Mechanical Loading-Driven Tumor Suppression Is Mediated by Lrp5-Dependent and Independent Mechanisms

 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Simple Summary
Abstract
1. Introduction
2. Results
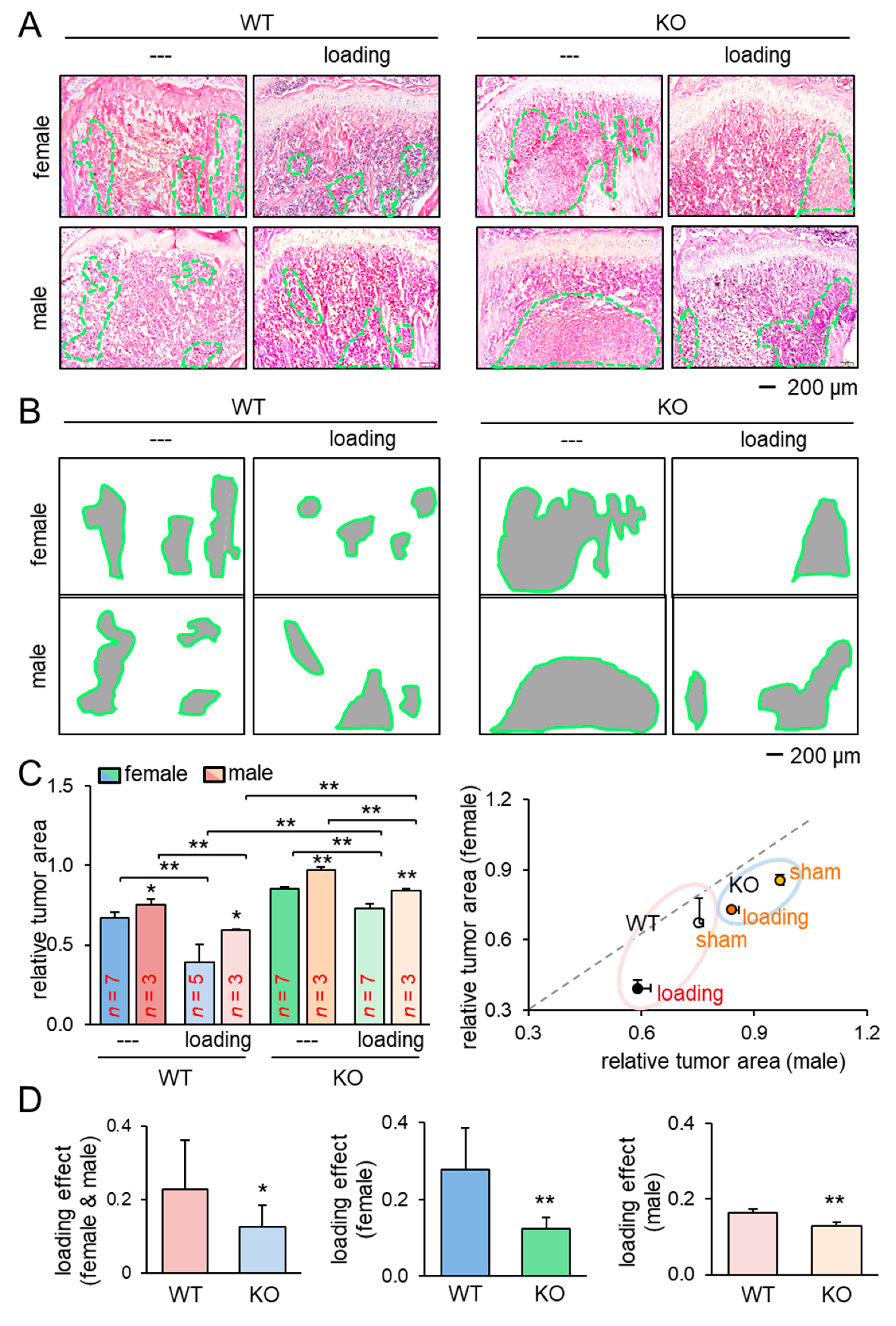
2.1. Inhibitory Effects of Knee Loading on Mammary Tumors
2.2. Protection of Tumor-Invaded Tibia by Knee Loading
2.3. Loading-Dependent Alterations in Dopamine and Cholesterol in the Serum and Urine
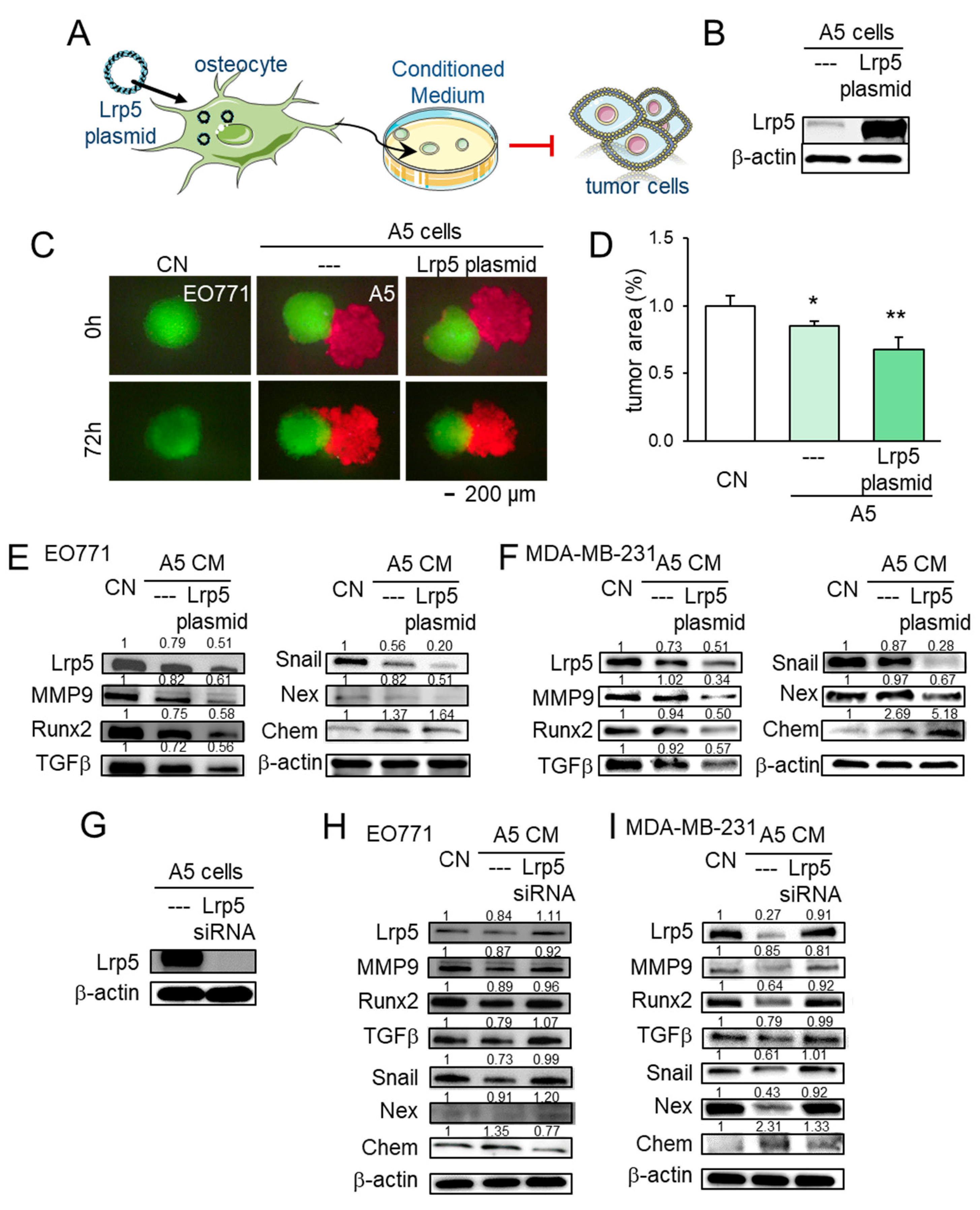
2.4. Regulation of Chemerin as a Tumor Suppressor and Nexin as a Tumor Promoter
2.5. Tumor-Suppressing Effect of TRAIL, p53, Chemerin, and Tumor-Promoting Effect of Nexin
2.6. Lrp5 as a Tumor-Suppressor in Osteocytes
2.7. Regulation of Osteoclastogenesis by Chemerin and Nexin
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. MTT, EdU, Invasion, Wound-Healing Scratch, and 3D Spheroid Competition Assay
4.3. Western Blotting, RNA Interference, Plasmid Transfection
4.4. ELISA and Protein Array Analysis
4.5. Osteoclast Differentiation Assay
4.6. Animal Models
4.7. Knee Loading
4.8. μCT Imaging and Histology
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schneider, A.P., 2nd; Zainer, C.M.; Kubat, C.K.; Mullen, N.K.; Windisch, A.K. The breast cancer epidemic: 10 facts. Linacre Q. 2014, 81, 244–277. [Google Scholar] [CrossRef] [PubMed]
- Castillejos-Molina, R.A.; Gabilondo-Navarro, F.B. Prostate cancer. Salud Publica Mexico 2016, 58, 279–284. [Google Scholar] [CrossRef] [PubMed]
- Haider, M.T.; Smit, D.J.; Taipaleenmaki, H. The endosteal niche in breast cancer bone metastasis. Front. Oncol. 2020, 10, 335. [Google Scholar] [CrossRef] [PubMed]
- Fornetti, J.; Welm, A.L.; Stewart, S.A. Understanding the bone in cancer metastasis. J. Bone Miner. Res. 2018, 33, 2099–2113. [Google Scholar] [CrossRef] [PubMed]
- Kamiya, N.; Suzuki, H.; Endo, T.; Yano, M.; Naoi, M.; Nishimi, D.; Kawamura, K.; Imamoto, T.; Ichikawa, T. Clinical usefulness of bone markers in prostate cancer with bone metastasis. Int. J. Urol. Off. J. Jpn. Urol. Assoc. 2012, 19, 968–979. [Google Scholar] [CrossRef]
- Lynch, M.E.; Brooks, D.; Mohanan, S.; Lee, M.J.; Polamraju, P.; Dent, K.; Bonassar, L.J.; van der Meulen, M.C.; Fischbach, C. In vivo tibial compression decreases osteolysis and tumor formation in a human metastatic breast cancer model. J. Bone Miner. Res. 2013, 28, 2357–2367. [Google Scholar] [CrossRef]
