Hormones Secretion and Rho GTPases in Neuroendocrine Tumors


Abstract
1. Introduction
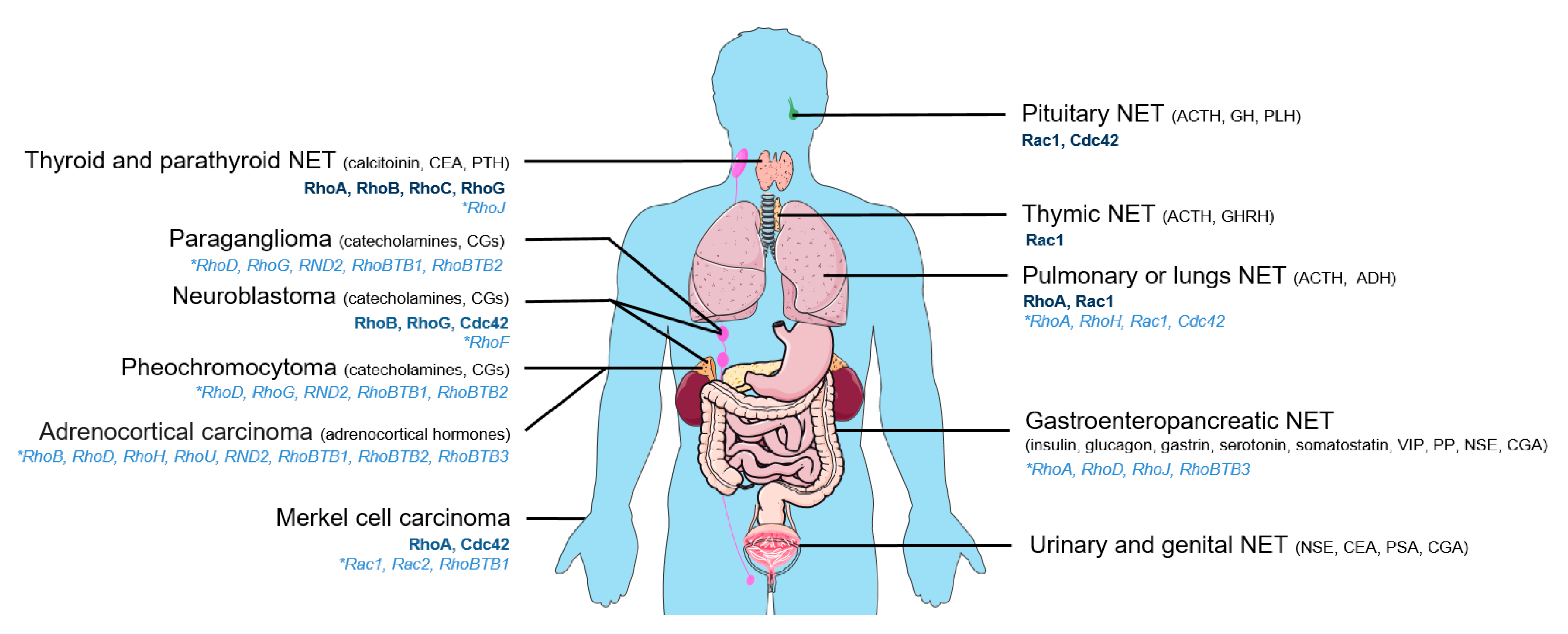
2. Neuroendocrine Tumors and Rho GTPases
3. Control of Rho Activity in NETs: Important Role of Rho GEFs and GAPs
4. Rho GTPases and Hormones Secretion
4.1. Control of Secretion through Actin Remodeling
4.2. Control of Secretion through Lipids Action
4.3. Rho GEFs and Rho GAPs at the Commands
5. Conclusions
Supplementary Materials
Funding
Acknowledgments
Conflicts of Interest
References
- Kulke, M.H.; Shah, M.H.; Benson, A.B., 3rd; Bergsland, E.; Berlin, J.D.; Blaszkowsky, L.S.; Emerson, L.; Engstrom, P.F.; Fanta, P.; Giordano, T.; et al. Neuroendocrine tumors, version 1.2015. J. Natl. Compr. Canc. Netw. 2015, 13, 78–108. [Google Scholar] [CrossRef] [PubMed]
- Zandee, W.T.; Kamp, K.; van Adrichem, R.C.; Feelders, R.A.; de Herder, W.W. Effect of hormone secretory syndromes on neuroendocrine tumor prognosis. Endocr. Relat. Cancer 2017, 24, R261–R274. [Google Scholar] [CrossRef] [PubMed]
- Juhlin, C.C.; Skoglund, S.; Juntti-Berggren, L.; Karlberg, M.; Calissendorff, J. Non-functioning neuroendocrine pancreatic tumors transforming to malignant insulinomas - four cases and review of the literature. Neuro. Endocrinol. Lett. 2019, 40, 175–183. [Google Scholar] [PubMed]
- Song, P.; Sekhon, H.S.; Jia, Y.; Keller, J.A.; Blusztajn, J.K.; Mark, G.P.; Spindel, E.R. Acetylcholine is synthesized by and acts as an autocrine growth factor for small cell lung carcinoma. Cancer Res. 2003, 63, 214–221. [Google Scholar]
- Brown, R.L.; Muzzafar, T.; Wollman, R.; Weiss, R.E. A pituitary carcinoma secreting TSH and prolactin: A non-secreting adenoma gone awry. Eur. J. Endocrinol. 2006, 154, 639–643. [Google Scholar] [CrossRef]
- Daems, T.; Verhelst, J.; Michotte, A.; Abrams, P.; De Ridder, D.; Abs, R. Modification of hormonal secretion in clinically silent pituitary adenomas. Pituitary 2009, 12, 80–86. [Google Scholar] [CrossRef]
- Lenders, N.; McCormack, A. Malignant transformation in non-functioning pituitary adenomas (pituitary carcinoma). Pituitary 2018, 21, 217–229. [Google Scholar] [CrossRef]
- Neumann, H.P.H.; Young, W.F., Jr.; Eng, C. Pheochromocytoma and Paraganglioma. N. Engl. J. Med. 2019, 381, 552–565. [Google Scholar] [CrossRef]
- Aspenstrom, P.; Ruusala, A.; Pacholsky, D. Taking Rho GTPases to the next level: The cellular functions of atypical Rho GTPases. Exp. Cell Res. 2007, 313, 3673–3679. [Google Scholar] [CrossRef]
- Boureux, A.; Vignal, E.; Faure, S.; Fort, P. Evolution of the Rho family of ras-like GTPases in eukaryotes. Mol. Biol. Evol. 2007, 24, 203–216. [Google Scholar] [CrossRef]
- Croise, P.; Estay-Ahumada, C.; Gasman, S.; Ory, S. Rho GTPases, phosphoinositides, and actin: A tripartite framework for efficient vesicular trafficking. Small GTPases 2014, 5, e29469. [Google Scholar] [CrossRef] [PubMed]
- Haga, R.B.; Ridley, A.J. Rho GTPases: Regulation and roles in cancer cell biology. Small GTPases 2016, 7, 207–221. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Peyrollier, K.; Kilic, G.; Brakebusch, C. Rho GTPases and cancer. Biofactors 2014, 40, 226–235. [Google Scholar] [CrossRef] [PubMed]
- Olayioye, M.A.; Noll, B.; Hausser, A. Spatiotemporal Control of Intracellular Membrane Trafficking by Rho GTPases. Cells 2019, 8, 1478. [Google Scholar] [CrossRef] [PubMed]
- Orgaz, J.L.; Herraiz, C.; Sanz-Moreno, V. Rho GTPases modulate malignant transformation of tumor cells. Small GTPases 2014, 5, e29019. [Google Scholar] [CrossRef]
- Phuyal, S.; Farhan, H. Multifaceted Rho GTPase Signaling at the Endomembranes. Front. Cell Dev. Biol. 2019, 7, 127. [Google Scholar] [CrossRef]
- Porter, A.P.; Papaioannou, A.; Malliri, A. Deregulation of Rho GTPases in cancer. Small GTPases 2016, 7, 123–138. [Google Scholar] [CrossRef]
- Warner, H.; Wilson, B.J.; Caswell, P.T. Control of adhesion and protrusion in cell migration by Rho GTPases. Curr. Opin. Cell. Biol. 2019, 56, 64–70. [Google Scholar] [CrossRef]
- Wilson, K.F.; Erickson, J.W.; Antonyak, M.A.; Cerione, R.A. Rho GTPases and their roles in cancer metabolism. Trends Mol. Med. 2013, 19, 74–82. [Google Scholar] [CrossRef]
- Alan, J.K.; Lundquist, E.A. Mutationally activated Rho GTPases in cancer. Small GTPases 2013, 4, 159–163. [Google Scholar] [CrossRef]
- Bustelo, X.R. RHO GTPases in cancer: Known facts, open questions, and therapeutic challenges. Biochem. Soc. Trans. 2018, 46, 741–760. [Google Scholar] [CrossRef] [PubMed]
- Kataoka, K.; Ogawa, S. Variegated RHOA mutations in human cancers. Exp. Hematol. 2016, 44, 1123–1129. [Google Scholar] [CrossRef] [PubMed]
