
Changes in the Proteome in the Development of Chronic Human Papillomavirus Infection—A Prospective Study in HIV Positive and HIV Negative Rwandan Women

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Description of the Cohort
2.2. Sample Preparation for Proteomic Analysis, LC-MS/MS Analysis and Data Analysis
2.3. Statistics, Venn Diagrams and Volcano Plots
2.4. Cluster Analysis
2.5. Protein-Protein Network Analysis
2.6. Ethical Statement
3. Results
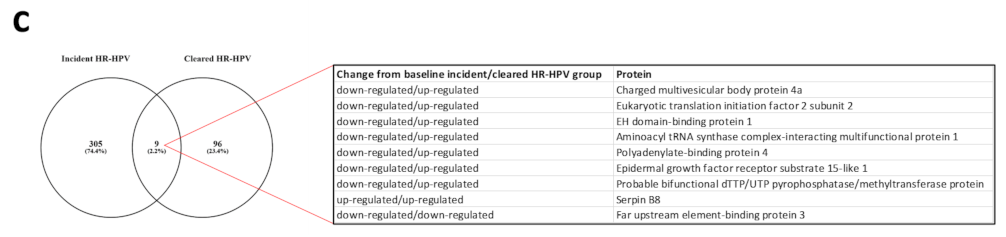
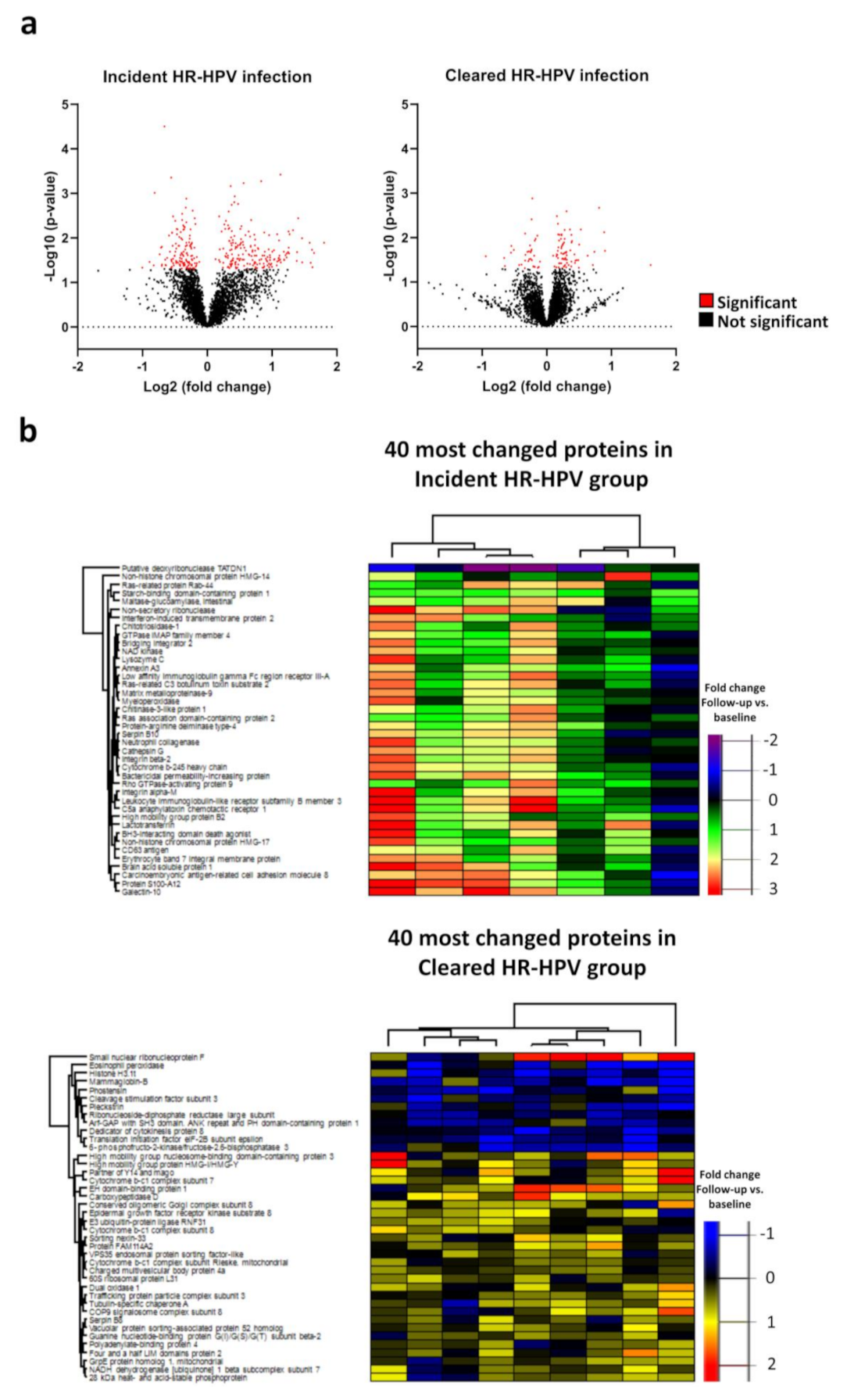
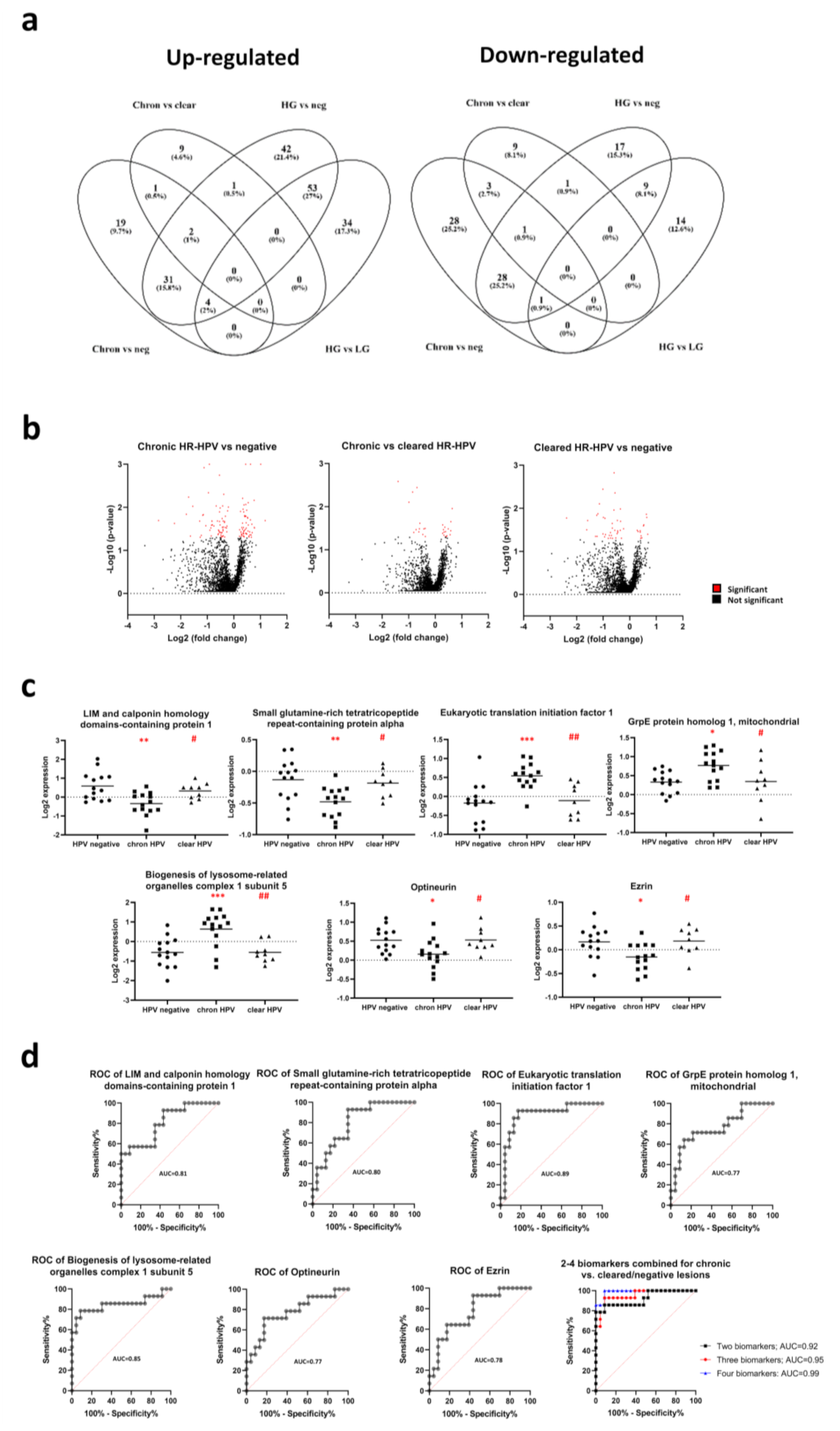
3.1. Proteome Changes in the Incident and Cleared HR-HPV Groups
3.2. Proteome Changes in the Chronic and Cleared HR-HPV Groups
3.3. Biomarker Panels Identifying Chronic HR-HPV Lesions
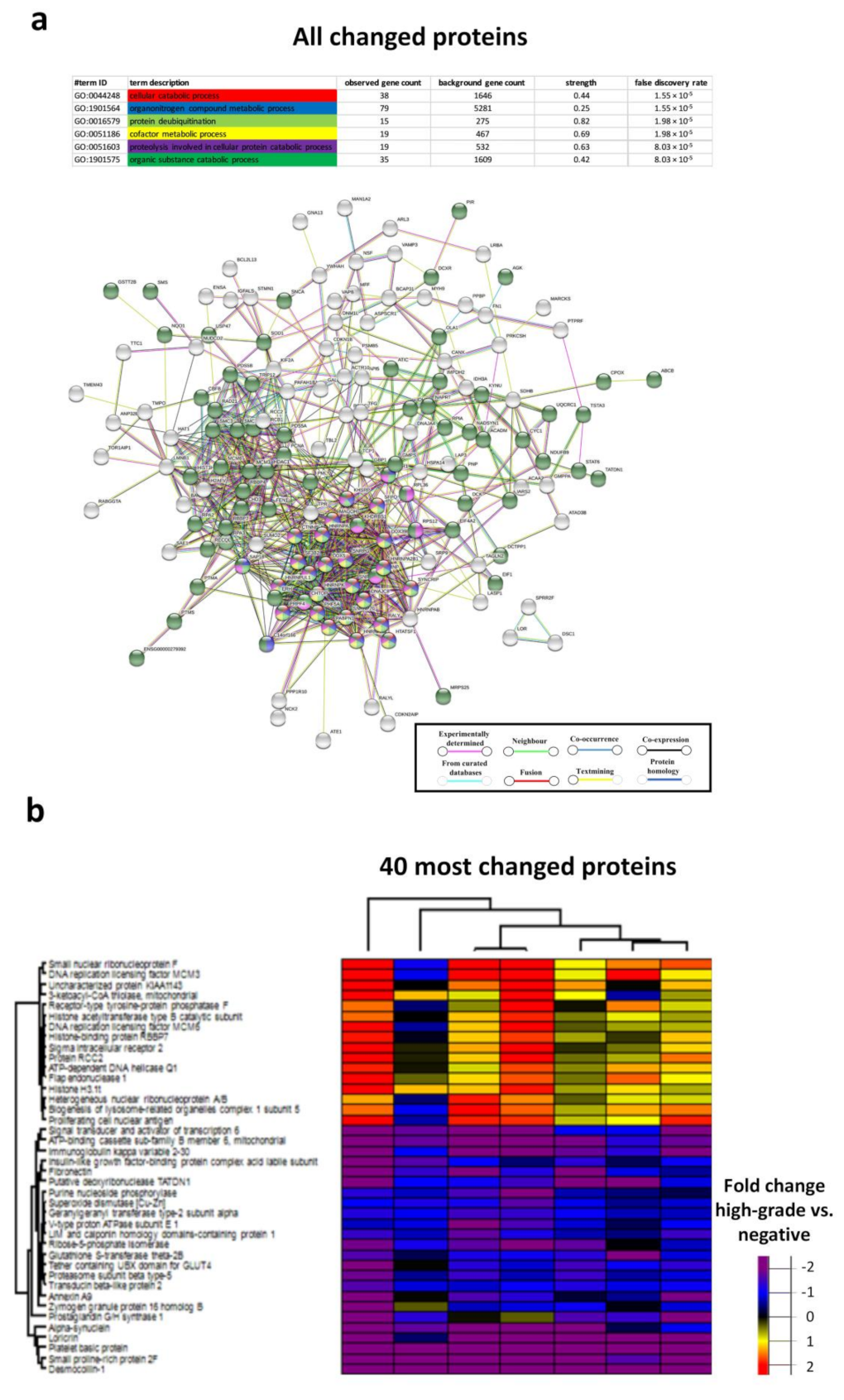
3.4. Biological Processes in the Chronic HR-HPV, Cleared HR-HPV and HPV Negative Groups
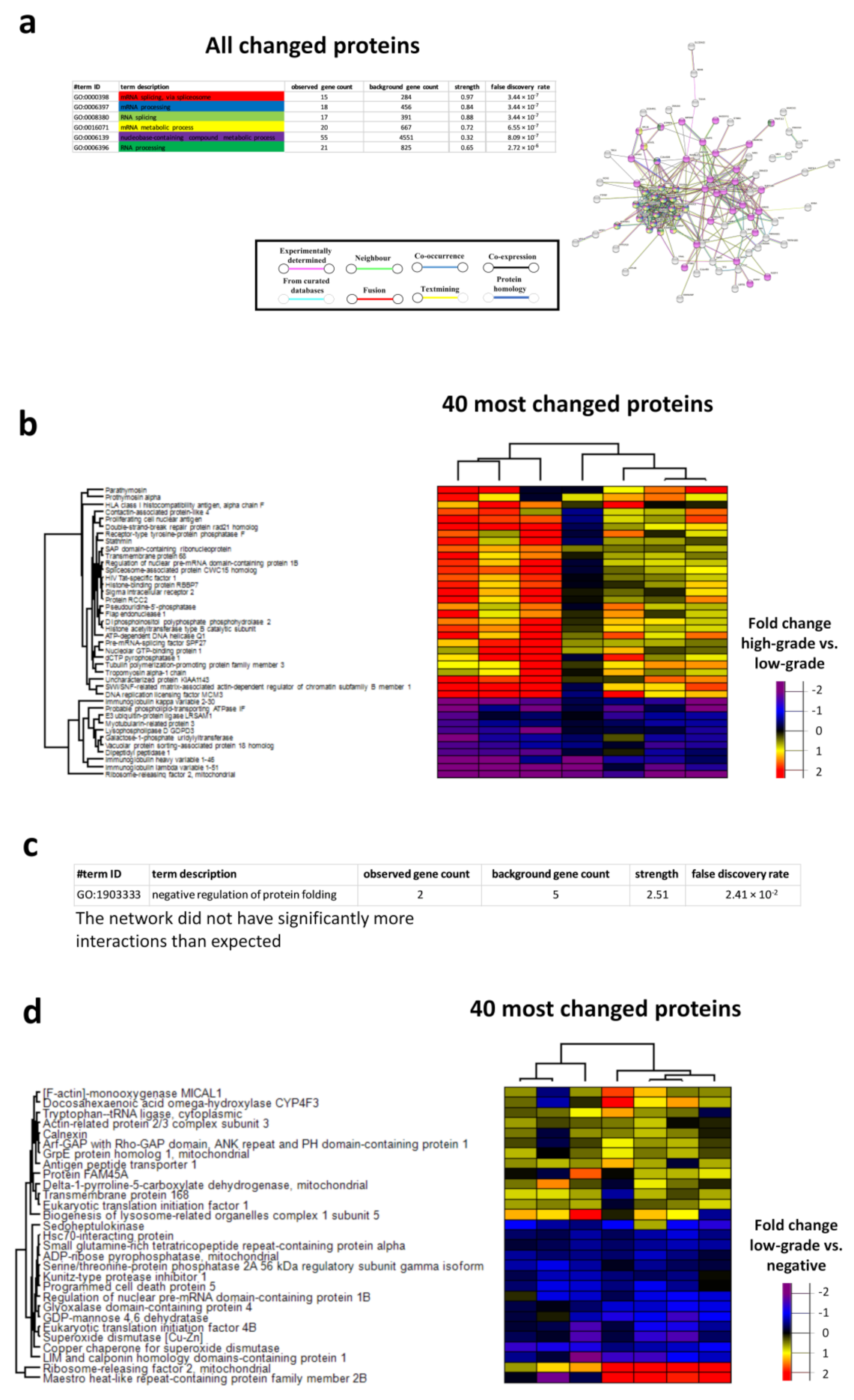
3.5. Proteome Changes in the Low-Grade and High-Grade Lesion Groups
3.6. Biomarker Panels Identifying High-Grade Lesions
3.7. Networks in High-Grade and Low-Grade Cervical Lesions
4. Discussion
4.1. The Immune System Is Involved in HPV Infection but Not in Clearance of Infection
4.2. Proteins Involved in the Development of HR-HPV Chronicity
4.3. Biomarkers Identifying Patients with Chronic HR-HPV Cervical Infections
4.4. RNA Processing in High-Grade Lesions
4.5. Biomarker Panels Identifying High-Grade Lesions
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [Green Version]
- Schiffman, M.H.; Bauer, H.M.; Hoover, R.N.; Glass, A.G.; Cadell, D.M.; Rush, B.B.; Scott, D.R.; Sherman, M.E.; Kurman, R.J.; Wacholder, S.; et al. Epidemiologic evidence showing that human papillomavirus infection causes most cervical intraepithelial neoplasia. J. Natl. Cancer Inst. 1993, 85, 958–964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukanyangezi, M.F.; Rugwizangoga, B.; Manzi, O.; Rulisa, S.; Hellstrand, K.; Tobin, G.; Martner, A.; Bienvenu, E.; Giglio, D. Persistence rate of cervical human papillomavirus infections and abnormal cytology in Rwanda. HIV Med. 2019, 20, 485–495. [Google Scholar] [CrossRef]
- Murenzi, G.; Tuyisenge, P.; Kanyabwisha, F.; Munyaneza, A.; Muhoza, B.; Kubwimana, G.; Murangwa, A.; Mutesa, L.; Anastos, K.; Castle, P.E. Type-specific persistence, clearance and incidence of high-risk HPV among screen-positive Rwandan women living with HIV. Infect. Agent Cancer 2021, 16, 16. [Google Scholar] [CrossRef]
