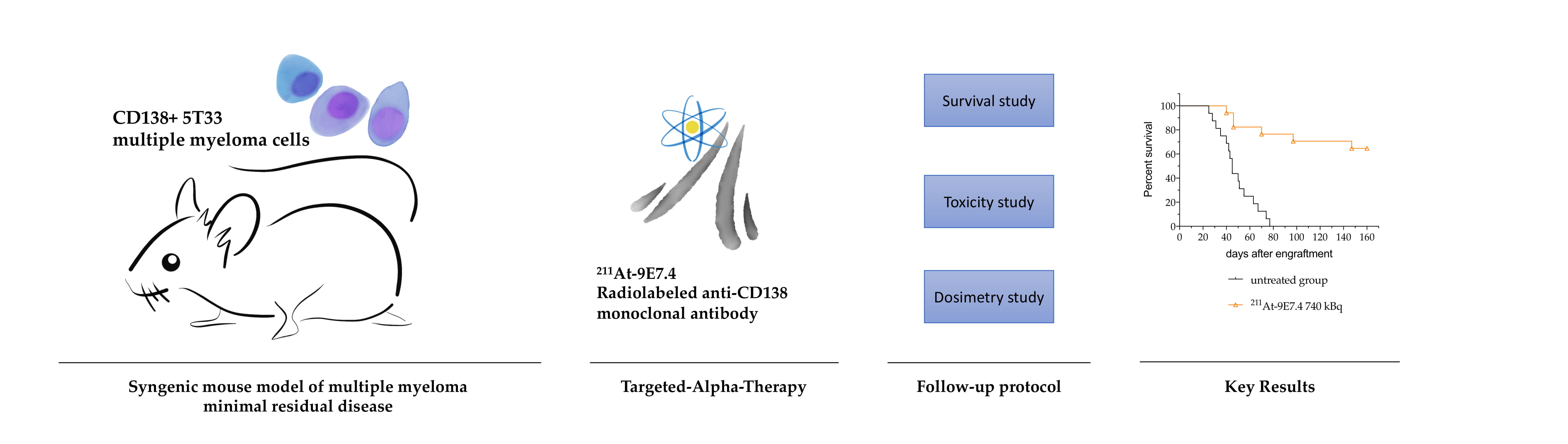
Targeted-Alpha-Therapy Combining Astatine-211 and anti-CD138 Antibody in a Preclinical Syngeneic Mouse Model of Multiple Myeloma Minimal Residual Disease

 , , , , , , ,
, , , , , , ,
Simple Summary
Abstract

1. Introduction
2. Results
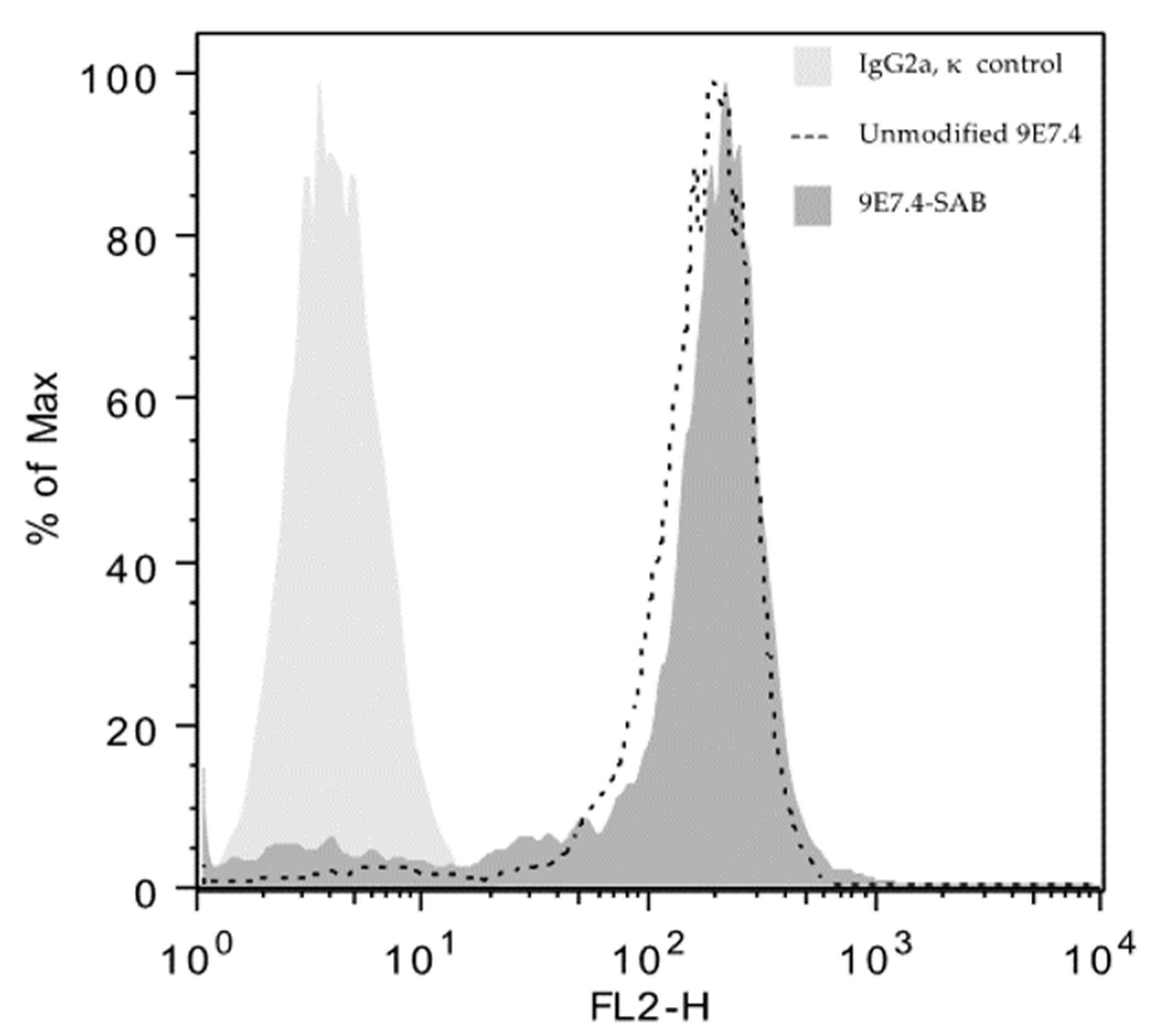
2.1. Anti-CD138 Antibody, 211At Radiolabeling, Immunoreactivity and Flow Cytometry
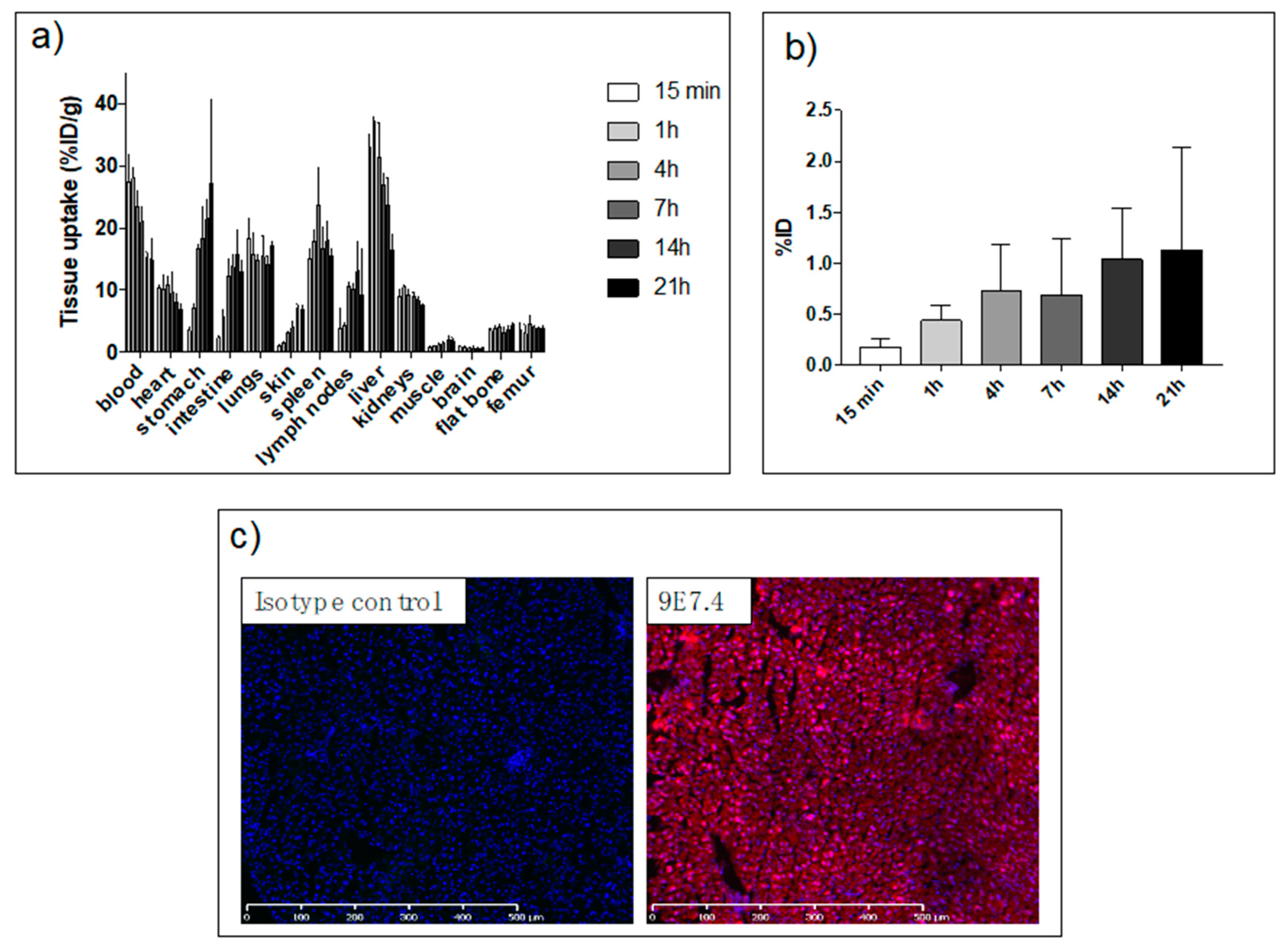
2.2. Biodistribution Study
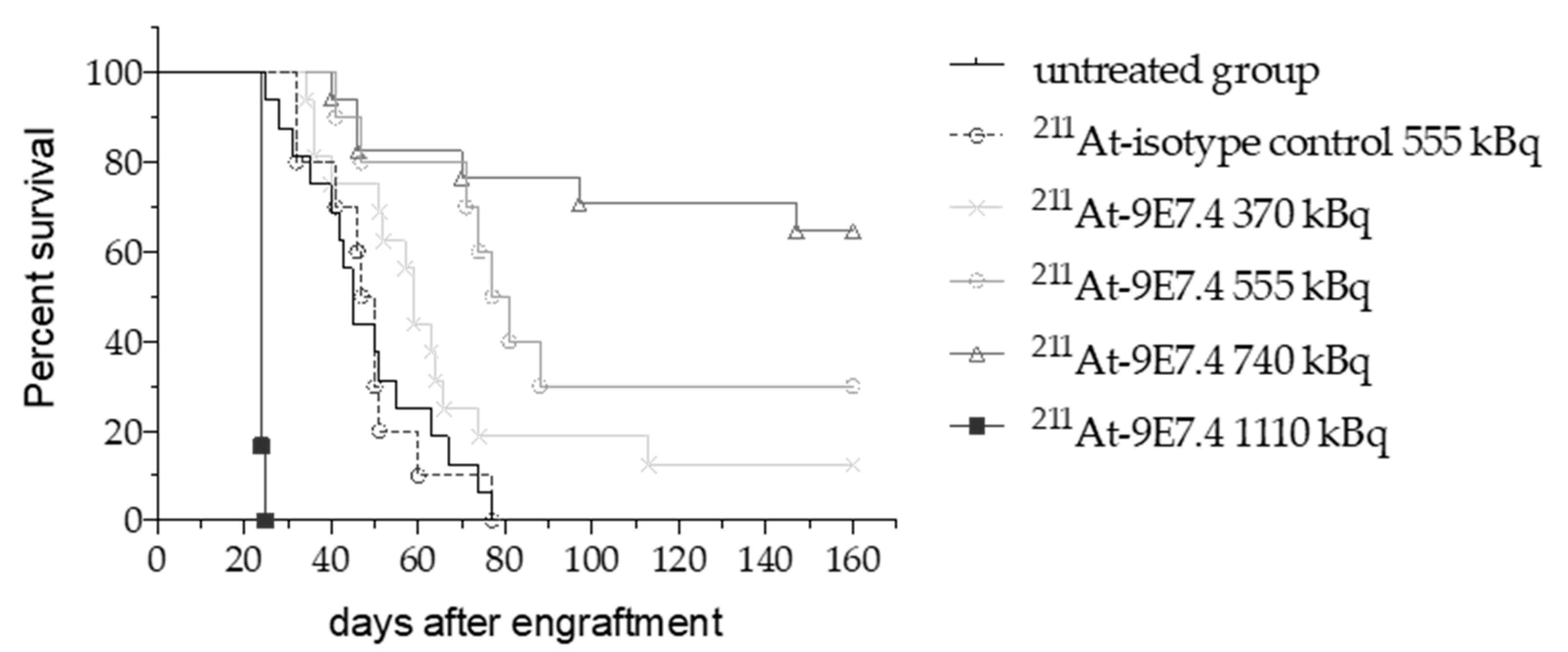
2.3. 211At-anti-mCD138 TAT in a Disseminated Murine MM
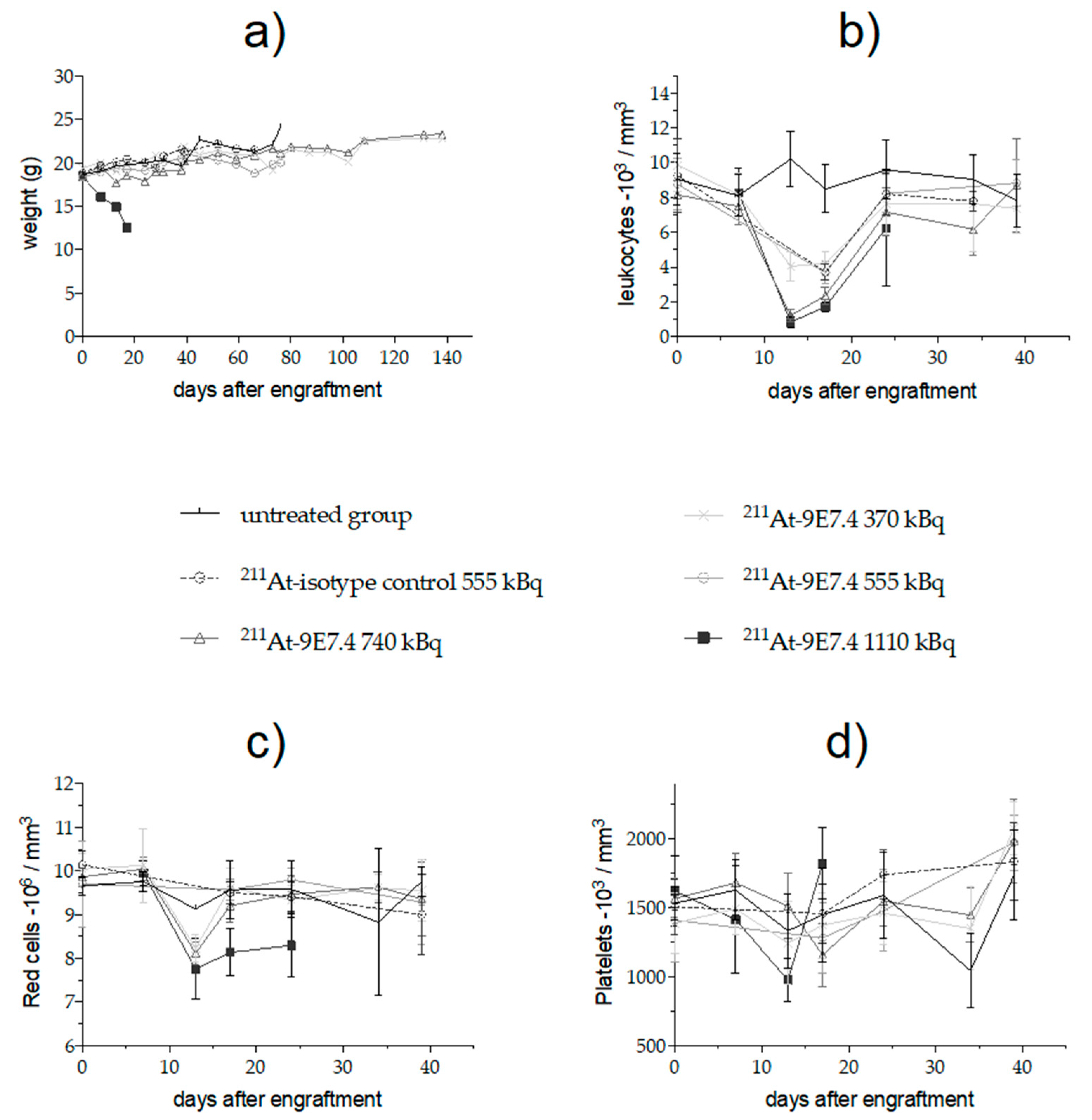
2.4. Monitoring of Early Toxicity of the Treatment
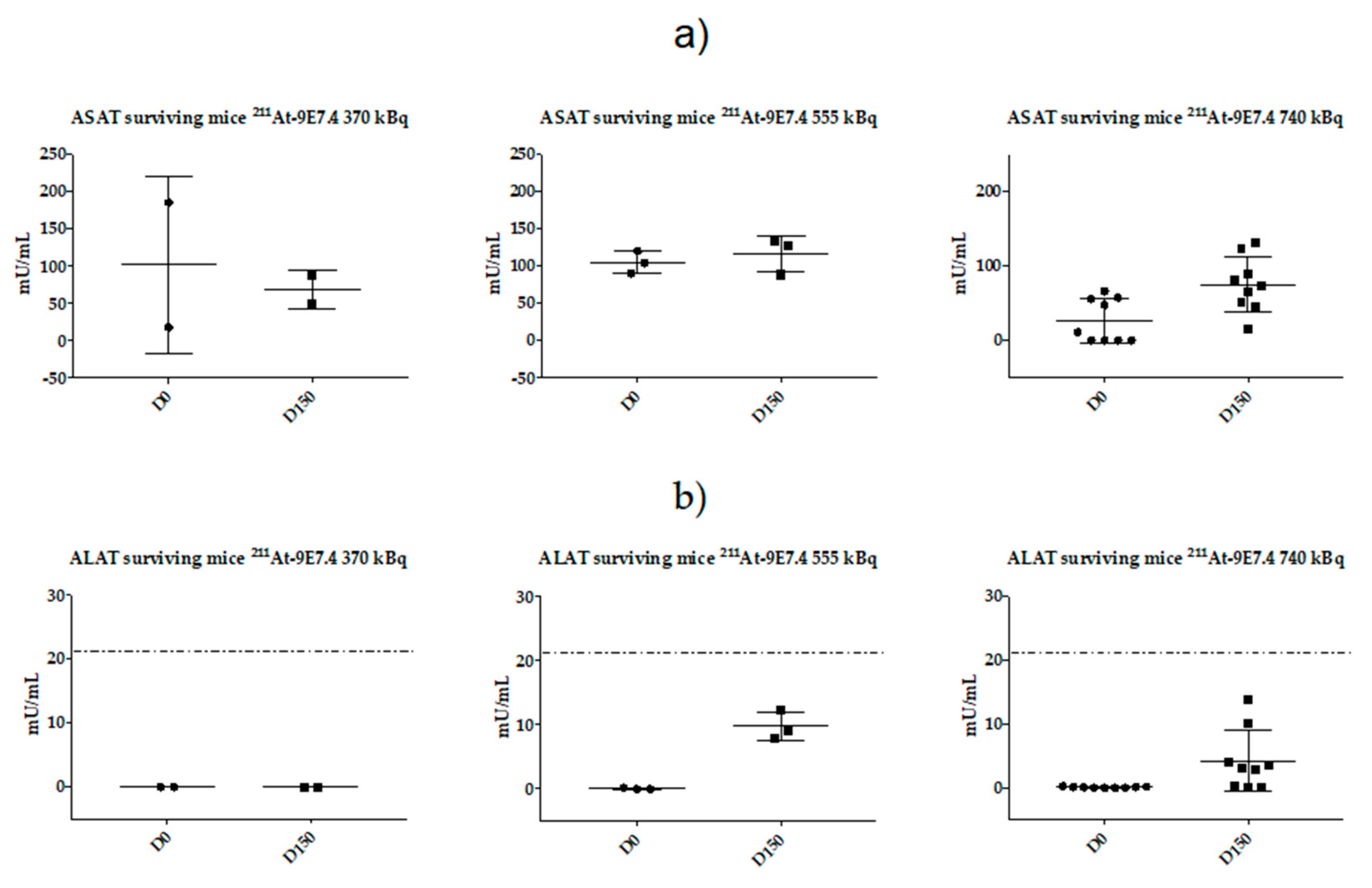
2.5. Monitoring of Late Toxicity of the Treatment by ASAT (Aspartate Aminotransferase) and ALAT (Alanine Aminotransferase) Enzymatic Assay
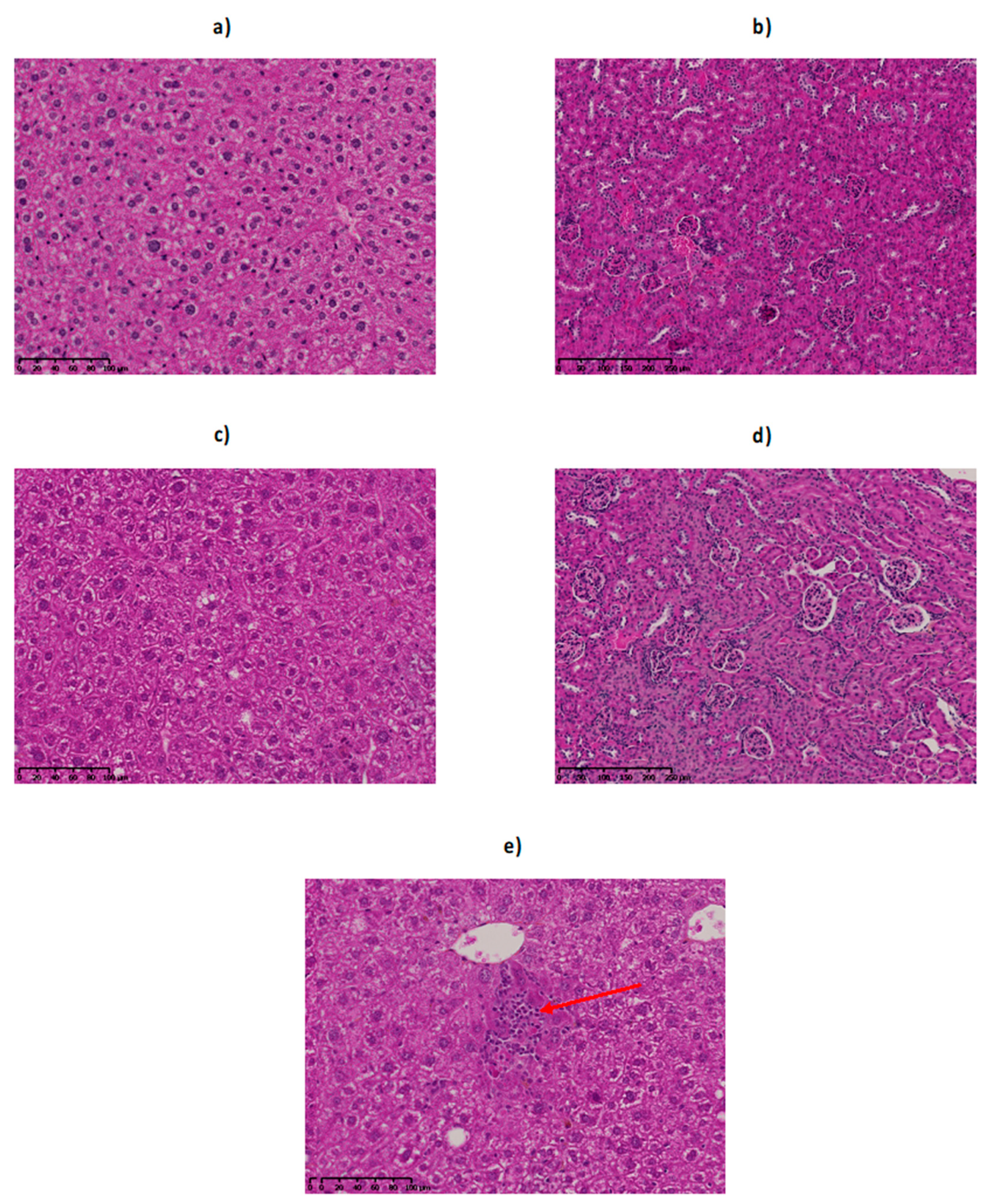
2.6. Monitoring of Late Toxicity of the Treatment by Histology
2.6.1. Histological Examination of the Liver
2.6.2. Histological Examination of the Kidney
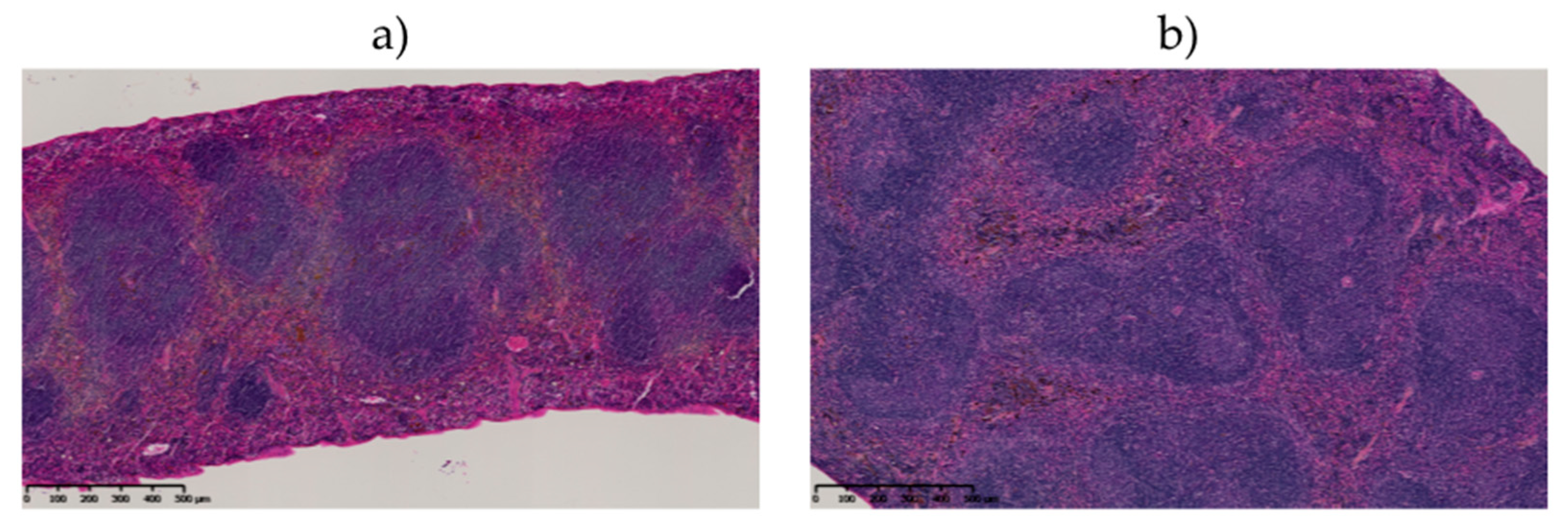
2.6.3. Histological Examination of the Spleen
2.7. Dosimetry Study
3. Discussion
4. Materials and Methods
4.1. Mice
4.2. 5T33 Cell Line
4.3. Antibodies
4.4. Flow Cytometry
4.5. 211At Antibody Radiolabeling
4.6. Immunoreactivity Assay
4.7. 211At-9E7.4 Biodistribution
4.8. Immunofluorescence
4.9. 211At-9E7.4 TAT Study
4.10. Serum Toxicity Assessments
4.11. Histological Examination of Mouse Organs
4.12. Quantification of Mouse Myeloma IgG2b
4.13. Dosimetry
4.14. Statistical Considerations
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Siegel, R.L.; Mph, K.D.M.; Jemal, A. Cancer statistics, 2016. CA A Cancer J. Clin. 2016, 66, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Rajkumar, S.V.; Rajkumar, V.; Kyle, R.A.; Van Duin, M.; Sonneveld, P.; Mateos, M.-V.; Gay, F.; Anderson, K.C. Multiple myeloma. Nat. Rev. Dis. Prim. 2017, 3, 17046. [Google Scholar] [CrossRef] [PubMed]
