |
| HER2 | human epidermal growth factor receptor-2 |
| HGFR | hepatocyte growth factor receptor |
| HNSCC | head and neck squamous cell carcinoma |
| PDAC | pancreatic ductal adenocarcinoma |
| RTK | receptor tyrosine kinase |
| STn | sialyl-GalNAcα-Ser/Thr |
| TF | Galβ1,3GalNAcα-ser/Thr (Thomsen–Friedenreich antigen) |
| Tn | GalNAcα-Ser/Thr |
| TRAIL | tumor necrosis factor-related apoptosis inducing ligand |
| VEGFR2 | Vascular endothelial growth factor-2 |
| VVA | Vicia Villosa lectin |
References
- Pinho, S.S.; Reis, C.A. Glycosylation in cancer: Mechanisms and clinical implications. Nat. Rev. Cancer 2015, 15, 540–555. [Google Scholar] [CrossRef] [PubMed]
- Reily, C.; Stewart, T.J.; Renfrow, M.B.; Novak, J. Glycosylation in health and disease. Nat. Rev. Nephrol. 2019, 15, 346–366. [Google Scholar] [CrossRef] [PubMed]
- Tran, D.T.; Ten Hagen, K.G. Mucin-type O-glycosylation during development. J. Biol. Chem. 2013, 288, 6921–6929. [Google Scholar] [CrossRef] [Green Version]
- Jensen, P.H.; Kolarich, D.; Packer, N.H. Mucin-type O-glycosylation—Putting the pieces together. FEBS J. 2010, 277, 81–94. [Google Scholar] [CrossRef]
- Haltiwanger, R.S.; Lowe, J.B. Role of glycosylation in development. Annu. Rev. Biochem. 2004, 73, 491–537. [Google Scholar] [CrossRef] [PubMed]
- Stowell, S.R.; Ju, T.; Cummings, R.D. Protein glycosylation in cancer. Annu. Rev. Pathol. 2015, 10, 473–510. [Google Scholar] [CrossRef] [Green Version]
- Cervoni, G.E.; Cheng, J.J.; Stackhouse, K.A.; Heimburg-Molinaro, J.; Cummings, R.D. O-glycan recognition and function in mice and human cancers. Biochem. J. 2020, 477, 1541–1564. [Google Scholar] [CrossRef] [PubMed]
- Ju, T.; Cummings, R.D. Core 1 β3Galactosyltransferase (C1GalT1, T-Synthase) and Its Specific Molecular Chaperone Cosmc (C1GalT1C1). In Handbook of Glycosyltransferases and Related Genes; Springer: Tokyo, Japan, 2014; pp. 149–169. [Google Scholar]
- Kudo, T.; Iwai, T.; Kubota, T.; Iwasaki, H.; Takayma, Y.; Hiruma, T.; Inaba, N.; Zhang, Y.; Gotoh, M.; Togayachi, A.; et al. Molecular cloning and characterization of a novel UDP-Gal:GalNAc(alpha) peptide beta 1,3-galactosyltransferase (C1Gal-T2), an enzyme synthesizing a core 1 structure of O-glycan. J. Biol. Chem. 2002, 277, 47724–47731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ju, T.; Brewer, K.; D’Souza, A.; Cummings, R.D.; Canfield, W.M. Cloning and expression of human core 1 beta1,3-galactosyltransferase. J. Biol. Chem. 2002, 277, 178–186. [Google Scholar] [CrossRef] [Green Version]
- Yu, L.G. The oncofetal Thomsen-Friedenreich carbohydrate antigen in cancer progression. Glycoconj. J. 2007, 24, 411–420. [Google Scholar] [CrossRef] [PubMed]
- Ju, T.; Cummings, R.D.; Canfield, W.M. Purification, characterization, and subunit structure of rat core 1 Beta1,3-galactosyltransferase. J. Biol. Chem. 2002, 277, 169–177. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Ju, T.; Ding, X.; Xia, B.; Wang, W.; Xia, L.; He, M.; Cummings, R.D. Cosmc is an essential chaperone for correct protein O-glycosylation. Proc. Natl. Acad. Sci. USA 2010, 107, 9228–9233. [Google Scholar] [CrossRef] [Green Version]
