The ITIM-Containing Receptor: Leukocyte-Associated Immunoglobulin-Like Receptor-1 (LAIR-1) Modulates Immune Response and Confers Poor Prognosis in Invasive Breast Carcinoma

 ,
, 

Abstract
Simple Summary
Abstract
1. Introduction
2. Results
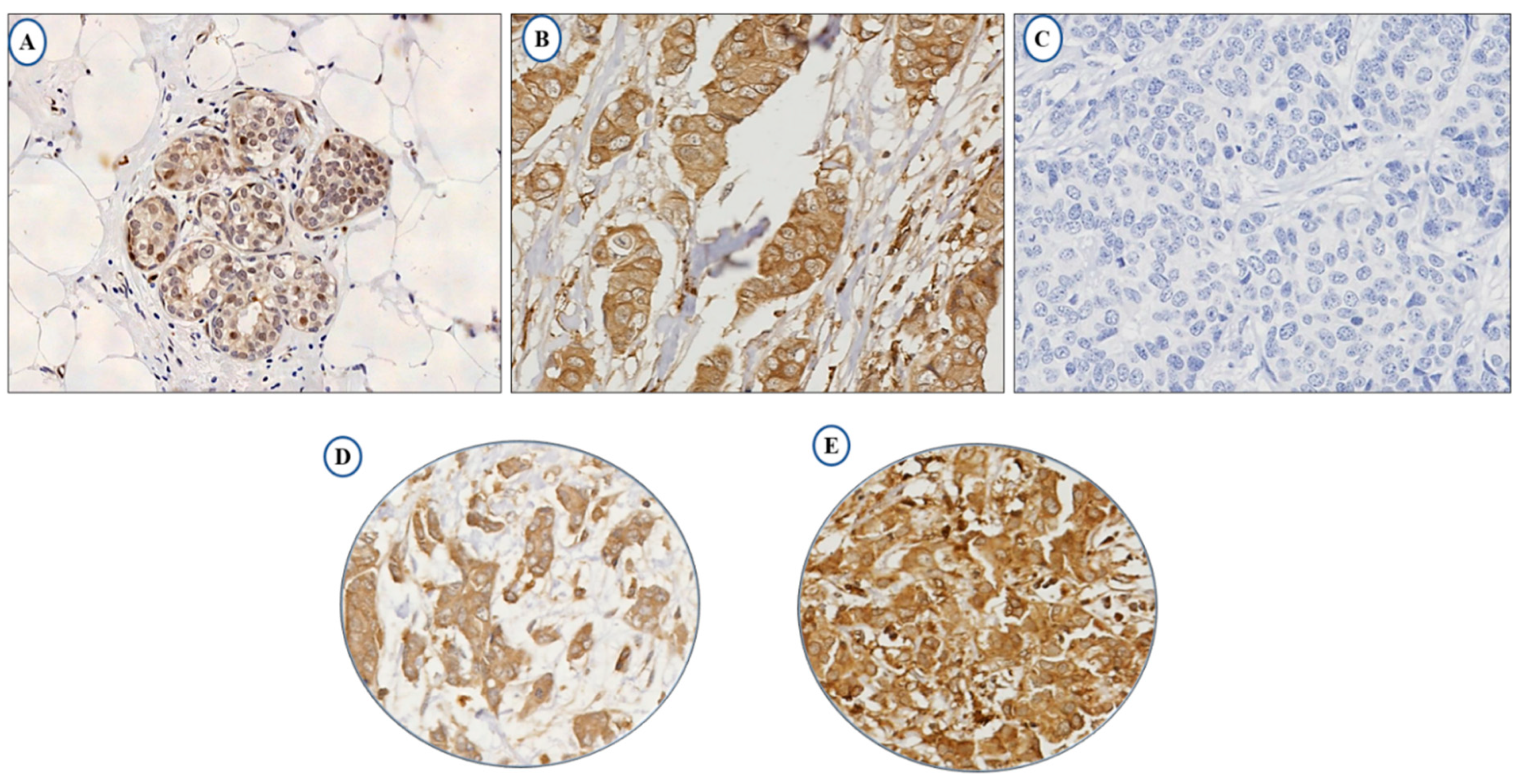
2.1. LAIR-1 Expression
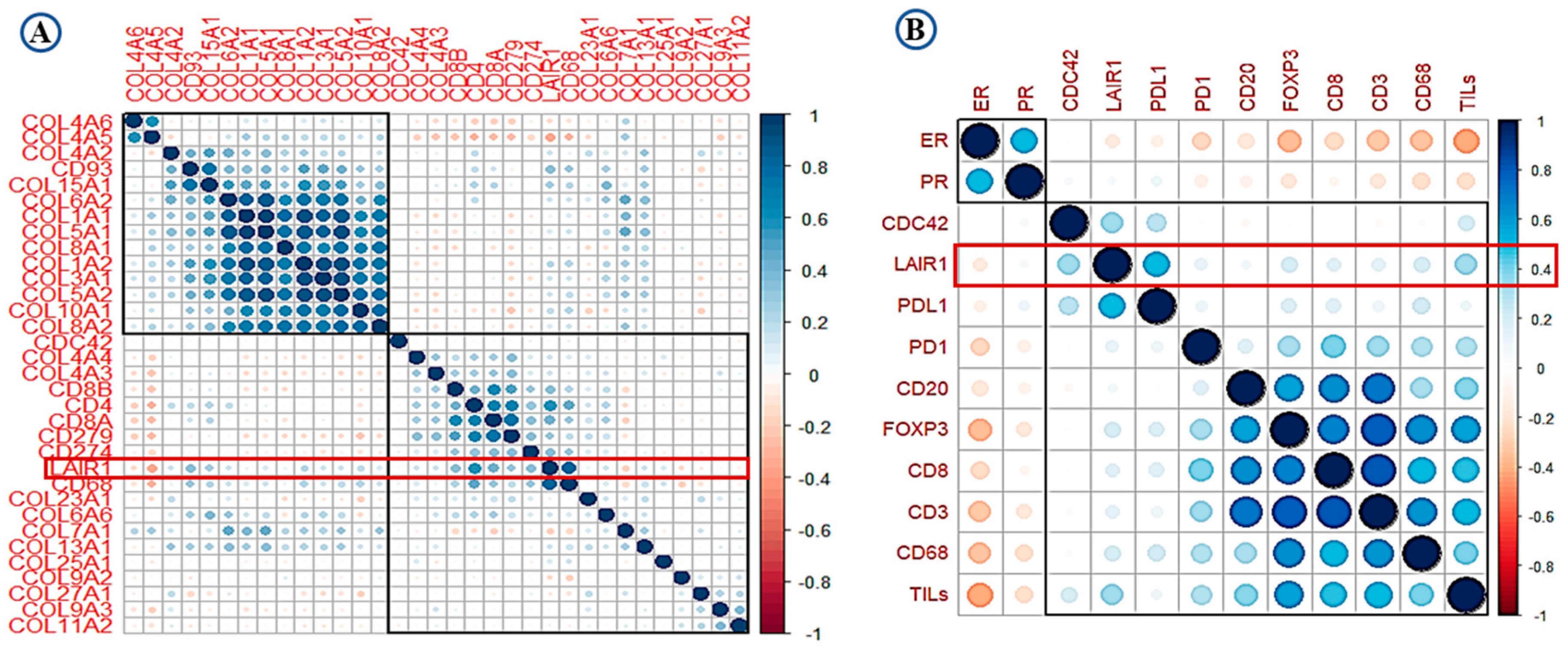
2.2. LAIR-1 Correlation with Collagens and Immune Cell Types
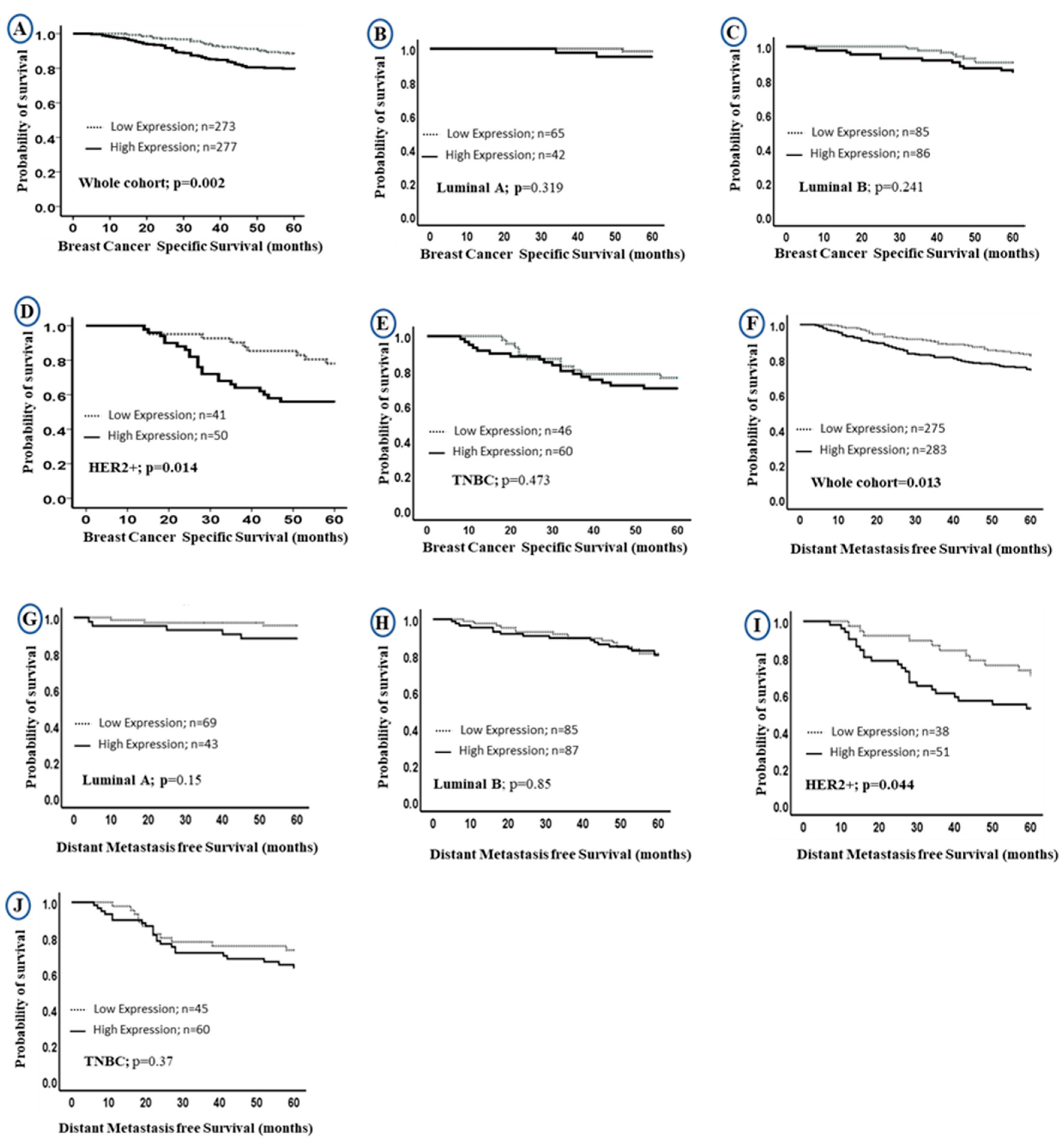
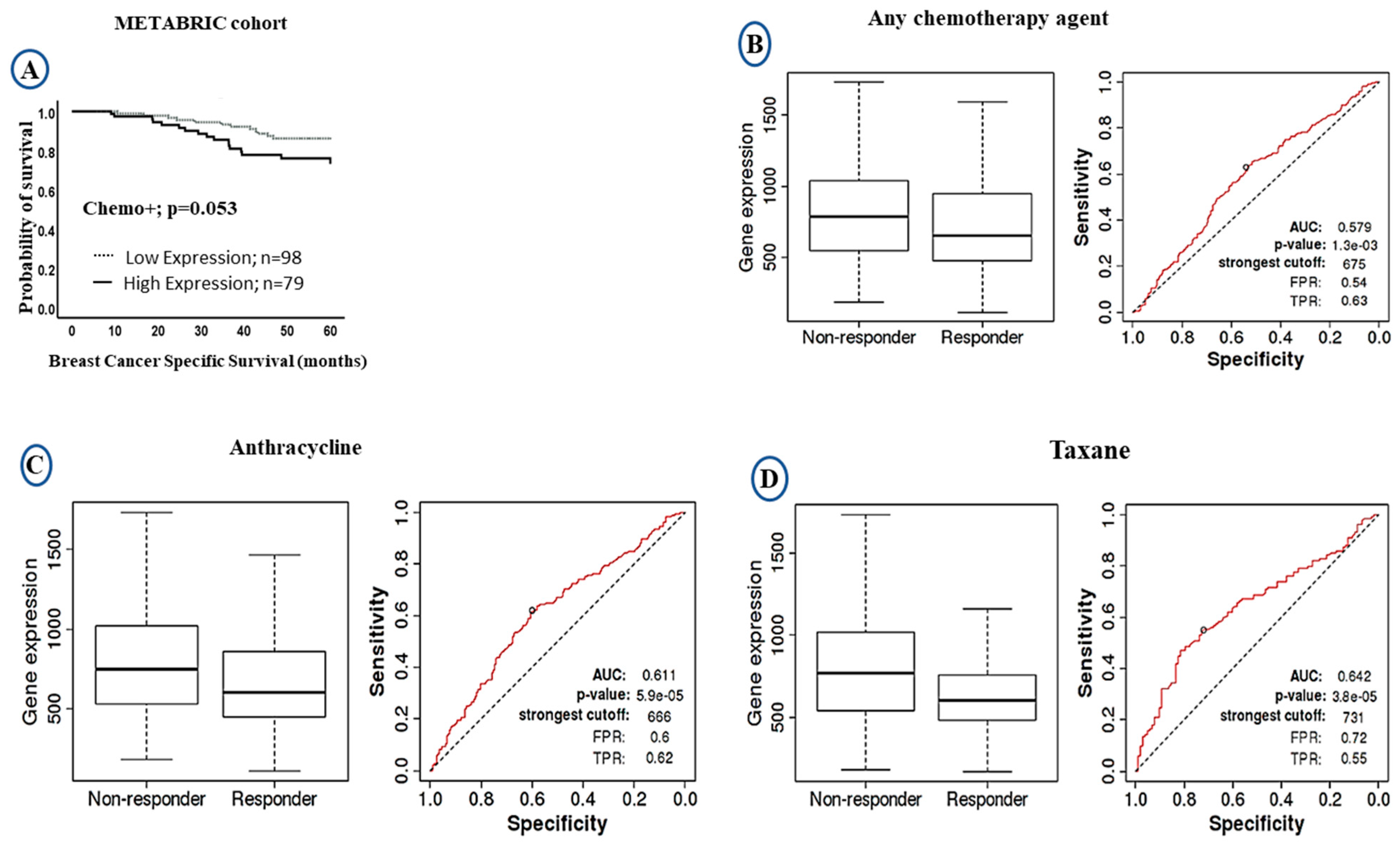
2.3. Prognostic Value of LAIR-1 Expression
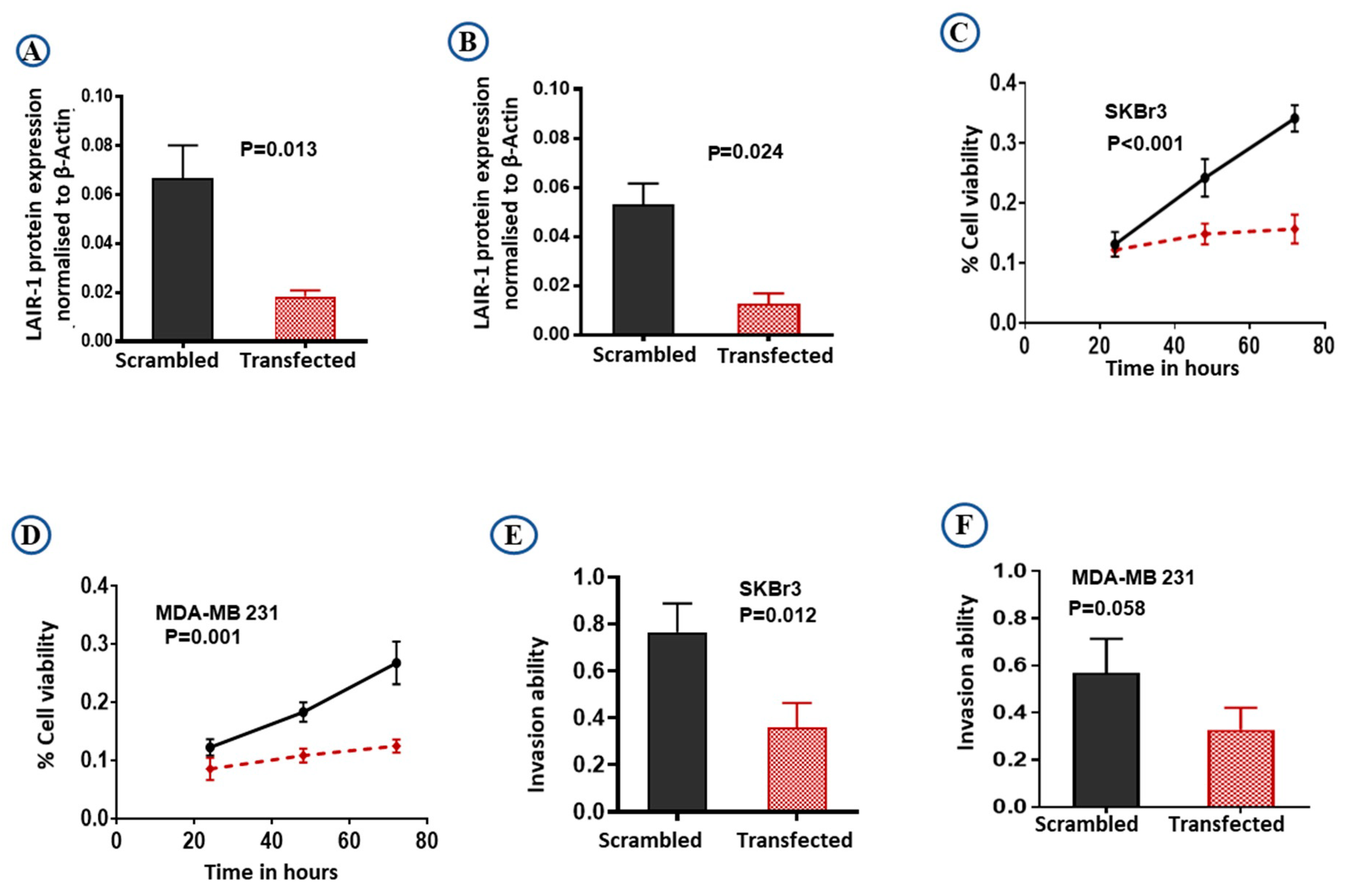
2.4. LAIR-1 Promotes Cell Proliferation and Invasion Ability in BC Cell Lines
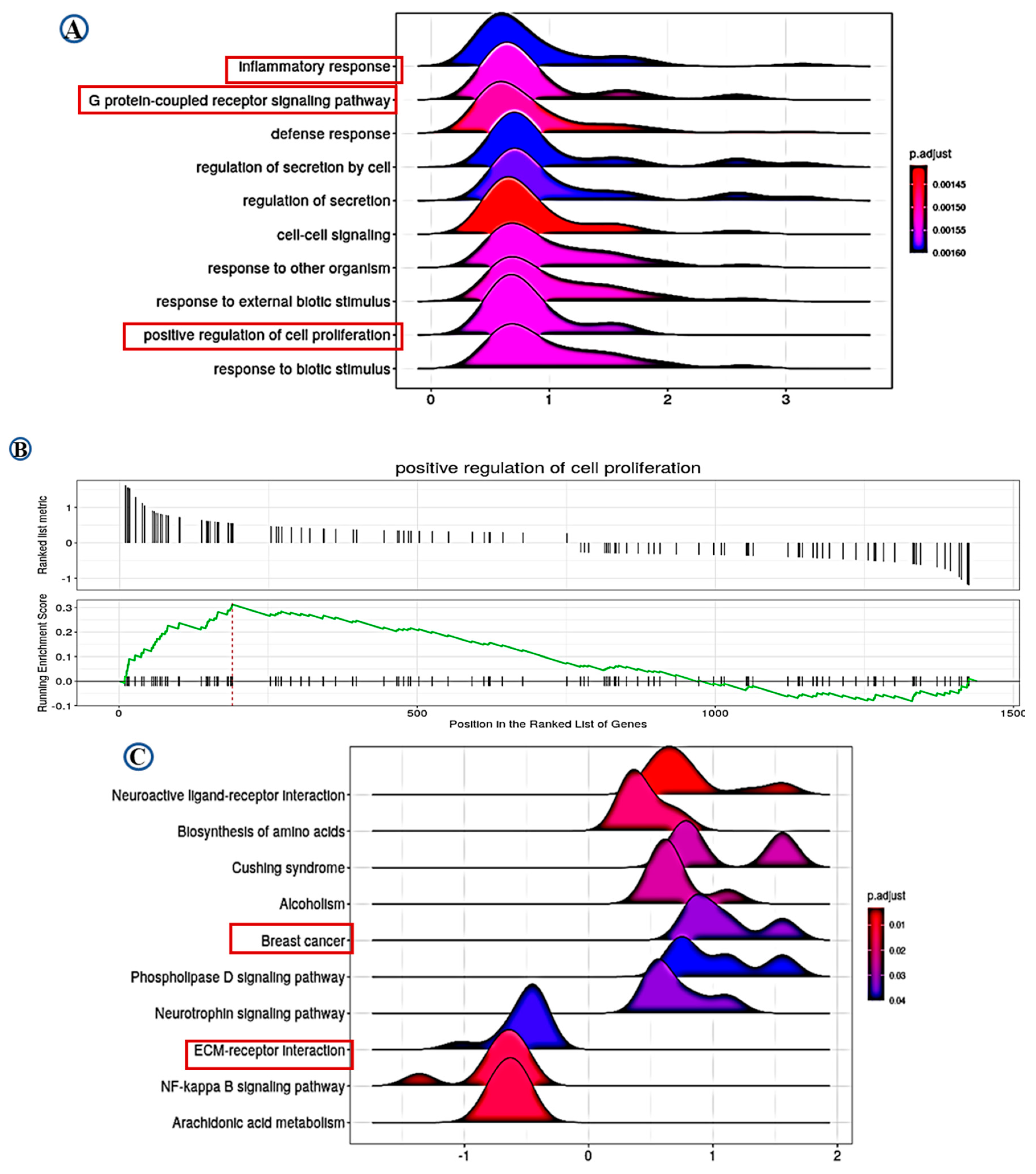
2.5. Pathway Analysis
3. Discussion
4. Materials and Methods
4.1. LAIR-1 mRNA Expression
4.2. LAIR-1 Protein Expression
4.2.1. Study Cohort
4.2.2. Immunohistochemistry (IHC)
4.2.3. In Vitro Studies
4.2.4. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Meyaard, L.; Hurenkamp, J.; Clevers, H.; Lanier, L.L.; Phillips, J.H. Leukocyte-associated Ig-like receptor-1 functions as an inhibitory receptor on cytotoxic T cells. J. Immunol. 1999, 162, 5800–5804. [Google Scholar] [PubMed]
- Singh, M.; Bhatia, P.; Shandilya, J.K.; Rawat, A.; Varma, N.; Sachdeva, M.S.; Trehan, A.; Bansal, D.; Jain, R.; Totadri, S. Low Expression of Leucocyte Associated Immunoglobulin Like Receptor-1 (LAIR-1/CD305) in a Cohort of Pediatric Acute Lymphoblastic Leukemia Cases. Asian Pac. J. Cancer Prev. 2018, 19, 3131–3135. [Google Scholar] [CrossRef] [PubMed]
- Lebbink, R.J.; De Ruiter, T.; Adelmeijer, J.; Brenkman, A.B.; Van Helvoort, J.M.; Koch, M.; Farndale, R.W.; Lisman, T.; Sonnenberg, A.; Lenting, P.J.; et al. Collagens are functional, high affinity ligands for the inhibitory immune receptor LAIR-1. J. Exp. Med. 2006, 203, 1419–1425. [Google Scholar] [CrossRef] [PubMed]
- Son, M.; Diamond, B. C1q-Mediated Repression of Human Monocytes Is Regulated by Leukocyte-Associated Ig-Like Receptor 1 (LAIR-1). Mol. Med. 2014, 20, 559–568. [Google Scholar] [CrossRef] [PubMed]
- Verbrugge, A.; Rijkers, E.S.K.; De Ruiter, T.; Meyaard, L. Leukocyte-associated Ig-like receptor-1 has SH2 domain-containing phosphatase-independent function and recruits C-terminal Src kinase. Eur. J. Immunol. 2006, 36, 190–198. [Google Scholar] [CrossRef] [PubMed]
- Fang, M.; Yuan, J.; Peng, C.; Li, Y. Collagen as a double-edged sword in tumor progression. Tumor Biol. 2014, 35, 2871–2882. [Google Scholar] [CrossRef]
- Brown, D.; Trowsdale, J.; Allen, R. The LILR family: Modulators of innate and adaptive immune pathways in health and disease. Tissue Antigens 2004, 64, 215–225. [Google Scholar] [CrossRef]
- Poggi, A.; Catellani, S.; Bruzzone, A.; Caligaris-Cappio, F.; Gobbi, M.; Zocchi, M.R. Lack of the leukocyte-associated Ig-like receptor-1 expression in high-risk chronic lymphocytic leukaemia results in the absence of a negative signal regulating kinase activation and cell division. Leukemia 2008, 22, 980–988. [Google Scholar] [CrossRef]
- Perbellini, O.; Falisi, E.; Giaretta, I.; Boscaro, E.; Novella, E.; Facco, M.; Fortuna, S.; Finotto, S.; Amati, E.; Maniscalco, F.; et al. Clinical significance of LAIR1 (CD305) as assessed by flow cytometry in a prospective series of patients with chronic lymphocytic leukemia. Haematologica 2014, 99, 881–887. [Google Scholar] [CrossRef]