- Olson, M.F. Rho GTPases, their post-translational modifications, disease-associated mutations and pharmacological inhibitors. Small GTPases 2018, 9, 203–215. [Google Scholar] [CrossRef] [PubMed]
- Svensmark, J.H.; Brakebusch, C. Rho GTPases in cancer: Friend or foe? Oncogene 2019, 38, 7447–7456. [Google Scholar] [CrossRef] [PubMed]
- Zandvakili, I.; Lin, Y.; Morris, J.C.; Zheng, Y. Rho GTPases: Anti- or pro-neoplastic targets? Oncogene 2017, 36, 3213–3222. [Google Scholar] [CrossRef]
- Davis, M.J.; Ha, B.H.; Holman, E.C.; Halaban, R.; Schlessinger, J.; Boggon, T.J. RAC1P29S is a spontaneously activating cancer-associated GTPase. Proc. Natl. Acad. Sci. USA 2013, 110, 912–917. [Google Scholar] [CrossRef]
- Kawazu, M.; Ueno, T.; Kontani, K.; Ogita, Y.; Ando, M.; Fukumura, K.; Yamato, A.; Soda, M.; Takeuchi, K.; Miki, Y.; et al. Transforming mutations of RAC guanosine triphosphatases in human cancers. Proc. Natl. Acad. Sci. USA 2013, 110, 3029–3034. [Google Scholar] [CrossRef]
- Zhang, H.; Schaefer, A.; Wang, Y.; Hodge, R.G.; Blake, D.R.; Diehl, J.N.; Papageorge, A.G.; Stachler, M.D.; Liao, J.; Zhou, J.; et al. Gain-of-Function RHOA Mutations Promote Focal Adhesion Kinase Activation and Dependency in Diffuse Gastric Cancer. Cancer Discov. 2020, 10, 288–305. [Google Scholar] [CrossRef]
- Wang, K.; Yuen, S.T.; Xu, J.; Lee, S.P.; Yan, H.H.; Shi, S.T.; Siu, H.C.; Deng, S.; Chu, K.M.; Law, S.; et al. Whole-genome sequencing and comprehensive molecular profiling identify new driver mutations in gastric cancer. Nat. Genet. 2014, 46, 573–582. [Google Scholar] [CrossRef]
- Augert, A.; Zhang, Q.; Bates, B.; Cui, M.; Wang, X.; Wildey, G.; Dowlati, A.; MacPherson, D. Small Cell Lung Cancer Exhibits Frequent Inactivating Mutations in the Histone Methyltransferase KMT2D/MLL2: CALGB 151111 (Alliance). J. Thorac. Oncol. 2017, 12, 704–713. [Google Scholar] [CrossRef]
- Wen, K.W.; Grenert, J.P.; Joseph, N.M.; Shafizadeh, N.; Huang, A.; Hosseini, M.; Kakar, S. Genomic profile of appendiceal goblet cell carcinoid is distinct compared to appendiceal neuroendocrine tumor and conventional adenocarcinoma. Hum. Pathol. 2018, 77, 166–174. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Zhou, W.; Li, L.; Wang, J.; Gao, Z.; Jiang, Y.; Jiang, X.; Shan, A.; Bailey, M.H.; Huang, K.L.; et al. Pan-cancer analysis of somatic mutations across 21 neuroendocrine tumor types. Cell Res. 2018, 28, 601–604. [Google Scholar] [CrossRef]
- Wilzen, A.; Rehammar, A.; Muth, A.; Nilsson, O.; Tesan Tomic, T.; Wangberg, B.; Kristiansson, E.; Abel, F. Malignant pheochromocytomas/paragangliomas harbor mutations in transport and cell adhesion genes. Int. J. Cancer 2016, 138, 2201–2211. [Google Scholar] [CrossRef] [PubMed]
- Flynn, A.; Benn, D.; Clifton-Bligh, R.; Robinson, B.; Trainer, A.H.; James, P.; Hogg, A.; Waldeck, K.; George, J.; Li, J.; et al. The genomic landscape of phaeochromocytoma. J. Pathol. 2015, 236, 78–89. [Google Scholar] [CrossRef]
- Scarpa, A.; Chang, D.K.; Nones, K.; Corbo, V.; Patch, A.M.; Bailey, P.; Lawlor, R.T.; Johns, A.L.; Miller, D.K.; Mafficini, A.; et al. Whole-genome landscape of pancreatic neuroendocrine tumours. Nature 2017, 543, 65–71. [Google Scholar] [CrossRef] [PubMed]
- George, J.; Walter, V.; Peifer, M.; Alexandrov, L.B.; Seidel, D.; Leenders, F.; Maas, L.; Muller, C.; Dahmen, I.; Delhomme, T.M.; et al. Integrative genomic profiling of large-cell neuroendocrine carcinomas reveals distinct subtypes of high-grade neuroendocrine lung tumors. Nat. Commun. 2018, 9, 1048. [Google Scholar] [CrossRef]
- Francis, J.M.; Kiezun, A.; Ramos, A.H.; Serra, S.; Pedamallu, C.S.; Qian, Z.R.; Banck, M.S.; Kanwar, R.; Kulkarni, A.A.; Karpathakis, A.; et al. Somatic mutation of CDKN1B in small intestine neuroendocrine tumors. Nat. Genet. 2013, 45, 1483–1486. [Google Scholar] [CrossRef]
- Harms, P.W.; Vats, P.; Verhaegen, M.E.; Robinson, D.R.; Wu, Y.M.; Dhanasekaran, S.M.; Palanisamy, N.; Siddiqui, J.; Cao, X.; Su, F.; et al. The Distinctive Mutational Spectra of Polyomavirus-Negative Merkel Cell Carcinoma. Cancer Res. 2015, 75, 3720–3727. [Google Scholar] [CrossRef]
- Castro-Vega, L.J.; Letouze, E.; Burnichon, N.; Buffet, A.; Disderot, P.H.; Khalifa, E.; Loriot, C.; Elarouci, N.; Morin, A.; Menara, M.; et al. Multi-omics analysis defines core genomic alterations in pheochromocytomas and paragangliomas. Nat. Commun. 2015, 6, 6044. [Google Scholar] [CrossRef]
- Ellenbroek, S.I.; Collard, J.G. Rho GTPases: Functions and association with cancer. Clin. Exp. Metastasis 2007, 24, 657–672. [Google Scholar] [CrossRef]