- Ju, T.; Lanneau, G.S.; Gautam, T.; Wang, Y.; Xia, B.; Stowell, S.R.; Willard, M.T.; Wang, W.; Xia, J.Y.; Zuna, R.E.; et al. Human tumor antigens Tn and sialyl Tn arise from mutations in Cosmc. Cancer Res. 2008, 68, 1636–1646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crew, V.K.; Singleton, B.K.; Green, C.; Parsons, S.F.; Daniels, G.; Anstee, D.J. New mutations in C1GALT1C1 in individuals with Tn positive phenotype. Br. J. Haematol. 2008, 142, 657–667. [Google Scholar] [CrossRef] [PubMed]
- Ju, T.; Wang, Y.; Aryal, R.P.; Lehoux, S.D.; Ding, X.; Kudelka, M.R.; Cutler, C.; Zeng, J.; Wang, J.; Sun, X.; et al. Tn and sialyl-Tn antigens, aberrant O-glycomics as human disease markers. Proteom. Clin. Appl. 2013, 7, 618–631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ju, T.; Aryal, R.P.; Stowell, C.J.; Cummings, R.D. Regulation of protein O-glycosylation by the endoplasmic reticulum-localized molecular chaperone Cosmc. J. Cell. Biol. 2008, 182, 531–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aryal, R.P.; Ju, T.; Cummings, R.D. The endoplasmic reticulum chaperone Cosmc directly promotes in vitro folding of T-synthase. J. Biol. Chem. 2010, 285, 2456–2462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aryal, R.P.; Ju, T.; Cummings, R.D. Tight complex formation between Cosmc chaperone and its specific client non-native T-synthase leads to enzyme activity and client-driven dissociation. J. Biol. Chem. 2012, 287, 15317–15329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aryal, R.P.; Ju, T.; Cummings, R.D. Identification of a novel protein binding motif within the T-synthase for the molecular chaperone Cosmc. J. Biol. Chem. 2014, 289, 11630–11641. [Google Scholar] [CrossRef] [Green Version]
- Vembar, S.S.; Brodsky, J.L. One step at a time: Endoplasmic reticulum-associated degradation. Nat. Rev. Mol. Cell. Biol. 2008, 9, 944–957. [Google Scholar] [CrossRef]
- Schoebel, S.; Mi, W.; Stein, A.; Ovchinnikov, S.; Pavlovicz, R.; DiMaio, F.; Baker, D.; Chambers, M.G.; Su, H.; Li, D.; et al. Cryo-EM structure of the protein-conducting ERAD channel Hrd1 in complex with Hrd3. Nature 2017, 548, 352–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, X.; Liu, Y.; Deng, X.; Shao, J.; Tian, S.; Chen, S.; Huang, R.; Lin, Z.; Chen, C.; Shen, L. C1GALT1, Negatively Regulated by miR-181d-5p, Promotes Tumor Progression via Upregulating RAC1 in Lung Adenocarcinoma. Front. Cell Dev. Biol. 2021, 9, 707970. [Google Scholar] [CrossRef] [PubMed]
- Hung, J.-S.; Huang, J.; Lin, Y.-C.; Huang, M.-J.; Lee, P.-H.; Lai, H.-S.; Liang, J.-T.; Huang, M.-C. C1GALT1 overexpression promotes the invasive behavior of colon cancer cells through modifying O-glycosylation of FGFR2. Oncotarget 2014, 5, 2096–2106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, J.; Che, M.I.; Lin, N.Y.; Hung, J.S.; Huang, Y.T.; Lin, W.C.; Huang, H.C.; Lee, P.H.; Liang, J.T.; Huang, M.C. The molecular chaperone Cosmc enhances malignant behaviors of colon cancer cells via activation of Akt and ERK. Mol. Carcinog. 2014, 53 (Suppl. S1), E62–E71. [Google Scholar] [CrossRef]
