Microfluidic and Microscale Assays to Examine Regenerative Strategies in the Neuro Retina


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
1.1. Characteristics of Microfluidics
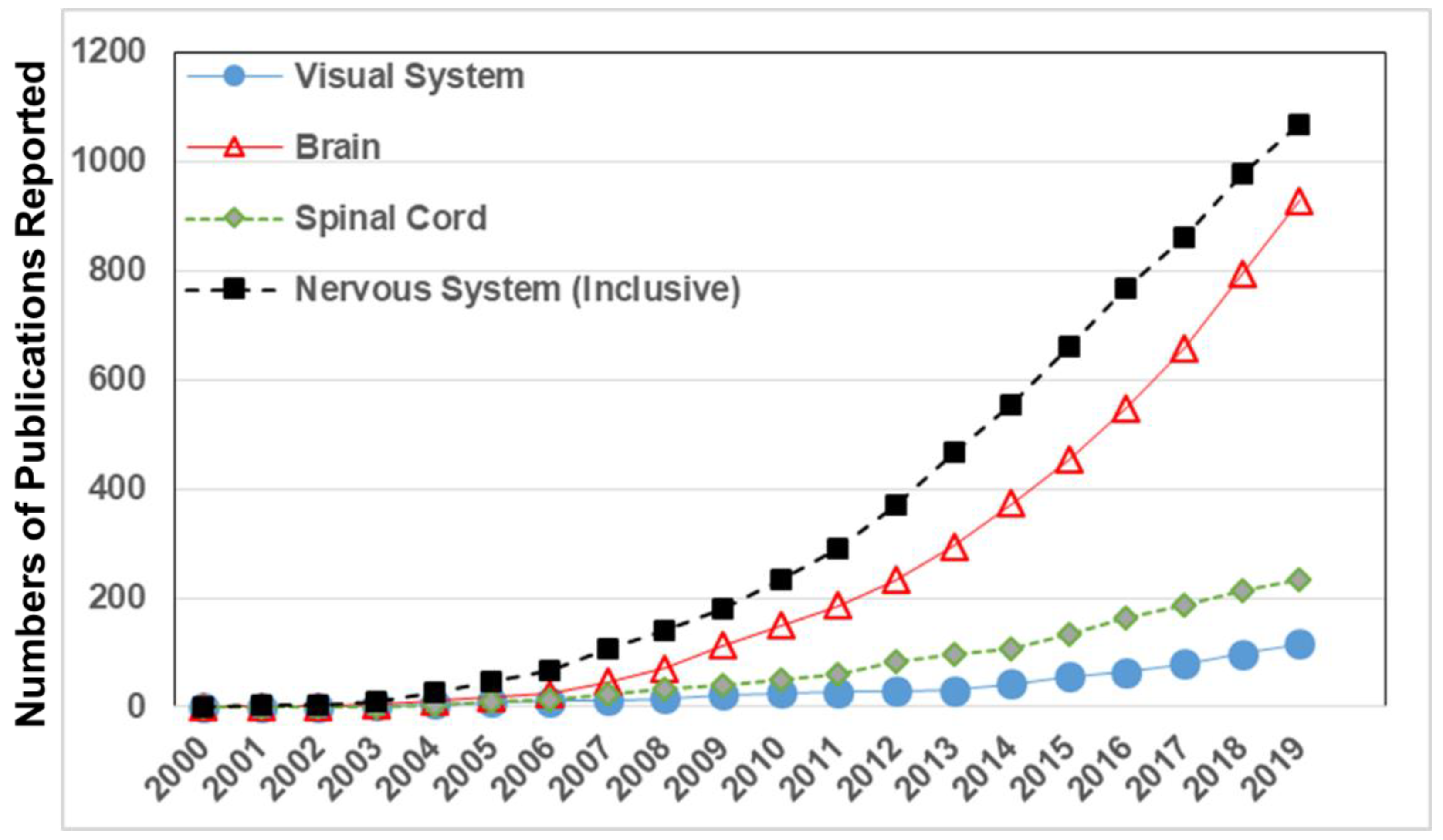
1.2. Applications in the Nervous System
1.3. Significance to the Visual System
1.4. Roles for Microfluidics in Neuro-Regenerative Medicine
1.5. Scope
2. The Visual System
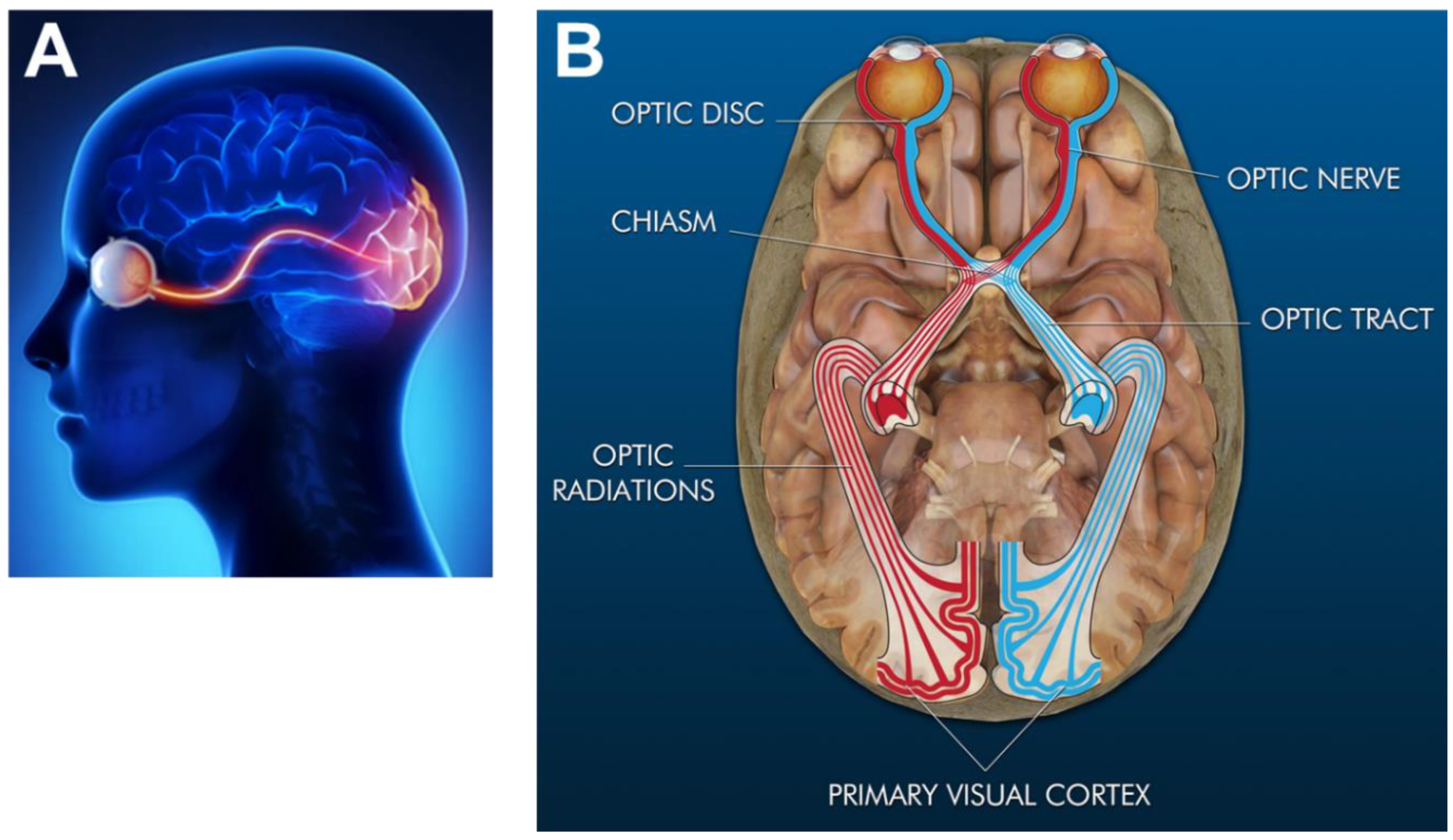
2.1. Anatomy and Light Path
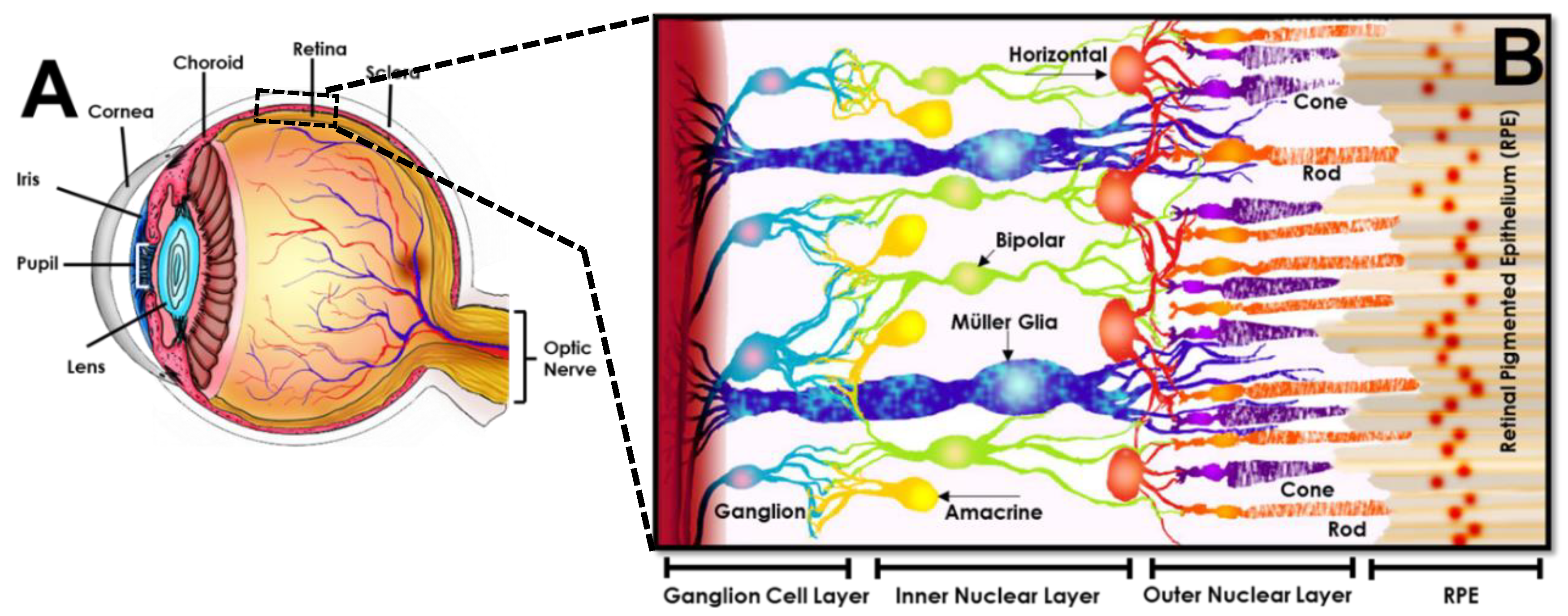
2.2. Retinal Tissue
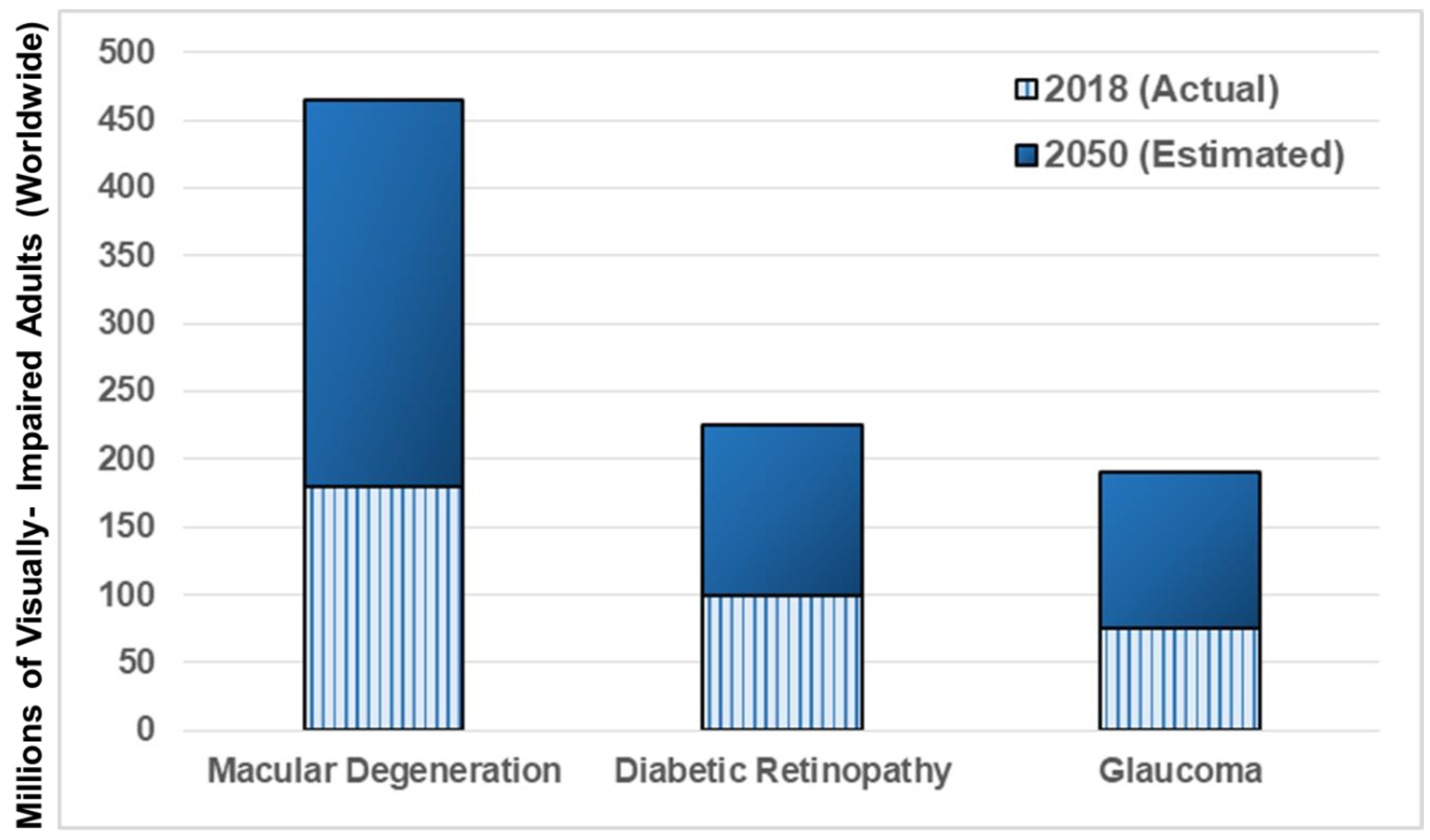
2.3. Visual Impairment and Loss of Vision
2.4. Opportunities for Microfluidics in Retinal Regeneration
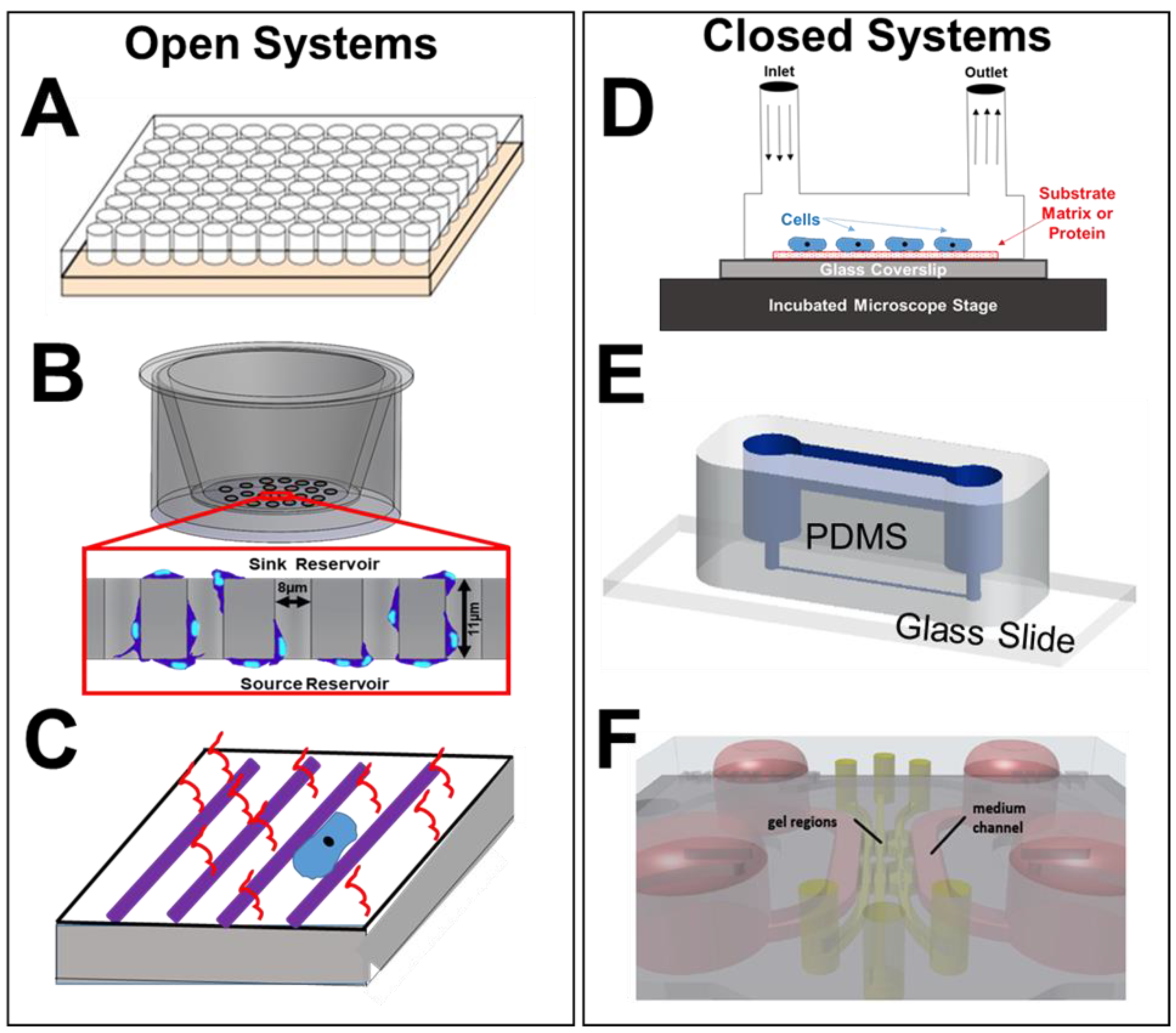
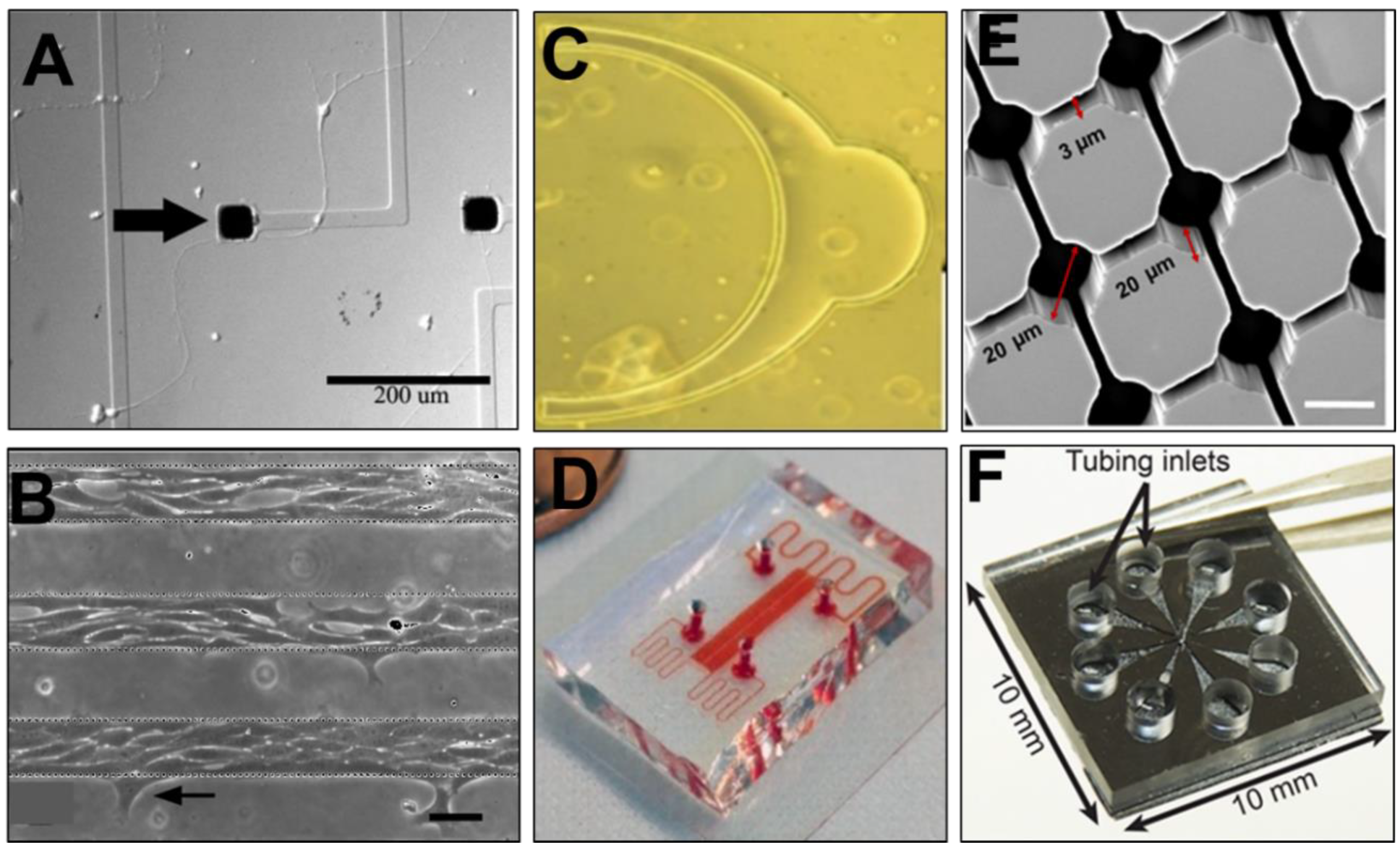
3. Microfluidic Assays and Microscale Systems
3.1. Microscale Cell Culture Chambers
3.2. Transwell Assays
3.3. Micropatterned Substrates
3.4. Microfluidic Channels
3.5. Microfluidic Perfusion Chambers
3.6. On-a-Chip Devices

4. Modeling of Retinal Cell Behaviors
4.1. Survival and Viability
4.2. Morphology and Cell Growth
4.3. Migration and Modalities
4.4. Apoptosis and Cell Death
4.5. Neuronal Connectivity
4.6. Neuronal-Glia Communication

5. Study of Developmental Processes
5.1. Cone Growth and Guidance
5.2. Regulation of Cell Fate
5.3. Cell Alignment and Positioning
5.4. Stem Cell Delivery
6. Future Directions
6.1. Macular Models
6.2. Retinal Blood Barriers
6.3. Hybrid Micro-Physiological Systems
6.4. Microfluidic Testing Models
Funding
Conflicts of Interest
Abbreviations
| AMD | Age-related Macular Degeneration (wet and dry) |
| CNTF | Ciliary Neurotrophic Factor |
| CTGF | Connective Tissue Growth Factor |
| Cx43 | Gap junction protein Connexin 43 |
| EGF | Epidermal Growth Factor |
| ESCs | Embryonic Stem Cells (mouse) |
| FGF | Fibroblast Growth Factor |
| GCL | Ganglion Cell Layer of Retina |
| GDNF | Glia-Derived Neurotrophic Factor |
| IGF | Insulin-like Growth Factor |
| INL | Inner Nuclear Layer of Retina |
| iPSCs | Induced Pluripotent Stem Cells (human) |
| I/R | Ischemia-Reperfusion Injury Model |
| MPS | Micro-Physiological Systems (or organoids) |
| MSCs | Mesenchymal Stem Cells |
| NrF2 | Nuclear Factor Erythroid 2-related factor 2 |
| NS | Nervous System (Central and Peripheral) |
| ONL | Outer Nuclear Layer of Retina |
| PDMS | Poly (dimethyl) siloxane |
| PCL | Polycaprolactone |
| PDGF | Platelet-Derived Growth Factor |
| PEDF | Pigment Epithelium-Derived Factor |
| PEM | Polyelectrolyte Multilayer |
| PLGA | Poly (lactic-co-glycolic acid) |
| PMMA | Poly (methyl-methacrylate) |
| SDF-1 | Stromal-Derived Growth Factor |
| TGF-b | Transforming Growth Factor Beta |
| VEGF | Vascular Endothelial Growth Factor |
| WHO | World Health Organization |
References
- Duncombe, T.A.; Tentori, A.M.; Herr, A.E. Microfluidics: Reframing biological enquiry. Nat. Rev. Mol. Cell Biol. 2015, 16, 554–567. [Google Scholar] [CrossRef] [PubMed]
- Mu, X.; Zheng, W.; Sun, J.; Zhang, W.; Jiang, X. Microfluidics for manipulating cells. Small 2013, 9, 9–21. [Google Scholar] [CrossRef]
- Bai, Y.; Gao, M.; Wen, L.; He, C.; Chen, Y.; Liu, C.; Fu, X.; Huang, S. Applications of Microfluidics in Quantitative Biology. Biotechnol. J. 2018, 13, e1700170. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Han, X.; Qin, L. Recent Progress of Microfluidics in Translational Applications. Adv. Healthc. Mater. 2016, 5, 871–888. [Google Scholar] [CrossRef]
- Sonnen, K.F.; Merten, C.A. Microfluidics as an Emerging Precision Tool in Developmental Biology. Dev. Cell 2019, 48, 293–311. [Google Scholar] [CrossRef] [PubMed]
- Gupta, N.; Renugopalakrishnan, V.; Liepmann, D.; Paulmurugan, R.; Malhotra, B.D. Cell-based biosensors: Recent trends, challenges and future perspectives. Biosens. Bioelectron. 2019, 141, 111435. [Google Scholar] [CrossRef]
- Levario, T.J.; Lim, B.; Shvartsman, S.Y.; Lu, H. Microfluidics for High-Throughput Quantitative Studies of Early Development. Annu. Rev. Biomed. Eng. 2016, 18, 285–309. [Google Scholar] [CrossRef]
- Rutkowski, J.M.; Swartz, M.A. A driving force for change: Interstitial flow as a morphoregulator. Trends Cell Biol. 2007, 17, 44–50. [Google Scholar] [CrossRef]
- Mofazzal Jahromi, M.A.; Abdoli, A.; Rahmanian, M.; Bardania, H.; Bayandori, M.; Moosavi Basri, S.M.; Kalbasi, A.; Aref, A.R.; Karimi, M.; Hamblin, M.R. Microfluidic Brain-on-a-Chip: Perspectives for Mimicking Neural System Disorders. Mol. Neurobiol. 2019, 56, 8489–8512. [Google Scholar] [CrossRef]
- van der Helm, M.W.; van der Meer, A.D.; Eijkel, J.C.; van den Berg, A.; Segerink, L.I. Microfluidic organ-on-chip technology for blood-brain barrier research. Tissue Barriers 2016, 4, e1142493. [Google Scholar] [CrossRef]
- Samal, P.; van Blitterswijk, C.; Truckenmuller, R.; Giselbrecht, S. Grow with the Flow: When Morphogenesis Meets Microfluidics. Adv. Mater. 2019, 31, e1805764. [Google Scholar] [CrossRef]
- Khademhosseini, A.; Langer, R.; Borenstein, J.; Vacanti, J.P. Microscale technologies for tissue engineering and biology. Proc. Natl. Acad. Sci. USA 2006, 103, 2480–2487. [Google Scholar] [CrossRef] [PubMed]
- Taylor, A.M.; Jeon, N.L. Micro-scale and microfluidic devices for neurobiology. Curr. Opin. Neurobiol. 2010, 20, 640–647. [Google Scholar] [CrossRef] [PubMed]
- Araci, I.E.; Brisk, P. Recent developments in microfluidic large scale integration. Curr. Opin. Biotechnol. 2014, 25, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Ren, L.; Li, L.; Liu, W.; Zhou, J.; Yu, W.; Tong, D.; Chen, S. Microfluidics: A new cosset for neurobiology. Lab Chip 2009, 9, 644–652. [Google Scholar] [CrossRef] [PubMed]
- Taylor, A.M.; Jeon, N.L. Microfluidic and compartmentalized platforms for neurobiological research. Crit. Rev. Biomed. Eng. 2011, 39, 185–200. [Google Scholar] [CrossRef] [PubMed]
- Berthiaume, F.; Maguire, T.J.; Yarmush, M.L. Tissue engineering and regenerative medicine: History, progress, and challenges. Annu. Rev. Chem. Biomol. Eng. 2011, 2, 403–430. [Google Scholar] [CrossRef]
- Lanza, R. Regenerative medicine: The last 10 years. Regen. Med. 2016, 11, 745–746. [Google Scholar] [CrossRef]
- Wang, Q.; Stern, J.H.; Temple, S. Regenerative Medicine: Solution in Sight. Adv. Exp. Med. Biol. 2016, 854, 543–548. [Google Scholar] [CrossRef]
- Whitesides, G.M. The origins and the future of microfluidics. Nature 2006, 442, 368–373. [Google Scholar] [CrossRef]
- Kartalov, E.P.; Anderson, W.F.; Scherer, A. The analytical approach to polydimethylsiloxane microfluidic technology and its biological applications. J. Nanosci. Nanotechnol. 2006, 6, 2265–2277. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Liu, Y.; Qu, W.; Jiang, X. Combining nanosurface chemistry and microfluidics for molecular analysis and cell biology. Anal. Chim. Acta 2009, 650, 98–105. [Google Scholar] [CrossRef] [PubMed]
- Berthier, E.; Young, E.W.; Beebe, D. Engineers are from PDMS-land, Biologists are from Polystyrenia. Lab Chip 2012, 12, 1224–1237. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, P.F.; Ben Azouz, A.; Vazquez, M.; Liu, J.; Marczak, S.; Slouka, Z.; Chang, H.C.; Diamond, D.; Brabazon, D. Advances in three-dimensional rapid prototyping of microfluidic devices for biological applications. Biomicrofluidics 2014, 8, 052112. [Google Scholar] [CrossRef] [PubMed]
- Whitesides, G.M.; Ostuni, E.; Takayama, S.; Jiang, X.; Ingber, D.E. Soft lithography in biology and biochemistry. Annu. Rev. Biomed. Eng. 2001, 3, 335–373. [Google Scholar] [CrossRef] [PubMed]
- Fallahi, H.; Zhang, J.; Phan, H.P.; Nguyen, N.T. Flexible Microfluidics: Fundamentals, Recent Developments, and Applications. Micromachines 2019, 10, 830. [Google Scholar] [CrossRef]
- Novotny, J.; Foret, F. Fluid manipulation on the micro-scale: Basics of fluid behavior in microfluidics. J. Sep. Sci. 2017, 40, 383–394. [Google Scholar] [CrossRef]
- Sackmann, E.K.; Fulton, A.L.; Beebe, D.J. The present and future role of microfluidics in biomedical research. Nature 2014, 507, 181–189. [Google Scholar] [CrossRef]