- Karlsson, R.; Pedersen, E.D.; Wang, Z.; Brakebusch, C. Rho GTPase function in tumorigenesis. Biochim. Biophys. Acta 2009, 1796, 91–98. [Google Scholar] [CrossRef]
- Gomez del Pulgar, T.; Benitah, S.A.; Valeron, P.F.; Espina, C.; Lacal, J.C. Rho GTPase expression in tumourigenesis: Evidence for a significant link. Bioessays 2005, 27, 602–613. [Google Scholar] [CrossRef] [PubMed]
- Sahai, E.; Marshall, C.J. RHO-GTPases and cancer. Nat. Rev. Cancer 2002, 2, 133–142. [Google Scholar] [CrossRef] [PubMed]
- Long, Y.; Lu, M.; Cheng, T.; Zhan, X.; Zhan, X. Multiomics-Based Signaling Pathway Network Alterations in Human Non-functional Pituitary Adenomas. Front. Endocrinol. (Lausanne) 2019, 10, 835. [Google Scholar] [CrossRef] [PubMed]
- Veschi, V.; Verona, F.; Thiele, C.J. Cancer Stem Cells and Neuroblastoma: Characteristics and Therapeutic Targeting Options. Front. Endocrinol. (Lausanne) 2019, 10, 782. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.R.; Song, Y.K.; Yu, L.R.; Wei, J.S.; Chung, J.Y.; Hewitt, S.M.; Veenstra, T.D.; Khan, J. Global genomic and proteomic analysis identifies biological pathways related to high-risk neuroblastoma. J. Proteome Res. 2010, 9, 373–382. [Google Scholar] [CrossRef]
- Coco, S.; Theissen, J.; Scaruffi, P.; Stigliani, S.; Moretti, S.; Oberthuer, A.; Valdora, F.; Fischer, M.; Gallo, F.; Hero, B.; et al. Age-dependent accumulation of genomic aberrations and deregulation of cell cycle and telomerase genes in metastatic neuroblastoma. Int. J. Cancer 2012, 131, 1591–1600. [Google Scholar] [CrossRef]
- Stigliani, S.; Coco, S.; Moretti, S.; Oberthuer, A.; Fischer, M.; Theissen, J.; Gallo, F.; Garavent, A.; Berthold, F.; Bonassi, S.; et al. High genomic instability predicts survival in metastatic high-risk neuroblastoma. Neoplasia 2012, 14, 823–832. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, C.; Contreras, B.; Iwanaga, T.; Takegami, Y.; Bakker, A.; Bronson, R.T.; Noda, M.; Loda, M.; Hunt, J.L.; Ewen, M.E. Nras loss induces metastatic conversion of Rb1-deficient neuroendocrine thyroid tumor. Nat. Genet. 2006, 38, 118–123. [Google Scholar] [CrossRef]
- Gawin, M.; Wojakowska, A.; Pietrowska, M.; Marczak, L.; Chekan, M.; Jelonek, K.; Lange, D.; Jaksik, R.; Gruca, A.; Widlak, P. Proteome profiles of different types of thyroid cancers. Mol. Cell. Endocrinol. 2018, 472, 68–79. [Google Scholar] [CrossRef]
- Rekhtman, N. Neuroendocrine tumors of the lung: An update. Arch. Pathol. Lab. Med. 2010, 134, 1628–1638. [Google Scholar] [CrossRef]
- Touge, H.; Chikumi, H.; Igishi, T.; Kurai, J.; Makino, H.; Tamura, Y.; Takata, M.; Yoneda, K.; Nakamoto, M.; Suyama, H.; et al. Diverse activation states of RhoA in human lung cancer cells: Contribution of G protein coupled receptors. Int. J. Oncol. 2007, 30, 709–715. [Google Scholar] [CrossRef] [PubMed]
- Varker, K.A.; Phelps, S.H.; King, M.M.; Williams, C.L. The small GTPase RhoA has greater expression in small cell lung carcinoma than in non-small cell lung carcinoma and contributes to their unique morphologies. Int. J. Oncol. 2003, 22, 671–681. [Google Scholar] [PubMed]
- Fujii, K.; Miyata, Y.; Takahashi, I.; Koizumi, H.; Saji, H.; Hoshikawa, M.; Takagi, M.; Nishimura, T.; Nakamura, H. Differential Proteomic Analysis between Small Cell Lung Carcinoma (SCLC) and Pulmonary Carcinoid Tumors Reveals Molecular Signatures for Malignancy in Lung Cancer. Proteom. Clin. Appl. 2018, 12, e1800015. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.X.; Wang, W.Q.; Ye, L.; Bi, Y.F.; Fang, H.; Cui, B.; Zhou, W.W.; Dai, M.; Zhang, J.; Li, X.Y.; et al. p21-activated kinase 3 is overexpressed in thymic neuroendocrine tumors (carcinoids) with ectopic ACTH syndrome and participates in cell migration. Endocrine 2010, 38, 38–47. [Google Scholar] [CrossRef]
- Stakaityte, G.; Nwogu, N.; Dobson, S.J.; Knight, L.M.; Wasson, C.W.; Salguero, F.J.; Blackbourn, D.J.; Blair, G.E.; Mankouri, J.; Macdonald, A.; et al. Merkel Cell Polyomavirus Small T Antigen Drives Cell Motility via Rho-GTPase-Induced Filopodium Formation. J. Virol. 2018, 92. [Google Scholar] [CrossRef]
- Croise, P.; Houy, S.; Gand, M.; Lanoix, J.; Calco, V.; Toth, P.; Brunaud, L.; Lomazzi, S.; Paramithiotis, E.; Chelsky, D.; et al. Cdc42 and Rac1 activity is reduced in human pheochromocytoma and correlates with FARP1 and ARHGEF1 expression. Endocr. Relat. Cancer 2016, 23, 281–293. [Google Scholar] [CrossRef]
- Croise, P.; Brunaud, L.; Toth, P.; Gasman, S.; Ory, S. Inhibition of Cdc42 and Rac1 activities in pheochromocytoma, the adrenal medulla tumor. Small GTPases 2017, 8, 122–127. [Google Scholar] [CrossRef][Green Version]