- Amin, S. Mechanical factors and bone health: Effects of weightlessness and neurologic injury. Curr. Rheumatol. Rep. 2010, 12, 170–176. [Google Scholar] [CrossRef]
- Goodman, C.A.; Hornberger, T.A.; Robling, A.G. Bone and skeletal muscle: Key players in mechanotransduction and potential overlapping mechanisms. Bone 2015, 80, 24–36. [Google Scholar] [CrossRef]
- Moore, E.R.; Zhu, Y.X.; Ryu, H.S.; Jacobs, C.R. Periosteal progenitors contribute to load-induced bone formation in adult mice and require primary cilia to sense mechanical stimulation. Stem Cell Res. Ther. 2018, 9, 190. [Google Scholar] [CrossRef]
- Liphardt, A.M.; Windahl, S.H.; Sehic, E.; Hannemann, N.; Gustafsson, K.L.; Bozec, A.; Schett, G.; Engdahl, C. Changes in mechanical loading affect arthritis-induced bone loss in mice. Bone 2020, 131, 115149. [Google Scholar] [CrossRef]
- Yang, S.; Liu, H.; Zhu, L.; Li, X.; Liu, D.; Song, X.; Yokota, H.; Zhang, P. Ankle loading ameliorates bone loss from breast cancer-associated bone metastasis. Off. Publ. Fed. Am. Soc. Exp. Biol. 2019, 33, 10742–10752. [Google Scholar] [CrossRef]
- Wang, X.; He, Y.; Tian, S.; Zhu, F.; Huang, B.; Zhang, J.; Chen, Z.; Wang, H. Fluid shear stress increases osteocyte and inhibits osteoclasts via downregulating receptor-activator of nuclear factor kappaB (RANK)/osteoprotegerin expression in myeloma microenvironment. Med Sci. Monit. Int. Med J. Exp. Clin. Res. 2019, 25, 5961–5968. [Google Scholar] [CrossRef]
- Takao, S.; Taya, M.; Chiew, C. Mechanical stress-induced cell death in breast cancer cells. Biol. Open 2019, 8. [Google Scholar] [CrossRef] [PubMed]
- Norwitz, N.G.; Mota, A.S.; Misra, M.; Ackerman, K.E. LRP5, Bone density, and mechanical stress: A case report and literature review. Front. Endocrinol. 2019, 10, 184. [Google Scholar] [CrossRef] [PubMed]
- Kang, K.S.; Hong, J.M.; Horan, D.J.; Lim, K.E.; Bullock, W.A.; Bruzzaniti, A.; Hann, S.; Warman, M.L.; Robling, A.G. Induction of Lrp5 HBM-causing mutations in Cathepsin-K expressing cells alters bone metabolism. Bone 2019, 120, 166–175. [Google Scholar] [CrossRef] [PubMed]
- Sawakami, K.; Robling, A.G.; Ai, M.; Pitner, N.D.; Liu, D.; Warden, S.J.; Li, J.; Maye, P.; Rowe, D.W.; Duncan, R.L.; et al. The Wnt co-receptor LRP5 is essential for skeletal mechanotransduction but not for the anabolic bone response to parathyroid hormone treatment. J. Biol. Chem. 2006, 281, 23698–23711. [Google Scholar] [CrossRef]