- Aminian, M.; Bernardi, F.; Camassa, R.; Harris, D.M.; McLaughlin, R.M. The Diffusion of Passive Tracers in Laminar Shear Flow. J. Vis. Exp. 2018. [Google Scholar] [CrossRef]
- Islam, M.Z.; Tsui, Y.Y. Quasi-3D Modeling and Efficient Simulation of Laminar Flows in Microfluidic Devices. Sensors 2016, 16, 1639. [Google Scholar] [CrossRef]
- Kuczenski, B.; LeDuc, P.R.; Messner, W.C. Pressure-driven spatiotemporal control of the laminar flow interface in a microfluidic network. Lab Chip 2007, 7, 647–649. [Google Scholar] [CrossRef] [PubMed]
- Kaneda, S.; Fujii, T. Integrated microfluidic systems. Adv. Biochem. Eng. Biotechnol. 2010, 119, 179–194. [Google Scholar] [CrossRef]
- Walsh, E.J.; Feuerborn, A.; Wheeler, J.H.R.; Tan, A.N.; Durham, W.M.; Foster, K.R.; Cook, P.R. Microfluidics with fluid walls. Nat. Commun. 2017, 8, 816. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; He, X. Interfacial tension based on-chip extraction of microparticles confined in microfluidic Stokes flows. Appl. Phys. Lett. 2014, 105, 143704. [Google Scholar] [CrossRef] [PubMed]
- Steegmans, M.L.; Warmerdam, A.; Schroen, K.G.; Boom, R.M. Dynamic interfacial tension measurements with microfluidic Y-junctions. Langmuir ACS J. Surf. Colloids 2009, 25, 9751–9758. [Google Scholar] [CrossRef]
- Lim, H.; Jafry, A.T.; Lee, J. Fabrication, Flow Control, and Applications of Microfluidic Paper-Based Analytical Devices. Molecules 2019, 24, 2869. [Google Scholar] [CrossRef]
- Hassan, S.U.; Tariq, A.; Noreen, Z.; Donia, A.; Zaidi, S.Z.J.; Bokhari, H.; Zhang, X. Capillary-Driven Flow Microfluidics Combined with Smartphone Detection: An Emerging Tool for Point-of-Care Diagnostics. Diagnostics 2020, 10, 509. [Google Scholar] [CrossRef]
- Channon, R.B.; Nguyen, M.P.; Henry, C.S.; Dandy, D.S. Multilayered Microfluidic Paper-Based Devices: Characterization, Modeling, and Perspectives. Anal. Chem. 2019, 91, 8966–8972. [Google Scholar] [CrossRef] [PubMed]
- Slater, G.W.; Tessier, F.; Kopecka, K. The Electroosmotic Flow (EOF). Methods Mol. Biol. 2010, 583, 121–134. [Google Scholar] [CrossRef]
- Zhao, C.; Yang, C. Electrokinetics of non-Newtonian fluids: A review. Adv. Colloid Interface Sci. 2013, 201–202, 94–108. [Google Scholar] [CrossRef]
- Vukasinovic, J.; Cullen, D.K.; LaPlaca, M.C.; Glezer, A. A microperfused incubator for tissue mimetic 3D cultures. Biomed. Microdevices 2009, 11, 1155–1165. [Google Scholar] [CrossRef] [PubMed]
- Neeves, K.B.; Lo, C.T.; Foley, C.P.; Saltzman, W.M.; Olbricht, W.L. Fabrication and characterization of microfluidic probes for convection enhanced drug delivery. J. Control. Release Off. J. Control. Release Soc. 2006, 111, 252–262. [Google Scholar] [CrossRef] [PubMed]
- Asghari, M.; Cao, X.; Mateescu, B.; van Leeuwen, D.; Aslan, M.K.; Stavrakis, S.; de Mello, A.J. Oscillatory Viscoelastic Microfluidics for Efficient Focusing and Separation of Nanoscale Species. ACS Nano 2020, 14, 422–433. [Google Scholar] [CrossRef]
- Hopkins, C.C.; Haward, S.J.; Shen, A.Q. Purely Elastic Fluid-Structure Interactions in Microfluidics: Implications for Mucociliary Flows. Small 2020, 16, e1903872. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Hjort, K.; Wicher, G.; Fex Svenningsen, A. Microfluidic high viability neural cell separation using viscoelastically tuned hydrodynamic spreading. Biomed. Microdevices 2008, 10, 631–638. [Google Scholar] [CrossRef] [PubMed]
- Mitrovski, S.M.; Nuzzo, R.G. An electrochemically driven poly(dimethylsiloxane) microfluidic actuator: Oxygen sensing and programmable flows and pH gradients. Lab Chip 2005, 5, 634–645. [Google Scholar] [CrossRef]
- Lyu, C.; Wang, J.; Rubinsky, B. Non-electrolytic microelectroporation. Biomed. Microdevices 2017, 19, 65. [Google Scholar] [CrossRef]
- Chang, H.; Khan, R.; Rong, Z.; Sapelkin, A.; Vadgama, P. Study of albumin and fibrinogen membranes formed by interfacial crosslinking using microfluidic flow. Biofabrication 2010, 2, 035002. [Google Scholar] [CrossRef]
- Harrison, H.; Lu, X.; Patel, S.; Thomas, C.; Todd, A.; Johnson, M.; Raval, Y.; Tzeng, T.R.; Song, Y.; Wang, J.; et al. Electrokinetic preconcentration of particles and cells in microfluidic reservoirs. Analyst 2015, 140, 2869–2875. [Google Scholar] [CrossRef]
- Huh, D.; Bahng, J.H.; Ling, Y.; Wei, H.H.; Kripfgans, O.D.; Fowlkes, J.B.; Grotberg, J.B.; Takayama, S. Gravity-driven microfluidic particle sorting device with hydrodynamic separation amplification. Anal. Chem. 2007, 79, 1369–1376. [Google Scholar] [CrossRef]
- Lu, Y.; Chan, Y.K.; Lau, L.H.; Chao, Y.; Shih, K.C.; Lai, S.M.; Wong, D.; Shum, H.C. Adhesion of silicone oil and emulsification: An in vitro assessment using a microfluidic device and ‘Eye-on-a-Chip’. Acta Ophthalmol. 2019, 97, 313–318. [Google Scholar] [CrossRef]
- Sakai, S.; Kawabata, K.; Ono, T.; Ijima, H.; Kawakami, K. Higher viscous solution induces smaller droplets for cell-enclosing capsules in a co-flowing stream. Biotechnol. Prog. 2005, 21, 994–997. [Google Scholar] [CrossRef]
- Sarkar, A.; Hou, H.W.; Mahan, A.E.; Han, J.; Alter, G. Multiplexed Affinity-Based Separation of Proteins and Cells Using Inertial Microfluidics. Sci. Rep. 2016, 6, 23589. [Google Scholar] [CrossRef] [PubMed]
- Mashimo, Y.; Yoshioka, M.; Tokunaga, Y.; Fockenberg, C.; Terada, S.; Koyama, Y.; Shibata-Seki, T.; Yoshimoto, K.; Sakai, R.; Hakariya, H.; et al. Fabrication of a Multiplexed Artificial Cellular MicroEnvironment Array. J. Vis. Exp. 2018. [Google Scholar] [CrossRef] [PubMed]
- Xiang, N.; Zhang, R.; Han, Y.; Ni, Z. A Multilayer Polymer-Film Inertial Microfluidic Device for High-Throughput Cell Concentration. Anal. Chem. 2019, 91, 5461–5468. [Google Scholar] [CrossRef] [PubMed]
- Vensi Basso, J.M.; Yurchenko, I.; Simon, M.; Rizzo, D.J.; Staii, C. Role of geometrical cues in neuronal growth. Phys. Rev. E 2019, 99, 022408. [Google Scholar] [CrossRef] [PubMed]
- Hardelauf, H.; Waide, S.; Sisnaiske, J.; Jacob, P.; Hausherr, V.; Schobel, N.; Janasek, D.; van Thriel, C.; West, J. Micropatterning neuronal networks. Analyst 2014, 139, 3256–3264. [Google Scholar] [CrossRef]
- Beduer, A.; Vieu, C.; Arnauduc, F.; Sol, J.C.; Loubinoux, I.; Vaysse, L. Engineering of adult human neural stem cells differentiation through surface micropatterning. Biomaterials 2012, 33, 504–514. [Google Scholar] [CrossRef]
- Reyes, D.R.; Perruccio, E.M.; Becerra, S.P.; Locascio, L.E.; Gaitan, M. Micropatterning neuronal cells on polyelectrolyte multilayers. Langmuir ACS J. Surf. Colloids 2004, 20, 8805–8811. [Google Scholar] [CrossRef]
- van de Wijdeven, R.; Ramstad, O.H.; Bauer, U.S.; Halaas, O.; Sandvig, A.; Sandvig, I. Structuring a multi-nodal neural network in vitro within a novel design microfluidic chip. Biomed. Microdevices 2018, 20, 9. [Google Scholar] [CrossRef]
- Li, Y.C.; Jodat, Y.A.; Samanipour, R.; Zorzi, G.; Zhu, K.; Hirano, M.; Chang, K.; Arnaout, A.; Hassan, S.; Matharu, N.; et al. Toward a neurospheroid niche model: Optimizing embedded 3D bioprinting for fabrication of neurospheroid brain-like co-culture constructs. Biofabrication 2020. [Google Scholar] [CrossRef] [PubMed]
- Nayagam, B.A.; Muniak, M.A.; Ryugo, D.K. The spiral ganglion: Connecting the peripheral and central auditory systems. Hear. Res. 2011, 278, 2–20. [Google Scholar] [CrossRef]
- Durante, M.A.; Kurtenbach, S.; Sargi, Z.B.; Harbour, J.W.; Choi, R.; Kurtenbach, S.; Goss, G.M.; Matsunami, H.; Goldstein, B.J. Single-cell analysis of olfactory neurogenesis and differentiation in adult humans. Nat. Neurosci. 2020, 23, 323–326. [Google Scholar] [CrossRef] [PubMed]
- Dewan, A.; Cichy, A.; Zhang, J.; Miguel, K.; Feinstein, P.; Rinberg, D.; Bozza, T. Single olfactory receptors set odor detection thresholds. Nat. Commun. 2018, 9, 2887. [Google Scholar] [CrossRef] [PubMed]
- Chaudhari, N.; Roper, S.D. The cell biology of taste. J. Cell Biol. 2010, 190, 285–296. [Google Scholar] [CrossRef]
- Barlow, L.A. Progress and renewal in gustation: New insights into taste bud development. Development 2015, 142, 3620–3629. [Google Scholar] [CrossRef] [PubMed]