- Muhimpundu, M.A.; Ngabo, F.; Sayinzoga, F.; Balinda, J.P.; Rusine, J.; Harward, S.; Eagan, A.; Krivacsy, S.; Bayingana, A.; Uwimbabazi, J.C.; et al. Screen, Notify, See, and Treat: Initial Results of Cervical Cancer Screening and Treatment in Rwanda. JCO Glob. Oncol. 2021, 7, 632–638. [Google Scholar] [CrossRef] [PubMed]
- Mukanyangezi, M.F.; Sengpiel, V.; Manzi, O.; Tobin, G.; Rulisa, S.; Bienvenu, E.; Giglio, D. Screening for human papillomavirus, cervical cytological abnormalities and associated risk factors in HIV-positive and HIV-negative women in Rwanda. HIV Med. 2018, 19, 152–166. [Google Scholar] [CrossRef] [Green Version]
- Serafin-Higuera, I.; Garibay-Cerdenares, O.L.; Illades-Aguiar, B.; Flores-Alfaro, E.; Jimenez-Lopez, M.A.; Sierra-Martinez, P.; Alarcon-Romero Ldel, C. Differential proteins among normal cervix cells and cervical cancer cells with HPV-16 infection, through mass spectrometry-based Proteomics (2D-DIGE) in women from Southern Mexico. Proteome Sci. 2016, 14, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiarini, A.; Liu, D.; Rassu, M.; Armato, U.; Eccher, C.; Dal Pra, I. Over Expressed TKTL1, CIP-2A, and B-MYB Proteins in Uterine Cervix Epithelium Scrapings as Potential Risk Predictive Biomarkers in HR-HPV-Infected LSIL/ASCUS Patients. Front. Oncol. 2019, 9, 213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Starodubtseva, N.L.; Brzhozovskiy, A.G.; Bugrova, A.E.; Kononikhin, A.S.; Indeykina, M.I.; Gusakov, K.I.; Chagovets, V.V.; Nazarova, N.M.; Frankevich, V.E.; Sukhikh, G.T.; et al. Label-free cervicovaginal fluid proteome profiling reflects the cervix neoplastic transformation. J. Mass Spectrom. 2019, 54, 693–703. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, A.L.; Lindberg, J.H.; Shevchenko, G.; Gustavsson, I.; Bergquist, J.; Gyllensten, U.; Enroth, S. Identification of Candidate Protein Biomarkers for CIN2+ Lesions from Self-Sampled, Dried Cervico-Vaginal Fluid Using LC-MS/MS. Cancers 2021, 13, 2592. [Google Scholar] [CrossRef]
- Tyanova, S.; Temu, T.; Sinitcyn, P.; Carlson, A.; Hein, M.Y.; Geiger, T.; Mann, M.; Cox, J. The Perseus computational platform for comprehensive analysis of (prote)omics data. Nat. Methods 2016, 13, 731–740. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [Green Version]