- Saxon, L.K.; Jackson, B.F.; Sugiyama, T.; Lanyon, L.E.; Price, J.S. Analysis of multiple bone responses to graded strains above functional levels, and to disuse, in mice in vivo show that the human Lrp5 G171V High Bone Mass mutation increases the osteogenic response to loading but that lack of Lrp5 activity reduces it. Bone 2011, 49, 184–193. [Google Scholar] [CrossRef][Green Version]
- Park, S.H.; Eber, M.R.; Shiozawa, Y. Models of prostate cancer bone metastasis. Methods Mol. Biol. 2019, 1914, 295–308. [Google Scholar] [CrossRef]
- Li, X.; Liu, D.; Li, J.; Yang, S.; Xu, J.; Yokota, H.; Zhang, P. Wnt3a involved in the mechanical loading on improvement of bone remodeling and angiogenesis in a postmenopausal osteoporosis mouse model. Off. Publ. Fed. Am. Soc. Exp. Biol. 2019, 33, 8913–8924. [Google Scholar] [CrossRef]
- Zengin, A.; Zhang, L.; Herzog, H.; Baldock, P.A.; Sainsbury, A. Neuropeptide Y and sex hormone interactions in humoral and neuronal regulation of bone and fat. Trends Endocrinol. Metab. 2010, 21, 411–418. [Google Scholar] [CrossRef]
- Igwe, J.C.; Jiang, X.; Paic, F.; Ma, L.; Adams, D.J.; Baldock, P.A.; Pilbeam, C.C.; Kalajzic, I. Neuropeptide Y is expressed by osteocytes and can inhibit osteoblastic activity. J. Cell. Biochem. 2009, 108, 621–630. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Wu, D.; Sun, X.; Fan, Y.; Zha, R.; Jalali, A.; Teli, M.; Sano, T.; Siegel, A.; Sudo, A.; et al. Mechanical stimulations can inhibit local and remote tumor progression by downregulating WISP1. Off. Publ. Fed. Am. Soc. Exp. Biol. 2020, 34, 12847–12859. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Fan, Y.; Liu, S.; Woollam, M.D.; Sun, X.; Murao, E.; Zha, R.; Prakash, R.; Park, C.; Siegel, A.P.; et al. Loading-induced antitumor capability of murine and human urine. Off. Publ. Fed. Am. Soc. Exp. Biol. 2020, 34, 7578–7592. [Google Scholar] [CrossRef] [PubMed]
- Qin, L.; Liu, W.; Cao, H.; Xiao, G. Molecular mechanosensors in osteocytes. Bone Res. 2020, 8, 23. [Google Scholar] [CrossRef] [PubMed]
- Kang, K.S.; Robling, A.G. New insights into Wnt-Lrp5/6-beta-catenin signaling in mechanotransduction. Front. Endocrinol. 2014, 5, 246. [Google Scholar] [CrossRef]