- Holmes, N.P.; Tame, L.; Beeching, P.; Medford, M.; Rakova, M.; Stuart, A.; Zeni, S. Locating primary somatosensory cortex in human brain stimulation studies: Experimental evidence. J. Neurophysiol. 2019, 121, 336–344. [Google Scholar] [CrossRef]
- Yin, P.Y.; Hao, M.Z.; Liu, X.D.; Cao, C.Y.; Niu, C.M.; Lan, N. Neural Correlation between Evoked Tactile Sensation and Central Activities in the Somatosensory Cortex. Annu. Int. Conf. IEEE Eng. Med. Biol. Soc. 2018, 2018, 2296–2299. [Google Scholar] [CrossRef]
- Miesfeld, J.B.; Brown, N.L. Eye organogenesis: A hierarchical view of ocular development. Curr. Top. Dev. Biol. 2019, 132, 351–393. [Google Scholar] [CrossRef]
- Atri, A. The Alzheimer’s Disease Clinical Spectrum: Diagnosis and Management. Med. Clin. N. Am. 2019, 103, 263–293. [Google Scholar] [CrossRef]
- Karam, C.Y.; Paganoni, S.; Joyce, N.; Carter, G.T.; Bedlack, R. Palliative Care Issues in Amyotrophic Lateral Sclerosis: An Evidenced-Based Review. Am. J. Hosp. Palliat. Care 2016, 33, 84–92. [Google Scholar] [CrossRef]
- Masel, B.E.; DeWitt, D.S. Traumatic brain injury: A disease process, not an event. J. Neurotrauma 2010, 27, 1529–1540. [Google Scholar] [CrossRef] [PubMed]
- Hatch, B.B.; Wood-Wentz, C.M.; Therneau, T.M.; Walker, M.G.; Payne, J.M.; Reeves, R.K. Factors predictive of survival and estimated years of life lost in the decade following nontraumatic and traumatic spinal cord injury. Spinal Cord 2017, 55, 540–544. [Google Scholar] [CrossRef] [PubMed]
- Flaxman, S.R.; Bourne, R.R.A.; Resnikoff, S.; Ackland, P.; Braithwaite, T.; Cicinelli, M.V.; Das, A.; Jonas, J.B.; Keeffe, J.; Kempen, J.H.; et al. Global causes of blindness and distance vision impairment 1990-2020: A systematic review and meta-analysis. Lancet Glob. Health 2017, 5, e1221–e1234. [Google Scholar] [CrossRef]
- Pelletier, A.L.; Rojas-Roldan, L.; Coffin, J. Vision Loss in Older Adults. Am. Fam. Physician 2016, 94, 219–226. [Google Scholar] [PubMed]
- Singh, R.; Cuzzani, O.; Binette, F.; Sternberg, H.; West, M.D.; Nasonkin, I.O. Pluripotent Stem Cells for Retinal Tissue Engineering: Current Status and Future Prospects. Stem Cell Rev. Rep. 2018, 14, 463–483. [Google Scholar] [CrossRef] [PubMed]
- Pena, J.S.; Vazquez, M. Reducing health disparities in adult vision loss via interfaces with emerging technology. Eye 2019, 33, 532–533. [Google Scholar] [CrossRef]
- Zarbin, M. Cell-Based Therapy for Degenerative Retinal Disease. Trends Mol. Med. 2016, 22, 115–134. [Google Scholar] [CrossRef]
- Sieving, P.A. Tissue engineering: NIH competition to create ‘eye in a dish’. Nature 2017, 546, 352. [Google Scholar] [CrossRef]
- Bourne, R.R.A.; Flaxman, S.R.; Braithwaite, T.; Cicinelli, M.V.; Das, A.; Jonas, J.B.; Keeffe, J.; Kempen, J.H.; Leasher, J.; Limburg, H.; et al. Magnitude, temporal trends, and projections of the global prevalence of blindness and distance and near vision impairment: A systematic review and meta-analysis. Lancet Glob. Health 2017, 5, e888–e897. [Google Scholar] [CrossRef]
- Leasher, J.L.; Braithwaite, T.; Furtado, J.M.; Flaxman, S.R.; Lansingh, V.C.; Silva, J.C.; Resnikoff, S.; Taylor, H.R.; Bourne, R.R.A. Prevalence and causes of vision loss in Latin America and the Caribbean in 2015: Magnitude, temporal trends and projections. Br. J. Ophthalmol. 2019, 103, 885–893. [Google Scholar] [CrossRef] [PubMed]
- Nangia, V.; Jonas, J.B.; George, R.; Lingam, V.; Ellwein, L.; Cicinelli, M.V.; Das, A.; Flaxman, S.R.; Keeffe, J.E.; Kempen, J.H.; et al. Prevalence and causes of blindness and vision impairment: Magnitude, temporal trends and projections in South and Central Asia. Br. J. Ophthalmol. 2019, 103, 871–877. [Google Scholar] [CrossRef] [PubMed]
- Bourne, R.R.A.; Jonas, J.B.; Bron, A.M.; Cicinelli, M.V.; Das, A.; Flaxman, S.R.; Friedman, D.S.; Keeffe, J.E.; Kempen, J.H.; Leasher, J.; et al. Prevalence and causes of vision loss in high-income countries and in Eastern and Central Europe in 2015: Magnitude, temporal trends and projections. Br. J. Ophthalmol. 2018, 102, 575–585. [Google Scholar] [CrossRef]
- Nemeth, J.; Toth, G.; Resnikoff, S.; de Faber, J.T. Preventing blindness and visual impairment in Europe: What do we have to do? Eur. J. Ophthalmol. 2019, 29, 129–132. [Google Scholar] [CrossRef] [PubMed]
- Zambelli-Weiner, A.; Crews, J.E.; Friedman, D.S. Disparities in adult vision health in the United States. Am. J. Ophthalmol. 2012, 154, S23–S30.e21. [Google Scholar] [CrossRef] [PubMed]
- Doyal, L.; Das-Bhaumik, R.G. Sex, gender and blindness: A new framework for equity. BMJ Open Ophthalmol. 2018, 3, e000135. [Google Scholar] [CrossRef]
- Mitchell, P.; Liew, G.; Gopinath, B.; Wong, T.Y. Age-related macular degeneration. Lancet 2018, 392, 1147–1159. [Google Scholar] [CrossRef]
- Desmettre, T.J. Epigenetics in Age-related Macular Degeneration (AMD). J. Fr. D’ophtalmologie 2018, 41, e407–e415. [Google Scholar] [CrossRef]
- Sabanayagam, C.; Banu, R.; Chee, M.L.; Lee, R.; Wang, Y.X.; Tan, G.; Jonas, J.B.; Lamoureux, E.L.; Cheng, C.Y.; Klein, B.E.K.; et al. Incidence and progression of diabetic retinopathy: A systematic review. Lancet Diabetes Endocrinol. 2019, 7, 140–149. [Google Scholar] [CrossRef]
- Ban, N.; Siegfried, C.J.; Apte, R.S. Monitoring Neurodegeneration in Glaucoma: Therapeutic Implications. Trends Mol. Med. 2018, 24, 7–17. [Google Scholar] [CrossRef]
- Li, W.; Liu, J.; Galvin, J.A. Epigenetics and Common Ophthalmic Diseases. Yale J. Biol. Med. 2016, 89, 597–600. [Google Scholar]
- Wang, T.; Pahlberg, J.; Cafaro, J.; Frederiksen, R.; Cooper, A.J.; Sampath, A.P.; Field, G.D.; Chen, J. Activation of Rod Input in a Model of Retinal Degeneration Reverses Retinal Remodeling and Induces Formation of Functional Synapses and Recovery of Visual Signaling in the Adult Retina. J. Neurosci. Off. J. Soc. Neurosci. 2019, 39, 6798–6810. [Google Scholar] [CrossRef]
- Fine, I.; Park, J.M. Blindness and Human Brain Plasticity. Annu. Rev. Vis. Sci. 2018, 4, 337–356. [Google Scholar] [CrossRef]
- Zhou, L.; Chen, W.; Lin, D.; Hu, W.; Tang, Z. Neuronal apoptosis, axon damage and synapse loss occur synchronously in acute ocular hypertension. Exp. Eye Res. 2019, 180, 77–85. [Google Scholar] [CrossRef]
- Garweg, J.G.; Tappeiner, C.; Halberstadt, M. Pathophysiology of proliferative vitreoretinopathy in retinal detachment. Surv. Ophthalmol. 2013, 58, 321–329. [Google Scholar] [CrossRef]
- Danesh-Meyer, H.V.; Yoon, J.J.; Lawlor, M.; Savino, P.J. Visual loss and recovery in chiasmal compression. Prog. Retin. Eye Res. 2019, 73, 100765. [Google Scholar] [CrossRef]
- Pinazo-Duran, M.D.; Gallego-Pinazo, R.; Garcia-Medina, J.J.; Zanon-Moreno, V.; Nucci, C.; Dolz-Marco, R.; Martinez-Castillo, S.; Galbis-Estrada, C.; Marco-Ramirez, C.; Lopez-Galvez, M.I.; et al. Oxidative stress and its downstream signaling in aging eyes. Clin. Interv. Aging 2014, 9, 637–652. [Google Scholar] [CrossRef]
- Bhatia-Dey, N.; Kanherkar, R.R.; Stair, S.E.; Makarev, E.O.; Csoka, A.B. Cellular Senescence as the Causal Nexus of Aging. Front. Genet. 2016, 7, 13. [Google Scholar] [CrossRef]
- Alavi, M.V. Aging and Vision. Adv. Exp. Med. Biol. 2016, 854, 393–399. [Google Scholar] [CrossRef]
- Brunello, C.A.; Jokinen, V.; Sakha, P.; Terazono, H.; Nomura, F.; Kaneko, T.; Lauri, S.E.; Franssila, S.; Rivera, C.; Yasuda, K.; et al. Microtechnologies to fuel neurobiological research with nanometer precision. J. Nanobiotechnol. 2013, 11, 11. [Google Scholar] [CrossRef]
- Millet, L.J.; Gillette, M.U. New perspectives on neuronal development via microfluidic environments. Trends Neurosci. 2012, 35, 752–761. [Google Scholar] [CrossRef] [PubMed]
- Chang, W.C.; Kliot, M.; Sretavan, D.W. Microtechnology and nanotechnology in nerve repair. Neurol. Res. 2008, 30, 1053–1062. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, J. Stem cells and regenerative medicine for neural repair. Curr. Opin. Biotechnol. 2018, 52, 102–108. [Google Scholar] [CrossRef]
- Gage, F.H.; Temple, S. Neural stem cells: Generating and regenerating the brain. Neuron 2013, 80, 588–601. [Google Scholar] [CrossRef]