- Al-Eitan, L.N.; Tarkhan, A.H.; Alghamdi, M.A.; Al-Qarqaz, F.A.; Al-Kofahi, H.S. Transcriptome analysis of HPV-induced warts and healthy skin in humans. BMC Med. Genom. 2020, 13, 35. [Google Scholar] [CrossRef] [Green Version]
- Xie, Y.; Feng, Y.; Li, W.; Zhan, F.; Huang, G.; Hu, H.; Xiong, Y.; Tan, B.; Chen, T. Revealing the Disturbed Vaginal Micobiota Caused by Cervical Cancer Using High-Throughput Sequencing Technology. Front. Cell Infect. Microbiol. 2020, 10, 538336. [Google Scholar] [CrossRef]
- Alvarez, K.L.F.; Beldi, M.; Sarmanho, F.; Rossetti, R.A.M.; Silveira, C.R.F.; Mota, G.R.; Andreoli, M.A.; Caruso, E.D.C.; Kamillos, M.F.; Souza, A.M.; et al. Local and systemic immunomodulatory mechanisms triggered by Human Papillomavirus transformed cells: A potential role for G-CSF and neutrophils. Sci. Rep. 2017, 7, 9002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DiGiuseppe, S.; Bienkowska-Haba, M.; Guion, L.G.; Sapp, M. Cruising the cellular highways: How human papillomavirus travels from the surface to the nucleus. Virus Res 2017, 231, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Kazemi, S.; Papadopoulou, S.; Li, S.; Su, Q.; Wang, S.; Yoshimura, A.; Matlashewski, G.; Dever, T.E.; Koromilas, A.E. Control of alpha subunit of eukaryotic translation initiation factor 2 (eIF2 alpha) phosphorylation by the human papillomavirus type 18 E6 oncoprotein: Implications for eIF2 alpha-dependent gene expression and cell death. Mol. Cell. Biol. 2004, 24, 3415–3429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hao, P.; Yu, J.; Ward, R.; Liu, Y.; Hao, Q.; An, S.; Xu, T. Eukaryotic translation initiation factors as promising targets in cancer therapy. Cell Commun. Signal. 2020, 18, 175. [Google Scholar] [CrossRef]
- Liu, Z.; Teng, L.; Gao, L.; Wang, H.; Su, Y.; Li, J. The role of eukaryotic translation initiation factor 5A-1 (eIF5A-1) gene in HPV 16 E6 induces cell growth in human cervical squamous carcinoma cells. Biochem. Biophys. Res. Commun. 2018, 504, 6–12. [Google Scholar] [CrossRef]
- Meznad, K.; Paget-Bailly, P.; Jacquin, E.; Peigney, A.; Aubin, F.; Guittaut, M.; Mougin, C.; Pretet, J.L.; Baguet, A. The exon junction complex core factor eIF4A3 is a key regulator of HPV16 gene expression. Biosci. Rep. 2021, 41. [Google Scholar] [CrossRef]
