
Vaccines against Botulism


Abstract
:1. Nucleic Acid-Based Vaccines against Botulism
Plasmid-Based Vaccines against Botulism
2. Viral Vector Vaccines
2.1. Adenovirus-Based Vectors
2.2. Influenza Virus-Based Vectors
2.3. Rabies Virus-Based Vectors
2.4. Alphaviruses
2.4.1. Semliki Forest Virus (SFV)-Based Plasmid DNA Replicon Vectors
2.4.2. Semliki Forest Virus-Based Viral Vectors
2.4.3. Venezuelan Equine Encephalitis Virus-Based Vectors
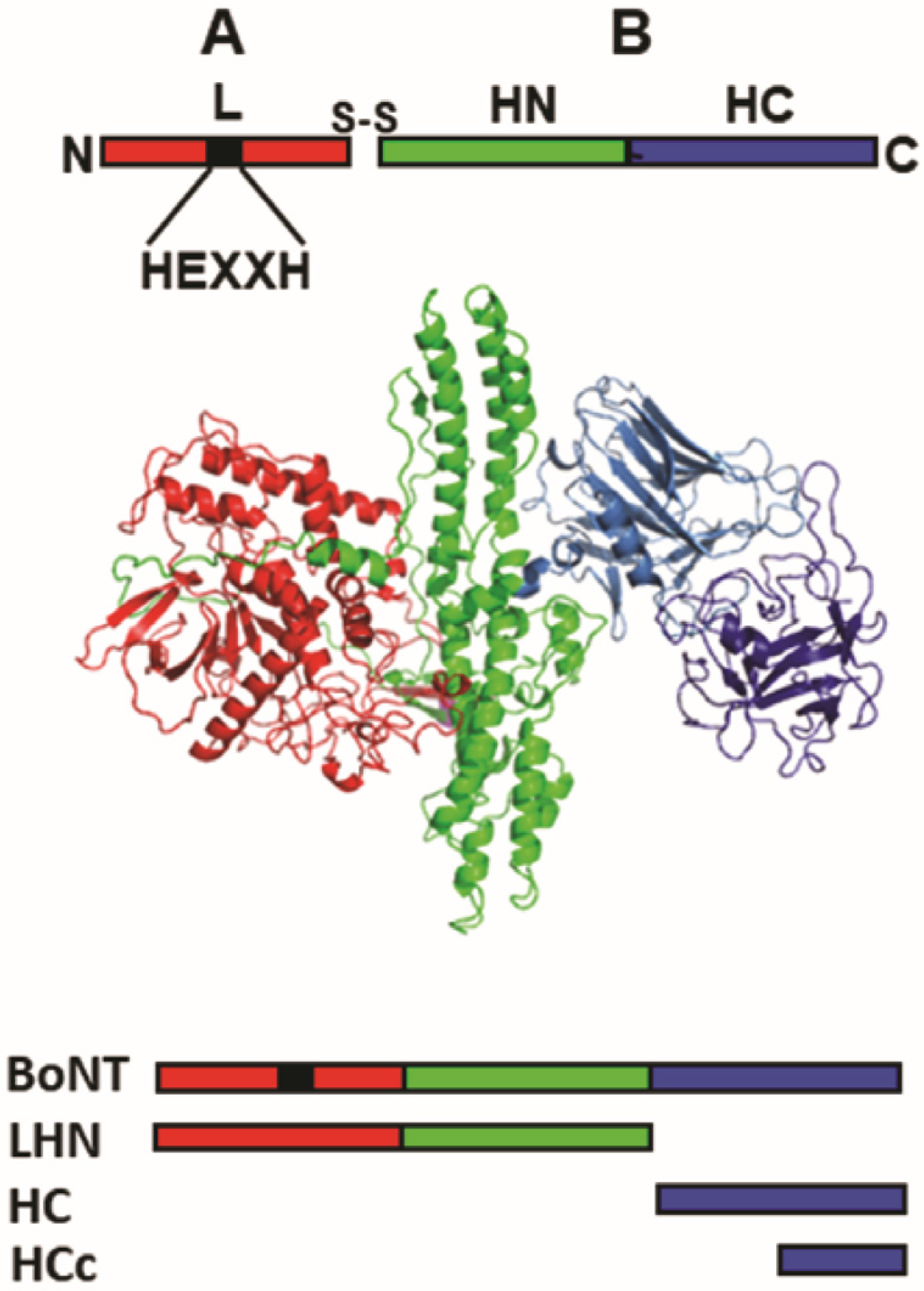
3. Protein-Based Vaccines against Botulism
3.1. Chemically Detoxified BoNT Vaccine
3.2. Recombinant BoNT Vaccines
3.2.1. Recombinant, HC-Derived Vaccines
3.2.2. Recombinant, LHN-Derived Vaccine
3.2.3. Engineered, Full-Length BoNT Vaccines
4. Veterinary BoNT Vaccines
5. Summary
Acknowledgments
Conflicts of Interest
References
- CDC, Botulism. Centers of Disease Control and Prevention 2017, 1600 Clifton Road Atlanta, GA 30329–4027 USA; CDC, Botulism: Atlanta, GA, USA, 2017.
- Gill, D.M. Bacterial toxins: A table of lethal amounts. Microbiol. Rev. 1982, 46, 86–94. [Google Scholar] [PubMed]
- Hill, K.K.; Smith, T.J.; Helma, C.H.; Ticknor, L.O.; Foley, B.T.; Svensson, R.T.; Brown, J.L.; Johnson, E.A.; Smith, L.A.; Okinaka, R.T.; et al. Genetic diversity among Botulinum Neurotoxin-producing clostridial strains. J. Bacteriol. 2007, 189, 818–832. [Google Scholar] [CrossRef] [PubMed]
- Schiavo, G.; Matteoli, M.; Montecucco, C. Neurotoxins affecting neuroexocytosis. Physiol. Rev. 2000, 80, 717–766. [Google Scholar] [PubMed]
- Montecucco, C.; Schiavo, G. Tetanus and botulism neurotoxins: A new group of zinc proteases. Trends Biochem. Sci. 1993, 18, 324–327. [Google Scholar] [CrossRef]
- Montecucco, C.; Rossetto, O.; Schiavo, G. Presynaptic receptor arrays for clostridial neurotoxins. Trends Microbiol. 2004, 12, 442–446. [Google Scholar] [CrossRef] [PubMed]
- Lam, K.H.; Yao, G.; Jin, R. Diverse binding modes, same goal: The receptor recognition mechanism of botulinum neurotoxin. Prog. Biophys. Mol. Biol. 2015, 117, 225–231. [Google Scholar] [CrossRef] [PubMed]
- Whitemarsh, R.C.; Tepp, W.H.; Johnson, E.A.; Pellett, S. Persistence of botulinum neurotoxin a subtypes 1–5 in primary rat spinal cord cells. PLoS ONE 2014, 9, e90252. [Google Scholar] [CrossRef] [PubMed]
- Hill, K.K.; Smith, T.J. Genetic diversity within Clostridium botulinum serotypes, botulinum neurotoxin gene clusters and toxin subtypes. Curr. Top. Microbiol. Immunol. 2013, 364, 1–20. [Google Scholar] [PubMed]