- Aladdad, A.M.; Kador, K.E. Adult Stem Cells, Tools for Repairing the Retina. Curr. Ophthalmol. Rep. 2019, 7, 21–29. [Google Scholar] [CrossRef]
- Reichenbach, A.; Bringmann, A. New functions of Muller cells. Glia 2013, 61, 651–678. [Google Scholar] [CrossRef]
- Mead, B.; Logan, A.; Berry, M.; Leadbeater, W.; Scheven, B.A. Concise Review: Dental Pulp Stem Cells: A Novel Cell Therapy for Retinal and Central Nervous System Repair. Stem Cells 2017, 35, 61–67. [Google Scholar] [CrossRef]
- Tso, D.; McKinnon, R.D. Cell replacement therapy for central nervous system diseases. Neural Regen. Res. 2015, 10, 1356–1358. [Google Scholar] [CrossRef]
- Levin, L.A.; Miller, J.W.; Zack, D.J.; Friedlander, M.; Smith, L.E.H. Special Commentary: Early Clinical Development of Cell Replacement Therapy: Considerations for the National Eye Institute Audacious Goals Initiative. Ophthalmology 2017, 124, 926–934. [Google Scholar] [CrossRef]
- Zarbin, M. Cell-Based Therapy for Retinal Disease: The New Frontier. Methods Mol. Biol. 2019, 1834, 367–381. [Google Scholar] [CrossRef]
- MacLaren, R.E.; Pearson, R.A.; MacNeil, A.; Douglas, R.H.; Salt, T.E.; Akimoto, M.; Swaroop, A.; Sowden, J.C.; Ali, R.R. Retinal repair by transplantation of photoreceptor precursors. Nature 2006, 444, 203–207. [Google Scholar] [CrossRef] [PubMed]
- MacLaren, R.E.; Bennett, J.; Schwartz, S.D. Gene Therapy and Stem Cell Transplantation in Retinal Disease: The New Frontier. Ophthalmology 2016, 123, S98–S106. [Google Scholar] [CrossRef] [PubMed]
- Singh, M.S.; Park, S.S.; Albini, T.A.; Canto-Soler, M.V.; Klassen, H.; MacLaren, R.E.; Takahashi, M.; Nagiel, A.; Schwartz, S.D.; Bharti, K. Retinal stem cell transplantation: Balancing safety and potential. Prog. Retin. Eye Res. 2020, 75, 100779. [Google Scholar] [CrossRef]
- Jayakody, S.A.; Gonzalez-Cordero, A.; Ali, R.R.; Pearson, R.A. Cellular strategies for retinal repair by photoreceptor replacement. Prog. Retin. Eye Res. 2015, 46, 31–66. [Google Scholar] [CrossRef] [PubMed]
- Ho, M.T.; Teal, C.J.; Shoichet, M.S. A hyaluronan/methylcellulose-based hydrogel for local cell and biomolecule delivery to the central nervous system. Brain Res. Bull. 2019, 148, 46–54. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, R.; Zarembinski, T.I.; Doty, N.; Jiang, C.; Regatieri, C.; Zhang, X.; Young, M.J. The application of hyaluronic acid hydrogels to retinal progenitor cell transplantation. Tissue Eng. Part A 2013, 19, 135–142. [Google Scholar] [CrossRef]
- Ballios, B.G.; Cooke, M.J.; Donaldson, L.; Coles, B.L.; Morshead, C.M.; van der Kooy, D.; Shoichet, M.S. A Hyaluronan-Based Injectable Hydrogel Improves the Survival and Integration of Stem Cell Progeny following Transplantation. Stem Cell Rep. 2015, 4, 1031–1045. [Google Scholar] [CrossRef]
- Pearson, R.A.; Barber, A.C.; Rizzi, M.; Hippert, C.; Xue, T.; West, E.L.; Duran, Y.; Smith, A.J.; Chuang, J.Z.; Azam, S.A.; et al. Restoration of vision after transplantation of photoreceptors. Nature 2012, 485, 99–103. [Google Scholar] [CrossRef] [PubMed]
- Reh, T.A. Photoreceptor Transplantation in Late Stage Retinal Degeneration. Investig. Ophthalmol. Vis. Sci. 2016, 57, ORSFg1–ORSFg7. [Google Scholar] [CrossRef] [PubMed]
- Hoon, M.; Okawa, H.; Della Santina, L.; Wong, R.O. Functional architecture of the retina: Development and disease. Prog. Retin. Eye Res. 2014, 42, 44–84. [Google Scholar] [CrossRef]
- Wallace, V.A. Concise review: Making a retina--from the building blocks to clinical applications. Stem Cells 2011, 29, 412–417. [Google Scholar] [CrossRef] [PubMed]
- Low, L.A.; Mummery, C.; Berridge, B.R.; Austin, C.P.; Tagle, D.A. Organs-on-chips: Into the next decade. Nat. Rev. Drug Discov. 2020. [Google Scholar] [CrossRef] [PubMed]
- Nakano, T.; Ando, S.; Takata, N.; Kawada, M.; Muguruma, K.; Sekiguchi, K.; Saito, K.; Yonemura, S.; Eiraku, M.; Sasai, Y. Self-formation of optic cups and storable stratified neural retina from human ESCs. Cell Stem Cell 2012, 10, 771–785. [Google Scholar] [CrossRef] [PubMed]
- Zhong, X.; Gutierrez, C.; Xue, T.; Hampton, C.; Vergara, M.N.; Cao, L.H.; Peters, A.; Park, T.S.; Zambidis, E.T.; Meyer, J.S.; et al. Generation of three-dimensional retinal tissue with functional photoreceptors from human iPSCs. Nat. Commun. 2014, 5, 4047. [Google Scholar] [CrossRef] [PubMed]
- Stenkamp, D.L. Development of the Vertebrate Eye and Retina. Prog. Mol. Biol. Transl. Sci. 2015, 134, 397–414. [Google Scholar] [CrossRef]
- Weiland, J.D.; Walston, S.T.; Humayun, M.S. Electrical Stimulation of the Retina to Produce Artificial Vision. Annu. Rev. Vis. Sci. 2016, 2, 273–294. [Google Scholar] [CrossRef]
- Zarbin, M.; Sugino, I.; Townes-Anderson, E. Concise Review: Update on Retinal Pigment Epithelium Transplantation for Age-Related Macular Degeneration. Stem Cells Transl. Med. 2019, 8, 466–477. [Google Scholar] [CrossRef]
- Yeste, J.; Garcia-Ramirez, M.; Illa, X.; Guimera, A.; Hernandez, C.; Simo, R.; Villa, R. A compartmentalized microfluidic chip with crisscross microgrooves and electrophysiological electrodes for modeling the blood-retinal barrier. Lab Chip 2017, 18, 95–105. [Google Scholar] [CrossRef]
- Chung, M.; Lee, S.; Lee, B.J.; Son, K.; Jeon, N.L.; Kim, J.H. Wet-AMD on a Chip: Modeling Outer Blood-Retinal Barrier In Vitro. Adv. Healthc. Mater. 2018, 7. [Google Scholar] [CrossRef]
- Pena, C.D.; Zhang, S.; Majeska, R.; Venkatesh, T.; Vazquez, M. Invertebrate Retinal Progenitors as Regenerative Models in a Microfluidic System. Cells 2019, 8, 1301. [Google Scholar] [CrossRef]
- Ail, D.; Perron, M. Retinal Degeneration and Regeneration-Lessons From Fishes and Amphibians. Curr. Pathobiol. Rep. 2017, 5, 67–78. [Google Scholar] [CrossRef] [PubMed]
- Fischer, A.J. Neural regeneration in the chick retina. Prog. Retin. Eye Res. 2005, 24, 161–182. [Google Scholar] [CrossRef] [PubMed]
- Prasad, S.; Galetta, S.L. Anatomy and physiology of the afferent visual system. Handb. Clin. Neurol. 2011, 102, 3–19. [Google Scholar] [CrossRef] [PubMed]
- Sabesan, R.; Schmidt, B.P.; Tuten, W.S.; Roorda, A. The elementary representation of spatial and color vision in the human retina. Sci. Adv. 2016, 2, e1600797. [Google Scholar] [CrossRef]
- Kaplan, H.J. Anatomy and function of the eye. Chem. Immunol. Allergy 2007, 92, 4–10. [Google Scholar] [CrossRef]
- Eghrari, A.O.; Riazuddin, S.A.; Gottsch, J.D. Overview of the Cornea: Structure, Function, and Development. Prog. Mol. Biol. Transl. Sci. 2015, 134, 7–23. [Google Scholar] [CrossRef]
- Quigley, H.A. The iris is a sponge: A cause of angle closure. Ophthalmology 2010, 117, 1–2. [Google Scholar] [CrossRef]
- Augusteyn, R.C. On the growth and internal structure of the human lens. Exp. Eye Res. 2010, 90, 643–654. [Google Scholar] [CrossRef]
- Uji, A.; Yoshimura, N. Microarchitecture of the Vitreous Body: A High-Resolution Optical Coherence Tomography Study. Am. J. Ophthalmol. 2016, 168, 24–30. [Google Scholar] [CrossRef]
- Lamb, T.D. Evolution of phototransduction, vertebrate photoreceptors and retina. Prog. Retin. Eye Res. 2013, 36, 52–119. [Google Scholar] [CrossRef]
- Nickla, D.L.; Wallman, J. The multifunctional choroid. Prog. Retin. Eye Res. 2010, 29, 144–168. [Google Scholar] [CrossRef] [PubMed]
- Watson, P.G.; Young, R.D. Scleral structure, organisation and disease. A review. Exp. Eye Res. 2004, 78, 609–623. [Google Scholar] [CrossRef]
- Vazquez, M. Electro-chemotactic stimuli for cell replacement therapy in neurosensory retina. Neural Regen. Res. 2020, 15, 450–452. [Google Scholar] [CrossRef] [PubMed]
- Ben M’Barek, K.; Habeler, W.; Monville, C. Stem Cell-Based RPE Therapy for Retinal Diseases: Engineering 3D Tissues Amenable for Regenerative Medicine. Adv. Exp. Med. Biol. 2018, 1074, 625–632. [Google Scholar] [CrossRef] [PubMed]
- Jha, B.S.; Bharti, K. Regenerating Retinal Pigment Epithelial Cells to Cure Blindness: A Road Towards Personalized Artificial Tissue. Curr. Stem Cell Rep. 2015, 1, 79–91. [Google Scholar] [CrossRef]