- Castillo, A.B.; Triplett, J.W.; Pavalko, F.M.; Turner, C.H. Estrogen receptor-beta regulates mechanical signaling in primary osteoblasts. Am. J. Physiol. Endocrinol. Metab. 2014, 306, E937–E944. [Google Scholar] [CrossRef]
- Swift, S.N.; Swift, J.M.; Bloomfield, S.A. Mechanical loading increases detection of estrogen receptor-alpha in osteocytes and osteoblasts despite chronic energy restriction. J. Appl. Physiol. 2014, 117, 1349–1355. [Google Scholar] [CrossRef]
- Lu, W.; Liu, C.C.; Thottassery, J.V.; Bu, G.; Li, Y. Mesd is a universal inhibitor of Wnt coreceptors LRP5 and LRP6 and blocks Wnt/beta-catenin signaling in cancer cells. Biochemistry 2010, 49, 4635–4643. [Google Scholar] [CrossRef]
- El-Sahli, S.; Xie, Y.; Wang, L.; Liu, S. Wnt signaling in cancer metabolism and immunity. Cancers 2019, 11, 904. [Google Scholar] [CrossRef]
- Shimojo, G.; Joseph, B.; Shah, R.; Consolim-Colombo, F.M.; De Angelis, K.; Ulloa, L. Exercise activates vagal induction of dopamine and attenuates systemic inflammation. Brain Behav. Immun. 2019, 75, 181–191. [Google Scholar] [CrossRef]
- Runegaard, A.H.; Fitzpatrick, C.M.; Woldbye, D.P.D.; Andreasen, J.T.; Sorensen, A.T.; Gether, U. Modulating dopamine signaling and behavior with chemogenetics: Concepts, progress, and challenges. Pharmacol. Rev. 2019, 71, 123–156. [Google Scholar] [CrossRef] [PubMed]
- Warner, R.L.; Johnston, C.; Hamilton, R.; Skolnick, M.H.; Wilson, O.B. Transcranial electrostimulation effects on rat opioid and neurotransmitter levels. Life Sci. 1994, 54, 481–490. [Google Scholar] [CrossRef]
- Liu, M.; Luo, J. Relationship between peripheral blood dopamine level and internet addiction disorder in adolescents: A pilot study. Int. J. Clin. Exp. Med. 2015, 8, 9943–9948. [Google Scholar] [PubMed]
- Suzuki, K.; Izumi, M.; Sakamoto, T.; Hayashi, M. Blood pressure and total cholesterol level are critical risks especially for hemorrhagic stroke in Akita, Japan. Cerebrovasc. Dis. 2011, 31, 100–106. [Google Scholar] [CrossRef] [PubMed]
- Martin, M.J.; Hulley, S.B.; Browner, W.S.; Kuller, L.H.; Wentworth, D. Serum cholesterol, blood pressure, and mortality: Implications from a cohort of 361,662 men. Lancet 1986, 2, 933–936. [Google Scholar] [CrossRef]
- Li, J.J.; Yin, H.K.; Guan, D.X.; Zhao, J.S.; Feng, Y.X.; Deng, Y.Z.; Wang, X.; Li, N.; Wang, X.F.; Cheng, S.Q.; et al. Chemerin suppresses hepatocellular carcinoma metastasis through CMKLR1-PTEN-Akt axis. Br. J. Cancer 2018, 118, 1337–1348. [Google Scholar] [CrossRef]
