
The Role of Circular RNAs in Keratinocyte Carcinomas




Abstract
Simple Summary
Abstract
1. Introduction
2. Pathogenesis/Oncogenesis
3. Non-Coding RNAs
4. CircRNAs in SCC
5. CircRNAs in BCC
6. Future Perspectives
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Karimkani, C.; Boyers, L.N.; Dellavalle, R.P.; Weinstock, M.A. It’s time for “keratinocyte carcinoma” to replace the term “nonmelanoma skin cancer”. J. Am. Acad. Dermatol. 2015, 72, 186–187. [Google Scholar] [CrossRef] [PubMed]
- Peris, K.; Fargnoli, M.C.; Garbe, C.; Kaufmann, R.; Bastholt, L.; Seguin, N.B.; Bataille, V.; del Marmol, V.; Dummer, R.; Harwood, C.A. Diagnosis and treatment of basal cell carcinoma: European consensus-based interdisciplinary guidelines. Eur. J. Cancer 2019, 118, 10–34. [Google Scholar] [CrossRef] [PubMed]
- Fitzgerald, K.; Tsai, K.K. Systemic therapy for advanced cutaneous squamous cell carcinoma. Semin. Cutan. Med. Surg. 2019, 38, E67–E74. [Google Scholar] [CrossRef]
- Gender, R.E.; Weijns, M.E.; Dekkers, O.M.; Plasmeijer, E.I. Metastasis of cutaneous squamous cell carcinoma in organ transplant recipients and the immunocompetent population: Is there a difference? a systematic review and meta-analysis. J. Eur. Acad. Dermatol. Venereol. 2019, 33, 828–841. [Google Scholar] [CrossRef]
- Venables, Z.; Autier, P.; Nijsten, T.; Wong, K.F.; Langan, S.M.; Rous, B.; Broggio, J.; Harwood, C.; Henson, K.; Proby, C.M.; et al. Incidence of metastatic cutaneous squamous cell carcinoma: A nation-wide study in England. JAMA Dermatol. 2019, 155, 298–305. [Google Scholar] [CrossRef]
- Rogers, H.W.; Weinstock, M.A.; Feldman, S.R.; Coldiron, B.M. Incidence Estimate of Nonmelanoma Skin Cancer (Keratinocyte Carcinomas) in the US Population, 2012. JAMA Dermatol. 2015, 151, 1081. [Google Scholar] [CrossRef]
- Skin Cancer Foundation. Skin Cancer Facts. Available online: http://www.skincancer.org/skin-cancerinformation/skin-cancer-facts (accessed on 23 June 2021).
- Lomas, A.; Leonardi-Bee, J.; Bath-Hextall, F. A systematic review of worldwide incidence of nonmelanoma skin cancer. Br. J. Dermatol. 2012, 166, 1069e80. [Google Scholar] [CrossRef]
- Staples, M.P.; Elwood, M.; Burton, R.C.; Williams, J.L.; Marks, R.; Giles, G.G. Non-melanoma skin cancer in Australia: The 2002 national survey and trends since 1985. Med. J. Aust. 2006, 184, 6–10. [Google Scholar] [CrossRef] [PubMed]
- Katalinic, A.; Kunze, U.; Schafer, T. Epidemiology of cutaneous melanoma and non-melanoma skin cancer in Schleswig-Holstein, Germany: Incidence, clinical subtypes, tumour stages and localization (epidemiology of skin cancer). Br. J. Dermatol 2003, 149, 1200–1206. [Google Scholar] [CrossRef]
- Stang, A.; Khil, L.; Kajüter, H.; Pandeya, N.; Schmults, C.D.; Ruiz, E.S.; Karia, P.S.; Green, A.C. Incidence and mortality for cutaneous squamous cell carcinoma: Comparison across three continents. J. Eur. Acad. Dermatol. Venereol. 2019, 33 (Suppl. 8), 6–10. [Google Scholar] [CrossRef] [PubMed]
- Brewster, D.H.; Bhatti, L.A.; Inglis, J.H.; Nairn, E.R.; Doherty, V.R. Recent trends in incidence of nonmelanoma skin cancer in the east of scotland, 1992–2003. Br. J. Dermatol. 2007, 156, 1295–1300. [Google Scholar] [CrossRef]
- Birch-Johansen, F.; Jensen, A.; Mortensen, L.; Olesen, A.B.; Kjær, S.K. Trends in the incidence of nonmelanoma skin cancer in Denmark 1978–2007: Rapid incidence increase among young Danish women. Int. J. Cancer 2010, 127, 2190–2198. [Google Scholar] [CrossRef] [PubMed]
- Christenson, L.J.; Borrowman, T.A.; Vachon, C.M.; Tollefson, M.M.; Otley, C.C.; Weaver, A.L.; Roenigk, R.K. Incidence of Basal Cell and Squamous Cell Carcinomas in a Population Younger Than 40 Years. JAMA 2005, 294, 681–690. [Google Scholar] [CrossRef] [PubMed]