- de Cubas, A.A.; Leandro-Garcia, L.J.; Schiavi, F.; Mancikova, V.; Comino-Mendez, I.; Inglada-Perez, L.; Perez-Martinez, M.; Ibarz, N.; Ximenez-Embun, P.; Lopez-Jimenez, E.; et al. Integrative analysis of miRNA and mRNA expression profiles in pheochromocytoma and paraganglioma identifies genotype-specific markers and potentially regulated pathways. Endocr. Relat. Cancer 2013, 20, 477–493. [Google Scholar] [CrossRef]
- Bossan, A.; Ottman, R.; Andl, T.; Hasan, M.F.; Mahajan, N.; Coppola, D.; Chakrabarti, R. Expression of FGD4 positively correlates with the aggressive phenotype of prostate cancer. BMC Cancer 2018, 18, 1257. [Google Scholar] [CrossRef]
- Shahid, M.; George, T.B.; Saller, J.; Haija, M.; Sayegh, Z.; Boulware, D.; Strosberg, J.; Chakrabarti, R.; Coppola, D. FGD4 (Frabin) Overexpression in Pancreatic Neuroendocrine Neoplasms. Pancreas 2019, 48, 1307–1311. [Google Scholar] [CrossRef]
- Lazer, G.; Idelchuk, Y.; Schapira, V.; Pikarsky, E.; Katzav, S. The haematopoietic specific signal transducer Vav1 is aberrantly expressed in lung cancer and plays a role in tumourigenesis. J. Pathol. 2009, 219, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Ziv, T.; Barnea, E.; Segal, H.; Sharon, R.; Beer, I.; Admon, A. Comparative proteomics of small cell lung carcinoma. Cancer Biomark. 2006, 2, 219–234. [Google Scholar] [CrossRef] [PubMed]
- Weissenrieder, J.S.; Reilly, J.E.; Neighbors, J.D.; Hohl, R.J. Inhibiting geranylgeranyl diphosphate synthesis reduces nuclear androgen receptor signaling and neuroendocrine differentiation in prostate cancer cell models. Prostate 2019, 79, 21–30. [Google Scholar] [CrossRef]
- Arya, A.K.; Bhadada, S.K.; Singh, P.; Dahiya, D.; Kaur, G.; Sharma, S.; Saikia, U.N.; Behera, A.; Rao, S.D.; Bhasin, M. Quantitative proteomics analysis of sporadic parathyroid adenoma tissue samples. J. Endocrinol. Investig. 2019, 42, 577–590. [Google Scholar] [CrossRef] [PubMed]
- Tsai, H.K.; Lehrer, J.; Alshalalfa, M.; Erho, N.; Davicioni, E.; Lotan, T.L. Gene expression signatures of neuroendocrine prostate cancer and primary small cell prostatic carcinoma. BMC Cancer 2017, 17, 759. [Google Scholar] [CrossRef] [PubMed]
- Bader, M.F.; Doussau, F.; Chasserot-Golaz, S.; Vitale, N.; Gasman, S. Coupling actin and membrane dynamics during calcium-regulated exocytosis: A role for Rho and ARF GTPases. Biochim. Biophys. Acta 2004, 1742, 37–49. [Google Scholar] [CrossRef]
- Gasman, S.; Chasserot-Golaz, S.; Bader, M.F.; Vitale, N. Regulation of exocytosis in adrenal chromaffin cells: Focus on ARF and Rho GTPases. Cell Signal. 2003, 15, 893–899. [Google Scholar] [CrossRef]
- Malacombe, M.; Bader, M.F.; Gasman, S. Exocytosis in neuroendocrine cells: New tasks for actin. Biochim. Biophys. Acta 2006, 1763, 1175–1183. [Google Scholar] [CrossRef]
- Momboisse, F.; Houy, S.; Ory, S.; Calco, V.; Bader, M.F.; Gasman, S. How important are Rho GTPases in neurosecretion? J. Neurochem. 2011, 117, 623–631. [Google Scholar] [CrossRef]
- Ory, S.; Gasman, S. Rho GTPases and exocytosis: What are the molecular links? Semin. Cell. Dev. Biol. 2011, 22, 27–32. [Google Scholar] [CrossRef] [PubMed]
- Bader, M.F.; Holz, R.W.; Kumakura, K.; Vitale, N. Exocytosis: The chromaffin cell as a model system. Ann. N. Y. Acad. Sci. 2002, 971, 178–183. [Google Scholar] [CrossRef]
- Westerink, R.H.; Ewing, A.G. The PC12 cell as model for neurosecretion. Acta. Physiol. (Oxf.) 2008, 192, 273–285. [Google Scholar] [CrossRef] [PubMed]
- Miyazaki, J.; Araki, K.; Yamato, E.; Ikegami, H.; Asano, T.; Shibasaki, Y.; Oka, Y.; Yamamura, K. Establishment of a pancreatic beta cell line that retains glucose-inducible insulin secretion: Special reference to expression of glucose transporter isoforms. Endocrinology 1990, 127, 126–132. [Google Scholar] [CrossRef]
- Flatt, P.R. Defective regulation of insulin secretion in diabetes and insulinoma. Biochem. Soc. Trans. 1990, 18, 124–127. [Google Scholar] [CrossRef] [PubMed]
- Okabayashi, T.; Shima, Y.; Sumiyoshi, T.; Kozuki, A.; Ito, S.; Ogawa, Y.; Kobayashi, M.; Hanazaki, K. Diagnosis and management of insulinoma. World J. Gastroenterol. 2013, 19, 829–837. [Google Scholar] [CrossRef] [PubMed]
- Gasman, S.; Chasserot-Golaz, S.; Popoff, M.R.; Aunis, D.; Bader, M.F. Involvement of Rho GTPases in calcium-regulated exocytosis from adrenal chromaffin cells. J. Cell. Sci. 1999, 112 Pt. 24, 4763–4771. [Google Scholar]
- Gasman, S.; Chasserot-Golaz, S.; Malacombe, M.; Way, M.; Bader, M.F. Regulated exocytosis in neuroendocrine cells: A role for subplasmalemmal Cdc42/N-WASP-induced actin filaments. Mol. Biol. Cell. 2004, 15, 520–531. [Google Scholar] [CrossRef] [PubMed]