- Pachynski, R.K.; Wang, P.; Salazar, N.; Zheng, Y.; Nease, L.; Rosalez, J.; Leong, W.I.; Virdi, G.; Rennier, K.; Shin, W.J.; et al. Chemerin Suppresses breast cancer growth by recruiting immune effector cells into the tumor microenvironment. Front. Immunol. 2019, 10, 983. [Google Scholar] [CrossRef]
- Dubois-Vedrenne, I.; De Henau, O.; Robert, V.; Langa, F.; Javary, J.; Al Delbany, D.; Vosters, O.; Angelats-Canals, E.; Vernimmen, M.; Luangsay, S.; et al. Expression of bioactive chemerin by keratinocytes inhibits late stages of tumor development in a chemical model of skin carcinogenesis. Front. Oncol. 2019, 9, 1253. [Google Scholar] [CrossRef]
- Pachynski, R.K.; Zabel, B.A.; Kohrt, H.E.; Tejeda, N.M.; Monnier, J.; Swanson, C.D.; Holzer, A.K.; Gentles, A.J.; Sperinde, G.V.; Edalati, A.; et al. The chemoattractant chemerin suppresses melanoma by recruiting natural killer cell antitumor defenses. J. Exp. Med. 2012, 209, 1427–1435. [Google Scholar] [CrossRef]
- Rennier, K.; Shin, W.J.; Krug, E.; Virdi, G.; Pachynski, R.K. Chemerin reactivates PTEN and suppresses PD-L1 in tumor cells via modulation of a novel CMKLR1-mediated signaling cascade. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2020, 26, 5019–5035. [Google Scholar] [CrossRef]
- Peng, Y.; Kajiyama, H.; Yuan, H.; Nakamura, K.; Yoshihara, M.; Yokoi, A.; Fujikake, K.; Yasui, H.; Yoshikawa, N.; Suzuki, S.; et al. PAI-1 secreted from metastatic ovarian cancer cells triggers the tumor-promoting role of the mesothelium in a feedback loop to accelerate peritoneal dissemination. Cancer Lett. 2019, 442, 181–192. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Zhang, W.; Tang, L.; Chen, W.; Guan, X. Epithelial-mesenchymal transition induced PAI-1 is associated with prognosis of triple-negative breast cancer patients. Gene 2018, 670, 7–14. [Google Scholar] [CrossRef] [PubMed]
- He, W.; Tan, R.; Dai, C.; Li, Y.; Wang, D.; Hao, S.; Kahn, M.; Liu, Y. Plasminogen activator inhibitor-1 is a transcriptional target of the canonical pathway of Wnt/beta-catenin signaling. J. Biol. Chem. 2010, 285, 24665–24675. [Google Scholar] [CrossRef] [PubMed]
- Kozlova, N.; Jensen, J.K.; Chi, T.F.; Samoylenko, A.; Kietzmann, T. PAI-1 modulates cell migration in a LRP1-dependent manner via beta-catenin and ERK1/2. Thromb. Haemost. 2015, 113, 988–998. [Google Scholar] [CrossRef] [PubMed]