- Brash, D.E.; Rudolph, J.A.; Simon, J.A.; Lin, A.; McKenna, G.J.; Baden, H.P.; Halperin, A.J.; Pontén, J. A role for sunlight in skin cancer: UV-induced p53 mutations in squamous cell carcinoma. Proc. Natl. Acad. Sci. USA 1991, 88, 10124–10128. [Google Scholar] [CrossRef] [PubMed]
- Bernard, J.J.; Gallo, R.L.; Krutmann, J. Photoimmunology: How ultraviolet radiation affects the immune system. Nat. Rev. Immunol. 2019, 19, 688–701. [Google Scholar] [CrossRef] [PubMed]
- Hart, P.H.; Norval, M. Ultraviolet radiation-induced immunosuppression and its relevance for skin carcinogenesis. Photochem. Photobiol. Sci. 2018, 17, 1872–1884. [Google Scholar] [CrossRef]
- Peterson, S.C.; Eberl, M.; Vagnozzi, A.N.; Belkadi, A.; Veniaminova, N.A.; Verhaegen, M.E.; Bichakjian, C.K.; Ward, N.L.; Dlugosz, A.A.; Wong, S.Y. Basal cell carcinoma preferentially arises from stem cells within hair follicle and mechanosensory niches. Cell Stem Cell 2015, 16, 400–412. [Google Scholar] [CrossRef]
- Pellegrini, C.; Maturo, M.G.; Di Nardo, L.; Ciciarelli, V.; Gutierrez Garcıa-Rodrigo, C.; Fargnoli, M.C. Understanding the molecular genetics of basal cell carcinoma. Int. J. Mol. Sci. 2017, 18, 2485. [Google Scholar] [CrossRef]
- Bonilla, X.; Parmentier, L.; King, B.; Bezrukov, F.; Kaya, G.; Zoete, V.; Seplyarskiy, V.B.; Sharpe, H.J.; McKee, T.; Letourneau, A.; et al. Genomic analysis identifies new drivers and progression pathways in skin basal cell carcinoma. Nat. Genet. 2016, 48, 398–406. [Google Scholar] [CrossRef]
- Saenz-Sardà, X.; Carrato, C.; Pérez-Roca, L.; Puig, L.; Ferrendiz, C.; Ariza, A.; Fernandez-Figueras, M.-T. Epithelial-to-mesenchymal transition contributes to invasion in squamous cell carcinomas originated from actinic keratosis through the differentiated pathway, whereas proliferation plays a more significant role in the classical pathway. J. Eur. Acad. Dermatol. Venereol. 2018, 32, 581–586. [Google Scholar] [CrossRef]
- Rodríguez-Paredes, M.; Bormann, F.; Raddatz, G.; Gutekunst, J.; Lucena-Porcel, C.; Köhler, F.; Wurzer, E.; Schmidt, K.; Gallinat, S.; Wenck, H.; et al. Methylation profiling identifies two subclasses of squamous cell carcinoma related to distinct cells of origin. Nat. Commun. 2018, 9, 577. [Google Scholar] [CrossRef]
- Nagarajan, P.; Asgari, M.M.; Green, A.C.; Guhan, S.M.; Arron, S.T.; Proby, C.M.; Rollison, D.E.; Harwood, C.A.; Toland, A.E. Keratinocyte Carcinomas: Current Concepts and Future Research Priorities. Clin. Cancer Res. 2019, 25, 2379–2391. [Google Scholar] [CrossRef] [PubMed]
- Que, S.K.T.; Zwald, F.O.; Schmults, C.D. Cutaneous squamous cell carcinoma: Incidence, risk factors, diagnosis, and staging. J. Am. Acad. Dermatol. 2018, 78, 237–247. [Google Scholar] [CrossRef] [PubMed]
- Proby, C.M.; Harwood, C.A.; Neale, R.E.; Green, A.C.; Euvrard, S.; Naldi, L.; Tessari, G.; Feltkamp, M.C.; de Koning, M.N.; Quint, W.G.; et al. A case-control study of betapapillomavirus infection and cutaneous squamous cell carcinoma in organ transplant recipients. Am. J. Transplant. 2011, 11, 1498–1508. [Google Scholar] [CrossRef]
- Arron, S.T.; Ruby, J.G.; Dybbro, E.; Ganem, D.; Derisi, J.L. Transcriptome sequencing demonstrates that human papillomavirus is not active in cutaneous squamous cell carcinoma. J. Investig. Dermatol. 2011, 131, 1745–1753. [Google Scholar] [CrossRef]
- Hodorogea, A.; Calinescu, A.; Antohe, M.; Balaban, M.; Nedelcu, R.I.; Turcu, G.; Ion, D.A.; Badarau, I.A.; Popescu, C.M.; Popescu, R.; et al. Epithelial-Mesenchymal Transition in Skin Cancers: A Review. Anal. Cell. Pathol. 2019, 385, 1576. [Google Scholar] [CrossRef] [PubMed]
- Lamouille, S.; Xu, J.; Derynck, R. Molecular mechanisms of epithelialmesenchymal transition. Nat. Rev. Mol. Cell Biol. 2014, 3, 178–196. [Google Scholar] [CrossRef]
- Barrette, K.; Van Kelst, S.; Wouters, J.; Marasigan, V.; Fieuws, S.; Agostinis, P.; van den Oord, J.; Garmyn, M. Epithelial-mesenchymal transition during invasion of cutaneous squamous cell carcinoma is paralleled by AKT activation. Br. J. Dermatol. 2014, 171, 1014–1021. [Google Scholar] [CrossRef] [PubMed]