- Pellett, S.; Tepp, W.H.; Bradshaw, M.; Kalb, S.R.; Dykes, J.K.; Lin, G.; Nawrocki, E.M.; Pier, C.L.; Barr, J.R.; Maslanka, S.E.; et al. Purification and Characterization of Botulinum Neurotoxin FA from a Genetically Modified Clostridium botulinum Strain. mSphere 2016, 1, e00100-15. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Masuyer, G.; Zhang, J.; Shen, Y.; Lundin, D.; Henriksson, L.; Miyashita, S.I.; Martinez-Carranza, M.; Dong, M.; Stenmark, P. Identification and characterization of a novel botulinum neurotoxin. Nat. Commun. 2017, 8, 14130. [Google Scholar] [CrossRef] [PubMed]
- Hill, K.K.; Xie, G.; Foley, B.T.; Smith, T.J. Genetic diversity within the botulinum neurotoxin-producing bacteria and their neurotoxins. Toxicon 2015, 107, 2–8. [Google Scholar] [CrossRef] [PubMed]
- Graham, R.; Thorp, F. The Effect of Formalin on Botulinum Toxins A, B and C. J. Immunol. 1929, 16, 391–401. [Google Scholar]
- CDC. Notice of CDC's discontinuation of investigational pentavalent (ABCDE) botulinum toxoid vaccine for workers at risk for occupational exposure to botulinum toxins. Morb. Mortal. Wkly. Rep. 2011, 60, 1454–1455. [Google Scholar]
- Rusnak, J.M.; Smith, L.A. Botulinum neurotoxin vaccines: Past history and recent developments. Hum. Vaccines 2009, 5, 794–805. [Google Scholar] [CrossRef]
- Scott, V.L.; Villarreal, D.O.; Hutnick, N.A.; Walters, J.N.; Ragwan, E.; Bdeir, K.; Yan, J.; Sardesai, N.Y.; Finnefrock, A.C.; Casimiro, D.R.; et al. DNA vaccines targeting heavy chain C-terminal fragments of Clostridium botulinum neurotoxin serotypes A, B, and E induce potent humoral and cellular immunity and provide protection from lethal toxin challenge. Hum. Vaccines Immunother. 2015, 11, 1961–1971. [Google Scholar] [CrossRef] [PubMed]
- Jathoul, A.P.; Holley, J.L.; Garmory, H.S. Efficacy of DNA vaccines expressing the type F botulinum toxin Hc fragment using different promoters. Vaccine 2004, 22, 3942–3946. [Google Scholar] [CrossRef] [PubMed]
- Bennett, A.M.; Perkins, S.D.; Holley, J.L. DNA vaccination protects against botulinum neurotoxin type F. Vaccine 2003, 21, 3110–3117. [Google Scholar] [CrossRef]
- Trollet, C.; Pereira, Y.; Burgain, A.; Litzler, E.; Mezrahi, M.; Seguin, J.; Manich, M.; Popoff, M.R.; Scherman, D.; Bigey, P. Generation of High-Titer Neutralizing Antibodies against Botulinum Toxins A, B, and E by DNA Electrotransfer. Infect. Immun. 2009, 77, 2221–2229. [Google Scholar] [CrossRef] [PubMed]