- Gasman, S.; Chasserot-Golaz, S.; Hubert, P.; Aunis, D.; Bader, M.F. Identification of a potential effector pathway for the trimeric Go protein associated with secretory granules. Go stimulates a granule-bound phosphatidylinositol 4-kinase by activating RhoA in chromaffin cells. J. Biol. Chem. 1998, 273, 16913–16920. [Google Scholar] [CrossRef] [PubMed]
- Gasman, S.; Chasserot-Golaz, S.; Popoff, M.R.; Aunis, D.; Bader, M.F. Trimeric G proteins control exocytosis in chromaffin cells. Go regulates the peripheral actin network and catecholamine secretion by a mechanism involving the small GTP-binding protein Rho. J. Biol. Chem. 1997, 272, 20564–20571. [Google Scholar] [CrossRef] [PubMed]
- Kalwat, M.A.; Thurmond, D.C. Signaling mechanisms of glucose-induced F-actin remodeling in pancreatic islet beta cells. Exp. Mol. Med. 2013, 45, e37. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Oh, E.; Thurmond, D.C. Glucose-stimulated Cdc42 signaling is essential for the second phase of insulin secretion. J. Biol. Chem. 2007, 282, 9536–9546. [Google Scholar] [CrossRef] [PubMed]
- Uenishi, E.; Shibasaki, T.; Takahashi, H.; Seki, C.; Hamaguchi, H.; Yasuda, T.; Tatebe, M.; Oiso, Y.; Takenawa, T.; Seino, S. Actin dynamics regulated by the balance of neuronal Wiskott-Aldrich syndrome protein (N-WASP) and cofilin activities determines the biphasic response of glucose-induced insulin secretion. J. Biol. Chem. 2013, 288, 25851–25864. [Google Scholar] [CrossRef] [PubMed]
- Kalwat, M.A.; Yoder, S.M.; Wang, Z.; Thurmond, D.C. A p21-activated kinase (PAK1) signaling cascade coordinately regulates F-actin remodeling and insulin granule exocytosis in pancreatic beta cells. Biochem. Pharmacol. 2013, 85, 808–816. [Google Scholar] [CrossRef] [PubMed]
- Bretou, M.; Jouannot, O.; Fanget, I.; Pierobon, P.; Larochette, N.; Gestraud, P.; Guillon, M.; Emiliani, V.; Gasman, S.; Desnos, C.; et al. Cdc42 controls the dilation of the exocytotic fusion pore by regulating membrane tension. Mol. Biol. Cell. 2014, 25, 3195–3209. [Google Scholar] [CrossRef]
- Gabel, M.; Delavoie, F.; Demais, V.; Royer, C.; Bailly, Y.; Vitale, N.; Bader, M.F.; Chasserot-Golaz, S. Annexin A2-dependent actin bundling promotes secretory granule docking to the plasma membrane and exocytosis. J. Cell. Biol. 2015, 210, 785–800. [Google Scholar] [CrossRef]
- Ma, W.; Chang, J.; Tong, J.; Ho, U.; Yau, B.; Kebede, M.A.; Thorn, P. Arp2/3 nucleates F-actin coating of fusing insulin granules in pancreatic beta cells to control insulin secretion. J. Cell. Sci. 2020, 133. [Google Scholar] [CrossRef]
- Gomes, E.R.; Jani, S.; Gundersen, G.G. Nuclear movement regulated by Cdc42, MRCK, myosin, and actin flow establishes MTOC polarization in migrating cells. Cell 2005, 121, 451–463. [Google Scholar] [CrossRef]
- Brooks, A.B.; Humphreys, D.; Singh, V.; Davidson, A.C.; Arden, S.D.; Buss, F.; Koronakis, V. MYO6 is targeted by Salmonella virulence effectors to trigger PI3-kinase signaling and pathogen invasion into host cells. Proc. Natl. Acad. Sci. USA 2017, 114, 3915–3920. [Google Scholar] [CrossRef]
- Kimura, K.; Ito, M.; Amano, M.; Chihara, K.; Fukata, Y.; Nakafuku, M.; Yamamori, B.; Feng, J.; Nakano, T.; Okawa, K.; et al. Regulation of myosin phosphatase by Rho and Rho-associated kinase (Rho-kinase). Science 1996, 273, 245–248. [Google Scholar] [CrossRef]
- Wilkinson, S.; Paterson, H.F.; Marshall, C.J. Cdc42-MRCK and Rho-ROCK signalling cooperate in myosin phosphorylation and cell invasion. Nat. Cell. Biol. 2005, 7, 255–261. [Google Scholar] [CrossRef] [PubMed]
- Arous, C.; Rondas, D.; Halban, P.A. Non-muscle myosin IIA is involved in focal adhesion and actin remodelling controlling glucose-stimulated insulin secretion. Diabetologia 2013, 56, 792–802. [Google Scholar] [CrossRef] [PubMed]
- Berberian, K.; Torres, A.J.; Fang, Q.; Kisler, K.; Lindau, M. F-actin and myosin II accelerate catecholamine release from chromaffin granules. J. Neurosci. 2009, 29, 863–870. [Google Scholar] [CrossRef] [PubMed]
- Neco, P.; Fernandez-Peruchena, C.; Navas, S.; Gutierrez, L.M.; de Toledo, G.A.; Ales, E. Myosin II contributes to fusion pore expansion during exocytosis. J. Biol. Chem. 2008, 283, 10949–10957. [Google Scholar] [CrossRef] [PubMed]
- Papadopulos, A.; Gomez, G.A.; Martin, S.; Jackson, J.; Gormal, R.S.; Keating, D.J.; Yap, A.S.; Meunier, F.A. Activity-driven relaxation of the cortical actomyosin II network synchronizes Munc18-1-dependent neurosecretory vesicle docking. Nat. Commun. 2015, 6, 6297. [Google Scholar] [CrossRef]