- Zhang, W.; Su, X.; Li, S.; Wang, Y.; Wang, Q.; Zeng, H. Inhibiting MNK Selectively Targets Cervical Cancer via Suppressing eIF4E-Mediated beta-Catenin Activation. Am. J. Med. Sci. 2019, 358, 227–234. [Google Scholar] [CrossRef]
- Jaiswal, P.K.; Koul, S.; Palanisamy, N.; Koul, H.K. Eukaryotic Translation Initiation Factor 4 Gamma 1 (EIF4G1): A target for cancer therapeutic intervention? Cancer Cell Int. 2019, 19, 224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, P.; Tan, Q.; Jiang, W.; Ou, Y.; Xu, P.; Yuan, L. Eukaryotic translation initiation factor 3B is overexpressed and correlates with deteriorated tumor features and unfavorable survival profiles in cervical cancer patients. Cancer Biomark. 2019, 26, 123–130. [Google Scholar] [CrossRef]
- Nakajo, A.; Yoshimura, S.; Togawa, H.; Kunii, M.; Iwano, T.; Izumi, A.; Noguchi, Y.; Watanabe, A.; Goto, A.; Sato, T.; et al. EHBP1L1 coordinates Rab8 and Bin1 to regulate apical-directed transport in polarized epithelial cells. J. Cell Biol. 2016, 212, 297–306. [Google Scholar] [CrossRef]
- Kobayashi, S.; Suzuki, T.; Kawaguchi, A.; Phongphaew, W.; Yoshii, K.; Iwano, T.; Harada, A.; Kariwa, H.; Orba, Y.; Sawa, H. Rab8b Regulates Transport of West Nile Virus Particles from Recycling Endosomes. J. Biol. Chem. 2016, 291, 6559–6568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krishnan, M.N.; Sukumaran, B.; Pal, U.; Agaisse, H.; Murray, J.L.; Hodge, T.W.; Fikrig, E. Rab 5 is required for the cellular entry of dengue and West Nile viruses. J. Virol. 2007, 81, 4881–4885. [Google Scholar] [CrossRef] [Green Version]
- Young, J.M.; Zine El Abidine, A.; Gomez-Martinez, R.A.; Ozbun, M.A. The Known and Potential Intersections of Rab-GTPases in Human Papillomavirus Infections. Front. Cell Dev. Biol. 2019, 7, 139. [Google Scholar] [CrossRef] [Green Version]
- Liang, D.; Tian, L.; You, R.; Halpert, M.M.; Konduri, V.; Baig, Y.C.; Paust, S.; Kim, D.; Kim, S.; Jia, F.; et al. AIMp1 Potentiates TH1 Polarization and Is Critical for Effective Antitumor and Antiviral Immunity. Front. Immunol. 2017, 8, 1801. [Google Scholar] [CrossRef] [PubMed]
- Halpert, M.M.; Konduri, V.; Liang, D.; Vazquez-Perez, J.; Hofferek, C.J.; Weldon, S.A.; Baig, Y.; Vedula, I.; Levitt, J.M.; Decker, W.K. MHC class I and II peptide homology regulates the cellular immune response. FASEB J. 2020, 34, 8082–8101. [Google Scholar] [CrossRef] [Green Version]