- Wu, X.; Zhang, L.; Zhou, J.; Liu, L.; Fu, Q.; Fu, A.; Feng, X.; Xin, R.; Liu, H.; Gao, Y.; et al. Clinicopathologic significance of LAIR-1 expression in hepatocellular carcinoma. Curr. Probl. Cancer 2019, 43, 18–26. [Google Scholar] [CrossRef]
- Rygiel, T.P.; Stolte, E.H.; De Ruiter, T.; Van De Weijer, M.L.; Meyaard, L. Tumor-expressed collagens can modulate immune cell function through the inhibitory collagen receptor LAIR-1. Mol. Immunol. 2011, 49, 402–406. [Google Scholar] [CrossRef] [PubMed]
- Koukourakis, M.I.; Bates, G.J.; Koukourakis, M.I.; Sivridis, E.; Gatter, K.C.; Harris, A.L.; Banham, A.H. The presence of tumor-infiltrating FOXP3+ lymphocytes correlates with intratumoral angiogenesis in endometrial cancer. Gynecol. Oncol. 2008, 110, 216–221. [Google Scholar] [CrossRef]
- Dieci, M.V.; Tsvetkova, V.; Orvieto, E.; Piacentini, F.; Ficarra, G.; Griguolo, G.; Miglietta, F.; Giarratano, T.; Omarini, C.; Bonaguro, S.; et al. Immune characterization of breast cancer metastases: Prognostic implications. Breast Cancer Res. 2018, 20, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y. Modelling the spatial heterogeneity and molecular correlates of lymphocytic infiltration in triple-negative breast cancer. J. R. Soc. Interface 2015, 12, 20141153. [Google Scholar] [CrossRef] [PubMed]
- Zegeye, M.M.; Lindkvist, M.; Fälker, K.; Kumawat, A.K.; Paramel, G.; Grenegård, M.; Sirsjö, A.; Ljungberg, L.U. Activation of the JAK/STAT3 and PI3K/AKT pathways are crucial for IL-6 trans-signaling-mediated pro-inflammatory response in human vascular endothelial cells. Cell Commun. Signal. 2018, 16, 55. [Google Scholar] [CrossRef] [PubMed]
- Hadrup, S.; Donia, M.; Straten, P.T. Effector CD4 and CD8 T Cells and Their Role in the Tumor Microenvironment. Cancer Microenviron. 2012, 6, 123–133. [Google Scholar] [CrossRef]
- Yu, X.; Zhang, Z.; Wang, Z.; Wu, P.; Qiu, F.; Huang, J. Prognostic and predictive value of tumor-infiltrating lymphocytes in breast cancer: A systematic review and meta-analysis. Clin. Transl. Oncol. 2015, 18, 497–506. [Google Scholar] [CrossRef]
- Keir, M.E.; Liang, S.C.; Guleria, I.; Latchman, Y.E.; Qipo, A.; Albacker, L.A.; Koulmanda, M.; Freeman, G.J.; Sayegh, M.H.; Sharpe, A.H. Tissue expression of PD-L1 mediates peripheral T cell tolerance. J. Exp. Med. 2006, 203, 883–895. [Google Scholar] [CrossRef]
- Freeman, G.J.; Long, A.J.; Iwai, Y.; Bourque, K.; Chernova, T.; Nishimura, H.; Fitz, L.J.; Malenkovich, N.; Okazaki, T.; Byrne, M.C.; et al. Engagement of the Pd-1 Immunoinhibitory Receptor by a Novel B7 Family Member Leads to Negative Regulation of Lymphocyte Activation. J. Exp. Med. 2000, 192, 1027–1034. [Google Scholar] [CrossRef]
- Feng, M.; Jiang, W.; Kim, B.Y.; Zhang, C.C.; Fu, Y.-X.; Weissman, I.L. Phagocytosis checkpoints as new targets for cancer immunotherapy. Nat. Rev. Cancer 2019, 19, 568–586. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, X.; Miao, F.; Cao, Y.; Xue, J.; Cao, Q.; Zhang, X. Clinical significance of leukocyte-associated immunoglobulin-like receptor-1 expression in human cervical cancer. Exp. Ther. Med. 2016, 12, 3699–3705. [Google Scholar] [CrossRef] [PubMed]
- Xua, L.; Wang, S.; Li, J.; Li, B. Cancer immunotherapy based on blocking immune suppression mediated by an immune modulator LAIR-1. OncoImmunology 2020, 9, 1–9. [Google Scholar] [CrossRef]
- Lanier, L.L. NK Cell Recognition. Annu. Rev. Immunol. 2005, 23, 225–274. [Google Scholar] [CrossRef] [PubMed]