- Kazandjian, D. Multiple myeloma epidemiology and survival: A unique malignancy. Semin. Oncol. 2016, 43, 676–681. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Zhang, Y.; Chen, R.; Tian, Z.; Zhai, Y.; Janz, S.; Gu, C.; Yang, Y. Chromosomal instability and acquired drug resistance in multiple myeloma. Oncotarget 2017, 8, 78234–78244. [Google Scholar] [CrossRef]
- Moreau, P. How I treat myeloma with new agents. Blood 2017, 130, 1507–1513. [Google Scholar] [CrossRef]
- Nooka, A.K.; Joseph, N.S.; Kaufman, J.L.; Heffner, L.T.; Gupta, V.A.; Gleason, C.; Boise, L.H.; Lonial, S. Clinical efficacy of daratumumab, pomalidomide, and dexamethasone in patients with relapsed or refractory myeloma: Utility of re-treatment with daratumumab among refractory patients. Cancer 2019, 125, 2991–3000. [Google Scholar] [CrossRef]
- Wijdenes, J.; Vooijs, W.C.; Clément, C.; Post, J.; Morard, F.; Vita, N.; Laurent, P.; Sun, R.-Y.; Klein, B.; Dore, J.-M. A plasmocyte selective monoclonal antibody (B-B4) recognizes syndecan-1. Br. J. Haematol. 1996, 94, 318–323. [Google Scholar] [CrossRef]
- Supiot, S.; Faivre-Chauvet, A.; Couturier, O.; Robillard, N.; Morandeau, L. Comparison of the biologic effects of MA5 and B-B4 monoclonal antibody labeled with iodine-131 and bismuth-213 on multiple myeloma. Cancer 2002, 94, 1202–1209. [Google Scholar] [CrossRef]
- Bayer-Garner, I.B.; Sanderson, R.D.; Dhodapkar, M.V.; Owens, R.B.; Wilson, C.S. Syndecan-1 (CD138) Immunoreactivity in Bone Marrow Biopsies of Multiple Myeloma: Shed Syndecan-1 Accumulates in Fibrotic Regions. Mod. Pathol. 2001, 14, 1052–1058. [Google Scholar] [CrossRef]
- Gharbaran, R. Advances in the molecular functions of syndecan-1 (SDC1/CD138) in the pathogenesis of malignancies. Crit. Rev. Oncol. 2015, 94, 1–17. [Google Scholar] [CrossRef]
- Pouget, J.-P.; Mather, S.J. General aspects of the cellular response to low- and high-LET radiation. Eur. J. Nucl. Med. Mol. Imaging 2001, 28, 541–561. [Google Scholar] [CrossRef] [PubMed]