- Toll, A.; Masferrer, E.; Hernandez-Ruiz, M.E.; Ferrandiz-Pulido, C.; Yébenes, M.; Jaka, A.; Tuneu, A.; Jucglà, A.; Gimeno, J.; Baró, T.; et al. Epithelial to mesenchymal transition markers are associated with an increased metastatic risk in primary cutaneous squamous cell carcinomas but are attenuated in lymph node metastases. J. Dermatol. Sci. 2013, 72, 93–102. [Google Scholar] [CrossRef]
- Murata, M.; Ito, T.; Tanaka, Y.; Yamamura, K.; Furue, K.; Furue, M. OVOL2-Mediated ZEB1 Downregulation May Prevent Promotion of Actinic Keratosis to Cutaneous Squamous Cell Carcinoma. J. Clin. Med. 2020, 9, 618. [Google Scholar] [CrossRef]
- Bakshi, A.; Shafi, R.; Nelson, J.; Cantrell, W.C.; Subhadarshani, S.; Andea, A.; Athar, M.; Elmets, C.A. The clinical course of actinic keratosis correlates with underlying molecular mechanisms. Br. J. Dermatol. 2020, 182, 995–1002. [Google Scholar] [CrossRef]
- Gambichler, T.; Steuke, A.K.; Schmitz, L.; Stockfleth, E.; Becker, J.C. Altered hydroxymethylation in cutaneous squamous cell carcinoma and keratoacanthoma. Br. J. Dermatol. 2020, 183, 955–957. [Google Scholar] [CrossRef]
- Sand, M.; Skrygan, M.; Georgas, D.; Sand, D.; Hahn, S.A.; Gambichler, T.; Altmeyer, P.; Bechara, F.G. Microarray analysis of microRNA expression in cutaneous squamous cell carcinoma. J. Dermatol. Sci. 2012, 68, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Sand, M.; Hessam, S.; Amur, S.; Skrygan, M.; Bromba, M.; Stockfleth, E.; Gambichler, T.; Bechara, F.G. Expression of oncogenic miR-17-92 and tumor suppressive miR-143-145 clusters in basal cell carcinoma and cutaneous squamous cell carcinoma. J. Dermatol. Sci. 2017, 86, 142–148. [Google Scholar] [CrossRef] [PubMed]
- Mizrahi, A.; Barzilai, A.; Gur-Wahnon, D.; Ben-Dov, I.Z.; Glassberg, S.; Meningher, T.; Elharar, E.; Masalha, M.; Jacob-Hirsch, J.; Tabibian-Keissar, H.; et al. Alterations of microRNAs throughout the malignant evolution of cutaneous squamous cell carcinoma: The role of miR-497 in epithelial to mesenchymal transition of keratinocytes. Oncogene 2018, 37, 218–230. [Google Scholar] [CrossRef] [PubMed]
- Dika, E.; Scarfì, F.; Ferracin, M.; Broseghini, E.; Marcelli, E.; Bortolani, B.; Campione, E.; Riefolo, M.; Ricci, C.; Lambertini, M. Basal Cell Carcinoma: A comprehensive review. Int. J. Mol. Sci. 2020, 21, 5572. [Google Scholar] [CrossRef]
- Chen, S.; Luo, L.; Chen, H.; He, C. The Current State of Research Regarding the Role of Non-Coding RNAs in Cutaneous Squamous Cell Carcinoma. Onco Targets Ther. 2020, 13, 13151–13158. [Google Scholar] [CrossRef]
- Zaravinos, A. The regulatory role of microRNAs in EMT and cancer. J. Oncol. 2015, 2015, 865816. [Google Scholar] [CrossRef]
- Sand, M.; Gambichler, T.; Sand, D.; Skrygan, M.; Altmeyer, P.; Bechara, F.G. MicroRNAs and the skin: Tiny players in the body’s largest organ. J. Dermatol. Sci. 2009, 53, 169–175. [Google Scholar] [CrossRef]
- Chen, H.; Xu, Z.; Liu, D. Small non-coding RNA and colorectal cancer. J. Cell. Mol. Med. 2019, 23, 3050–3057. [Google Scholar] [CrossRef]
- Schickel, R.; Boyerinas, B.; Park, S.M.; Peter, M.E. MicroRNAs: Key players in the immune system, differentiation, tumorigenesis and cell death. Oncogene 2008, 27, 5959–5974. [Google Scholar] [CrossRef]
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanisms, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef]
- Mogilyansky, E.; Rigoutsos, I. The miR-17/92 cluster: A comprehensive update on its genomics, genetics, functions and increasingly important and numerous roles in health and disease. Cell Death Differ. 2013, 12, 1603–1614. [Google Scholar] [CrossRef]