- Chou, C.-H.; Huang, M.-J.; Chen, C.-H.; Shyu, M.-K.; Huang, J.; Hung, J.-S.; Huang, C.-S.; Huang, M.-C. Up-regulation of C1GALT1 promotes breast cancer cell growth. Oncotarget 2015, 6, 6123–6135. [Google Scholar] [CrossRef] [Green Version]
- Du, T.; Jia, X.; Dong, X.; Ru, X.; Li, L.; Wang, Y.; Liu, J.; Feng, G.; Wen, T. Cosmc Disruption-Mediated Aberrant O-glycosylation Suppresses Breast Cancer Cell Growth via Impairment of CD44. Cancer Manag. Res. 2020, 12, 511–522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, P.C.; Chen, S.T.; Kuo, T.C.; Lin, T.C.; Lin, M.C.; Huang, J.; Hung, J.S.; Hsu, C.L.; Juan, H.F.; Lee, P.H.; et al. C1GALT1 is associated with poor survival and promotes soluble Ephrin A1-mediated cell migration through activation of EPHA2 in gastric cancer. Oncogene 2020, 39, 2724–2740. [Google Scholar] [CrossRef] [Green Version]
- Dong, X.; Chen, C.; Deng, X.; Liu, Y.; Duan, Q.; Peng, Z.; Luo, Z.; Shen, L. A novel mechanism for C1GALT1 in the regulation of gastric cancer progression. Cell Biosci. 2021, 11, 166. [Google Scholar] [CrossRef]
- Lin, M.C.; Chien, P.H.; Wu, H.Y.; Chen, S.T.; Juan, H.F.; Lou, P.J.; Huang, M.C. C1GALT1 predicts poor prognosis and is a potential therapeutic target in head and neck cancer. Oncogene 2018, 37, 5780–5793. [Google Scholar] [CrossRef]
- Kuo, T.C.; Wu, M.H.; Yang, S.H.; Chen, S.T.; Hsu, T.W.; Jhuang, J.Y.; Liao, Y.Y.; Tien, Y.W.; Huang, M.C. C1GALT1 high expression is associated with poor survival of patients with pancreatic ductal adenocarcinoma and promotes cell invasiveness through integrin alphav. Oncogene 2021, 40, 1242–1254. [Google Scholar] [CrossRef]
- Zhang, C.; Deng, X.; Qiu, L.; Peng, F.; Geng, S.; Shen, L.; Luo, Z. Knockdown of C1GalT1 inhibits radioresistance of human esophageal cancer cells through modifying beta1-integrin glycosylation. J. Cancer 2018, 9, 2666–2677. [Google Scholar] [CrossRef]
- Tzeng, S.F.; Tsai, C.H.; Chao, T.K.; Chou, Y.C.; Yang, Y.C.; Tsai, M.H.; Cha, T.L.; Hsiao, P.W. O-Glycosylation-mediated signaling circuit drives metastatic castration-resistant prostate cancer. FASEB J. 2018, 32, 6869–6882. [Google Scholar] [CrossRef] [PubMed]
- Chou, C.-H.; Huang, M.-J.; Liao, Y.-Y.; Chen, C.-H.; Huang, M.-C. C1GALT1 Seems to Promote In Vitro Disease Progression in Ovarian Cancer. Int. J. Gynecol. Cancer 2017, 27, 863–871. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.H.; Hu, R.H.; Huang, M.J.; Lai, I.R.; Chen, C.H.; Lai, H.S.; Wu, Y.M.; Huang, M.C. C1GALT1 promotes invasive phenotypes of hepatocellular carcinoma cells by modulating integrin beta1 glycosylation and activity. PLoS ONE 2014, 9, e94995. [Google Scholar] [CrossRef] [Green Version]
- Dong, X.; Luo, Z.; Wang, Y.; Meng, L.; Duan, Q.; Qiu, L.; Peng, F.; Shen, L. Altered O-glycosylation is associated with inherent radioresistance and malignancy of human laryngeal carcinoma. Exp. Cell Res. 2018, 362, 302–310. [Google Scholar] [CrossRef]