- Kennel, S.J.; Stabin, M.; Roeske, J.C.; Foote, L.J.; Lankford, P.K.; Terzaghi-Howe, M.; Patterson, H.; Barkenbus, J.; Popp, D.M.; Boll, R.; et al. Radiotoxicity of bismuth-213 bound to membranes of monolayer and spheroid cultures of tumor cells. Radiat. Res. 1999, 151, 244–256. [Google Scholar] [CrossRef] [PubMed]
- Ballangrud, Å.M.; Yang, W.H.; Charlton, D.E.; McDevitt, M.R.; Hamacher, K.A.; Panageas, K.S.; Ma, D.; Bander, N.H.; Scheinberg, D.A.; Sgouros, G. Response of LNCaP spheroids after treatment with an alpha-particle emitter (213Bi)-labeled anti-prostate-specific membrane antigen antibody (J591). Cancer Res. 2001, 61, 2008–2014. [Google Scholar] [PubMed]
- Lindegren, S.; Albertsson, P.; Bäck, T.; Jensen, H.; Palm, S.; Aneheim, E. Realizing Clinical Trials with Astatine-211: The Chemistry Infrastructure. Cancer Biother. Radiopharm. 2020. [Google Scholar] [CrossRef]
- Zalutsky, M.R.; Reardon, D.A.; Akabani, G.; Coleman, R.E.; Friedman, A.H.; Friedman, H.S.; McLendon, R.E.; Wong, T.Z.; Bigner, D.D. Clinical Experience with α-Particle–Emitting 211At: Treatment of Recurrent Brain Tumor Patients with 211At-Labeled Chimeric Antitenascin Monoclonal Antibody 81C6. J. Nucl. Med. 2007, 49, 30–38. [Google Scholar] [CrossRef]
- Andersson, H.; Cederkrantz, E.; Divgi, C.; Elgqvist, J.; Himmelman, J.; Horvath, G.; Jacobsson, L.; Jensen, H.; Lindegren, S.; Palm, S.; et al. Intraperitoneal -Particle Radioimmunotherapy of Ovarian Cancer Patients: Pharmacokinetics and Dosimetry of 211At-MX35 F(ab’)2--A Phase I Study. J. Nucl. Med. 2009, 50, 1153–1160. [Google Scholar] [CrossRef]
- Hallqvist, A.; Bergmark, K.; Bäck, T.; Andersson, H.; Dahm-Kähler, P.; Johansson, M.; Lindegren, S.; Jensen, H.; Jacobsson, L.; Hultborn, R.; et al. Intraperitoneal α-Emitting Radioimmunotherapy with 211At in Relapsed Ovarian Cancer: Long-Term Follow-up with Individual Absorbed Dose Estimations. J. Nucl. Med. 2019, 60, 1073–1079. [Google Scholar] [CrossRef]