- Tomatis, V.M.; Josh, P.; Papadopulos, A.; Gormal, R.S.; Lanoue, V.; Martin, S.; Meunier, F.A. ENA/VASP proteins regulate exocytosis by mediating myosin VI-dependent recruitment of secretory granules to the cortical actin network. Mol. Cell. Neurosci. 2017, 84, 100–111. [Google Scholar] [CrossRef]
- Gasman, S.; Vitale, N. Lipid remodelling in neuroendocrine secretion. Biol. Cell 2017. [Google Scholar] [CrossRef]
- Momboisse, F.; Lonchamp, E.; Calco, V.; Ceridono, M.; Vitale, N.; Bader, M.F.; Gasman, S. betaPIX-activated Rac1 stimulates the activation of phospholipase D, which is associated with exocytosis in neuroendocrine cells. J. Cell. Sci. 2009, 122, 798–806. [Google Scholar] [CrossRef]
- Zeniou-Meyer, M.; Zabari, N.; Ashery, U.; Chasserot-Golaz, S.; Haeberle, A.M.; Demais, V.; Bailly, Y.; Gottfried, I.; Nakanishi, H.; Neiman, A.M.; et al. Phospholipase D1 production of phosphatidic acid at the plasma membrane promotes exocytosis of large dense-core granules at a late stage. J. Biol. Chem. 2007, 282, 21746–21757. [Google Scholar] [CrossRef]
- Hughes, W.E.; Elgundi, Z.; Huang, P.; Frohman, M.A.; Biden, T.J. Phospholipase D1 regulates secretagogue-stimulated insulin release in pancreatic beta-cells. J. Biol. Chem. 2004, 279, 27534–27541. [Google Scholar] [CrossRef]
- Metz, S.A.; Dunlop, M. Stimulation of insulin release by phospholipase D. A potential role for endogenous phosphatidic acid in pancreatic islet function. Biochem. J. 1990, 270, 427–435. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Cambronero, J. Phospholipase D in cell signaling: From a myriad of cell functions to cancer growth and metastasis. J. Biol. Chem. 2014, 289, 22557–22566. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Frohman, M.A. Cellular and physiological roles for phospholipase D1 in cancer. J. Biol. Chem. 2014, 289, 22567–22574. [Google Scholar] [CrossRef] [PubMed]
- Ursino, G.M.; Fu, Y.; Cottle, D.L.; Mukhamedova, N.; Jones, L.K.; Low, H.; Tham, M.S.; Gan, W.J.; Mellett, N.A.; Das, P.P.; et al. ABCA12 regulates insulin secretion from beta-cells. EMBO Rep. 2020, 21, e48692. [Google Scholar] [CrossRef]
- Gong, L.W.; Di Paolo, G.; Diaz, E.; Cestra, G.; Diaz, M.E.; Lindau, M.; De Camilli, P.; Toomre, D. Phosphatidylinositol phosphate kinase type I gamma regulates dynamics of large dense-core vesicle fusion. Proc. Natl. Acad. Sci. USA 2005, 102, 5204–5209. [Google Scholar] [CrossRef]
- Milosevic, I.; Sorensen, J.B.; Lang, T.; Krauss, M.; Nagy, G.; Haucke, V.; Jahn, R.; Neher, E. Plasmalemmal phosphatidylinositol-4,5-bisphosphate level regulates the releasable vesicle pool size in chromaffin cells. J. Neurosci. 2005, 25, 2557–2565. [Google Scholar] [CrossRef]
- Waselle, L.; Gerona, R.R.; Vitale, N.; Martin, T.F.; Bader, M.F.; Regazzi, R. Role of phosphoinositide signaling in the control of insulin exocytosis. Mol. Endocrinol. 2005, 19, 3097–3106. [Google Scholar] [CrossRef]
- Gubar, O.; Morderer, D.; Tsyba, L.; Croise, P.; Houy, S.; Ory, S.; Gasman, S.; Rynditch, A. Intersectin: The Crossroad between Vesicle Exocytosis and Endocytosis. Front. Endocrinol. (Lausanne) 2013, 4, 109. [Google Scholar] [CrossRef]
- Malacombe, M.; Ceridono, M.; Calco, V.; Chasserot-Golaz, S.; McPherson, P.S.; Bader, M.F.; Gasman, S. Intersectin-1L nucleotide exchange factor regulates secretory granule exocytosis by activating Cdc42. Embo. J. 2006, 25, 3494–3503. [Google Scholar] [CrossRef]
- Zheng, Y. Dbl family guanine nucleotide exchange factors. Trends Biochem. Sci. 2001, 26, 724–732. [Google Scholar] [CrossRef]
- Audebert, S.; Navarro, C.; Nourry, C.; Chasserot-Golaz, S.; Lecine, P.; Bellaiche, Y.; Dupont, J.L.; Premont, R.T.; Sempere, C.; Strub, J.M.; et al. Mammalian Scribble Forms a Tight Complex with the betaPIX Exchange Factor. Curr. Biol. 2004, 14, 987–995. [Google Scholar] [CrossRef]
- Zhou, W.; Li, X.; Premont, R.T. Expanding functions of GIT Arf GTPase-activating proteins, PIX Rho guanine nucleotide exchange factors and GIT-PIX complexes. J. Cell Sci. 2016, 129, 1963–1974. [Google Scholar] [CrossRef] [PubMed]
- Dufurrena, Q.; Back, N.; Mains, R.E.; Hodgson, L.; Tanowitz, H.; Mandela, P.; Eipper, E.; Kuliawat, R. Kalirin/Trio Rho GDP/GTP exchange factors regulate proinsulin and insulin secretion. J. Mol. Endocrinol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Kowluru, A. GPCRs, G Proteins, and Their Impact on beta-cell Function. Compr. Physiol. 2020, 10, 453–490. [Google Scholar] [CrossRef] [PubMed]
- Veluthakal, R.; Madathilparambil, S.V.; McDonald, P.; Olson, L.K.; Kowluru, A. Regulatory roles for Tiam1, a guanine nucleotide exchange factor for Rac1, in glucose-stimulated insulin secretion in pancreatic beta-cells. Biochem. Pharmacol. 2009, 77, 101–113. [Google Scholar] [CrossRef] [PubMed]