- Bringmann, A.; Pannicke, T.; Grosche, J.; Francke, M.; Wiedemann, P.; Skatchkov, S.N.; Osborne, N.N.; Reichenbach, A. Muller cells in the healthy and diseased retina. Prog. Retin. Eye Res. 2006, 25, 397–424. [Google Scholar] [CrossRef]
- Bhutto, I.; Lutty, G. Understanding age-related macular degeneration (AMD): Relationships between the photoreceptor/retinal pigment epithelium/Bruch’s membrane/choriocapillaris complex. Mol. Asp. Med. 2012, 33, 295–317. [Google Scholar] [CrossRef]
- De Jong, P. Elusive drusen and changing terminology of AMD. Eye 2018, 32, 904–914. [Google Scholar] [CrossRef]
- Shaw, P.X.; Stiles, T.; Douglas, C.; Ho, D.; Fan, W.; Du, H.; Xiao, X. Oxidative stress, innate immunity, and age-related macular degeneration. Aims Mol. Sci. 2016, 3, 196–221. [Google Scholar] [CrossRef]
- Lechner, J.; O’Leary, O.E.; Stitt, A.W. The pathology associated with diabetic retinopathy. Vis. Res. 2017, 139, 7–14. [Google Scholar] [CrossRef]
- Schnichels, S.; Paquet-Durand, F.; Loscher, M.; Tsai, T.; Hurst, J.; Joachim, S.C.; Klettner, A. Retina in a dish: Cell cultures, retinal explants and animal models for common diseases of the retina. Prog. Retin. Eye Res. 2020. [Google Scholar] [CrossRef] [PubMed]
- Weinreb, R.N.; Aung, T.; Medeiros, F.A. The pathophysiology and treatment of glaucoma: A review. JAMA 2014, 311, 1901–1911. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Choi, B. Nonlinear Relationship Between Blood Pressure and Glaucoma in US Adults. Am. J. Hypertens. 2019, 32, 308–316. [Google Scholar] [CrossRef] [PubMed]
- Paguirigan, A.L.; Beebe, D.J. Microfluidics meet cell biology: Bridging the gap by validation and application of microscale techniques for cell biological assays. Bioessays News Rev. Mol. Cell. Dev. Biol. 2008, 30, 811–821. [Google Scholar] [CrossRef] [PubMed]
- Seigel, G.M. The golden age of retinal cell culture. Mol. Vis. 1999, 5, 4. [Google Scholar]
- Seigel, G.M. Review: R28 retinal precursor cells: The first 20 years. Mol. Vis. 2014, 20, 301–306. [Google Scholar]
- Wheway, G.; Nazlamova, L.; Turner, D.; Cross, S. 661W Photoreceptor Cell Line as a Cell Model for Studying Retinal Ciliopathies. Front. Genet. 2019, 10, 308. [Google Scholar] [CrossRef]
- Tan, E.; Ding, X.Q.; Saadi, A.; Agarwal, N.; Naash, M.I.; Al-Ubaidi, M.R. Expression of cone-photoreceptor-specific antigens in a cell line derived from retinal tumors in transgenic mice. Investig. Ophthalmol. Vis. Sci. 2004, 45, 764–768. [Google Scholar] [CrossRef]
- Gomez-Vicente, V.; Flores, A.; Lax, P.; Murciano, C.; Yanez, A.; Gil, M.L.; Cuenca, N.; Gozalbo, D.; Maneu, V. Characterization of a new murine retinal cell line (MU-PH1) with glial, progenitor and photoreceptor characteristics. Exp. Eye Res. 2013, 110, 125–135. [Google Scholar] [CrossRef]
- Bejjani, A.; Choi, M.R.; Cassidy, L.; Collins, D.W.; O’Brien, J.M.; Murray, T.; Ksander, B.R.; Seigel, G.M. RB116: An RB1+ retinoblastoma cell line expressing primitive markers. Mol. Vis. 2012, 18, 2805–2813. [Google Scholar]
- Madreperla, S.A.; Bookstein, R.; Jones, O.W.; Lee, W.H. Retinoblastoma cell lines Y79, RB355 and WERI-Rb27 are genetically related. Ophthalmic Paediatr. Genet. 1991, 12, 49–56. [Google Scholar] [CrossRef]
- Trisler, D.; Rutin, J.; Pessac, B. Retinal engineering: Engrafted neural cell lines locate in appropriate layers. Proc. Natl. Acad. Sci. USA 1996, 93, 6269–6274. [Google Scholar] [CrossRef] [PubMed]
- Krishnamoorthy, R.R.; Clark, A.F.; Daudt, D.; Vishwanatha, J.K.; Yorio, T. A forensic path to RGC-5 cell line identification: Lessons learned. Investig. Ophthalmol. Vis. Sci. 2013, 54, 5712–5719. [Google Scholar] [CrossRef] [PubMed]
- Hammang, J.P.; Baetge, E.E.; Behringer, R.R.; Brinster, R.L.; Palmiter, R.D.; Messing, A. Immortalized retinal neurons derived from SV40 T-antigen-induced tumors in transgenic mice. Neuron 1990, 4, 775–782. [Google Scholar] [CrossRef]
- Phelan, K.; May, K.M. Basic Techniques in Mammalian Cell Tissue Culture. Curr. Protoc. Toxicol. 2016, 70, A 3B 1–A 3B 22. [Google Scholar] [CrossRef]
- Baust, J.M.; Buehring, G.C.; Campbell, L.; Elmore, E.; Harbell, J.W.; Nims, R.W.; Price, P.; Reid, Y.A.; Simione, F. Best practices in cell culture: An overview. In Vitro Cell. Dev. Biol. Anim. 2017, 53, 669–672. [Google Scholar] [CrossRef]
- Pfeffer, B.A.; Fliesler, S.J. Streamlined duplex live-dead microplate assay for cultured cells. Exp. Eye Res. 2017, 161, 17–29. [Google Scholar] [CrossRef]
- Chen, H.C. Boyden chamber assay. Methods Mol. Biol. 2005, 294, 15–22. [Google Scholar] [CrossRef]
- Chaubey, S.; Ridley, A.J.; Wells, C.M. Using the Dunn chemotaxis chamber to analyze primary cell migration in real time. Methods Mol. Biol. 2011, 769, 41–51. [Google Scholar] [CrossRef]
- Taylor, L.; Recio, C.; Greaves, D.R.; Iqbal, A.J. In Vitro Migration Assays. Methods Mol. Biol. 2018, 1784, 197–214. [Google Scholar] [CrossRef]
- Able, R.A., Jr.; Ngnabeuye, C.; Beck, C.; Holland, E.C.; Vazquez, M. Low Concentration Microenvironments Enhance the Migration of Neonatal Cells of Glial Lineage. Cell. Mol. Bioeng. 2012, 5. [Google Scholar] [CrossRef] [PubMed]
- Kong, Q.; Majeska, R.J.; Vazquez, M. Migration of connective tissue-derived cells is mediated by ultra-low concentration gradient fields of EGF. Exp. Cell Res. 2011, 317, 1491–1502. [Google Scholar] [CrossRef]
- Sun, X.; Liu, X.; Zhang, Y.; Kuang, X.; Lv, B.; Ge, J. A simple and effective pressure culture system modified from a transwell cell culture system. Biol. Res. 2013, 46, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Palama, I.E.; D′Amone, S.; Coluccia, A.M.; Biasiucci, M.; Gigli, G. Cell self-patterning on uniform PDMS-surfaces with controlled mechanical cues. Integr. Biol. Quant. Biosci. Nano Macro 2012, 4, 228–236. [Google Scholar] [CrossRef] [PubMed]
- Tay, C.Y.; Koh, C.G.; Tan, N.S.; Leong, D.T.; Tan, L.P. Mechanoregulation of stem cell fate via micro-/nano-scale manipulation for regenerative medicine. Nanomedicine 2013, 8, 623–638. [Google Scholar] [CrossRef]
- Falconnet, D.; Csucs, G.; Grandin, H.M.; Textor, M. Surface engineering approaches to micropattern surfaces for cell-based assays. Biomaterials 2006, 27, 3044–3063. [Google Scholar] [CrossRef]
- Li, N.; Tourovskaia, A.; Folch, A. Biology on a chip: Microfabrication for studying the behavior of cultured cells. Crit. Rev. Biomed. Eng. 2003, 31, 423–488. [Google Scholar] [CrossRef]
- Holmes, D.; Gawad, S. The application of microfluidics in biology. Methods Mol. Biol. 2010, 583, 55–80. [Google Scholar] [CrossRef]
- Tehranirokh, M.; Kouzani, A.Z.; Francis, P.S.; Kanwar, J.R. Microfluidic devices for cell cultivation and proliferation. Biomicrofluidics 2013, 7, 51502. [Google Scholar] [CrossRef]
- Warrick, J.W.; Murphy, W.L.; Beebe, D.J. Screening the cellular microenvironment: A role for microfluidics. IEEE Rev. Biomed. Eng. 2008, 1, 75–93. [Google Scholar] [CrossRef]
- Chung, B.G.; Manbachi, A.; Saadi, W.; Lin, F.; Jeon, N.L.; Khademhosseini, A. A gradient-generating microfluidic device for cell biology. J. Vis. Exp. 2007. [Google Scholar] [CrossRef] [PubMed]
- Mehling, M.; Tay, S. Microfluidic cell culture. Curr. Opin. Biotechnol. 2014, 25, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Hattori, K.; Sugiura, S.; Kanamori, T. Microfluidic perfusion culture. Methods Mol. Biol. 2014, 1104, 251–263. [Google Scholar] [CrossRef] [PubMed]
- Karimi, M.; Bahrami, S.; Mirshekari, H.; Basri, S.M.; Nik, A.B.; Aref, A.R.; Akbari, M.; Hamblin, M.R. Microfluidic systems for stem cell-based neural tissue engineering. Lab Chip 2016, 16, 2551–2571. [Google Scholar] [CrossRef]
- Bai, J.; Wang, C. Organoids and Microphysiological Systems: New Tools for Ophthalmic Drug Discovery. Front. Pharmacol. 2020, 11, 407. [Google Scholar] [CrossRef] [PubMed]
- Singh, T.; Vazquez, M. Time-Dependent Addition of Neuronal and Schwann Cells Increase Myotube Viability and Length in an In Vitro Tri-culture Model of the Neuromuscular Junction. Regen. Eng. Transl. Med. 2019, 5, 402–413. [Google Scholar] [CrossRef]