- Milde-Langosch, K.; Schutze, D.; Oliveira-Ferrer, L.; Wikman, H.; Muller, V.; Lebok, P.; Pantel, K.; Schroder, C.; Witzel, I.; Schumacher, U. Relevance of betaGal-betaGalNAc-containing glycans and the enzymes involved in their synthesis for invasion and survival in breast cancer patients. Breast Cancer Res. Treat. 2015, 151, 515–528. [Google Scholar] [CrossRef]
- Wu, Y.M.; Liu, C.H.; Huang, M.J.; Lai, H.S.; Lee, P.H.; Hu, R.H.; Huang, M.C. C1GALT1 enhances proliferation of hepatocellular carcinoma cells via modulating MET glycosylation and dimerization. Cancer Res. 2013, 73, 5580–5590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smyth, E.C.; Nilsson, M.; Grabsch, H.I.; van Grieken, N.C.T.; Lordick, F. Gastric cancer. Lancet 2020, 396, 635–648. [Google Scholar] [CrossRef]
- Chugh, S.; Barkeer, S.; Rachagani, S.; Nimmakayala, R.K.; Perumal, N.; Pothuraju, R.; Atri, P.; Mahapatra, S.; Thapa, I.; Talmon, G.A.; et al. Disruption of C1galt1 Gene Promotes Development and Metastasis of Pancreatic Adenocarcinomas in Mice. Gastroenterology 2018, 155, 1608–1624. [Google Scholar] [CrossRef]
- Hofmann, B.T.; Schluter, L.; Lange, P.; Mercanoglu, B.; Ewald, F.; Folster, A.; Picksak, A.S.; Harder, S.; El Gammal, A.T.; Grupp, K.; et al. COSMC knockdown mediated aberrant O-glycosylation promotes oncogenic properties in pancreatic cancer. Mol. Cancer 2015, 14, 109. [Google Scholar] [CrossRef] [Green Version]
- Luo, L.; Zhou, X.; Kolosov, V.P.; Perelman, J.M. The Cosmc-mediated effects of neutrophil elastase on T antigen expression in BEAS-2B cells. Respir. Physiol. Neurobiol. 2020, 281, 103496. [Google Scholar] [CrossRef]
- Vizcaino, C.; Mansilla, S.; Portugal, J. Sp1 transcription factor: A long-standing target in cancer chemotherapy. Pharmacol. Ther. 2015, 152, 111–124. [Google Scholar] [CrossRef] [Green Version]
- Chu, S.; Ferro, T.J. Sp1: Regulation of gene expression by phosphorylation. Gene 2005, 348, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Ren, Y.; Zhang, H.; Jiang, P. MicroRNA-382 inhibits cell growth and migration in colorectal cancer by targeting SP1. Biol. Res. 2018, 51, 51. [Google Scholar] [CrossRef] [PubMed]
- Malsy, M.; Graf, B.; Almstedt, K. The active role of the transcription factor Sp1 in NFATc2-mediated gene regulation in pancreatic cancer. BMC Biochem. 2019, 20, 2. [Google Scholar] [CrossRef] [PubMed]
- Beishline, K.; Azizkhan-Clifford, J. Sp1 and the ‘hallmarks of cancer’. FEBS J. 2015, 282, 224–258. [Google Scholar] [CrossRef] [PubMed]
- Barrow, H.; Tam, B.; Duckworth, C.A.; Rhodes, J.M.; Yu, L.G. Suppression of core 1 Gal-transferase is associated with reduction of TF and reciprocal increase of Tn, sialyl-Tn and Core 3 glycans in human colon cancer cells. PLoS ONE 2013, 8, e59792. [Google Scholar] [CrossRef] [Green Version]