- Nakamae, H.; Wilbur, D.S.; Hamlin, D.K.; Thakar, M.S.; Santos, E.B.; Fisher, D.R.; Kenoyer, A.L.; Pagel, J.M.; Press, O.W.; Storb, R.; et al. Biodistributions, Myelosuppression and Toxicities in Mice Treated with an Anti-CD45 Antibody Labeled with the α-Emitting Radionuclides Bismuth-213 or Astatine-211. Cancer Res. 2009, 69, 2408–2415. [Google Scholar] [CrossRef]
- Orozco, J.J.; Bäck, T.; Kenoyer, A.; Balkin, E.R.; Hamlin, D.K.; Wilbur, D.S.; Fisher, D.R.; Frayo, S.L.; Hylarides, M.D.; Green, D.J.; et al. Anti-CD45 radioimmunotherapy using 211At with bone marrow transplantation prolongs survival in a disseminated murine leukemia model. Blood 2013, 121, 3759–3767. [Google Scholar] [CrossRef]
- Bäck, T.; Chouin, N.; Lindegren, S.; Kahu, H.; Jensen, H.; Albertsson, P.; Palm, S. Cure of human ovarian carcinoma solid xenografts by fractionated [211At] alpha-radioimmunotherapy: Influence of tumor absorbed dose and effect on long-term survival. J. Nucl. Med. 2016, 58, 598–604. [Google Scholar] [CrossRef]
- Chérel, M.; Gouard, S.; Gaschet, J.; Saï-Maurel, C.; Bruchertseifer, F.; Morgenstern, A.; Bourgeois, M.; Gestin, J.-F.; Kraeber-Bodéré, F.; Barbet, J.; et al. 213Bi Radioimmunotherapy with an Anti-mCD138 Monoclonal Antibody in a Murine Model of Multiple Myeloma. J. Nucl. Med. 2013, 54, 1597–1604. [Google Scholar] [CrossRef] [PubMed]
- Fichou, N.; Gouard, S.; Maurel, C.; Barbet, J.; Ferrer, L.; Morgenstern, A.; Bruchertseifer, F.; Faivre-Chauvet, A.; Bigot-Corbel, E.; Davodeau, F.; et al. Single-Dose Anti-CD138 Radioimmunotherapy: Bismuth-213 is More Efficient than Lutetium-177 for Treatment of Multiple Myeloma in a Preclinical Model. Front. Med. 2015, 2, 76. [Google Scholar] [CrossRef] [PubMed]
- Bailly, C.; Gouard, S.; Lacombe, M.; Saec, P.R.-L.; Chalopin, B.; Bourgeois, M.; Chouin, N.; Tripier, R.; Halime, Z.; Haddad, F.; et al. Comparison of Immuno-PET of CD138 and PET imaging with 64CuCl2 and 18F-FDG in a preclinical syngeneic model of multiple myeloma. Oncotarget 2018, 9, 9061–9072. [Google Scholar] [CrossRef] [PubMed]