- Ota, A.; Tagawa, H.; Karnan, S.; Tsuzuki, S.; Karpas, A.; Kira, S.; Yoshida, Y.; Seto, M. Identification and characterization of a novel gene, C13orf25, as a target for 13q31-q32 amplification in malignant lymphoma. Cancer Res. 2004, 64, 3087–3095. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Huang, K.A.I.; Yu, J. Inhibition of microRNA-21 upregulates the expression of programmed cell death 4 and phosphatase tensin homologue in the A431 squamous cell carcinoma cell line. Oncol. Lett. 2014, 8, 203–207. [Google Scholar] [CrossRef]
- Darido, C.; Georgy, S.R.; Wilanowski, T.; Dworkin, S.; Auden, A.; Zhao, Q.; Rank, G.; Srivastava, S.; Finlay, M.J.; Papenfuss, A.T.; et al. Targeting of the tumor suppressor GRHL3 by a miR-21-dependent proto-oncogenic network results in PTEN loss and tumorigenesis. Cancer Cell 2011, 20, 635–648. [Google Scholar] [CrossRef] [PubMed]
- Lefort, K.; Brooks, Y.; Ostano, P.; Cario-André, M.; Calpini, V.; Guinea-Viniegra, J.; Albinger-Hegyi, A.; Hoetzenecker, W.; Kolfschoten, I.; Wagner, E.F.; et al. A miR-34a-SIRT6 axis in the squamous cell differentiation network. EMBO J. 2013, 32, 2248–2263. [Google Scholar] [CrossRef]
- Guo, X.; Gao, L.; Wang, Y.; Chiu, D.K.; Wang, T.; Deng, Y. Advances in long noncoding RNAs: Identification, structure prediction and function annotation. Brief. Funct. Genomics 2016, 15, 38–46. [Google Scholar] [CrossRef]
- Bergmann, J.H.; Spector, D.L. Long non-coding RNAs: Modulators of nuclear structure and function. Curr. Opin. Cell Biol. 2014, 26, 10–18. [Google Scholar] [CrossRef]
- Li, X.; Wu, Z.; Fu, X.; Han, W. LncRNAs: Insights into their function and mechanics in underlying disorders. Mutat. Res. Rev. Mutat. Res. 2014, 762, 1–21. [Google Scholar] [CrossRef]
- Mazar, J.; Sinha, S.; Dinger, M.E.; Mattick, J.S.; Perera, R.J. Proteincoding and non-coding gene expression analysis in differentiating human keratinocytes using a three-dimensional epidermal equivalent. Mol. Genet. Genomics 2010, 284, 1–9. [Google Scholar] [CrossRef]
- Sand, M.; Bechara, F.G.; Sand, D.; Gambichler, T.; Hahn, S.A.; Bromba, M.; Stockfleth, E.; Hessam, S. Expression profiles of long noncoding RNAs in cutaneous squamous cell carcinoma. Epigenomics 2016, 8, 501–518. [Google Scholar] [CrossRef]
- Yu, G.-J.; Sun, Y.; Zhang, D.-W.; Zhang, P. Long non-coding RNA HOTAIR functions as a competitive endogenous RNA to regulate PRAF2 expression by sponging miR-326 in cutaneous squamous cell carcinoma. Cancer Cell Int. 2019, 19, 270. [Google Scholar] [CrossRef] [PubMed]
- Qian, Z.; Wei, B.; Zhou, Y.; Wang, Q.; Wang, J.; Sun, Y.; Gao, Y.; Chen, X. PRAF2 overexpression predicts poor prognosis and promotes tumorigenesis in esophageal squamous cell carcinoma. BMC Cancer 2019, 19, 585. [Google Scholar] [CrossRef] [PubMed]
- Das Mahapatra, K.; Pasquali, L.; Søndergaard, J.N.; Lapins, J.; Nemeth, I.B.; Baltás, E.; Kemény, L.; Homey, B.; Moldovan, L.I.; Kjems, J.; et al. A comprehensive analysis of coding and non-coding transcriptomic changes in cutaneous squamous cell carcinoma. Sci. Rep. 2020, 10, 3637. [Google Scholar] [CrossRef]
- Li, J.; Yang, J.; Zhou, P.; Le, Y.; Zhou, C.; Wang, S.; Xu, D.; Lin, H.K.; Gong, Z. Circular RNAs in cancer: Novel insights into origins, properties, functions and implications. Am. J. Cancer Res. 2015, 15, 472–480. [Google Scholar]
- Dragomir, M.; Calin, G.A. Circular RNAs in Cancer—Lessons Learned From microRNAs. Front. Oncol. 2018, 28, 179. [Google Scholar] [CrossRef]
- Jeck, W.R.; Sorrentino, J.A.; Wang, K.; Slevin, M.K.; Burd, C.E.; Liu, J.; Marzluff, W.F.; Sharpless, N.E. Circular RNAs are abundant, conserved, and associated with ALU repeats. RNA 2013, 19, 141–157. [Google Scholar] [CrossRef] [PubMed]
- Meng, S.; Zhou, H.; Feng, Z.; Xu, Z.; Tang, Y.; Li, P.; Wu, M. CircRNA: Functions and properties of a novel potential biomarker for cancer. Mol. Cancer 2017, 16, 94. [Google Scholar] [CrossRef]
- Verduci, L.; Strano, S.; Yarden, Y.; Blandino, G. The circRNAmicroRNA code: Emerging implications for cancer diagnosis and treatment. Mol. Oncol. 2019, 13, 669–680. [Google Scholar] [CrossRef]