- Halle, M.K.; Sodal, M.; Forsse, D.; Engerud, H.; Woie, K.; Lura, N.G.; Wagner-Larsen, K.S.; Trovik, J.; Bertelsen, B.I.; Haldorsen, I.S.; et al. A 10-gene prognostic signature points to LIMCH1 and HLA-DQB1 as important players in aggressive cervical cancer disease. Br. J. Cancer 2021, 124, 1690–1698. [Google Scholar] [CrossRef] [PubMed]
- Xi, M.; Tang, W. Knockdown of Ezrin inhibited migration and invasion of cervical cancer cells in vitro. Int. J. Immunopathol. Pharmacol. 2020, 34, 2058738420930899. [Google Scholar] [CrossRef]
- Kong, J.; Li, Y.; Liu, S.; Jin, H.; Shang, Y.; Quan, C.; Li, Y.; Lin, Z. High expression of ezrin predicts poor prognosis in uterine cervical cancer. BMC Cancer 2013, 13, 520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zacapala-Gomez, A.E.; Navarro-Tito, N.; Alarcon-Romero, L.D.C.; Ortuno-Pineda, C.; Illades-Aguiar, B.; Castaneda-Saucedo, E.; Ortiz-Ortiz, J.; Garibay-Cerdenares, O.L.; Jimenez-Lopez, M.A.; Mendoza-Catalan, M.A. Ezrin and E-cadherin expression profile in cervical cytology: A prognostic marker for tumor progression in cervical cancer. BMC Cancer 2018, 18, 349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inokuchi, S.; Yoshizumi, T.; Toshima, T.; Itoh, S.; Yugawa, K.; Harada, N.; Mori, H.; Fukuhara, T.; Matsuura, Y.; Mori, M. Suppression of optineurin impairs the progression of hepatocellular carcinoma through regulating mitophagy. Cancer Med. 2021, 10, 1501–1514. [Google Scholar] [CrossRef] [PubMed]
- Kamiyama, H.; Izumida, M.; Umemura, Y.; Hayashi, H.; Matsuyama, T.; Kubo, Y. Role of Ezrin Phosphorylation in HIV-1 Replication. Front. Microbiol. 2018, 9, 1912. [Google Scholar] [CrossRef] [PubMed]
- Dutta, S.; Tan, Y.J. Structural and functional characterization of human SGT and its interaction with Vpu of the human immunodeficiency virus type 1. Biochemistry 2008, 47, 10123–10131. [Google Scholar] [CrossRef] [PubMed]
- Graham, S.V.; Faizo, A.A.A. Control of human papillomavirus gene expression by alternative splicing. Virus Res. 2017, 231, 83–95. [Google Scholar] [CrossRef] [Green Version]