- Bailly, C.; Gouard, S.; Guérard, F.; Chalopin, B.; Carlier, T.; Faivre-Chauvet, A.; Saëc, P.R.-L.; Bourgeois, M.; Chouin, N.; Rbah-Vidal, L.; et al. What is the Best Radionuclide for Immuno-PET of Multiple Myeloma? A Comparison Study Between 89Zr- and 64Cu-Labeled Anti-CD138 in a Preclinical Syngeneic Model. Int. J. Mol. Sci. 2019, 20, 2564. [Google Scholar] [CrossRef] [PubMed]
- Guérard, F.; Navarro, L.; Lee, Y.-S.; Roumesy, A.; Alliot, C.; Chérel, M.; Brechbiel, M.; Gestin, J.-F. Bifunctional aryliodonium salts for highly efficient radioiodination and astatination of antibodies. Bioorganic Med. Chem. 2017, 25, 5975–5980. [Google Scholar] [CrossRef] [PubMed]
- Larsen, R.H.; Slade, S.; Zalutsky, M.R. Blocking [211At]astatide accumulation in normal tissues: Preliminary evaluation of seven potential compounds. Nucl. Med. Biol. 1998, 25, 351–357. [Google Scholar] [CrossRef]
- Fernández, I.; Peña, A.; Del Teso, N.; Pérez, V.; Rodríguez-Cuesta, J. Rodríguez-Cuesta, Clinical biochemistry parameters in C57BL/6J mice after blood collection from the submandibular vein and retroorbital plexus. J. Am. Assoc. Lab. Anim. Sci. JAALAS 2010, 49, 202–206. [Google Scholar]
- Thoolen, B.; Maronpot, R.R.; Harada, T.; Nyska, A.; Rousseaux, C.; Nolte, T.; Malarkey, D.E.; Kaufmann, W.; Küttler, K.; Deschl, U.; et al. Proliferative and Nonproliferative Lesions of the Rat and Mouse Hepatobiliary System. Toxicol. Pathol. 2010, 38, 5S–81S. [Google Scholar] [CrossRef]
- Frazier, K.S.; Seely, J.C.; Hard, G.C.; Betton, G.; Burnett, R.; Nakatsuji, S.; Nishikawa, A.; Durchfeld-Meyer, B.; Bube, A. Proliferative and Nonproliferative Lesions of the Rat and Mouse Urinary System. Toxicol. Pathol. 2012, 40, 14S–86S. [Google Scholar] [CrossRef]