- Veluthakal, R.; Tunduguru, R.; Arora, D.K.; Sidarala, V.; Syeda, K.; Vlaar, C.P.; Thurmond, D.C.; Kowluru, A. VAV2, a guanine nucleotide exchange factor for Rac1, regulates glucose-stimulated insulin secretion in pancreatic beta cells. Diabetologia 2015, 58, 2573–2581. [Google Scholar] [CrossRef] [PubMed]
- Xin, X.; Ferraro, F.; Back, N.; Eipper, B.A.; Mains, R.E. Cdk5 and Trio modulate endocrine cell exocytosis. J. Cell Sci. 2004, 117, 4739–4748. [Google Scholar] [CrossRef] [PubMed]
- Khelfaoui, M.; Gambino, F.; Houbaert, X.; Ragazzon, B.; Muller, C.; Carta, M.; Lanore, F.; Srikumar, B.N.; Gastrein, P.; Lepleux, M.; et al. Lack of the presynaptic RhoGAP protein oligophrenin1 leads to cognitive disabilities through dysregulation of the cAMP/PKA signalling pathway. Philos. Trans. R Soc. Lond. B Biol. Sci. 2014, 369, 20130160. [Google Scholar] [CrossRef]
- Khelfaoui, M.; Pavlowsky, A.; Powell, A.D.; Valnegri, P.; Cheong, K.W.; Blandin, Y.; Passafaro, M.; Jefferys, J.G.; Chelly, J.; Billuart, P. Inhibition of RhoA pathway rescues the endocytosis defects in Oligophrenin1 mouse model of mental retardation. Hum. Mol. Genet. 2009, 18, 2575–2583. [Google Scholar] [CrossRef]
- Nadif Kasri, N.; Nakano-Kobayashi, A.; Malinow, R.; Li, B.; Van Aelst, L. The Rho-linked mental retardation protein oligophrenin-1 controls synapse maturation and plasticity by stabilizing AMPA receptors. Genes Dev. 2009, 23, 1289–1302. [Google Scholar] [CrossRef]
- Nakano-Kobayashi, A.; Kasri, N.N.; Newey, S.E.; Van Aelst, L. The Rho-linked mental retardation protein OPHN1 controls synaptic vesicle endocytosis via endophilin A1. Curr. Biol. 2009, 19, 1133–1139. [Google Scholar] [CrossRef] [PubMed]
- Powell, A.D.; Gill, K.K.; Saintot, P.P.; Jiruska, P.; Chelly, J.; Billuart, P.; Jefferys, J.G. Rapid reversal of impaired inhibitory and excitatory transmission but not spine dysgenesis in a mouse model of mental retardation. J. Physiol. 2012, 590, 763–776. [Google Scholar] [CrossRef] [PubMed]
- Houy, S.; Estay-Ahumada, C.; Croise, P.; Calco, V.; Haeberle, A.M.; Bailly, Y.; Billuart, P.; Vitale, N.; Bader, M.F.; Ory, S.; et al. Oligophrenin-1 Connects Exocytotic Fusion to Compensatory Endocytosis in Neuroendocrine Cells. J. Neurosci. 2015, 35, 11045–11055. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; O’Connor, K.L.; Hellmich, M.R.; Greeley, G.H., Jr.; Townsend, C.M., Jr.; Evers, B.M. The role of protein kinase D in neurotensin secretion mediated by protein kinase C-alpha/-delta and Rho/Rho kinase. J. Biol. Chem. 2004, 279, 28466–28474. [Google Scholar] [CrossRef] [PubMed]
- Arias-Romero, L.E.; Chernoff, J. Targeting Cdc42 in cancer. Expert. Opin. Ther. Targets 2013, 17, 1263–1273. [Google Scholar] [CrossRef]
- Cardama, G.A.; Gonzalez, N.; Maggio, J.; Menna, P.L.; Gomez, D.E. Rho GTPases as therapeutic targets in cancer (Review). Int. J. Oncol. 2017, 51, 1025–1034. [Google Scholar] [CrossRef]
- Evelyn, C.R.; Ferng, T.; Rojas, R.J.; Larsen, M.J.; Sondek, J.; Neubig, R.R. High-throughput screening for small-molecule inhibitors of LARG-stimulated RhoA nucleotide binding via a novel fluorescence polarization assay. J. Biomol. Screen 2009, 14, 161–172. [Google Scholar] [CrossRef]
- Friesland, A.; Zhao, Y.; Chen, Y.H.; Wang, L.; Zhou, H.; Lu, Q. Small molecule targeting Cdc42-intersectin interaction disrupts Golgi organization and suppresses cell motility. Proc. Natl. Acad. Sci. USA 2013, 110, 1261–1266. [Google Scholar] [CrossRef]
- Gao, Y.; Dickerson, J.B.; Guo, F.; Zheng, J.; Zheng, Y. Rational design and characterization of a Rac GTPase-specific small molecule inhibitor. Proc. Natl. Acad. Sci. USA 2004, 101, 7618–7623. [Google Scholar] [CrossRef]
- Lee, M.H.; Kundu, J.K.; Chae, J.I.; Shim, J.H. Targeting ROCK/LIMK/cofilin signaling pathway in cancer. Arch. Pharm. Res. 2019, 42, 481–491. [Google Scholar] [CrossRef]
- Lin, Y.; Zheng, Y. Approaches of targeting Rho GTPases in cancer drug discovery. Expert Opin. Drug Discov. 2015, 10, 991–1010. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Du, W.; Shang, X.; Wang, L.; Evelyn, C.; Florian, M.C.; Ryan, M.A.; Rayes, A.; Zhao, X.; Setchell, K.; et al. Rational identification of a Cdc42 inhibitor presents a new regimen for long-term hematopoietic stem cell mobilization. Leukemia 2019, 33, 749–761. [Google Scholar] [CrossRef] [PubMed]
- Mardilovich, K.; Olson, M.F.; Baugh, M. Targeting Rho GTPase signaling for cancer therapy. Future Oncol. 2012, 8, 165–177. [Google Scholar] [CrossRef]