- Pottier, C.; Kriegsmann, M.; Alberts, D.; Smargiasso, N.; Baiwir, D.; Mazzucchelli, G.; Herfs, M.; Fresnais, M.; Casadonte, R.; Delvenne, P.; et al. Microproteomic Profiling of High-Grade Squamous Intraepithelial Lesion of the Cervix: Insight into Biological Mechanisms of Dysplasia and New Potential Diagnostic Markers. Proteomics Clin. Appl. 2019, 13, e1800052. [Google Scholar] [CrossRef] [Green Version]
- Chand, K.; Iyer, K.; Mitra, D. Comparative analysis of differential gene expression of HSP40 and HSP70 family isoforms during heat stress and HIV-1 infection in T-cells. Cell Stress Chaperones 2021, 26, 403–416. [Google Scholar] [CrossRef] [PubMed]
- Lefranc, M.P. Immunoglobulin and T Cell Receptor Genes: IMGT((R)) and the Birth and Rise of Immunoinformatics. Front. Immunol. 2014, 5, 22. [Google Scholar] [CrossRef] [Green Version]
- Pang, K.; Zhang, Z.; Hao, L.; Shi, Z.; Chen, B.; Zang, G.; Dong, Y.; Li, R.; Liu, Y.; Wang, J.; et al. The ERH gene regulates migration and invasion in 5637 and T24 bladder cancer cells. BMC Cancer 2019, 19, 225. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Chu, Y.J.; Song, K.J.; Chen, Y.L.; Liu, W.; Lv, T.; Wang, J.; Zhao, H.; Ren, Y.Z.; Xu, J.X.; et al. Knockdown of enhancer of rudimentary homolog inhibits proliferation and metastasis in ovarian cancer by regulating epithelial-mesenchymal transition. Biomed. Pharmacother. 2020, 125, 109974. [Google Scholar] [CrossRef]
- Xiao, M.; Tang, N.; Yan, Y.; Li, Z.; Shi, S.; He, S.; Chen, Z.; Cao, K.; Chen, J.; Zhou, J.; et al. Knockdown of enhancer of rudimentary homolog expression attenuates proliferation, cell cycle and apoptosis of melanoma cells. Melanoma Res. 2021, 31, 309–318. [Google Scholar] [CrossRef] [PubMed]
- Zeng, C.; Riad, A.; Mach, R.H. The Biological Function of Sigma-2 Receptor/TMEM97 and Its Utility in PET Imaging Studies in Cancer. Cancers 2020, 12, 1877. [Google Scholar] [CrossRef]
- Brito, M.J.; Sequeira, P.; Silva, I.; Quintas, A.; Martins, C.; Felix, A. CD4(+) and CD8(+) cell populations in HIV-positive women with cervical squamous intra-epithelial lesions and squamous cell carcinoma. Int. J. Infect. Dis. 2021, 103, 370–377. [Google Scholar] [CrossRef]
- Lucena, A.A.; Guimaraes, M.V.; Michelin, M.A.; Lodi, C.T.; Lima, M.I.; Murta, E.F.; Melo, V.H. Evaluation of T, B and natural killer lymphocyte in the cervical stroma of HIV-positive and negative patients with cervical intraepithelial neoplasia. Immunol. Lett. 2016, 169, 98–103. [Google Scholar] [CrossRef]