- Liu-Chittenden, Y.; Jain, M.; Gaskins, K.; Wang, S.; Merino, M.J.; Kotian, S.; Kumar Gara, S.; Davis, S.; Zhang, L.; Kebebew, E. RARRES2 functions as a tumor suppressor by promoting beta-catenin phosphorylation/degradation and inhibiting p38 phosphorylation in adrenocortical carcinoma. Oncogene 2017, 36, 3541–3552. [Google Scholar] [CrossRef]
- Kaiser, A.M.; Attardi, L.D. Deconstructing networks of p53-mediated tumor suppression in vivo. Cell Death Differ. 2018, 25, 93–103. [Google Scholar] [CrossRef]
- Pishavar, E.; Attaranzadeh, A.; Alibolandi, M.; Ramezani, M.; Hashemi, M. Modified PAMAM vehicles for effective TRAIL gene delivery to colon adenocarcinoma: in vitro and in vivo evaluation. Artif. Cells Nanomed. Biotechnol. 2018, 46, S503–S513. [Google Scholar] [CrossRef]
- Kim, H.; Lee, J.H.; Lee, S.K.; Song, N.Y.; Son, S.H.; Kim, K.R.; Chung, W.Y. Chemerin treatment inhibits the growth and bone invasion of breast cancer cells. Int. J. Mol. Sci. 2020, 21, 2871. [Google Scholar] [CrossRef]
- Jin, G.; Aobulikasimu, A.; Piao, J.; Aibibula, Z.; Koga, D.; Sato, S.; Ochi, H.; Tsuji, K.; Nakabayashi, T.; Miyata, T.; et al. A small-molecule PAI-1 inhibitor prevents bone loss by stimulating bone formation in a murine estrogen deficiency-induced osteoporosis model. FEBS Open Biol. 2018, 8, 523–532. [Google Scholar] [CrossRef]
- Brinker, A.E.; Vivian, C.J.; Beadnell, T.C.; Koestler, D.C.; Teoh, S.T.; Lunt, S.Y.; Welch, D.R. Mitochondrial haplotype of the host stromal microenvironment alters metastasis in a non-cell autonomous manner. Cancer Res. 2020, 80, 1118–1129. [Google Scholar] [CrossRef]
- Foster, B.A.; Gingrich, J.R.; Kwon, E.D.; Madias, C.; Greenberg, N.M. Characterization of prostatic epithelial cell lines derived from transgenic adenocarcinoma of the mouse prostate (TRAMP) model. Cancer Res. 1997, 57, 3325–3330. [Google Scholar] [PubMed]
- Johnstone, C.N.; Pattison, A.D.; Harrison, P.F.; Powell, D.R.; Lock, P.; Ernst, M.; Anderson, R.L.; Beilharz, T.H. FGF13 promotes metastasis of triple-negative breast cancer. Int. J. Cancer 2020, 147, 230–243. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wang, Y.; Chen, A.; Jalali, A.; Liu, S.; Guo, Y.; Na, S.; Nakshatri, H.; Li, B.Y.; Yokota, H. Effects of a checkpoint kinase inhibitor, AZD7762, on tumor suppression and bone remodeling. Int. J. Oncol. 