- Manning, L.; Berger, J.; O’Donoghue, H.; Sheridan, G.; Claringbold, P.; Turner, J.H. A model of multiple myeloma: Culture of 5T33 murine myeloma cells and evaluation of tumorigenicity in the C57BL/KaLwRij mouse. Br. J. Cancer 1992, 66, 1088–1093. [Google Scholar] [CrossRef]
- Turner, J.H.; Claringbold, P.G.; Manning, L.S.; O’Donoghue, H.L.; Berger, J.D.; Glancy, R.J. Radiopharmaceutical Therapy of 5T33 Murine Myeloma by Sequential Treatment With Samarium-153 Ethylenediaminetetramethylene Phosphonate, Melphalan, and Bone Marrow Transplantation. J. Natl. Cancer Inst. 1993, 85, 1508–1513. [Google Scholar] [CrossRef] [PubMed]
- Frassanito, M.A.; Rao, L.; Moschetta, M.; Ria, R.; Di Marzo, L.; De Luisi, A.; Racanelli, V.; Catacchio, I.; Berardi, S.; Basile, A.; et al. Bone marrow fibroblasts parallel multiple myeloma progression in patients and mice: In vitro and in vivo studies. Leukemia 2013, 28, 904–916. [Google Scholar] [CrossRef]
- Sonneveld, P.; De Wit, E.; Moreau, P. How have evolutions in strategies for the treatment of relapsed/refractory multiple myeloma translated into improved outcomes for patients? Crit. Rev. Oncol. 2017, 112, 153–170. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.-C.; Lin, S.-F. Mechanisms of Drug Resistance in Relapse and Refractory Multiple Myeloma. BioMed Res. Int. 2015, 2015, 341430. [Google Scholar] [CrossRef] [PubMed]
- Gao, M.; Kong, Y.; Yang, G.; Gao, L.; Shi, J. Multiple myeloma cancer stem cells. Oncotarget 2016, 7, 35466–35477. [Google Scholar] [CrossRef] [PubMed]
- Abdi, J.; Jian, H.; Chang, H. Role of micro-RNAs in drug resistance of multiple myeloma. Oncotarget 2016, 7, 60723–60735. [Google Scholar] [CrossRef]
- Roscher, M.; Hormann, I.; Leib, O.; Marx, S.; Moreno, J.; Miltner, E.; Friesen, C. Targeted alpha-therapy using [Bi-213]anti-CD20 as novel treatment option for radio- and chemoresistant non-Hodgkin lymphoma cells. Oncotarget 2013, 4, 218–230. [Google Scholar] [CrossRef]