- Perez-Riverol, Y.; Csordas, A.; Bai, J.; Bernal-Llinares, M.; Hewapathirana, S.; Kundu, D.J.; Inuganti, A.; Griss, J.; Mayer, G.; Eisenacher, M.; et al. The PRIDE database and related tools and resources in 2019: Improving support for quantification data. Nucleic Acids Res. 2019, 47, D442–D450. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group A. HPV Incident | |||||
|---|---|---|---|---|---|
| Age at Inclusion | HIV Status | Cytology Baseline | Cytology Follow-Up | HPV Types Follow-Up | |
| Woman 1 | 45 | Positive | Normal | Normal | 66, 87 |
| Woman 2 | 61 | Positive | Normal | Not determined | 52 |
| Woman 3 | 40 | Positive | Normal | Normal | 45 |
| Woman 4 | 60 | Positive | Normal | Normal | 68 |
| Woman 5 | 48 | Positive | ASCUS | Normal | 45 (6) |
| Woman 6 | 47 | Positive | Normal | Normal | 31 |
| Woman 7 | 31 | Positive | Normal | Normal | 16 (74) |
| Group B. HPV Cleared | |||||
| Age at Inclusion | HIV Status | Cytology Baseline | Cytology Follow-Up | HPV Types Baseline | |
| Woman 8 | 49 | Positive | Normal | Normal | 53, 91 (16) |
| Woman 9 | 55 | Positive | Normal | Normal | 58 |
| Woman 10 | 45 | Positive | Normal | Normal | 16 |
| Woman 11 | 44 | Positive | Normal | ASCUS | 56 |
| Woman 12 | 50 | Positive | Normal | Normal | 69 |
| Woman 13 | 45 | Positive | HSIL | ASCUS | 16 |
| Woman 14 | 43 | Negative | Normal | Normal | 52 |
| Woman 15 | 38 | Negative | Normal | Normal | 16 |
| Woman 16 | 26 | Negative | LSIL | Normal | 51, 87 |
| Group C. HPV Chronic | |||||
| Age at Inclusion | HIV Status | HPV Types | Cytology | Subgroup | |
| Woman 17 | 51 | Positive | 16 | Normal | Low-grade |
| Woman 18 | 38 | Positive | 68, 70, 83, 86 (6, 31) | Normal | Low-grade |
| Woman 19 | 42 | Positive | 52, 53 | ASCUS | Low-grade |
| Woman 20 | 48 | Positive | 52 (67, 90) | ASCUS | Low-grade |
| Woman 21 | 43 | Positive | 39, 52, 53 | ASCUS | Low-grade |
| Woman 22 | 37 | Positive | 68 (87) | LSIL | Low-grade |
| Woman 23 | 36 | Positive | 70 (35) | LSIL | Low-grade |
| Woman 24 | 39 | Positive | 58 | HSIL | High-grade |
| Woman 25 | 52 | Positive | 52, 58 | HSIL | High-grade |
| Woman 26 | 53 | Positive | 31 | HSIL | High-grade |
| Woman 27 | 47 | Positive | 31 | HSIL | High-grade |
| Woman 28 | 43 | Positive | 16, 51, 52 | HSIL | High-grade |