- Itoh, K.; Akimoto, Y.; Kondo, S.; Ichimiya, T.; Aoki, K.; Tiemeyer, M.; Nishihara, S. Glucuronylated core 1 glycans are required for precise localization of neuromuscular junctions and normal formation of basement membranes on Drosophila muscles. Dev. Biol. 2018, 436, 108–124. [Google Scholar] [CrossRef]
- Xia, L.; McEver, R.P. Targeted Disruption of the Gene Encoding Core 1 β1-3-Galactosyltransferase (T-Synthase) Causes Embryonic Lethality and Defective Angiogenesis in Mice. Meth. Enzymol. 2006, 416, 314–331. [Google Scholar]
- Singh, R.; Campbell, B.J.; Yu, L.G.; Fernig, D.G.; Milton, J.D.; Goodlad, R.A.; FitzGerald, A.J.; Rhodes, J.M. Cell surface-expressed Thomsen-Friedenreich antigen in colon cancer is predominantly carried on high molecular weight splice variants of CD44. Glycobiology 2001, 11, 587–592. [Google Scholar] [CrossRef] [Green Version]
- Slovin, S.F.; Ragupathi, G.; Musselli, C.; Fernandez, C.; Diani, M.; Verbel, D.; Danishefsky, S.; Livingston, P.; Scher, H.I. Thomsen-Friedenreich (TF) antigen as a target for prostate cancer vaccine: Clinical trial results with TF cluster (c)-KLH plus QS21 conjugate vaccine in patients with biochemically relapsed prostate cancer. Cancer Immunol. Immunother. 2005, 54, 694–702. [Google Scholar] [CrossRef] [PubMed]
- Kölbl, A.C.; Jeschke, U.; Friese, K.; Andergassen, U. The role of TF- and Tn-antigens in breast cancer metastasis. Histol. Histopathol. 2016, 31, 613–621. [Google Scholar] [PubMed]
- Byrd, J.C.; Bresalier, R.S. Mucins and mucin binding proteins in colorectal cancer. Cancer Metastasis Rev. 2004, 23, 77–99. [Google Scholar] [CrossRef] [PubMed]
- Fu, C.; Zhao, H.; Wang, Y.; Cai, H.; Xiao, Y.; Zeng, Y.; Chen, H. Tumor-associated antigens: Tn antigen, sTn antigen, and T antigen. HLA 2016, 88, 275–286. [Google Scholar] [CrossRef]
- Cornelissen, L.A.M.; Blanas, A.; Zaal, A.; van der Horst, J.C.; Kruijssen, L.J.W.; O’Toole, T.; van Kooyk, Y.; van Vliet, S.J. Tn Antigen Expression Contributes to an Immune Suppressive Microenvironment and Drives Tumor Growth in Colorectal Cancer. Front. Oncol. 2020, 10, 1622. [Google Scholar] [CrossRef] [PubMed]
- Storr, S.J.; Royle, L.; Chapman, C.J.; Hamid, U.M.; Robertson, J.F.; Murray, A.; Dwek, R.A.; Rudd, P.M. The O-linked glycosylation of secretory/shed MUC1 from an advanced breast cancer patient’s serum. Glycobiology 2008, 18, 456–462. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Glinskii, O.V.; Mooney, B.P.; Rittenhouse-Olson, K.; Pienta, K.J.; Glinsky, V.V. Cell surface Thomsen-Friedenreich proteome profiling of metastatic prostate cancer cells reveals potential link with cancer stem cell-like phenotype. Oncotarget 2017, 8, 98598–98608. [Google Scholar] [CrossRef] [Green Version]
- Mulholland, D.J.; Xin, L.; Morim, A.; Lawson, D.; Witte, O.; Wu, H. Lin-Sca-1+CD49fhigh stem/progenitors are tumor-initiating cells in the Pten-null prostate cancer model. Cancer Res. 2009, 69, 8555–8562. [Google Scholar] [CrossRef] [Green Version]
- Collins, A.T.; Berry, P.A.; Hyde, C.; Stower, M.J.; Maitland, N.J. Prospective identification of tumorigenic prostate cancer stem cells. Cancer Res. 2005, 65, 10946–10951. [Google Scholar] [CrossRef] [Green Version]