- Iizasa, T. Overexpression of Collagen XVIII Is Associated with Poor Outcome and Elevated Levels of Circulating Serum Endostatin in Non-Small Cell Lung Cancer. Clin. Cancer Res. 2004, 10, 5361–5366. [Google Scholar] [CrossRef]
- Banyard, J.; Bao, L.; Zetter, B.R. Type XXIII Collagen, a New Transmembrane Collagen Identified in Metastatic Tumor Cells. J. Biol. Chem. 2003, 278, 20989–20994. [Google Scholar] [CrossRef]
- Jingushi, K.; Uemura, M.; Nakano, K.; Hayashi, Y.; Wang, C.; Ishizuya, Y.; Yamamoto, Y.; Hayashi, T.; Kinouchi, T.; Matsuzaki, K.; et al. Leukocyte-associated immunoglobulin-like receptor 1 promotes tumorigenesis in RCC. Oncol. Rep. 2018, 41, 1293–1303. [Google Scholar] [CrossRef]
- Cao, Q.; Fu, A.; Yang, S.; He, X.; Wang, Y.; Zhang, X.; Zhou, J.; Luan, X.; Yu, W.; Xue, J. Leukocyte-associated immunoglobulin-like receptor-1 expressed in epithelial ovarian cancer cells and involved in cell proliferation and invasion. Biochem. Biophys. Res. Commun. 2015, 458, 399–404. [Google Scholar] [CrossRef]
- Triulzi, T.; Forte, L.; Regondi, V.; Di Modica, M.; Ghirelli, C.; Carcangiu, M.L.; Sfondrini, L.; Balsari, A.; Tagliabue, E. HER2 signaling regulates the tumor immune microenvironment and trastuzumab efficacy. OncoImmunology 2019, 8, e1512942. [Google Scholar] [CrossRef]
- Green, A.R.; Aleskandarany, M.A.; Agarwal, D.; Elsheikh, S.; Nolan, C.C.; Diez-Rodriguez, M.; Macmillan, R.D.; Ball, G.R.; Caldas, C.; Madhusudan, S.; et al. MYC functions are specific in biological subtypes of breast cancer and confers resistance to endocrine therapy in luminal tumours. Br. J. Cancer 2016, 114, 917–928. [Google Scholar] [CrossRef]
- Bray, K.; Gillette, M.; Young, J.; Loughran, E.; Hwang, M.; Sears, J.C.; Vargo-Gogola, T. Cdc42 overexpression induces hyperbranching in the developing mammary gland by enhancing cell migration. Breast Cancer Res. 2013, 15, R91. [Google Scholar] [CrossRef]
- Tsiatas, M.; Kalogeras, K.T.; Manousou, K.; Wirtz, R.M.; Gogas, H.; Veltrup, E.; Zagouri, F.; Lazaridis, G.; Koutras, A.; Christodoulou, C.; et al. Evaluation of the prognostic value of CD3, CD8, and FOXP3 mRNA expression in early-stage breast cancer patients treated with anthracycline-based adjuvant chemotherapy. Cancer Med. 2018, 7, 5066–5082. [Google Scholar] [CrossRef] [PubMed]
- Mahmoud, S.M.A.; Paish, E.C.; Powe, D.G.; Macmillan, R.D.; Lee, A.H.S.; Ellis, I.O.; Green, A.R. An evaluation of the clinical significance of FOXP3+ infiltrating cells in human breast cancer. Breast Cancer Res. Treat. 2010, 127, 99–108. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Sun, H.; Zhao, S.; Wang, Y.; Pu, H.; Zhang, Q. Expression of PD-L1 and prognosis in breast cancer: A meta-analysis. Oncotarget 2017, 8, 31347–31354. [Google Scholar] [CrossRef] [PubMed]
- Bolland, S.; Ravetch, J.V. Inhibitory pathways triggered by ITIM-containing receptors. Adv. Immunol. 1999, 72, 149–177. [Google Scholar] [CrossRef]
- Hasebe, T.; Tsuda, H.; Tsubono, Y.; Imoto, S.; Mukai, K. Fibrotic Focus in Invasive Ductal Carcinoma of the Breast: A Histopathological Prognostic Parameter for Tumor Recurrence and Tumor Death within Three Years after the Initial Operation. Jpn. J. Cancer Res. 1997, 88, 590–599. [Google Scholar] [CrossRef]
- Adams, S.; Loi, S.; Toppmeyer, D.; Cescon, D.; De Laurentiis, M.; Nanda, R.; Winer, E.; Mukai, H.; Tamura, K.; Armstrong, A.; et al. Pembrolizumab monotherapy for previously untreated, PD-L1-positive, metastatic triple-negative breast cancer: Cohort B of the phase II KEYNOTE-086 study. Ann. Oncol. 2019, 30, 405–411. [Google Scholar] [CrossRef]