| Woman 29 | 43 | Positive | 33 (59) | AIS | High-grade |
| Woman 30 | 50 | Positive | 45, 70 (6, 11, 70, 89, 51) | SCC | High-grade |
| Group D. Normal Cytology | |||||
| Age at Inclusion | HIV Status | HPV Types | Cytology | ||
| Woman 31 | 41 | Positive | Negative | Normal | |
| Woman 32 | 61 | Positive | Negative | Normal | |
| Woman 33 | 56 | Positive | Negative | Normal | |
| Woman 34 | 50 | Positive | Negative | Normal | |
| Woman 35 | 45 | Positive | Negative | Normal | |
| Woman 36 | 51 | Positive | Negative | Normal | |
| Woman 37 | 39 | Positive | Negative | Normal | |
| Woman 38 | 40 | Positive | Negative | Normal | |
| Woman 39 | 40 | Negative | Negative | Normal | |
| Woman 40 | 42 | Negative | Negative | Normal | |
| Woman 41 | 55 | Negative | Negative | Normal | |
| Woman 42 | 41 | Negative | Negative | Normal | |
| Woman 43 | 33 | Unknown | Negative | Normal | |
| Woman 44 | 40 | Negative | Negative | Normal |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bienvenu, E.; Mukanyangezi, M.F.; Rulisa, S.; Martner, A.; Hasséus, B.; Vorontsov, E.; Tobin, G.; Giglio, D. Changes in the Proteome in the Development of Chronic Human Papillomavirus Infection—A Prospective Study in HIV Positive and HIV Negative Rwandan Women. Cancers 2021, 13, 5983. https://doi.org/10.3390/cancers13235983
Bienvenu E, Mukanyangezi MF, Rulisa S, Martner A, Hasséus B, Vorontsov E, Tobin G, Giglio D. Changes in the Proteome in the Development of Chronic Human Papillomavirus Infection—A Prospective Study in HIV Positive and HIV Negative Rwandan Women. Cancers. 2021; 13(23):5983. https://doi.org/10.3390/cancers13235983
Chicago/Turabian StyleBienvenu, Emile, Marie Francoise Mukanyangezi, Stephen Rulisa, Anna Martner, Bengt Hasséus, Egor Vorontsov, Gunnar Tobin, and Daniel Giglio. 2021. "Changes in the Proteome in the Development of Chronic Human Papillomavirus Infection—A Prospective Study in HIV Positive and HIV Negative Rwandan Women" Cancers 13, no. 23: 5983. https://doi.org/10.3390/cancers13235983
APA StyleBienvenu, E., Mukanyangezi, M. F., Rulisa, S., Martner, A., Hasséus, B., Vorontsov, E., Tobin, G., & Giglio, D. (2021). Changes in the Proteome in the Development of Chronic Human Papillomavirus Infection—A Prospective Study in HIV Positive and HIV Negative Rwandan Women. Cancers, 13(23), 5983. https://doi.org/10.3390/cancers13235983