- Noto, J.J.; Schmidt, C.A.; Matera, A.G. Engineering and expressing circular RNAs via tRNA splicing. RNA Biol. 2017, 14, 978–984. [Google Scholar] [CrossRef]
- Kristensen, L.S.; Hansen, T.B.; Veno, M.T.; Kjems, J. Circular RNAs in cancer: Opportunities and challenges in the field. Oncogene 2018, 37, 555–565. [Google Scholar] [CrossRef]
- Chen, G.; Shi, Y.; Liu, M.; Sun, J. circHIPK3 regulates cell proliferation and migration by sponging miR-124 and regulating AQP3 expression in hepatocellular carcinoma. Cell Death Dis. 2018, 9, 1–13. [Google Scholar] [CrossRef]
- Li, Y.; Zheng, F.; Xiao, X.; Xie, F.; Tao, D.; Huang, C. CircHIPK3 sponges miR-558 to suppress heparanase expression in bladder cancer cells. EMBO Rep. 2017, 18, 1646–1659. [Google Scholar] [CrossRef] [PubMed]
- Xie, H.; Ren, X.; Xin, S.; Lan, X.; Lu, G.; Lin, Y.; Yang, S.; Zeng, Z.; Liao, W.; Ding, Y.-Q.; et al. Emerging roles of circRNA_001569 targeting miR-145 in the proliferation and invasion of colorectal cancer. Oncotarget 2016, 7, 26680–26691. [Google Scholar] [CrossRef] [PubMed]
- Sand, M.; Bechara, F.G.; Gambichler, T.; Sand, D.; Bromba, M.; Hahn, S.A.; Stockfleth, E.; Hessam, S.J. Circular RNA expression in cutaneous squamous cell carcinoma. Dermatol. Sci. 2016, 83, 210–218. [Google Scholar] [CrossRef] [PubMed]
- An, X.; Liu, X.; Ma, G.; Li, C. Upregulated circular RNA circ_0070934 facilitates cutaneous squamous cell carcinoma cell growth and invasion by sponging miR-1238 and miR-1247-5p. Biochem. Biophys. Res. Commun. 2019, 513, 380–385. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.; Jin, H.J.; Zhang, D.; Lin, Q. Silencing circRNA_001937 may inhibit cutaneous squamous cellcarcinoma proliferation and induce apoptosis by preventing the sponging of the miRNA-597-3p/FOSL2 pathway. Int. J. Mol. Med. 2020, 46, 1653–1660. [Google Scholar] [CrossRef]
- Chen, S.; Ding, J.; Wang, Y.; Lu, T.; Wang, L.; Gao, X.; Chen, H.; Qu, L.; He, C. RNA-Seq Profiling of Circular RNAs and the Oncogenic Role of circPVT1 in Cutaneous Squamous Cell Carcinoma. Onco Targets Ther. 2020, 13, 6777–6788. [Google Scholar] [CrossRef]
- Zhang, D.W.; Wu, H.Y.; Zhu, C.R.; Wu, D.D. CircRNA hsa_circ_0070934 functions as a competitive endogenous RNA to regulate HOXB7 expression by sponging miR-1236-3p in cutaneous squamous cell carcinoma. Int. J. Oncol. 2020, 57, 478–487. [Google Scholar] [CrossRef]
- Chen, P.; Li, C.; Huang, H.; Liang, L.; Zhang, J.; Li, Q.; Wang, Q.; Zhang, S.; Zeng, K.; Zhang, X.; et al. Circular RNA profiles and the potential involvement of down-expression of hsa_circ_0001360 in cutaneous squamous cell carcinogenesis. J. FEBS Open Bio 2021, 11, 1209–1222. [Google Scholar] [CrossRef]
- Lu, X.; Gan, Q.; Gan, C. Circular RNA circSEC24A promotes Cutaneous Squamous Cell Carcinoma Progression by Regulating miR-1193/MAP3K9 Axis. Onco Targets Ther. 2021, 14, 653–666. [Google Scholar] [CrossRef]
- Li, X.; Kong, Y.; Li, H.; Xu, M.; Jiang, M.; Sun, W.; Xu, S. CircRNA circ_0067772 aggravates the malignant progression of cutaneous squamous cell carcinoma by regulating miR-1238-3p/FOXG1 axis. Genes Genomics 2021, 43, 491–501. [Google Scholar] [CrossRef]
- Cai, L.; Wang, Y.; Wu, J.; Wu, G. Hsa_circ_0008234 facilitates proliferation of cutaneous squamous cell carcinoma through targeting miR-127-5p to regulate ADCY7. Arch. Dermatol. Res. 2021. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Guo, H.; Yang, W.; Li, J. Exosomal circular RNA, RNA-seq profiling and the carcinogenic role of exosomal circ-CYP24A1 in cutaneous squamous cell carcinoma. Front. Med. 2021, 8, 675842. [Google Scholar] [CrossRef]
- Sand, M.; Bechara, F.G.; Sand, D.; Gambichler, T.; Hahn, S.A.; Bromba, M.; Stockfleth, E.; Hessam, S. Circular RNA expression in basal cell carcinoma. Epigenomics 2016, 8, 619–632. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Li, Y.; Li, L. Circular RNA hsa_Circ_0005795 mediates cell proliferation of cutaneous basal cell carcinoma via sponging miR-1231. Arch. Dermatol. Res. 2021. [Google Scholar] [CrossRef] [PubMed]