- Shyu, R.-H.; Shaio, M.-F.; Tang, S.-S.; Shyu, H.-F.; Lee, C.-F.; Tsai, M.-H.; Smith, J.E.; Huang, H.-H.; Wey, J.-J.; Huang, J.-L.; et al. DNA Vaccination Using the Fragment C of Botulinum Neurotoxin Type A Provided Protective Immunity in Mice. J. Biomed. Sci. 2000, 7, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Clayton, J.; Middlebrook, J.L. Vaccination of mice with DNA encoding a large fragment of botulinum neurotoxin serotype A. Vaccine 2000, 18, 1855–1862. [Google Scholar] [CrossRef]
- Lasaro, M.O.; Ertl, H.C.J. New Insights on Adenovirus as Vaccine Vectors. Mol. Ther. 2009, 17, 1333–1339. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Pichichero, M.E.; Simpson, L.L.; Elias, M.; Smith, L.A.; Zeng, M. An adenoviral vector-based mucosal vaccine is effective in protection against botulism. Gene Ther. 2009, 16, 367–375. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Xu, Q.; Zeng, M. Oral vaccination with an adenovirus-vectored vaccine protects against botulism. Vaccine 2013, 31, 1009–1011. [Google Scholar] [CrossRef] [PubMed]
- Zeng, M.; Xu, Q.; Elias, M.; Pichichero, M.E.; Simpson, L.L.; Smith, L.A. Protective immunity against botulism provided by a single dose vaccination with an adenovirus-vectored vaccine. Vaccine 2007, 25, 7540–7548. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Diaz-Arévalo, D.; Chen, Y.; Zeng, M. Intranasal Vaccination with an Engineered Influenza Virus Expressing the Receptor Binding Subdomain of Botulinum Neurotoxin Provides Protective Immunity Against Botulism and Influenza. Front. Immunol. 2015, 6, 170. [Google Scholar] [CrossRef] [PubMed]
- Mustafa, W.; Al-Saleem, F.H.; Nasser, Z.; Olson, R.M.; Mattis, J.A.; Simpson, L.L.; Schnell, M.J. Immunization of mice with the non-toxic HC50 domain of botulinum neurotoxin presented by rabies virus particles induces a strong immune response affording protection against high-dose botulinum neurotoxin challenge. Vaccine 2011, 29, 4638–4645. [Google Scholar] [CrossRef] [PubMed]
- Hudacek, A.W.; Al-Saleem, F.H.; Willet, M.; Eisemann, T.; Mattis, J.A.; Simpson, L.L.; Schnell, M.J. Recombinant rabies virus particles presenting botulinum neurotoxin antigens elicit a protective humoral response in vivo. Mol. Ther. Methods Clin. Dev. 2014, 1, 14046. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Liu, S.; Ma, Y.; Gong, Z.-W.; Wang, S.; Sun, Z.