- Adams, S.; Schmid, P.; Rugo, H.; Winer, E.; Loirat, D.; Awada, A.; Cescon, D.; Iwata, H.; Campone, M.; Nanda, R.; et al. Pembrolizumab monotherapy for previously treated metastatic triple-negative breast cancer: Cohort A of the phase II KEYNOTE-086 study. Ann. Oncol. 2019, 30, 397–404. [Google Scholar] [CrossRef]
- Syed, Y.Y.; Syed, Y.Y. Atezolizumab (in Combination with Nab-Paclitaxel): A Review in Advanced Triple-Negative Breast Cancer. Drugs 2020, 80, 601–607. [Google Scholar] [CrossRef]
- Schmid, P.; Chui, S.Y.; Emens, L.A. Atezolizumab and Nab-Paclitaxel in Advanced Triple-Negative Breast Cancer. N. Engl. J. Med. 2019, 380, 985–988. [Google Scholar] [CrossRef]
- Hurd, P.J.; Nelson, C.J. Advantages of next-generation sequencing versus the microarray in epigenetic research. Brief. Funct. Genom. Proteom. 2009, 8, 174–183. [Google Scholar] [CrossRef]
- Wolff, A.; Bayerlová, M.; Gaedcke, J.; Kube, D.; Beißbarth, T. A comparative study of RNA-Seq and microarray data analysis on the two examples of rectal-cancer patients and Burkitt Lymphoma cells. PLoS ONE 2018, 13, e0197162. [Google Scholar] [CrossRef] [PubMed]
- Curtis, C.; Shah, S.P.; Chin, S.-F.; Turashvili, G.; Rueda, O.M.; Dunning, M.J.; Speed, D.; Lynch, A.G.; Samarajiwa, S.A. The genomic and transcriptomic architecture of 2000 breast tumours reveals novel subgroups. Nature 2012, 486, 346–352. [Google Scholar] [CrossRef] [PubMed]
- Jézéquel, P.; Frénel, J.-S.; Campion, L.; Guérin-Charbonnel, C.; Gouraud, W.; Ricolleau, G.; Campone, M. bc-GenExMiner 3.0: New mining module computes breast cancer gene expression correlation analyses. Database 2013, 2013, bas060. [Google Scholar] [CrossRef] [PubMed]
- Cerami, E.; Gao, J.; Dogrusoz, U.; Gross, B.E.; Sumer, S.O.; Aksoy, B.A.; Jacobsen, A.; Byrne, C.J.; Heuer, M.L.; Larsson, E.; et al. The cBio Cancer Genomics Portal: An Open Platform for Exploring Multidimensional Cancer Genomics Data: Figure 1. Cancer Discov. 2012, 2, 401–404. [Google Scholar] [CrossRef]
- Fekete, J.T.; Györffy, B. ROCplot.org: Validating predictive biomarkers of chemotherapy/hormonal therapy/anti-HER2 therapy using transcriptomic data of 3,104 breast cancer patients. Int. J. Cancer 2019, 145, 3140–3151. [Google Scholar] [CrossRef]
- Rakha, E.A.; Boyce, R.W.G.; El-Rehim, D.A.; Kurien, T.; Green, A.R.; Paish, E.C.; Robertson, J.F.R.; Ellis, I.O. Expression of mucins (MUC1, MUC2, MUC3, MUC4, MUC5AC and MUC6) and their prognostic significance in human breast cancer. Mod. Pathol. 2005, 18, 1295–1304. [Google Scholar] [CrossRef]
- El Ansari, R.; Craze, M.L.; Miligy, I.; Diez-Rodriguez, M.; Nolan, C.C.; Ellis, I.O.; Rakha, E.A.; Green, A.R. The amino acid transporter SLC7A5 confers a poor prognosis in the highly proliferative breast cancer subtypes and is a key therapeutic target in luminal B tumours. Breast Cancer Res. 2018, 20, 1–17. [Google Scholar] [CrossRef]
- Rakha, E.A.; Pinder, S.E.; Bartlett, J.M.S.; Ibrahim, M.; Starczynski, J.; Carder, P.J.; Provenzano, E.; Hanby, A.; Hales, S.; Lee, A.H.S.; et al. Updated UK Recommendations for HER2 assessment in breast cancer. J. Clin. Pathol. 2014, 68, 93–99. [Google Scholar] [CrossRef]
- Rakha, E.A.; Agarwal, D.; Green, A.R.; Ashankyty, I.; Ellis, I.O.; Ball, G.; Aleskandarany, M.A. Prognostic stratification of oestrogen receptor-positive HER2-negative lymph node-negative class of breast cancer. Histopathology 2016, 70, 622–631. [Google Scholar] [CrossRef]