- Yamane, K.; Jinnin, M.; Etoh, T.; Kobayashi, Y.; Shimozono, N.; Fukushima, S.; Masuguchi, S.; Maruo, K.; Inoue, Y.; Ishihara, T.; et al. Down-regulation of miR-124/-214 in cutaneous squamous cell carcinoma mediates abnormal cell proliferation via the induction of ERK. J. Mol. Med. 2013, 91, 69–81. [Google Scholar] [CrossRef]
- White, R.A.; Neiman, J.M.; Reddi, A.; Han, G.; Birlea, S.; Mitra, D.; Dionne, L.; Fernandez, P.; Murao, K.; Ban, L.; et al. Epithelial stem cell mutations that promote squamous cell carcinoma metastasis. J. Clin. Investig. 2013, 123, 4390–4404. [Google Scholar] [CrossRef]
- Errico, M.C.; Jin, K.; Sukumar, S.; Care, A. The Widening Sphere of Influence of HOXB7 in Solid Tumors. Cancer Res. 2016, 76, 2857–2862. [Google Scholar] [CrossRef]
- Wang, J.; Li, C.; Xu, L.; Yang, C.; Zhang, X. MiR-1193 was sponged by LINC00963 and inhibited cutaneous squamous cell carcinoma progression by targeting SOX4. Pathology 2019, 215, 152600. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Li, Y.; Zheng, Q.P.; Bao, C.Y.; He, J.; Chen, B.; Lyu, D.; Zheng, B.; Xu, Y.; Long, Z.; et al. Circular RNA profile identifies circPVT1 as a proliferative factor and prognostic marker in gastric cancer. Cancer Lett. 2017, 388, 208–219. [Google Scholar] [CrossRef]
- Verduci, L.; Ferraiuolo, M.; Sacconi, A.; Ganci, F.; Vitale, J.; Colombo, T.; Paci, P.; Strano, S.; Macino, G.; Rajewski, N.; et al. The oncogenic role of circPVT1 in head and neck squamous cell carcinoma is mediated through the mutant p53/YAP/TEAD transcription-competent complex. Genome Biol. 2017, 18, 237. [Google Scholar] [CrossRef] [PubMed]
- Xu, N.; Zhang, L.; Meisgen, F.; Harada, M.; Heilborn, J.; Homey, B.; Grander, D.; Stahle, M.; Sonkoly, E.; Pivarcsi, A. MicroRNA-125b down-regulates matrix metallopeptidase 13 and inhibits cutaneous squamous cell carcinoma cell proliferation, migration, and invasion. J. Biol. Chem 2012, 287, 29899–29908. [Google Scholar] [CrossRef] [PubMed]
- Chan, D.W.; Liu, V.W.; To, R.M.; Chiu, P.M.; Lee, W.Y.; Yao, K.M.; Cheung, A.N.; Ngan, H.Y. Overexpression of FOXG1 contributes to TGFbeta resistance through inhibition of p21WAF1/CIP1 expression in ovarian cancer. Br. J. Cancer 2009, 101, 1433–1443. [Google Scholar] [CrossRef][Green Version]
- Dai, J.; Su, Y.; Zhong, S.; Cong, L.; Liu, B.; Yang, J.; Tao, Y.; He, Z.; Chen, C.; Jiang, Y. Exosomes: Key players in cancer and potential therapeutic strategy. Signal Transduct. Target. Ther. 2020, 5, 145. [Google Scholar] [CrossRef]
- Chen, B.; Wie, W.; Huang, X.; Xie, X.; Kong, Y.; Dai, D.; Yang, L.; Wang, J.; Tang, H.; Xie, X. circEPSTI1 as a Prognostic Marker and Mediator of Triple-Negative Breast Cancer Progression. Theranostics 2018, 8, 4003–4015. [Google Scholar] [CrossRef]
- Tan, X.; Tan, D.; Li, H.; Lin, Y.; Wen, Z.; Zeng, C. circEPSTI1 Acts as a ceRNA to Regulate the Progression of Osteosarcoma. Curr. Cancer Drug Targets 2020, 20, 288–294. [Google Scholar] [CrossRef]
- Wu, P.; Li, X.; Ye, D.; Yu, K.; Li, Y.; Tang, H.; Xu, G.; Yi, S.; Zhang, Z. Circular RNA circEPSTI1 accelerates cervical cancer progression via miR-375/409-3P/515-5p-SLC7A11 axis. Aging (Albany NY) 2021, 13, 4663–4673. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Wang, S.; Li, G.; Zhao, X.; Jiang, F.; Liu, J.; Tan, W. circEPSTI1 regulates ovarian cancer progression via decoying miR-942. J. Cell. Mol. Med. 2019, 23, 3597–3602. [Google Scholar] [CrossRef]
- Barbagallo, D.; Condorelli, A.; Ragusa, M.; Salito, L.; Sammito, M.; Banelli, B.; Caltabiano, R.; Barbagallo, G.; Zappalà, A.; Battaglia, R.; et al. Dysregulated miR-671-5p/CDR1-AS/CDR1/VSNL1 axis is involved in glioblastoma multiforme. Oncotarget 2016, 7, 4746–4759. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Gong, X.; Sun, L.; Zhou, Q.; Lu, B.; Zhu, L. The circular RNA Cdr1as act as an oncogene in hepatocellular carcinoma through targeting miR-7 expression. PLoS ONE 2016, 11, e0158347. [Google Scholar] [CrossRef]