2018, 53, 1001–1012. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Jalali, A.; Chen, A.; Zhao, X.; Liu, S.; Teli, M.; Guo, Y.; Li, F.; Li, J.; Siegel, A.; et al. Skeletal loading regulates breast cancer-associated osteolysis in a loading intensity-dependent fashion. Bone Res. 2020, 8, 9. [Google Scholar] [CrossRef]
- Li, F.; Chen, A.; Reeser, A.; Wang, Y.; Fan, Y.; Liu, S.; Zhao, X.; Prakash, R.; Kota, D.; Li, B.Y.; et al. Vinculin force sensor detects tumor-osteocyte interactions. Sci. Rep. 2019, 9, 5615. [Google Scholar] [CrossRef]
- Liu, S.; Fan, Y.; Chen, A.; Jalali, A.; Minami, K.; Ogawa, K.; Nakshatri, H.; Li, B.Y.; Yokota, H. Osteocyte-driven downregulation of snail restrains effects of Drd2 inhibitors on mammary tumor cells. Cancer Res. 2018, 78, 3865–3876. [Google Scholar] [CrossRef]
- Zhao, L.; Shim, J.W.; Dodge, T.R.; Robling, A.G.; Yokota, H. Inactivation of Lrp5 in osteocytes reduces young’s modulus and responsiveness to the mechanical loading. Bone 2013, 54, 35–43. [Google Scholar] [CrossRef]
- Hamamura, K.; Zhang, P.; Zhao, L.; Shim, J.W.; Chen, A.; Dodge, T.R.; Wan, Q.; Shih, H.; Na, S.; Lin, C.C.; et al. Knee loading reduces MMP13 activity in the mouse cartilage. BMC Musculoskelet. Disord. 2013, 14, 312. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feng, Y.; Liu, S.; Zha, R.; Sun, X.; Li, K.; Robling, A.; Li, B.; Yokota, H. Mechanical Loading-Driven Tumor Suppression Is Mediated by Lrp5-Dependent and Independent Mechanisms. Cancers 2021, 13, 267. https://doi.org/10.3390/cancers13020267
Feng Y, Liu S, Zha R, Sun X, Li K, Robling A, Li B, Yokota H. Mechanical Loading-Driven Tumor Suppression Is Mediated by Lrp5-Dependent and Independent Mechanisms. Cancers. 2021; 13(2):267. https://doi.org/10.3390/cancers13020267
Chicago/Turabian StyleFeng, Yan, Shengzhi Liu, Rongrong Zha, Xun Sun, Kexin Li, Alexander Robling, Baiyan Li, and Hiroki Yokota. 2021. "Mechanical Loading-Driven Tumor Suppression Is Mediated by Lrp5-Dependent and Independent Mechanisms" Cancers 13, no. 2: 267. https://doi.org/10.3390/cancers13020267
APA StyleFeng, Y., Liu, S., Zha, R., Sun, X., Li, K., Robling, A., Li, B., & Yokota, H. (2021). Mechanical Loading-Driven Tumor Suppression Is Mediated by Lrp5-Dependent and Independent Mechanisms. Cancers, 13(2), 267. https://doi.org/10.3390/cancers13020267