- Fragomeni, S.M.; Sciallis, A.; Jeruss, J.S. Molecular Subtypes and Local-Regional Control of Breast Cancer. Surg. Oncol. Clin. N. Am. 2018, 27, 95–120. [Google Scholar] [CrossRef]
- Joseph, C.; Arshad, M.; Kurozomi, S.; Althobiti, M.; Miligy, I.; Al-Izzi, S.; Toss, M.S.; Goh, F.Q.; Johnston, S.J.; Martin, S.G.; et al. Overexpression of the cancer stem cell marker CD133 confers a poor prognosis in invasive breast cancer. Breast Cancer Res. Treat. 2019, 174, 387–399. [Google Scholar] [CrossRef] [PubMed]
- McCarty, K.S., Jr.; Miller, L.S.; Cox, E.B.; Konrath, J.; McCarty, K.S., Sr. Estrogen receptor analyses. Correlation of biochemical and immunohistochemical methods using monoclonal antireceptor antibodies. Arch. Pathol. Lab. Med. 1985, 109, 716–721. [Google Scholar] [PubMed]
- Althobiti, M.; Aleskandarany, M.A.; Joseph, C.; Toss, M.; Mongan, N.; Diez-Rodriguez, M.; Nolan, C.C.; Ashankyty, I.; Ellis, I.O.; Green, A.R.; et al. Heterogeneity of tumour-infiltrating lymphocytes in breast cancer and its prognostic significance. Histopathology 2018, 73, 887–896. [Google Scholar] [CrossRef] [PubMed]
- Mahmoud, S.; Lee, A.; Ellis, I.; Green, A. CD8+T lymphocytes infiltrating breast cancer. OncoImmunology 2012, 1, 364–365. [Google Scholar] [CrossRef]
- Mahmoud, S.M.A.; Lee, A.H.; Paish, E.C.; Macmillan, R.D.; Ellis, I.O.; Green, A.R. Tumour-infiltrating macrophages and clinical outcome in breast cancer. J. Clin. Pathol. 2011, 65, 159–163. [Google Scholar] [CrossRef]
- Habashy, H.O.; Powe, D.G.; Staka, C.M.; Rakha, E.A.; Ball, G.; Green, A.R.; Aleskandarany, M.; Paish, E.C.; Macmillan, R.D.; Nicholson, R.I.; et al. Transferrin receptor (CD71) is a marker of poor prognosis in breast cancer and can predict response to tamoxifen. Breast Cancer Res. Treat. 2009, 119, 283–293. [Google Scholar] [CrossRef]
- Chrysanthou, E.; Gorringe, K.L.; Joseph, C.; Craze, M.; Nolan, C.C.; Diez-Rodriguez, M.; Green, A.R.; Rakha, E.A.; Ellis, I.O.; Mukherjee, A. Phenotypic characterisation of breast cancer: The role of CDC42. Breast Cancer Res. Treat. 2017, 164, 317–325. [Google Scholar] [CrossRef]
- Aleskandarany, M.A.; Rakha, E.A.; Macmillan, R.D.; Powe, D.G.; Ellis, I.O.; Green, A.R. MIB1/Ki-67 labelling index can classify grade 2 breast cancer into two clinically distinct subgroups. Breast Cancer Res. Treat. 2010, 127, 591–599. [Google Scholar] [CrossRef]
- El-Rehim, D.M.A.; Pinder, S.E.; Paish, C.E.; Bell, J.A.; Rampaul, R.S.; Blamey, R.W.; Robertson, J.F.R.; Nicholson, R.I.; Ellis, I.O. Expression and co-expression of the members of the epidermal growth factor receptor (EGFR) family in invasive breast carcinoma. Br. J. Cancer 2004, 91, 1532–1542. [Google Scholar] [CrossRef]
- Aleskandarany, M.A.; Rakha, E.A.; Ahmed, M.A.H.; Powe, D.G.; Paish, E.C.; Macmillan, R.D.; Ellis, I.O.; Green, A.R. PIK3CA expression in invasive breast cancer: A biomarker of poor prognosis. Breast Cancer Res. Treat. 2009, 122, 45–53. [Google Scholar] [CrossRef]
- Yu, G.; Wang, L.-G.; Han, Y.; He, Q.-Y. clusterProfiler: An R Package for Comparing Biological Themes Among Gene Clusters. OMICS A J. Integr. Biol. 2012, 16, 284–287. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clinicopathological Criteria | LAIR-1 mRNA Expression N (%) | χ2 (p Value) | |
|---|---|---|---|
| Negative/Low Expression | High Expression | ||
| Age at Diagnosis | |||
| ≤50 | 234 (55.0) | 190 (45.0) | 0.091 (0.764) |
| >50 | 846 (54.0) | 710 (46.0) | |