- Tang, W.; Ji, M.; He, G.; Yang, L.; Niu, Z.; Jian, M.; Wie, Y.; Ren, L.; Xu, J. Silencing CDR1as inhibits colorectal cancer progression through regulating microRNA-7. Onco Targets Ther. 2017, 10, 2045–2056. [Google Scholar] [CrossRef] [PubMed]
- Russell, M.R.; Penikis, A.; Oldridge, D.A.; Alvarez-Dominguez, J.R.; McDaniel, L.; Diamond, M.; Padovan, O.; Raman, P.; Li, Y.; Wie, J.S.; et al. CASC15-S Is a Tumor Suppressor lncRNA at the 6p22 Neuroblastoma Susceptibility Locus. Cancer Res. 2015, 75, 3155–3166. [Google Scholar] [CrossRef]
- Sheng, L.; Wei, R. Long Non-Coding RNA-CASC15 Promotes Cell Proliferation, Migration, and Invasion by Activating Wnt/β-Catenin Signaling Pathway in Melanom. Pathobiology 2020, 87, 20–29. [Google Scholar] [CrossRef] [PubMed]
- Sand, M.; Skrygan, M.; Sand, D.; Georgas, D.; Hahn, S.A.; Gambichler, T.; Altmeyer, P.; Bechara, F.G. Expression of microRNAs in basal cell carcinoma. Br. J. Dermatol. 2012, 167, 847–855. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Thomson, J.M.; Hemann, M.T.; Hernando-Monge, E.; Mu, D.; Goodson, S.; Powers, S.; Cordon-Cardo, C.; Lowe, S.W.; Hannon, G.J.; et al. A microRNA polycistron as a potential human oncogene. Nature 2005, 435, 828–833. [Google Scholar] [CrossRef] [PubMed]
- Jeck, W.R.; Sharpless, N.E. Detecting and characterizing circular RNAs. Nat. Biotechnol. 2014, 32, 453–461. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Wan, J.; Wang, Z.; Li, L.; Jia, H.; Xing, S.; Chen, S.; Fan, X.; Li, R. miR-3196 acts as a Tumor Suppressor and Predicts Survival Outcomes in Patients With Gastric Cancer. Technol. Cancer Res. Treat. 2020, 19, 1533033820923427. [Google Scholar] [CrossRef]
- Li, Y.; Tan, W.; Neo, T.W.L.; Aung, M.O.; Wasser, S.; Lim, S.G.; Tan, T.M.C. Role of the miR-106b-25 microRNA cluster in hepatocellular carcinoma. Cancer Sci. 2009, 100, 1234–1242. [Google Scholar] [CrossRef]
- Jiang, Q.; Liu, C.; Li, C.-P.; Xu, S.-S.; Yao, M.-D.; Ge, H.-M.; Sun, Y.-N.; Li, X.-M.; Zhang, S.J.; Shan, K.; et al. Circular RNA-ZNF532 regulates diabetes-induced retinal pericyte degeneration and vascular dysfunction. J. Clin. Investig. 2020, 130, 3833–3847. [Google Scholar] [CrossRef]
- Hanniford, D.; Ulloa-Morales, A.; Karz, A.; Berzoti-Coelho, M.G.; Moubarak, R.S.; Sanchez-Sendra, B.; Kloetgen, A.; Davalos, V.; Imig, J.; Wu, P.; et al. Epigenetic silencing of CDR1as drives IGF2BP3-mediated melanoma invasion and metastasis. Cancer Cell 2020, 37, 55–70. [Google Scholar] [CrossRef]
- He, A.T.; Liu, J.; Li, F.; Yang, B.B. Targeting circular RNAs as a therapeutic approach: Current strategies and challenges. Signal Transduct. Target Ther. 2021, 6, 185. [Google Scholar] [CrossRef]
- Wahane, A.; Waghmode, A.; Kapphahn, A.; Dhuri, K.; Gupta, A.; Bahal, R. Role of Lipid-Based and Polymer-Based Non-Viral Vectors in Nucleic Acid Delivery for Next-Generation Gene Therapy. Molecules 2020, 25, 2866. [Google Scholar] [CrossRef] [PubMed]
- Tay, F.C.; Lim, J.K.; Zhu, H.; Hin, L.C.; Wang, S. Using artificial microRNA sponges to achieve microRNA loss-of-function in cancer cells. Adv. Drug Deliv. Rev. 2015, 81, 117–127. [Google Scholar] [CrossRef] [PubMed]
- Piwecka, M.; Glažar, P.; Hernandez-Miranda, L.R.; Memczak, S.; Wolf, S.A.; Rybak-Wolf, A.; Filipchyk, A.; Klironomos, F.; Cerda Jara, C.-A.; Fenske, P.; et al. Loss of a mammalian circular RNA locus causes miRNA deregulation and affects brain function. Science 2017, 357, eaam8526. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Li, X.; Xue, W.; Zhang, L.; Yang, L.Z.; Cao, S.M.; Lei, Y.N.; Liu, C.X.; Guo, S.K.; Lin, S.; et al. Screening for functional circular RNAs using the CRIPR-Cas 13 system. Nat. Methods 2021, 18, 51–59. [Google Scholar] [CrossRef]