- Kong, Q.; Able, R.A., Jr.; Dudu, V.; Vazquez, M. A microfluidic device to establish concentration gradients using reagent density differences. J. Biomech. Eng. 2010, 132, 121012. [Google Scholar] [CrossRef]
- Karzbrun, E.; Kshirsagar, A.; Cohen, S.R.; Hanna, J.H.; Reiner, O. Human Brain Organoids on a Chip Reveal the Physics of Folding. Nat. Phys. 2018, 14, 515–522. [Google Scholar] [CrossRef]
- Pena, J.S.; Vazquez, M. VEGF Upregulates EGFR Expression to Stimulate Chemotactic Behaviors in the rMC-1 Model of Muller Glia. Brain Sci. 2020, 10, 330. [Google Scholar] [CrossRef]
- Pena, J.S.; Robles, D.; Zhang, S.; Vazquez, M. A Milled Microdevice to Advance Glia-Mediated Therapies in the Adult Nervous System. Micromachines 2019, 10, 513. [Google Scholar] [CrossRef]
- Lo Furno, D.; Mannino, G.; Pellitteri, R.; Zappala, A.; Parenti, R.; Gili, E.; Vancheri, C.; Giuffrida, R. Conditioned Media From Glial Cells Promote a Neural-Like Connexin Expression in Human Adipose-Derived Mesenchymal Stem Cells. Front. Physiol. 2018, 9, 1742. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.T.; Tsai, M.J.; Hsieh, N.; Lo, M.J.; Lee, M.J.; Cheng, H.; Huang, W.C. The superiority of conditioned medium derived from rapidly expanded mesenchymal stem cells for neural repair. Stem Cell Res. Ther. 2019, 10, 390. [Google Scholar] [CrossRef] [PubMed]
- Traverso, V.; Kinkl, N.; Grimm, L.; Sahel, J.; Hicks, D. Basic fibroblast and epidermal growth factors stimulate survival in adult porcine photoreceptor cell cultures. Investig. Ophthalmol. Vis. Sci. 2003, 44, 4550–4558. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Siffroi-Fernandez, S.; Felder-Schmittbuhl, M.P.; Khanna, H.; Swaroop, A.; Hicks, D. FGF19 exhibits neuroprotective effects on adult mammalian photoreceptors in vitro. Investig. Ophthalmol. Vis. Sci. 2008, 49, 1696–1704. [Google Scholar] [CrossRef]
- Hauck, S.M.; Gloeckner, C.J.; Harley, M.E.; Schoeffmann, S.; Boldt, K.; Ekstrom, P.A.; Ueffing, M. Identification of paracrine neuroprotective candidate proteins by a functional assay-driven proteomics approach. Mol. Cell. Proteom. MCP 2008, 7, 1349–1361. [Google Scholar] [CrossRef]
- Unterlauft, J.D.; Claudepierre, T.; Schmidt, M.; Muller, K.; Yafai, Y.; Wiedemann, P.; Reichenbach, A.; Eichler, W. Enhanced survival of retinal ganglion cells is mediated by Muller glial cell-derived PEDF. Exp. Eye Res. 2014, 127, 206–214. [Google Scholar] [CrossRef]
- Leung, D.W.; Lindlief, L.A.; Laabich, A.; Vissvesvaran, G.P.; Kamat, M.; Lieu, K.L.; Fawzi, A.; Kubota, R. Minocycline protects photoreceptors from light and oxidative stress in primary bovine retinal cell culture. Investig. Ophthalmol. Vis. Sci. 2007, 48, 412–421. [Google Scholar] [CrossRef]
- Luthra, S.; Narayanan, R.; Marques, L.E.; Chwa, M.; Kim, D.W.; Dong, J.; Seigel, G.M.; Neekhra, A.; Gramajo, A.L.; Brown, D.J.; et al. Evaluation of in vitro effects of bevacizumab (Avastin) on retinal pigment epithelial, neurosensory retinal, and microvascular endothelial cells. Retina 2006, 26, 512–518. [Google Scholar] [CrossRef]
- McUsic, A.C.; Lamba, D.A.; Reh, T.A. Guiding the morphogenesis of dissociated newborn mouse retinal cells and hES cell-derived retinal cells by soft lithography-patterned microchannel PLGA scaffolds. Biomaterials 2012, 33, 1396–1405. [Google Scholar] [CrossRef]
- Piccolino, M. Cajal and the retina: A 100-year retrospective. Trends Neurosci. 1988, 11, 521–525. [Google Scholar] [CrossRef]
- Kong, Q.; Vazquez, M. Flow-induced shear stresses increase the number of cell-cell contacts within extracellular matrix. J. Biomed. Mater. Res. Part A 2009, 89, 968–979. [Google Scholar] [CrossRef] [PubMed]
- Tiryaki, V.M.; Adia-Nimuwa, U.; Ayres, V.M.; Ahmed, I.; Shreiber, D.I. Texture-based segmentation and a new cell shape index for quantitative analysis of cell spreading in AFM images. Cytom. Part A J. Int. Soc. Anal. Cytol. 2015, 87, 1090–1100. [Google Scholar] [CrossRef] [PubMed]
- Kunzevitzky, N.J.; Willeford, K.T.; Feuer, W.J.; Almeida, M.V.; Goldberg, J.L. Amacrine cell subtypes differ in their intrinsic neurite growth capacity. Investig. Ophthalmol. Vis. Sci. 2013, 54, 7603–7613. [Google Scholar] [CrossRef]
- Tsukamoto, Y.; Omi, N. Classification of Mouse Retinal Bipolar Cells: Type-Specific Connectivity with Special Reference to Rod-Driven AII Amacrine Pathways. Front. Neuroanat. 2017, 11, 92. [Google Scholar] [CrossRef] [PubMed]
- Gogia, P.; Schneeweis, D.M. Neurite extension from rod bipolar cells in retinal cell and explant cultures. Conf. Proc. Annu. Int. Conf. IEEE Eng. Med. Biol. Soc. 2004, 2004, 4229–4232. [Google Scholar] [CrossRef]
- Mehenti, N.Z.; Tsien, G.S.; Leng, T.; Fishman, H.A.; Bent, S.F. A model retinal interface based on directed neuronal growth for single cell stimulation. Biomed. Microdevices 2006, 8, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Peterman, M.C.; Mehenti, N.Z.; Bilbao, K.V.; Lee, C.J.; Leng, T.; Noolandi, J.; Bent, S.F.; Blumenkranz, M.S.; Fishman, H.A. The Artificial Synapse Chip: A flexible retinal interface based on directed retinal cell growth and neurotransmitter stimulation. Artif. Organs 2003, 27, 975–985. [Google Scholar] [CrossRef] [PubMed]
- Bloch, E.; Luo, Y.; da Cruz, L. Advances in retinal prosthesis systems. Ther. Adv. Ophthalmol. 2019, 11, 2515841418817501. [Google Scholar] [CrossRef] [PubMed]
- Paknahad, J.; Loizos, K.; Humayun, M.; Lazzi, G. Targeted Stimulation of Retinal Ganglion Cells in Epiretinal Prostheses: A Multiscale Computational Study. IEEE Trans. Neural Syst. Rehabil. Eng. 2020. [Google Scholar] [CrossRef] [PubMed]
- Gomis, S.; Labib, M.; Coles, B.L.K.; van der Kooy, D.; Sargent, E.H.; Kelley, S.O. Single-Cell Tumbling Enables High-Resolution Size Profiling of Retinal Stem Cells. ACS Appl. Mater. Interfaces 2018, 10, 34811–34816. [Google Scholar] [CrossRef] [PubMed]
- Waldron, P.V.; Di Marco, F.; Kruczek, K.; Ribeiro, J.; Graca, A.B.; Hippert, C.; Aghaizu, N.D.; Kalargyrou, A.A.; Barber, A.C.; Grimaldi, G.; et al. Transplanted Donor- or Stem Cell-Derived Cone Photoreceptors Can Both Integrate and Undergo Material Transfer in an Environment-Dependent Manner. Stem Cell Rep. 2018, 10, 406–421. [Google Scholar] [CrossRef] [PubMed]
- Szabo, A.; Mayor, R. Mechanisms of Neural Crest Migration. Annu. Rev. Genet. 2018, 52, 43–63. [Google Scholar] [CrossRef] [PubMed]
- Jiang, C.; Klassen, H.; Zhang, X.; Young, M. Laser injury promotes migration and integration of retinal progenitor cells into host retina. Mol. Vis. 2010, 16, 983–990. [Google Scholar] [PubMed]
- Thakur, A.; Mishra, S.; Pena, J.; Zhou, J.; Redenti, S.; Majeska, R.; Vazquez, M. Collective adhesion and displacement of retinal progenitor cells upon extracellular matrix substrates of transplantable biomaterials. J. Tissue Eng. 2018, 9, 2041731417751286. [Google Scholar] [CrossRef]
- Abedin Zadeh, M.; Khoder, M.; Al-Kinani, A.A.; Younes, H.M.; Alany, R.G. Retinal cell regeneration using tissue engineered polymeric scaffolds. Drug Discov. Today 2019, 24, 1669–1678. [Google Scholar] [CrossRef] [PubMed]
- Kundu, J.; Michaelson, A.; Baranov, P.; Young, M.J.; Carrier, R.L. Approaches to cell delivery: Substrates and scaffolds for cell therapy. Dev. Ophthalmol. 2014, 53, 143–154. [Google Scholar] [CrossRef]
- Sip, C.G.; Bhattacharjee, N.; Folch, A. Microfluidic transwell inserts for generation of tissue culture-friendly gradients in well plates. Lab Chip 2014, 14, 302–314. [Google Scholar] [CrossRef]
- Unachukwu, U.J.; Sauane, M.; Vazquez, M.; Redenti, S. Microfluidic generated EGF-gradients induce chemokinesis of transplantable retinal progenitor cells via the JAK/STAT and PI3kinase signaling pathways. PLoS ONE 2013, 8, e83906. [Google Scholar] [CrossRef]
- Mishra, S.; Thakur, A.; Redenti, S.; Vazquez, M. A model microfluidics-based system for the human and mouse retina. Biomed. Microdevices 2015, 17, 107. [Google Scholar] [CrossRef]
- McCutcheon, S.; Unachukwu, U.; Thakur, A.; Majeska, R.; Redenti, S.; Vazquez, M. In vitro formation of neuroclusters in microfluidic devices and cell migration as a function of stromal-derived growth factor 1 gradients. Cell Adhes. Migr. 2017, 11, 1–12. [Google Scholar] [CrossRef] [PubMed]