- Yard, B.D.; Gopal, P.; Bannik, K.; Siemeister, G.; Hagemann, U.B.; Abazeed, M.E. Cellular and Genetic Determinants of the Sensitivity of Cancer to α-Particle Irradiation. Cancer Res. 2019, 79, 5640–5651. [Google Scholar] [CrossRef]
- Sgouros, G. α-Particle-Emitter Radiopharmaceutical Therapy: Resistance Is Futile. Cancer Res. 2019, 79, 5479–5481. [Google Scholar] [CrossRef]
- Gorin, J.-B.; Gouard, S.; Ménager, J.; Morgenstern, A.; Bruchertseifer, F.; Faivre-Chauvet, A.; Guilloux, Y.; Chérel, M.; Davodeau, F.; Gaschet, J. Alpha Particles Induce Autophagy in Multiple Myeloma Cells. Front. Med. 2015, 2, 74. [Google Scholar] [CrossRef]
- Chouin, N.; Bernardeau, K.; Bardies, M.; Faivre-Chauvet, A.; Bourgeois, M.; Apostolidis, C.; Morgenstern, A.; Lisbona, A.; Cherel, M.; Davodeau, F. Evidence of Extranuclear Cell Sensitivity to Alpha-Particle Radiation Using a Microdosimetric Model. II. Application of the Microdosimetric Model to Experimental Results. Radiat. Res. 2009, 171, 664–673. [Google Scholar] [CrossRef] [PubMed]
- Hoyos, V.; Borrello, I. The immunotherapy era of myeloma: Monoclonal antibodies, vaccines, and adoptive T-cell therapies. Blood 2016, 128, 1679–1687. [Google Scholar] [CrossRef] [PubMed]
- Sherbenou, D.W.; Mark, T.M.; Forsberg, P. Monoclonal Antibodies in Multiple Myeloma: A New Wave of the Future. Clin. Lymphoma Myeloma Leuk. 2017, 17, 545–554. [Google Scholar] [CrossRef] [PubMed]
- Ghai, A.; Maji, D.; Cho, N.; Chanswangphuwana, C.; Rettig, M.; Shen, D.; DiPersio, J.; Akers, W.; Dehdashti, F.; Achilefu, S.; et al. Preclinical Development of CD38-Targeted [89Zr]Zr-DFO-Daratumumab for Imaging Multiple Myeloma. J. Nucl. Med. 2017, 59, 216–222. [Google Scholar] [CrossRef] [PubMed]
- Cannons, J.L.; Tangye, S.; Schwartzberg, P.L. SLAM Family Receptors and SAP Adaptors in Immunity. Annu. Rev. Immunol. 2011, 29, 665–705. [Google Scholar] [CrossRef]
- Tissue Expression of SLAMF7—Summary—The Human Protein Atlas. Available online: https://www.proteinatlas.org/ENSG00000026751-SLAMF7/tissue (accessed on 30 March 2020).