- Cao, Y.; Merling, A.; Karsten, U.; Goletz, S.; Punzel, M.; Kraft, R.; Butschak, G.; Schwartz-Albiez, R. Expression of CD175 (Tn), CD175s (sialosyl-Tn) and CD176 (Thomsen-Friedenreich antigen) on malignant human hematopoietic cells. Int. J. Cancer 2008, 123, 89–99. [Google Scholar] [CrossRef]
- Yu, L.G.; Andrews, N.; Zhao, Q.; McKean, D.; Williams, J.F.; Connor, L.J.; Gerasimenko, O.V.; Hilkens, J.; Hirabayashi, J.; Kasai, K.; et al. Galectin-3 interaction with Thomsen-Friedenreich disaccharide on cancer-associated MUC1 causes increased cancer cell endothelial adhesion. J. Biol. Chem. 2007, 282, 773–781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khaldoyanidi, S.K.; Glinsky, V.V.; Sikora, L.; Glinskii, A.B.; Mossine, V.V.; Quinn, T.P.; Glinsky, G.V.; Sriramarao, P. MDA-MB-435 human breast carcinoma cell homo- and heterotypic adhesion under flow conditions is mediated in part by Thomsen-Friedenreich antigen-galectin-3 interactions. J. Biol. Chem. 2003, 278, 4127–4134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sindrewicz, P.; Lian, L.Y.; Yu, L.G. Interaction of the Oncofetal Thomsen-Friedenreich Antigen with Galectins in Cancer Progression and Metastasis. Front. Oncol. 2016, 6, 79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glinskii, O.V.; Sud, S.; Mossine, V.V.; Mawhinney, T.P.; Anthony, D.C.; Glinsky, G.V.; Pienta, K.J.; Glinsky, V.V. Inhibition of prostate cancer bone metastasis by synthetic TF antigen mimic/galectin-3 inhibitor lactulose-L-leucine. Neoplasia 2012, 14, 65–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Q.; Barclay, M.; Hilkens, J.; Guo, X.; Barrow, H.; Rhodes, J.M.; Yu, L.G. Interaction between circulating galectin-3 and cancer-associated MUC1 enhances tumour cell homotypic aggregation and prevents anoikis. Mol. Cancer 2010, 9, 154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Srinivasan, N.; Bane, S.M.; Ahire, S.D.; Ingle, A.D.; Kalraiya, R.D. Poly N-acetyllactosamine substitutions on N- and not O-oligosaccharides or Thomsen-Friedenreich antigen facilitate lung specific metastasis of melanoma cells via galectin-3. Glycoconj. J. 2009, 26, 445–456. [Google Scholar] [CrossRef]
- Radhakrishnan, P.; Dabelsteen, S.; Madsen, F.B.; Francavilla, C.; Kopp, K.L.; Steentoft, C.; Vakhrushev, S.Y.; Olsen, J.V.; Hansen, L.; Bennett, E.P.; et al. Immature truncated O-glycophenotype of cancer directly induces oncogenic features. Proc. Natl. Acad. Sci. USA 2014, 111, E4066–E4075. [Google Scholar] [CrossRef] [Green Version]
- Tarbell, J.M.; Cancel, L.M. The glycocalyx and its significance in human medicine. J. Intern. Med. 2016, 280, 97–113. [Google Scholar] [CrossRef] [Green Version]
- Taylor-Papadimitriou, J.; Burchell, J.; Miles, D.W.; Dalziel, M. MUC1 and cancer. Biochim. Biophys. Acta 1999, 1455, 301–313. [Google Scholar] [CrossRef] [Green Version]
- Guzman-Aranguez, A.; Argüeso, P. Structure and biological roles of mucin-type O-glycans at the ocular surface. Ocul. Surf. 2010, 8, 8–17. [Google Scholar] [CrossRef] [Green Version]