- McCutcheon, S.; Majeska, R.; Schaffler, M.; Vazquez, M. A multiscale fluidic device for the study of dendrite-mediated cell to cell communication. Biomed. Microdevices 2017, 19, 71. [Google Scholar] [CrossRef]
- Mishra, S.; Pena, J.S.; Redenti, S.; Vazquez, M. A novel electro-chemotactic approach to impact the directional migration of transplantable retinal progenitor cells. Exp. Eye Res. 2019, 185, 107688. [Google Scholar] [CrossRef]
- Mishra, S.; Vazquez, M. A Gal-MmicroS Device to Evaluate Cell Migratory Response to Combined Galvano-Chemotactic Fields. Biosensors 2017, 7, 54. [Google Scholar] [CrossRef]
- Vazquez, M.; Pena, J.S.; Mut, S. An Ex Vivo Eye Facsimile System (EVES) to evaluate transplantation strategies for cell replacement therapy. Investig. Ophthalmol. Vis. Sci. 2020, 61, 795. [Google Scholar]
- Romo, P.; Madigan, M.C.; Provis, J.M.; Cullen, K.M. Differential effects of TGF-beta and FGF-2 on in vitro proliferation and migration of primate retinal endothelial and Muller cells. Acta Ophthalmol. 2011, 89, e263–e268. [Google Scholar] [CrossRef] [PubMed]
- Willbold, E.; Layer, P.G. Muller glia cells and their possible roles during retina differentiation in vivo and in vitro. Histol. Histopathol. 1998, 13, 531–552. [Google Scholar] [CrossRef]
- Pena, J.; Dulger, N.; Singh, T.; Zhou, J.; Majeska, R.; Redenti, S.; Vazquez, M. Controlled microenvironments to evaluate chemotactic properties of cultured Muller glia. Exp. Eye Res. 2018, 173, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Remé, C.E.; Grimm, C.; Hafezi, F.; Wenzel, A.; Williams, T.P. Apoptosis in the Retina: The Silent Death of Vision. News Physiol. Sci. 2000, 15. [Google Scholar] [CrossRef] [PubMed]
- Clark, A.J.; Yang, P.; Khaderi, K.R.; Moshfeghi, A.A. Ocular Tolerance of Contemporary Electronic Display Devices. Ophthalmic Surg. Lasers Imaging Retin. 2018, 49, 346–354. [Google Scholar] [CrossRef]
- Du, Y.; Hirooka, K.; Miyamoto, O.; Itano, T.; Tokuda, M.; Shiraga, F. Both amacrine and bipolar cells release glutamate in the rat retina after ischemia/reperfusion insult in vitro. Curr. Eye Res. 2008, 33, 782–788. [Google Scholar] [CrossRef]
- Kuse, Y.; Ogawa, K.; Tsuruma, K.; Shimazawa, M.; Hara, H. Damage of photoreceptor-derived cells in culture induced by light emitting diode-derived blue light. Sci. Rep. 2014, 4, 5223. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Cho, H.; Hartsock, M.J.; Mitchell, K.L.; Gong, J.; Wu, L.; Wei, Y.; Wang, S.; Thimmulappa, R.K.; Sporn, M.B.; et al. Neuroprotective role of Nrf2 for retinal ganglion cells in ischemia-reperfusion. J. Neurochem. 2015, 133, 233–241. [Google Scholar] [CrossRef] [PubMed]
- Nunez-Alvarez, C.; Osborne, N.N. Blue light exacerbates and red light counteracts negative insults to retinal ganglion cells in situ and R28 cells in vitro. Neurochem. Int. 2019, 125, 187–196. [Google Scholar] [CrossRef]
- Du, Z.; Wu, X.; Song, M.; Li, P.; Wang, L. Oxidative damage induces MCP-1 secretion and macrophage aggregation in age-related macular degeneration (AMD). Graefe’s Arch. Clin. Exp. Ophthalmol. 2016, 254, 2469–2476. [Google Scholar] [CrossRef]
- Mencl, S.; Trifunovic, D.; Zrenner, E.; Paquet-Durand, F. PKG-Dependent Cell Death in 661W Cone Photoreceptor-like Cell Cultures (Experimental Study). Adv. Exp. Med. Biol. 2018, 1074, 511–517. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Wang, W.; Li, X.; Huang, C.; Zhang, Z.; Yuan, M.; Li, X. High hydrostatic pressure induces apoptosis of retinal ganglion cells via regulation of the NGF signalling pathway. Mol. Med. Rep. 2019, 19, 5321–5334. [Google Scholar] [CrossRef]
- Liu, B.; Ma, X.; Guo, D.; Guo, Y.; Chen, N.; Bi, H. Neuroprotective effect of alpha-lipoic acid on hydrostatic pressure-induced damage of retinal ganglion cells in vitro. Neurosci. Lett. 2012, 526, 24–28. [Google Scholar] [CrossRef]
- Cui, Y.; Xu, N.; Xu, W.; Xu, G. Mesenchymal stem cells attenuate hydrogen peroxide-induced oxidative stress and enhance neuroprotective effects in retinal ganglion cells. Vitr. Cell. Dev. Biol. Anim. 2017, 53, 328–335. [Google Scholar] [CrossRef]
- Kung, F.; Wang, J.; Perez-Castillejos, R.; Townes-Anderson, E. Position along the nasal/temporal plane affects synaptic development by adult photoreceptors, revealed by micropatterning. Integr. Biol. Quant. Biosci. Nano Macro 2015, 7, 313–323. [Google Scholar] [CrossRef]
- Curcio, C.A.; Allen, K.A.; Sloan, K.R.; Lerea, C.L.; Hurley, J.B.; Klock, I.B.; Milam, A.H. Distribution and morphology of human cone photoreceptors stained with anti-blue opsin. J. Comp. Neurol. 1991, 312, 610–624. [Google Scholar] [CrossRef]
- Curcio, C.A.; Sloan, K.R.; Kalina, R.E.; Hendrickson, A.E. Human photoreceptor topography. J. Comp. Neurol. 1990, 292, 497–523. [Google Scholar] [CrossRef]
- Chan, T.L.; Martin, P.R.; Clunas, N.; Grunert, U. Bipolar cell diversity in the primate retina: Morphologic and immunocytochemical analysis of a new world monkey, the marmoset Callithrix jacchus. J. Comp. Neurol. 2001, 437, 219–239. [Google Scholar] [CrossRef]
- Schulte, D.; Peters, M.A.; Sen, J.; Cepko, C.L. The rod photoreceptor pattern is set at the optic vesicle stage and requires spatially restricted cVax expression. J. Neurosci. Off. J. Soc. Neurosci. 2005, 25, 2823–2831. [Google Scholar] [CrossRef]
- Ma, Y.; Han, X.; de Castro, R.B.; Zhang, P.; Zhang, K.; Hu, Z.; Qin, L. Analysis of the bystander effect in cone photoreceptors via a guided neural network platform. Sci. Adv. 2018, 4, eaas9274. [Google Scholar] [CrossRef]
- Rozental, R.; Giaume, C.; Spray, D.C. Gap junctions in the nervous system. Brain Res. Rev. 2000, 32, 11–15. [Google Scholar] [CrossRef]
- Cusato, K.; Bosco, A.; Rozental, R.; Guimaraes, C.A.; Reese, B.E.; Linden, R.; Spray, D.C. Gap junctions mediate bystander cell death in developing retina. J. Neurosci. Off. J. Soc. Neurosci. 2003, 23, 6413–6422. [Google Scholar] [CrossRef]
- Su, P.J.; Liu, Z.; Zhang, K.; Han, X.; Saito, Y.; Xia, X.; Yokoi, K.; Shen, H.; Qin, L. Retinal synaptic regeneration via microfluidic guiding channels. Sci. Rep. 2015, 5, 13591. [Google Scholar] [CrossRef]
- Rountree, C.M.; Raghunathan, A.; Troy, J.B.; Saggere, L. Prototype chemical synapse chip for spatially patterned neurotransmitter stimulation of the retina ex vivo. Microsyst. Nanoeng. 2017, 3, 17052. [Google Scholar] [CrossRef] [PubMed]
- Peterman, M.C.; Noolandi, J.; Blumenkranz, M.S.; Fishman, H.A. Localized chemical release from an artificial synapse chip. Proc. Natl. Acad. Sci. USA 2004, 101, 9951–9954. [Google Scholar] [CrossRef]
- Vohra, R.; Aldana, B.I.; Bulli, G.; Skytt, D.M.; Waagepetersen, H.; Bergersen, L.H.; Kolko, M. Lactate-Mediated Protection of Retinal Ganglion Cells. J. Mol. Biol. 2019, 431, 1878–1888. [Google Scholar] [CrossRef] [PubMed]
- Skytt, D.M.; Toft-Kehler, A.K.; Braendstrup, C.T.; Cejvanovic, S.; Gurubaran, I.S.; Bergersen, L.H.; Kolko, M. Glia-Neuron Interactions in the Retina Can Be Studied in Cocultures of Muller Cells and Retinal Ganglion Cells. Biomed Res. Int. 2016, 2016, 1087647. [Google Scholar] [CrossRef] [PubMed]
- Ruzafa, N.; Vecino, E. Effect of Muller cells on the survival and neuritogenesis in retinal ganglion cells. Arch. De La Soc. Esp. De Oftalmol. 2015, 90, 522–526. [Google Scholar] [CrossRef]
- Abrahan, C.E.; Insua, M.F.; Politi, L.E.; German, O.L.; Rotstein, N.P. Oxidative stress promotes proliferation and dedifferentiation of retina glial cells in vitro. J. Neurosci. Res. 2009, 87, 964–977. [Google Scholar] [CrossRef]
- Navneet, S.; Zhao, J.; Wang, J.; Mysona, B.; Barwick, S.; Ammal Kaidery, N.; Saul, A.; Kaddour-Djebbar, I.; Bollag, W.B.; Thomas, B.; et al. Hyperhomocysteinemia-induced death of retinal ganglion cells: The role of Muller glial cells and NRF2. Redox Biol. 2019, 24, 101199. [Google Scholar] [CrossRef] [PubMed]
- Goldman, D. Muller glial cell reprogramming and retina regeneration. Nat. Rev. Neurosci. 2014, 15, 431–442. [Google Scholar] [CrossRef] [PubMed]