-W. Pentavalent replicon vaccines against botulinum neurotoxins and tetanus toxin using DNA-based Semliki Forest virus replicon vectors. Hum. Vaccines Immunother. 2014, 10, 1874–1879. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.-Z.; Guo, J.-P.; An, H.-J.; Zhang, S.-M.; Wang, S.; Yu, W.-Y.; Sun, Z.-W. Potent tetravalent replicon vaccines against botulinum neurotoxins using DNA-based Semliki Forest virus replicon vectors. Vaccine 2013, 31, 2427–2432. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.-Z.; Zhang, S.-M.; Sun, Z.-W.; Wang, S.; Yu, W.-Y. Enhanced immune responses using plasmid DNA replicon vaccine encoding the Hc domain of Clostridium botulinum neurotoxin serotype A. Vaccine 2007, 25, 8843–8850. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Yu, J.; Li, N.; Wang, S.; Yu, W.; Sun, Z. Individual and bivalent vaccines against botulinum neurotoxin serotypes A and B using DNA-based Semliki Forest virus vectors. Vaccine 2009, 27, 6148–6153. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.S.; Pushko, P.; Parker, M.D.; Dertzbaugh, M.T.; Smith, L.A.; Smith, J.F. Candidate vaccine against botulinum neurotoxin serotype A derived from a Venezuelan equine encephalitis virus vector system. Infect. Immun. 2001, 69, 5709–5715. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.S.; Groebner, J.L.; Hadjipanayis, A.G.; Negley, D.L.; Schmaljohn, A.L.; Welkos, S.L.; Smith, L.A.; Smith, J.F. Multiagent vaccines vectored by Venezuelan equine encephalitis virus replicon elicits immune responses to Marburg virus and protection against anthrax and botulinum neurotoxin in mice. Vaccine 2006, 24, 6886–6892. [Google Scholar] [CrossRef] [PubMed]
- Appaiahgari, M.; Pandey, R.; Vrati, S. Seroprevalence of Neutralizing Antibodies to Adenovirus Type 5 among Children in India: Implications for Recombinant Adenovirus-Based Vaccines. Clin. Vaccine Immunol. 2007, 14, 1053–1055. [Google Scholar] [CrossRef] [PubMed]
- Chirmule, N.; Propert, K.J.; Magosin, S.A.; Qian, Y.; Qian, R.; Wilson, J.M. Immune responses to adenovirus and adeno-associated virus in humans. Gene Ther. 1999, 6, 1074–1083. [Google Scholar] [CrossRef] [PubMed]
- Nwanegbo, E.; Vardas, E.; Gao, W.; Whittle, H.; Sun, H.; Rowe, D.; Robbins, P.D.; Gambotto, A. Prevalence of Neutralizing Antibodies to Adenoviral Serotypes 5 and 35 in the Adult Populations of The Gambia, South Africa, and the United States. Clin. Diagn. Lab. Immunol. 2004, 11, 351–357. [Google Scholar] [CrossRef] [PubMed]