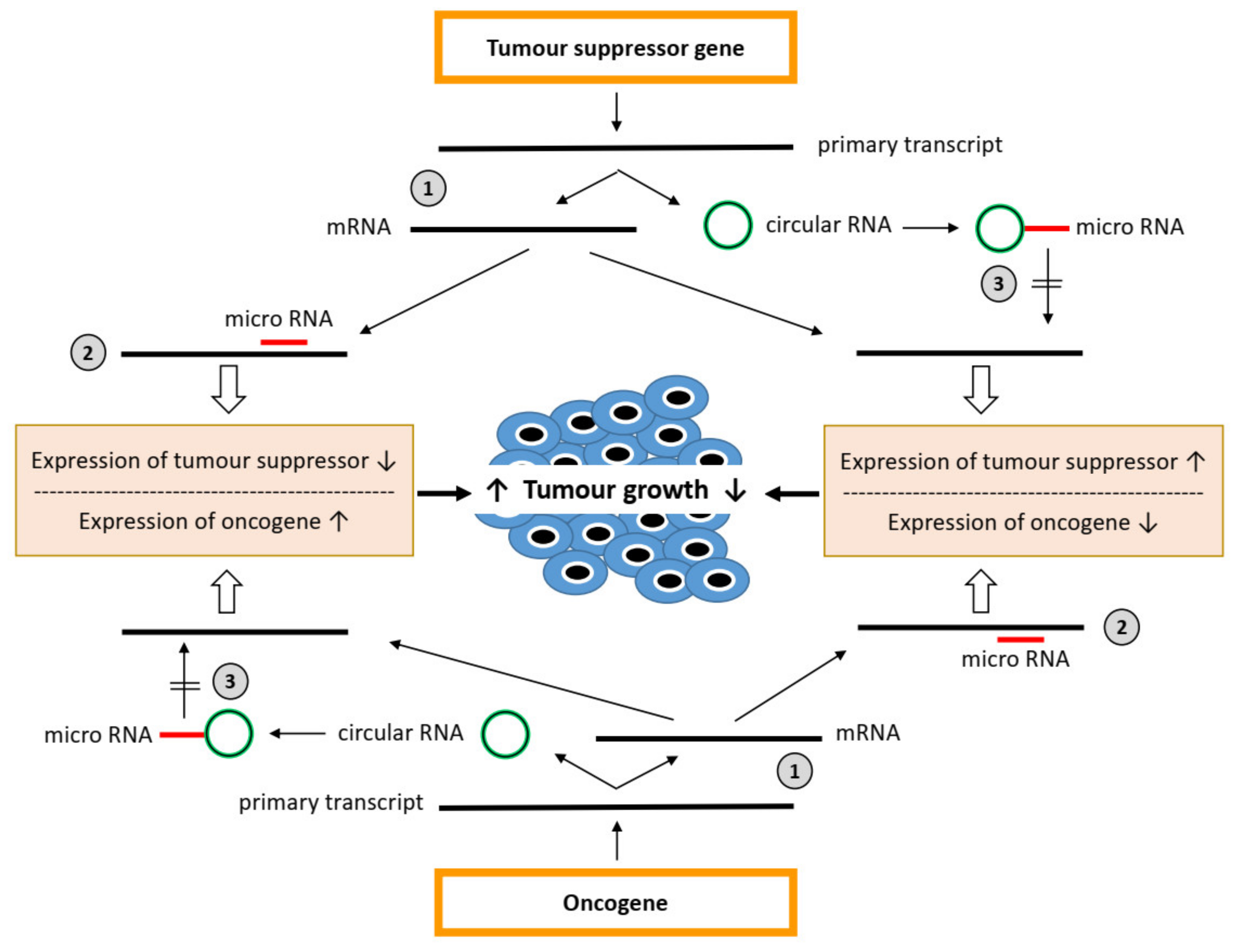{kind=link}
| cSCC | Reference | |
|---|---|---|
| Sand M et al. 2016 | Comparison of circular RNA expression in 3 cSCC and 3 healthy control tissues by microarray analysis | [67] |
| An X et al. 2019 | Functional characterization of hsa_circ_0070934 | [68] |
| Gao L et al. 2020 | Comparison of circular RNA expression in 3 cSCC and 3 healthy control tissues by microarray analysis and functional characterization of circRNA_001937 | [69] |
| Chen S et al. 2020 | Comparison of circRNAs in 30 paired cSCC and adjacent nontumorous tissues RNA sequencing and functional characterization of circ_PVT1 | [70] |
| Zhang et al. 2020 | Functional characterization of hsa_circ_0070934 | [71] |
| Das Mahapatra K et al. 2020 | Comparison of circRNAs, miRNAs, and lncRNAs in 9 cSCCs and 7 healthy skin samples by RNA sequencing | [56] |
| Chen P et al. 2021 | Comparison of circRNAs in 3 paired cSCC and adjacent non-tumorous tissues RNA sequencing and functional characterization of hsa_circ_0001360 | [72] |
| Lu X et al. 2021 | Functional characterization of circSEC24A | [73] |
| Li X et al. 2021 | Functional characterization of has_circ_0067772 | [74] |
| Cai L et al. 2021 | Functional characterization of hsa_circ_0008234 | [75] |
| Zhang Z et al. 2021 | Functional characterization of exosomal circ-CYP24A1 | [76] |
| BCC | ||
| Sand M et al. 2016 | Comparison of circular RNA expression in 3 BCC and 3 healthy control tissues by microarray analysis | [77] |
| Li Y et al. 2021 | Comparison of expression of hsa_circ_0005795 in 30 BCC and adjacent non-tumor tissues and functional characterization of hsa_circ_0005795 | [78] |
| Circular RNA | Dysregulation in KC | Interacting miRNAs | Cellular Effects | Ref. |
|---|---|---|---|---|
| hsa_circ_0070933 | Upregulated in cSCC | Not analyzed experimentally | Not analyzed experimentally | [67,71] |
| hsa_circ_0070934 | Upregulated in cSCC | miR-1238, miR-1247-5p miR-1236-3p | Increased cell proliferation, migration, and invasion, inhibits apoptosis | [67,68,71] |
| hsa_circ_0003528 (circSEC24A) | Upregulated in cSCC | miR-1193 | Increased cell proliferation, migration, and invasion, inhibits apoptosis | [73] |
| hsa_circ_001937 | Upregulated in cSCC | miR-597-3p | Increased cell proliferation | [69] |
| hsa_circ_0001821 (circPVT1) | Upregulated in cSCC | Not analyzed experimentally | Increased migration and invasion | [70] |
| hsa_circ_001360 | Downregulated in cSCC | Not analyzed experimentally | Reduced cell proliferation, migration, and invasion | [72] |
| hsa_circ_0067772 | Upregulated in cSCC | miR-1238-3p | Increased cell proliferation, migration and invasion | [74] |
| hsa_circ_0008234 | Upregulated in cSCC | miR-127-5p | Increased cell proliferation | [75] |
| Circ-CYP24A1 | Upregulated in cSCC | Not analyzed | Increased cell proliferation, migration, and invasion, reduced apoptosis | [76] |
| hsa_circ_0005795 | Upregulated in BCC | miR-1231 | Increased cell proliferation, inhibits apoptosis | [78] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meyer, T.; Sand, M.; Schmitz, L.; Stockfleth, E. The Role of Circular RNAs in Keratinocyte Carcinomas. Cancers 2021, 13, 4240. https://doi.org/10.3390/cancers13164240
Meyer T, Sand M, Schmitz L, Stockfleth E. The Role of Circular RNAs in Keratinocyte Carcinomas. Cancers. 2021; 13(16):4240. https://doi.org/10.3390/cancers13164240
Chicago/Turabian StyleMeyer, Thomas, Michael Sand, Lutz Schmitz, and Eggert Stockfleth. 2021. "The Role of Circular RNAs in Keratinocyte Carcinomas" Cancers 13, no. 16: 4240. https://doi.org/10.3390/cancers13164240
APA StyleMeyer, T., Sand, M., Schmitz, L., & Stockfleth, E. (2021). The Role of Circular RNAs in Keratinocyte Carcinomas. Cancers, 13(16), 4240. https://doi.org/10.3390/cancers13164240