| Tumour Size (cm) | |||
| ≤2.0 | 488 (57.0) | 370 (43.0) | 3.280 (0.070) |
| >2.0 | 581 (53.0) | 520 (47.0) | |
| Tumour Stage | |||
| I | 290 (58.0) | 211 (42.0) | 17.033 (0.001) |
| II | 417 (51.0) | 408 (49.0) | |
| III | 53 (45.0) | 65 (55.0) | |
| Histological Grade | |||
| Grade 1 | 112 (66.0) | 58 (34.0) | 56.994 (<0.001) |
| Grade 2 | 479 (62.0) | 291 (38.0) | |
| Grade 3 | 435 (46.0) | 517 (54.0) | |
| Tumour Type | |||
| Ductal | 926 (54.0) | 775 (46.0) | 11.279 (0.024) |
| Lobular | 92 (62.0) | 55 (38.0) | |
| Other special types | 29 (60.0) | 20 (40.0) | |
| Mixed tumours | 12 (57.0) | 9 (43.0) | |
| Nottingham Prognostic Index groups | |||
| Good | 438 (64.0) | 242 (36.0) | 50.501 (<0.001) |
| Moderate | 564 (51.0) | 537 (49.0) | |
| Poor | 78 (39.0) | 121 (61.0) | |
| PAM50 Subtype | |||
| Luminal A | 478 (67.0) | 240 (33.0) | 81.734 (<0.001) |
| Luminal B | 254 (52.0) | 234 (48.0) | |
| Basal | 127 (39.0 | 202 (61.0) | |
| Her2 enriched | 116 (48.0) | 124 (52.0) | |
| Normal like | 101 (51.0) | 98 (49.0) | |
| IntClustMemb | |||
| IntClustMemb 1 | 71 (51.0) | 68 (49.0) | 126.426 (<0.001) |
| IntClustMemb 2 | 33 (46.0) | 39 (54.0) | |
| IntClustMemb 3 | 178 (61.0) | 112 (39.0) | |
| IntClustMemb 4 | 145 (42.0) | 198 (58.0) | |
| IntClustMemb 5 | 94 (49.0) | 96 (51.0) | |
| IntClustMemb 6 | 52 (61.0) | 33 (39.0) | |
| IntClustMemb 7 | 130 (68.0) | 60 (32.0) | |
| IntClustMemb 8 | 221 (74.0) | 78 (26.0) | |
| IntClustMemb 9 | 76 (52.0) | 70 (48.0) | |
| IntClustMemb 10 | 80 (35.0) | 146 (65.0) | |
| Oestrogen Receptor (ER) | |||
| Negative | 189 (43.0) | 250 (57.0) | 30.649 (<0.001) |
| Positive | 869 (58.0) | 629 (42.0) | |
| Progesterone Receptor (PR) | |||
| Negative | 432 (45.0) | 517 (55.0) | 65.768 (<0.001) |
| Positive | 657 (63.0) | 383 (37.0) | |
| Human Epidermal Growth Factor Receptor 2(HER2) | |||
| Negative | 963 (56.0) | 770 (44.0) | 5.863 (0.015) |
| Positive | 117 (48.0) | 130 (52.0) | |
| Parameters | LAIR-1 Cytoplasmic Expression | ||
|---|---|---|---|
| Negative/Low Expression N (%) | High Expression N (%) | p Value (χ2) | |
| Age at Diagnosis (years) | |||
| <50 | 101 (49.0) | 105 (51.0) | 0.898 (0.016) |
| ≥50 | 180 (49.0) | 183 (51.0) | |
| Histological Grade | |||
| 1 | 41 (54.7) | 34 (45.3) | <0.0001 (23.004) |
| 2 | 115 (62.2) | 70 (37.8) | |
| 3 | 122 (40.3) | 181 (59.7) | |
| Stage | |||
| I | 175 (51.0) | 167 (49.0) | 0.763 (1.158) |
| II | 84 (47.0) | 95 (53.0) | |
| III | 19 (45.0) | 23 (55.0) | |
| Tumour Size | |||
| ˂2.0 cm | 125 (48.0) | 133 (52.0) | 0.657 (0.197) |
| ≥2.0 cm | 155 (50.0) | 153 (50.0) | |
| Histological Type | |||
| Ductal (including mixed) | 225 (47.0) | 256 (53.0) | <0.001 (25.715) |
| Lobular | 44 (79.0) | 12 (21.0) | |
| Special type | 10 (50.0) | 10 (50.0) | |
| IHC Subtypes | |||
| ER+/HER2-Low Proliferation | 69 (61.6) | 43 (38.4) | 0.019 (0.065) |
| ER+/HER2- High Proliferation | 86 (49.4) | 88 (50.6) | |
| Triple Negative | 46 (42.2) | 63 (57.8) | |
| HER2+ | 40 (43.0) | 53 (57.0) | |
| Nottingham Prognostic Index | |||
| GPG | 88 (60.0) | 59 (40.0) | 0.012 (8.866) |
| MPG | 153 (47.0) | 176 (53.0) | |
| PPG | 39 (43.0) | 51 (57.0) | |
| Oestrogen (ER) Status | |||
| Negative | 64 (41.0) | 92 (59.0) | 0.013 (6.143) |
| Positive | 216 (52.0) | 194 (48.0) | |
| Progesterone (PR) Status | |||
| Negative | 112(47.0) | 126 (53.0) | 0.345 (0.891) |
| Positive | 161 (51.0) | 154 (49.0) | |