- Senapati, S.; Chaturvedi, P.; Chaney, W.G.; Chakraborty, S.; Gnanapragassam, V.S.; Sasson, A.R.; Batra, S.K. Novel INTeraction of MUC4 and galectin: Potential pathobiological implications for metastasis in lethal pancreatic cancer. Clin. Cancer Res. 2011, 17, 267–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piyush, T.; Chacko, A.R.; Sindrewicz, P.; Hilkens, J.; Rhodes, J.M.; Yu, L.-G. Interaction of galectin-3 with MUC1 on cell surface promotes EGFR dimerization and activation in human epithelial cancer cells. Cell Death Differ. 2017, 24, 1937–1947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, Z.; Lovly, C.M. Mechanisms of receptor tyrosine kinase activation in cancer. Mol. Cancer 2018, 17, 58. [Google Scholar] [CrossRef] [PubMed]
- Segaliny, A.I.; Tellez-Gabriel, M.; Heymann, M.F.; Heymann, D. Receptor tyrosine kinases: Characterisation, mechanism of action and therapeutic interests for bone cancers. J. Bone Oncol. 2015, 4, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Lemmon, M.A.; Schlessinger, J. Cell signaling by receptor tyrosine kinases. Cell 2010, 141, 1117–1134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robinson, D.R.; Wu, Y.M.; Lin, S.F. The protein tyrosine kinase family of the human genome. Oncogene 2000, 19, 5548–5557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blume-Jensen, P.; Hunter, T. Oncogenic kinase signalling. Nature 2001, 411, 355–365. [Google Scholar] [CrossRef]
- Itkonen, H.M.; Mills, I.G. Studying N-linked glycosylation of receptor tyrosine kinases. Methods Mol. Biol. 2015, 1233, 103–109. [Google Scholar] [CrossRef]
- Ferreira, I.G.; Pucci, M.; Venturi, G.; Malagolini, N.; Chiricolo, M.; Dall’Olio, F. Glycosylation as a Main Regulator of Growth and Death Factor Receptors Signaling. Int. J. Mol. Sci. 2018, 19, 580. [Google Scholar] [CrossRef] [Green Version]
- Contessa, J.N.; Bhojani, M.S.; Freeze, H.H.; Rehemtulla, A.; Lawrence, T.S. Inhibition of N-linked glycosylation disrupts receptor tyrosine kinase signaling in tumor cells. Cancer Res. 2008, 68, 3803–3809. [Google Scholar] [CrossRef] [Green Version]
- Mezu-Ndubuisi, O.J.; Maheshwari, A. The role of integrins in inflammation and angiogenesis. Pediatr. Res. 2021, 89, 1619–1626. [Google Scholar] [CrossRef]
- Marsico, G.; Russo, L.; Quondamatteo, F.; Pandit, A. Glycosylation and Integrin Regulation in Cancer. Trends Cancer 2018, 4, 537–552. [Google Scholar] [CrossRef] [PubMed]
- Desgrosellier, J.S.; Cheresh, D.A. Integrins in cancer: Biological implications and therapeutic opportunities. Nat. Rev. Cancer 2010, 10, 9–22. [Google Scholar] [CrossRef] [Green Version]
- Assoian, R.K.; Klein, E.A. Growth control by intracellular tension and extracellular stiffness. Trends Cell Biol. 2008, 18, 347–352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cooper, J.; Giancotti, F.G. Integrin Signaling in Cancer: Mechanotransduction, Stemness, Epithelial Plasticity, and Therapeutic Resistance. Cancer Cell 2019, 35, 347–367. [Google Scholar] [CrossRef]