- Quelven, I.; Monteil, J.; Sage, M.; Saidi, A.; Mounier, J.; Bayout, A.; Garrier, J.; Cogne, M.; Durand-Panteix, S. 212Pb Alpha-Radioimmunotherapy targeting CD38 in Multiple Myeloma: A preclinical study. J. Nucl. Med. 2019. [Google Scholar] [CrossRef]
- O’Steen, S.; Comstock, M.L.; Orozco, J.J.; Hamlin, D.K.; Wilbur, D.S.; Jones, J.; Kenoyer, A.; Nartea, M.E.; Lin, Y.; Miller, B.W.; et al. The α-emitter astatine-211 targeted to CD38 can eradicate multiple myeloma in a disseminated disease model. Blood 2019, 134, 1247–1256. [Google Scholar] [CrossRef]
- Gouard, S.; Pallardy, A.; Gaschet, J.; Faivre-Chauvet, A.; Bruchertseifer, F.; Morgenstern, A.; Maurel, C.; Matous, E.; Kraeber-Bodéré, F.; Davodeau, F.; et al. Comparative analysis of multiple myeloma treatment by CD138 antigen targeting with bismuth-213 and Melphalan chemotherapy. Nucl. Med. Biol. 2014, 41, e30–e35. [Google Scholar] [CrossRef]
- Wu, C.; Zhang, L.; Brockman, Q.R.; Zhan, F.; Chen, L. Chimeric antigen receptor T cell therapies for multiple myeloma. J. Hematol. Oncol. 2019, 12, 120. [Google Scholar] [CrossRef]
- Hobbs, R.F.; Howell, R.W.; Song, H.; Baechler, S.; Sgouros, G. Redefining relative biological effectiveness in the context of the EQDX formalism: Implications for alpha-particle emitter therapy. Radiat. Res. 2014, 181, 90–98. [Google Scholar] [CrossRef]
- Dorso, L.; Bigot-Corbel, E.; Abadie, J.; Diab, M.; Gouard, S.; Bruchertseifer, F.; Morgenstern, A.; Maurel, C.; Chérel, M.; Davodeau, F. Long-Term Toxicity of 213Bi-Labelled BSA in Mice. PLoS ONE 2016, 11, e0151330. [Google Scholar] [CrossRef] [PubMed]
- Rousseau, C.; Ferrer, L.; Supiot, S.; Bardies, M.; Davodeau, F.; Faivre-Chauvet, A.; Baumgartner, P.; Wijdenes, J.; Lacombe, M.; Barbet, J.; et al. Dosimetry results suggest feasibility of radioimmunotherapy using anti-CD138 (B-B4) antibody in multiple myeloma patients. Tumor Biol. 2012, 33, 679–688. [Google Scholar] [CrossRef] [PubMed]
- Navarro, L.; Berdal, M.; Chérel, M.; Pecorari, F.; Gestin, J.-F.; Guérard, F. Prosthetic groups for radioiodination and astatination of peptides and proteins: A comparative study of five potential bioorthogonal labeling strategies. Bioorganic Med. Chem. 2019, 27, 167–174. [Google Scholar] [CrossRef] [PubMed]
- Eckerman, K.F.; Endo, A. MIRD: Radionuclide Data and Decay Schemes; Society of Nuclear Medicine: Reston, VA, USA, 2008. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Mice/Group | Surviving Mice | Dead Mice | Cause of Death | ||||
|---|---|---|---|---|---|---|---|---|
| Acute Toxicity | Paraplegia | Extramedullary Lesions | No Macroscopic Lesion but High Serum IgG Level | Undetermined Death | ||||
| Control | 16 | 0 | 16 | - | 7 | 9 (2) | 1 | 1 |
| 9E7.4 at 370 kBq | 16 | 2 | 14 | - | 4 | 10 (2) | - | 2 |
| 9E7.4 at 555 kBq | 10 | 3 | 7 | - | 4 | 5 (3) | - | 1 |
| 9E7.4 at 740 kBq | 16 | 10 | 6 | - | 3 | 4 (1) | - | - |
| 9E7.4 at 1110 kBq | 6 | 0 | 6 | 6 | - | - | - | - |
| Isotype control at 555 kBq | 10 | 0 | 10 | - | 5 | 5 (1) | 1 | - |
| Organs | Absorbed Dose (Gy/MBq) | SE (Gy/MBq) |
|---|---|---|
| Blood | 8.0 | 0.7 |
| Flat Bone | 1.5 | 0.2 |
| Liver | 10.9 | 1.0 |
| Kidneys | 3.6 | 0.3 |
| Gut | 6.6 | 1.3 |
| Lungs | 5.8 | 0.8 |
| Muscle | 0.8 | 0.3 |
| Spleen | 8.2 | 1.7 |
| Skin | 2.6 | 0.9 |
| Brain | 0.2 | 0.0 |
| Heart | 3.8 | 0.6 |
| Femur | 1.7 | 0.3 |
| Stomach | 9.5 | 4.7 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gouard, S.; Maurel, C.; Marionneau-Lambot, S.; Dansette, D.; Bailly, C.; Guérard, F.; Chouin, N.; Haddad, F.; Alliot, C.; Gaschet, J.; et al. Targeted-Alpha-Therapy Combining Astatine-211 and anti-CD138 Antibody in a Preclinical Syngeneic Mouse Model of Multiple Myeloma Minimal Residual Disease. Cancers 2020, 12, 2721. https://doi.org/10.3390/cancers12092721
Gouard S, Maurel C, Marionneau-Lambot S, Dansette D, Bailly C, Guérard F, Chouin N, Haddad F, Alliot C, Gaschet J, et al. Targeted-Alpha-Therapy Combining Astatine-211 and anti-CD138 Antibody in a Preclinical Syngeneic Mouse Model of Multiple Myeloma Minimal Residual Disease. Cancers. 2020; 12(9):2721. https://doi.org/10.3390/cancers12092721
Chicago/Turabian StyleGouard, Sébastien, Catherine Maurel, Séverine Marionneau-Lambot, Delphine Dansette, Clément Bailly, François Guérard, Nicolas Chouin, Ferid Haddad, Cyril Alliot, Joëlle Gaschet, and et al. 2020. "Targeted-Alpha-Therapy Combining Astatine-211 and anti-CD138 Antibody in a Preclinical Syngeneic Mouse Model of Multiple Myeloma Minimal Residual Disease" Cancers 12, no. 9: 2721. https://doi.org/10.3390/cancers12092721
APA StyleGouard, S., Maurel, C., Marionneau-Lambot, S., Dansette, D., Bailly, C., Guérard, F., Chouin, N., Haddad, F., Alliot, C., Gaschet, J., Eychenne, R., Kraeber-Bodéré, F., & Chérel, M. (2020). Targeted-Alpha-Therapy Combining Astatine-211 and anti-CD138 Antibody in a Preclinical Syngeneic Mouse Model of Multiple Myeloma Minimal Residual Disease. Cancers, 12(9), 2721. https://doi.org/10.3390/cancers12092721