| Human Epidermal Growth Factor Receptor 2 (HER2) | |||
| Negative | 233 (50.0) | 227 (50.0) | 0.091 (1.113) |
| Positive | 42 (45.0) | 52 (56.0) | |
| Epidermal Growth Factor Receptor (EGFR) (>10% as Positive Expression) | |||
| Negative | 225 (51.0) | 213 (49.0) | 0.058 (3.604) |
| Positive | 49 (42.0) | 69 (58.0) | |
| Phosphatidylinositol-4,5-Bisphosphate 3-Kinase, Catalytic Subunit Alpha (PIK3CA) (H-score > 100 as Positive Expression) | |||
| Negative | 58 (57.0) | 44 (43.0) | 0.048 (3.837) |
| Positive | 160 (46.0) | 189 (54.0) | |
| Myc Proto-Oncogene Protein (c-MYC) (H-score > 35 as Positive Expression) | |||
| Negative | 202 (55.0) | 166 (45.0) | <0.001 (27.38) |
| Positive | 28 (26.0) | 79 (74.0) | |
| Cell Division Cycle 42 (Cdc42)(H-score > 150 as Positive Expression) | |||
| Negative | 130 (57.0) | 98 (43.0) | <0.001 (16.24) |
| Positive | 58 (36.0) | 102 (64.0) | |
| Ki67 (>10% as Positive Expression) | |||
| Negative | 91 (57.0) | 69 (43.0) | 0.080 (3.071) |
| Positive | 135 (48.0) | 145 (52.0) | |
| Cyclin B1 (>1% as Positive Expression) | |||
| Negative | 113 (54.0) | 97 (46.0) | 0.010 (6.630) |
| Positive | 57 (40.0) | 86 (60.0) | |
| Variable | Breast Cancer-Specific Survival | Distant Metastasis-Free Interval | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Univariate | Multivariate | Univariate | Multivariate | |||||||||
| HR | 95% CI | p Value | HR | 95% CI | p Value | HR | 95% CI | p Value | HR | 95% CI | p Value | |
| Whole Cohort | ||||||||||||
| Tumour size | 2.6 | 2.0–3.4 | <0.0001 | 3.0 | 1.6–5.7 | 0.0004 | 2.3 | 1.8–2.8 | <0.0001 | 2.3 | 1.4–3.9 | 0.002 |
| Tumour stage | 2.5 | 2.6–2.9 | <0.0001 | 2.4 | 1.7–3.5 | <0.0001 | 2.6 | 2.3–2.9 | <0.0001 | 2.2 | 1.6–3.0 | <0.0001 |
| Grade | 4.2 | 3.2–5.5 | <0.0001 | 7.0 | 2.2–20.8 | 0.001 | 3.0 | 2.5–3.5 | <0.0001 | 2.8 | 1.6–5.1 | <0.001 |
| CD8 | 0.7 | 0.5–1.0 | 0.084 | 0.5 | 0.3–1.1 | 0.019 | 0.9 | 0.7–1.2 | 0.470 | 0.9 | 0.4–1.8 | 0.673 |
| CD3 | 1.0 | 0.7–1.6 | 0.851 | 1.1 | 0.5–3.0 | 0.7807 | 0.9 | 0.6–1.3 | 0.452 | 0.7 | 0.4–1.4 | 0.297 |
| Her2+ | 3.1 | 2.4–4.1 | <0.0001 | 2.1 | 1.2–3.5 | 0.008 | 2.7 | 2.1–3.5 | <0.0001 | 1.8 | 1.1–3.1 | 0.009 |
| Chemotherapy | 2.1 | 1.6–2.8 | <0.0001 | 0.9 | 0.5–1.7 | 0.880 | 2.0 | 1.6–2.6 | <0.0001 | 0.7 | 0.4–1.2 | 0.172 |
| LAIR-1 | 2.0 | 1.3–3.0 | 0.002 | 2.0 | 1.1–3.4 | 0.017 | 1.6 | 1.1–2.3 | 0.013 | 1.6 | 1.0–2.5 | 0.046 |
| ** LUMINAL A | ||||||||||||
| Tumour size | 3.5 | 1.3–9.4 | 0.013 | 1.8 | 0.1–22.0 | 0.647 | 4.2 | 2.0–9.3 | 0.0003 | 7.3 | 0.7–17.0 | 0.088 |
| Tumour stage | 3.6 | 1.9–6.7 | <0.001 | 3.4 | 0.3–33.0 | 0.292 | 3.0 | 1.8–4.8 | 0.0001 | 0.3 | 0.1–3.2 | 0.338 |
| Grade | 3.0 | 1.5–6.2 | 0.003 | 5.7 | 0.5–16.0 | 0.149 | 3.0 | 1.7–5.2 | 0.0001 | 1.1 | 0.3–3.6 | 0.890 |
| Chemotherapy | 1.8 | 0.2–13.6 | 0.580 | 2.9 | 0.2–41.0 | 0.414 | 2.3 | 0.6–10.0 | 0.250 | 2.2 | 0.2–28.0 | 0.557 |
| LAIR-1 | 3.1 | 0.3–34.0 | 0.346 | 2.1 | 0.1–46.0 | 0.638 | 2.7 | 0.7–11.0 | 0.166 | 5.0 | 0.6–40.0 | 0.135 |
| LUMINAL B | ||||||||||||
| Tumour size | 1.8 | 1.1–2.9 | 0.013 | 1.7 | 0.6–5.2 | 0.337 | 2.1 | 1.5–3.2 | <0.001 | 2.3 | 0.9–6.9 | 0.055 |
| Tumour stage | 2.5 | 1.8–3.3 | <0.0001 | 1.2 | 0.4–3.1 | 0.788 | 2.3 | 1.8–3.0 | <0.0001 | 1.0 | 0.4–2.3 | 0.989 |
| Grade | 3.1 | 1.9–5.2 | <0.0001 | 10.1 | 1.3–75 | 0.025 | 2.2 | 1.6–3.1 | 0.0001 | 2.3 | 0.9–5.4 | 0.066 |
| CD8 | 0.7 | 0.4–1.3 | 0.268 | 1.3 | 0.3–5.1 | 0.756 | 0.8 | 0.5–1.3 | 0.401 | 1.1 | 0.3–3.7 | 0.873 |