- Montalvo-Ortiz, B.L.; Castillo-Pichardo, L.; Hernandez, E.; Humphries-Bickley, T.; De la Mota-Peynado, A.; Cubano, L.A.; Vlaar, C.P.; Dharmawardhane, S. Characterization of EHop-016, novel small molecule inhibitor of Rac GTPase. J. Biol. Chem. 2012, 287, 13228–13238. [Google Scholar] [CrossRef]
- Nassar, N.; Cancelas, J.; Zheng, J.; Williams, D.A.; Zheng, Y. Structure-function based design of small molecule inhibitors targeting Rho family GTPases. Curr. Top. Med. Chem. 2006, 6, 1109–1116. [Google Scholar] [CrossRef] [PubMed]
- Pajic, M.; Herrmann, D.; Vennin, C.; Conway, J.R.; Chin, V.T.; Johnsson, A.K.; Welch, H.C.; Timpson, P. The dynamics of Rho GTPase signaling and implications for targeting cancer and the tumor microenvironment. Small GTPases 2015, 6, 123–133. [Google Scholar] [CrossRef]
- Shang, X.; Marchioni, F.; Sipes, N.; Evelyn, C.R.; Jerabek-Willemsen, M.; Duhr, S.; Seibel, W.; Wortman, M.; Zheng, Y. Rational design of small molecule inhibitors targeting RhoA subfamily Rho GTPases. Chem. Biol. 2012, 19, 699–710. [Google Scholar] [CrossRef]
- Xiao, X.H.; Lv, L.C.; Duan, J.; Wu, Y.M.; He, S.J.; Hu, Z.Z.; Xiong, L.X. Regulating Cdc42 and Its Signaling Pathways in Cancer: Small Molecules and MicroRNA as New Treatment Candidates. Molecules 2018, 23, 787. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, J.; Lai, X.N.; Jiao, X.Q.; Xiong, J.P.; Xiong, L.X. Focus on Cdc42 in Breast Cancer: New Insights, Target Therapy Development and Non-Coding RNAs. Cells 2019, 8, 146. [Google Scholar] [CrossRef]
- Rath, N.; Olson, M.F. Rho-associated kinases in tumorigenesis: Re-considering ROCK inhibition for cancer therapy. EMBO Rep. 2012, 13, 900–908. [Google Scholar] [CrossRef]
- Wang, Y.; Lei, R.; Zhuang, X.; Zhang, N.; Pan, H.; Li, G.; Hu, J.; Pan, X.; Tao, Q.; Fu, D.; et al. DLC1-dependent parathyroid hormone-like hormone inhibition suppresses breast cancer bone metastasis. J. Clin. Investig. 2014, 124, 1646–1659. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Gene/Tumor | ACC | GCC-AC | MCC (−) | NBL | PLCNEC | PNET | PPGL | PTC | SCLC | SINET | References |
|---|---|---|---|---|---|---|---|---|---|---|---|
| RHOA | Y42C | E40K | [30,31] | ||||||||
| D49H | |||||||||||
| RHOB | D28H | [32] | |||||||||
| RHOD | R174Q | R110Q | E149K | [32,33,34,35] | |||||||
| A165T | A173P | ||||||||||
| RHOF | K112 (fs) | [32] | |||||||||
| RHOG | P139L | [32,34] | |||||||||
| RHOH | A76E | P110A | S90 * | [30,32,36] | |||||||
| RHOJ | D8N | V25V | [32,37] | ||||||||
| RHOU | S249Y | [32] | |||||||||
| RND2 | E168K | R169S | [32] | ||||||||
| R137T | |||||||||||
| RAC1 | P29S | P92P | [36,38] | ||||||||
| RAC2 | R68W | [38] | |||||||||
| CDC42 | K184N | [30] | |||||||||
| RHOBTB1 | R670 * | P366S | A575C (fs) | [32,38,39] | |||||||
| RHOBTB2 | Q12H | D461G | [32,39] | ||||||||
| RHOBTB3 | R607H | S50T | [32,35] |
| GEFs/GAPs | Protein/Gene | Expression Variation | Tumors with Expression Modifications | Preferential Targets of the GEFs and GAPs | References |
|---|---|---|---|---|---|
| GEFs | ARHGEF1 | PCC vs. non-tumor | RhoA | [57,58] | |
| ARHGEF10L | NBL MYCN− vs. MYCN+ short survivors (gene) | RhoA, B, C | [48] | ||
| ARHGEF17 | NFPA vs. non-tumor | RhoA | [44] | ||
| FARP1 | PCC vs. non-tumor | Rac1 | [57,58] | ||
| FGD4 | PNET grade 2, 3 vs. 1 | Cdc42 | [61] | ||
| NEPC (gene) | [60] | ||||
| RCC2 | SCLC | Rac1 | [54] | ||
| VAV1 | uSCLC | RhoA, Rac1 | [62] | ||
| SCLC cell lines vs. non-SCLC cell lines | [63] | ||||
| VAV3 | CRPC-NEPC | RhoA, RhoG, Rac1 | [64] | ||
| GAPs | ARHGAP6 | PHPT vs. non-tumor | RhoA | [65] | |
| NEPC (gene) | [66] | ||||
| ARHGAP11A | NEPC (gene) | RhoA | [66] | ||
| ARHGAP11B | NEPC (gene) | RhoA, Cdc42 | [66] | ||
| ARHGAP18 | NFPA vs. non-tumor | RhoA, B, C | [44] | ||
| PCC (miRNA) | [59] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Streit, L.; Brunaud, L.; Vitale, N.; Ory, S.; Gasman, S. Hormones Secretion and Rho GTPases in Neuroendocrine Tumors. Cancers 2020, 12, 1859. https://doi.org/10.3390/cancers12071859
Streit L, Brunaud L, Vitale N, Ory S, Gasman S. Hormones Secretion and Rho GTPases in Neuroendocrine Tumors. Cancers. 2020; 12(7):1859. https://doi.org/10.3390/cancers12071859
Chicago/Turabian StyleStreit, Laura, Laurent Brunaud, Nicolas Vitale, Stéphane Ory, and Stéphane Gasman. 2020. "Hormones Secretion and Rho GTPases in Neuroendocrine Tumors" Cancers 12, no. 7: 1859. https://doi.org/10.3390/cancers12071859
APA StyleStreit, L., Brunaud, L., Vitale, N., Ory, S., & Gasman, S. (2020). Hormones Secretion and Rho GTPases in Neuroendocrine Tumors. Cancers, 12(7), 1859. https://doi.org/10.3390/cancers12071859