- Fischer, A.J.; Bongini, R. Turning Muller glia into neural progenitors in the retina. Mol. Neurobiol. 2010, 42, 199–209. [Google Scholar] [CrossRef] [PubMed]
- Lahne, M.; Nagashima, M.; Hyde, D.R.; Hitchcock, P.F. Reprogramming Muller Glia to Regenerate Retinal Neurons. Annu. Rev. Vis. Sci. 2020, 6, 171–193. [Google Scholar] [CrossRef] [PubMed]
- Pereiro, X.; Ruzafa, N.; Acera, A.; Fonollosa, A.; Rodriguez, F.D.; Vecino, E. Dexamethasone protects retinal ganglion cells but not Muller glia against hyperglycemia in vitro. PLoS ONE 2018, 13, e0207913. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.Y.; Song, J.Y.; Fan, B.; Wang, Y.; Pan, Y.R.; Che, L.; Sun, Y.J.; Li, G.Y. Resveratrol protects photoreceptors by blocking caspase- and PARP-dependent cell death pathways. Free Radic. Biol. Med. 2018, 129, 569–581. [Google Scholar] [CrossRef]
- Taki, K.; Horie, T.; Kida, T.; Mimura, M.; Ikeda, T.; Oku, H. Impairment of Autophagy Causes Superoxide Formation and Caspase Activation in 661 W Cells, a Cell Line for Cone Photoreceptors, under Hyperglycemic Conditions. Int. J. Mol. Sci. 2020, 21, 4240. [Google Scholar] [CrossRef]
- Bai, J.; Khajavi, M.; Sui, L.; Fu, H.; Tarakkad Krishnaji, S.; Birsner, A.E.; Bazinet, L.; Kamm, R.D.; D′Amato, R.J. Angiogenic responses in a 3D micro-engineered environment of primary endothelial cells and pericytes. Angiogenesis 2020. [Google Scholar] [CrossRef]
- Li, Y.; Lu, Y.; Chen, Q.; Kang, Y.; Yu, L. Probing of peripheral blood mononuclear cells anchoring on TNF-alpha challenged-vascular endothelia in an in vitro model of the retinal microvascular. Biomed. Microdevices 2017, 19, 54. [Google Scholar] [CrossRef]
- Jo, D.H.; Kim, J.H.; Kim, J.H. A platform of integrative studies from in vitro to in vivo experiments: Towards drug development for ischemic retinopathy. Biomed. Pharmacother. 2015, 69, 367–373. [Google Scholar] [CrossRef] [PubMed]
- Dodson, K.H.; Echevarria, F.D.; Li, D.; Sappington, R.M.; Edd, J.F. Retina-on-a-chip: A microfluidic platform for point access signaling studies. Biomed. Microdevices 2015, 17, 114. [Google Scholar] [CrossRef] [PubMed]
- Djojosubroto, M.W.; Arsenijevic, Y. Retinal stem cells: Promising candidates for retina transplantation. Cell Tissue Res. 2008, 331, 347–357. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.K.; Nasonkin, I.O. Limitations and Promise of Retinal Tissue From Human Pluripotent Stem Cells for Developing Therapies of Blindness. Front. Cell. Neurosci. 2020, 14, 179. [Google Scholar] [CrossRef]
- Lang, S.; von Philipsborn, A.C.; Bernard, A.; Bonhoeffer, F.; Bastmeyer, M. Growth cone response to ephrin gradients produced by microfluidic networks. Anal. Bioanal. Chem. 2008, 390, 809–816. [Google Scholar] [CrossRef][Green Version]
- Li, Y.; Tao, W.; Luo, L.; Huang, D.; Kauper, K.; Stabila, P.; Lavail, M.M.; Laties, A.M.; Wen, R. CNTF induces regeneration of cone outer segments in a rat model of retinal degeneration. PLoS ONE 2010, 5, e9495. [Google Scholar] [CrossRef]
- Fan, B.; Sun, Y.J.; Liu, S.Y.; Che, L.; Li, G.Y. Neuroprotective Strategy in Retinal Degeneration: Suppressing ER Stress-Induced Cell Death via Inhibition of the mTOR Signal. Int. J. Mol. Sci. 2017, 18, 201. [Google Scholar] [CrossRef]
- Teotia, P.; Van Hook, M.J.; Fischer, D.; Ahmad, I. Human retinal ganglion cell axon regeneration by recapitulating developmental mechanisms: Effects of recruitment of the mTOR pathway. Development 2019, 146. [Google Scholar] [CrossRef]
- Blagovic, K.; Kim, L.Y.; Voldman, J. Microfluidic perfusion for regulating diffusible signaling in stem cells. PLoS ONE 2011, 6, e22892. [Google Scholar] [CrossRef] [PubMed]
- Guild, J.; Haque, A.; Gheibi, P.; Gao, Y.; Son, K.J.; Foster, E.; Dumont, S.; Revzin, A. Embryonic Stem Cells Cultured in Microfluidic Chambers Take Control of Their Fate by Producing Endogenous Signals Including LIF. Stem Cells 2016, 34, 1501–1512. [Google Scholar] [CrossRef] [PubMed]
- Abdolvand, N.; Tostoes, R.; Raimes, W.; Kumar, V.; Szita, N.; Veraitch, F. Long-Term Retinal Differentiation of Human Induced Pluripotent Stem Cells in a Continuously Perfused Microfluidic Culture Device. Biotechnol. J. 2019, 14, e1800323. [Google Scholar] [CrossRef] [PubMed]
- Steedman, M.R.; Tao, S.L.; Klassen, H.; Desai, T.A. Enhanced differentiation of retinal progenitor cells using microfabricated topographical cues. Biomed. Microdevices 2010, 12, 363–369. [Google Scholar] [CrossRef] [PubMed]
- Neeley, W.L.; Redenti, S.; Klassen, H.; Tao, S.; Desai, T.; Young, M.J.; Langer, R. A microfabricated scaffold for retinal progenitor cell grafting. Biomaterials 2008, 29, 418–426. [Google Scholar] [CrossRef] [PubMed]
- Redenti, S.; Tao, S.; Yang, J.; Gu, P.; Klassen, H.; Saigal, S.; Desai, T.; Young, M.J. Retinal tissue engineering using mouse retinal progenitor cells and a novel biodegradable, thin-film poly(e-caprolactone) nanowire scaffold. J. Ocul. Biol. Dis. Inform. 2008, 1, 19–29. [Google Scholar] [CrossRef]
- Yao, J.; Ko, C.W.; Baranov, P.Y.; Regatieri, C.V.; Redenti, S.; Tucker, B.A.; Mighty, J.; Tao, S.L.; Young, M.J. Enhanced differentiation and delivery of mouse retinal progenitor cells using a micropatterned biodegradable thin-film polycaprolactone scaffold. Tissue Eng. Part A 2015, 21, 1247–1260. [Google Scholar] [CrossRef]
- Li, K.; Zhong, X.; Yang, S.; Luo, Z.; Li, K.; Liu, Y.; Cai, S.; Gu, H.; Lu, S.; Zhang, H.; et al. HiPSC-derived retinal ganglion cells grow dendritic arbors and functional axons on a tissue-engineered scaffold. Acta Biomater. 2017, 54, 117–127. [Google Scholar] [CrossRef]
- Blong, C.C.; Jeon, C.J.; Yeo, J.Y.; Ye, E.A.; Oh, J.; Callahan, J.M.; Law, W.D.; Mallapragada, S.K.; Sakaguchi, D.S. Differentiation and behavior of human neural progenitors on micropatterned substrates and in the developing retina. J. Neurosci. Res. 2010, 88, 1445–1456. [Google Scholar] [CrossRef]
- Rieke, M.; Gottwald, E.; Weibezahn, K.F.; Layer, P.G. Tissue reconstruction in 3D-spheroids from rodent retina in a motion-free, bioreactor-based microstructure. Lab Chip 2008, 8, 2206–2213. [Google Scholar] [CrossRef]
- Wang, Y.; Pan, J.; Wang, D.; Liu, J. The Use of Stem Cells in Neural Regeneration: A Review of Current Opinion. Curr. Stem Cell Res. Ther. 2018, 13, 608–617. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Prinsen, P.; Wang, H.; Bai, Z.; Wang, H.; Luque, R.; Xuan, J. Macroporous materials: Microfluidic fabrication, functionalization and applications. Chem. Soc. Rev. 2017, 46, 855–914. [Google Scholar] [CrossRef] [PubMed]
- Chung, B.G.; Lee, K.H.; Khademhosseini, A.; Lee, S.H. Microfluidic fabrication of microengineered hydrogels and their application in tissue engineering. Lab Chip 2012, 12, 45–59. [Google Scholar] [CrossRef] [PubMed]
- Tezcaner, A.; Hicks, D. In vitro characterization of micropatterned PLGA-PHBV8 blend films as temporary scaffolds for photoreceptor cells. J. Biomed. Mater. Res. Part A 2008, 86, 170–181. [Google Scholar] [CrossRef]
- Jung, Y.H.; Phillips, M.J.; Lee, J.; Xie, R.; Ludwig, A.L.; Chen, G.; Zheng, Q.; Kim, T.J.; Zhang, H.; Barney, P.; et al. 3D Microstructured Scaffolds to Support Photoreceptor Polarization and Maturation. Adv. Mater. 2018, 30, e1803550. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vazquez, M. Microfluidic and Microscale Assays to Examine Regenerative Strategies in the Neuro Retina. Micromachines 2020, 11, 1089. https://doi.org/10.3390/mi11121089
Vazquez M. Microfluidic and Microscale Assays to Examine Regenerative Strategies in the Neuro Retina. Micromachines. 2020; 11(12):1089. https://doi.org/10.3390/mi11121089
Chicago/Turabian StyleVazquez, Maribel. 2020. "Microfluidic and Microscale Assays to Examine Regenerative Strategies in the Neuro Retina" Micromachines 11, no. 12: 1089. https://doi.org/10.3390/mi11121089
APA StyleVazquez, M. (2020). Microfluidic and Microscale Assays to Examine Regenerative Strategies in the Neuro Retina. Micromachines, 11(12), 1089. https://doi.org/10.3390/mi11121089