| CD3 | 0.9 | 0.4–2.1 | 0.923 | 1.3 | 0.1–12 | 0.834 | 0.8 | 0.4–1.4 | 0.450 | 1.2 | 0.3–6.0 | 0.789 |
| Chemotherapy | 1.2 | 0.6–2.2 | 0.658 | 0.6 | 0.1–2.3 | 0.531 | 1.1 | 0.7–2.0 | 0.666 | 0.4 | 0.1–2.0 | 0.264 |
| LAIR-1 | 1.7 | 0.7–4.1 | 0.247 | 1.7 | 0.6–5.5 | 0.331 | 1.1 | 0.5–2.1 | 0.859 | 1.1 | 0.4–2.8 | 0.770 |
| HER2+ | ||||||||||||
| Tumour size | 2.8 | 1.7–4.8 | 0.0001 | 5.0 | 1.3–18.4 | 0.016 | 2.0 | 1.3–3.1 | 0.003 | 1.8 | 0.7–5.1 | 0.245 |
| Tumour stage | 2.5 | 1.9–3.4 | <0.0001 | 7.3 | 3.0–18.0 | <0.0001 | 2.4 | 1.8–3.2 | <0.0001 | 7.0 | 3.2–15.0 | <0.0001 |
| Grade | 2.0 | 0.9–4.1 | 0.048 | 3.3 | 0.4–27.0 | 0.261 | 1.5 | 0.9–2.6 | 0.167 | 0.6 | 0.1–1.9 | 0.311 |
| CD8 | 0.6 | 0.4–1.2 | 0.143 | 0.1 | 0.1–0.5 | 0.002 | 0.6 | 0.4–1.1 | 0.126 | 0.4 | 0.2–0.6 | 0.113 |
| CD3 | 0.9 | 0.3–2.7 | 0.944 | 2.2 | 0.3–15.0 | 0.405 | 0.7 | 0.3–1.6 | 0.427 | 2.4 | 0.4–16.3 | 0.369 |
| Chemotherapy | 1.5 | 0.9–2.5 | 0.144 | 2.6 | 0.9–7.0 | 0.070 | 1.3 | 0.8–2.2 | 0.232 | 0.9 | 0.4–2.3 | 0.866 |
| LAIR-1 | 2.5 | 1.1–5.3 | 0.014 | 4.8 | 1.8–13.0 | 0.003 | 2.0 | 1.0–4.1 | 0.044 | 5.8 | 1.9–17.0 | 0.002 |
| TNBC | ||||||||||||
| Tumour size | 1.3 | 0.8–2.1 | 0.308 | 2.8 | 0.9–9.0 | 0.086 | 1.1 | 0.7–1.8 | 0.553 | 1.6 | 0.5–4.5 | 0.364 |
| Tumour stage | 2.0 | 1.5–2.7 | <0.0001 | 3.2 | 1.6–6.5 | 0.001 | 2.0 | 1.5–2.6 | <0.0001 | 2.8 | 1.4–5.2 | 0.002 |
| CD8 | 0.5 | 0.2–0.8 | 0.011 | 0.3 | 0.1–1.3 | 0.194 | 0.5 | 0.3–1.0 | 0.061 | 0.4 | 0.1–1.7 | 0.222 |
| CD3 | 0.5 | 0.2–0.10 | 0.050 | 0.7 | 0.1–5.6 | 0.764 | 0.5 | 0.2–1.1 | 0.077 | 1.3 | 0.2–9.2 | 0.815 |
| Chemotherapy | 0.9 | 0.6–1.5 | 0.741 | 0.5 | 0.2–1.3 | 0.145 | 0.9 | 0.5–1.3 | 0.431 | 0.5 | 0.1–1.3 | 0.163 |
| LAIR-1 | 1.3 | 0.6–2.8 | 0.476 | 1.1 | 0.4–3.1 | 0.872 | 1.4 | 0.7–2.9 | 0.322 | 1.0 | 0.3–2.7 | 0.982 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Joseph, C.; Alsaleem, M.A.; Toss, M.S.; Kariri, Y.A.; Althobiti, M.; Alsaeed, S.; Aljohani, A.I.; Narasimha, P.L.; Mongan, N.P.; Green, A.R.; et al. The ITIM-Containing Receptor: Leukocyte-Associated Immunoglobulin-Like Receptor-1 (LAIR-1) Modulates Immune Response and Confers Poor Prognosis in Invasive Breast Carcinoma. Cancers 2021, 13, 80. https://doi.org/10.3390/cancers13010080
Joseph C, Alsaleem MA, Toss MS, Kariri YA, Althobiti M, Alsaeed S, Aljohani AI, Narasimha PL, Mongan NP, Green AR, et al. The ITIM-Containing Receptor: Leukocyte-Associated Immunoglobulin-Like Receptor-1 (LAIR-1) Modulates Immune Response and Confers Poor Prognosis in Invasive Breast Carcinoma. Cancers. 2021; 13(1):80. https://doi.org/10.3390/cancers13010080
Chicago/Turabian StyleJoseph, Chitra, Mansour A. Alsaleem, Michael S. Toss, Yousif A. Kariri, Maryam Althobiti, Sami Alsaeed, Abrar I. Aljohani, Pavan L. Narasimha, Nigel P. Mongan, Andrew R. Green, and et al. 2021. "The ITIM-Containing Receptor: Leukocyte-Associated Immunoglobulin-Like Receptor-1 (LAIR-1) Modulates Immune Response and Confers Poor Prognosis in Invasive Breast Carcinoma" Cancers 13, no. 1: 80. https://doi.org/10.3390/cancers13010080
APA StyleJoseph, C., Alsaleem, M. A., Toss, M. S., Kariri, Y. A., Althobiti, M., Alsaeed, S., Aljohani, A. I., Narasimha, P. L., Mongan, N. P., Green, A. R., & Rakha, E. A. (2021). The ITIM-Containing Receptor: Leukocyte-Associated Immunoglobulin-Like Receptor-1 (LAIR-1) Modulates Immune Response and Confers Poor Prognosis in Invasive Breast Carcinoma. Cancers, 13(1), 80. https://doi.org/10.3390/cancers13010080