- Jiang, Y.; Wen, T.; Yan, R.; Kim, S.R.; Stowell, S.R.; Wang, W.; Wang, Y.; An, G.; Cummings, R.D.; Ju, T. O-glycans on death receptors in cells modulate their sensitivity to TRAIL-induced apoptosis through affecting on their stability and oligomerization. FASEB J. 2020, 34, 11786–11801. [Google Scholar] [CrossRef]
- Xu, F.; Wang, D.; Cui, J.; Li, J.; Jiang, H. Demethylation of the Cosmc Promoter Alleviates the Progression of Breast Cancer Through Downregulation of the Tn and Sialyl-Tn Antigens. Cancer Manag. Res. 2020, 12, 1017–1027. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
| Cancer Type | C1GalT1 Expression | Effects on Cancer Cell Behaviors | Effects on Cell Signaling | Patient Survival | References |
|---|---|---|---|---|---|
| Colon | Increased | proliferation↑; migration↑; invasion↑; sphere formation↑ | EGF-ERK; FGFR2; PI3K-Akt | lower | [24,25] |
| Breast | Increased | viability↑; proliferation↑; migration↑; invasion↑ | CD44 inducing ERK-MAPK, p38/SAPKs, JNK; | lower | [26,27] |
| Pancreatic | Increased | viability↑; migration↑; invasion↑; | Integrin α5-FAk; Integrin αV-FAk Nucleoin, Grp-78, α-enolase; annexin A2; MUC16 inducing p-EGFR, α4 integrin and p-HER2 | lower | [31] |
| Decreased | migration↓; proliferation↓ | [40,41] | |||
| Hepatocellular | Increased | adhesion↑; migration↑; invasion↑ | HCF/MET; β1 integrin-FAK | lower | [35,38] |
| Gastric | Increased | viability↑; proliferation↑; migration↑; invasion↑ | ephrinA1-EphA2 Integrin α5-FAk; PI3K-AKt | lower | [28,29] |
| Head and neck | Increased | viability↑; migration↑; invasion↑ | EGF-EGFR | lower | [30] |
| Oesophageal | Increased | radiotherapy resistance↑ | β1 integrin-FAk | lower, and increased resistance to radiotherapy | [32] |
| Laryngeal | Increased | radiotherapy resistance↑ | β1 integrin-FAk | lower, and increased resistance to radiotherapy | [36] |
| Lung | Increased | proliferation↑; migration↑; colony formation↑ | Neutrophil Elastase (NE) via MUC5AC; PI3K, EGFR, Ras, p85; RAC1 | lower | [23,42] |
| Prostate | Increased | colony formation↑; sphere formation↑; proliferation↑ | Co-effect with galectin-4 to HER2 | lower | [33] |
| Ovarian | Increased | proliferation↑; migration↑; sphere formation↑ | CD133, CD24, Oct4, Nanog and SNAI2 | lower | [34] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wan, Y.; Yu, L.-G. Expression and Impact of C1GalT1 in Cancer Development and Progression. Cancers 2021, 13, 6305. https://doi.org/10.3390/cancers13246305
Wan Y, Yu L-G. Expression and Impact of C1GalT1 in Cancer Development and Progression. Cancers. 2021; 13(24):6305. https://doi.org/10.3390/cancers13246305
Chicago/Turabian StyleWan, Yangu, and Lu-Gang Yu. 2021. "Expression and Impact of C1GalT1 in Cancer Development and Progression" Cancers 13, no. 24: 6305. https://doi.org/10.3390/cancers13246305
APA StyleWan, Y., & Yu, L.-G. (2021). Expression and Impact of C1GalT1 in Cancer Development and Progression. Cancers, 13(24), 6305. https://doi.org/10.3390/cancers13246305