- Xiang, Z.; Li, Y.; Cun, A.; Yang, W.; Ellenberg, S.; Switzer, W.M.; Kalish, M.L.; Ertl, H. Chimpanzee Adenovirus Antibodies in Humans, Sub-Saharan Africa. Emerg. Infect. Dis. 2006, 12, 1596–1599. [Google Scholar] [CrossRef] [PubMed]
- Santibanez, T.A.; Kahn, K.E.; Zhai, Y.; O’Halloran, A.; Liu, L.; Bridges, C.B.; Lu, P.-J.; Greby, S.M.; Williams, W.W.; Singleton, J.A. Flu Vaccination Coverage, United States 2015–2016 Influenza Season. In Center Disease Control and Prevention; CDC: Atlanta, GA, USA, 2015. [Google Scholar]
- Grimaud, G.; Georges, A.J.; Mada, A.; Mathiot, C.C.; Daguisy, A.; Bouquety, J.; Garry, P. An Outbreak of Human Semliki Forest Virus Infections in Central African Republic. Am. J. Trop. Med. Hyg. 1990, 42, 386–393. [Google Scholar]
- Aguilar, P.V.; Estrada-Franco, J.G.; Navarro-Lopez, R.; Ferro, C.; Haddow, A.D.; Weaver, S.C. Endemic Venezuelan equine encephalitis in the Americas: Hidden under the dengue umbrella. Future Virol. 2011, 6, 721–740. [Google Scholar] [CrossRef]
- Lee, J.S.; Hadjipanayis, A.G.; Parker, M.D. Viral vectors for use in the development of biodefense vaccines. Adv. Drug Deliv. Rev. 2005, 57, 1293–1314. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.; Chang, J. Viral vectors for vaccine applications. Clin. Exp. Vaccine Res. 2013, 2, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Gomme, E.A.; Wanjalla, C.N.; Wirblich, C.; Schnell, M.J. Advances in Virus Research. Adv. Virus Res. 2011, 79, 139–164. [Google Scholar] [PubMed]
- Perrin, P.; Morgeaux, S. Inactivation of DNA by beta-propiolactone. Biologicals 1995, 23, 207–211. [Google Scholar] [CrossRef] [PubMed]
- Lundstrom, K. Alphavirus-Based Vaccines. Viruses 2014, 6, 2392–2415. [Google Scholar] [CrossRef] [PubMed]
- Harui, A.; Suzuki, S.; Kochanek, S.; Mitani, K. Frequency and stability of chromosomal integration of adenovirus vectors. J. Virol. 1999, 73, 6141–6146. [Google Scholar] [PubMed]
- Kobayashi, R.; Kohda, T.; Kataoka, K.; Ihara, H.; Kozaki, S.; Pascual, D.W.; Staats, H.F.; Kiyono, H.; McGhee, J.R.; Fujihashi, K. A novel neurotoxoid vaccine prevents mucosal botulism. J. Immun. 2005, 174, 2190–2195. [Google Scholar] [CrossRef] [PubMed]
- Torii, Y.; Tokumaru, Y.; Kawaguchi, S.; Izumi, N.; Maruyama, S.; Mukamoto, M.; Kozaki, S.; Takahashi, M. Production and immunogenic efficacy of botulinum tetravalent (A, B, E, F) toxoid. Vaccine 2002, 20, 2556–2561. [Google Scholar] [CrossRef]
- Torii, Y.; Sugimoto, N.; Kohda, T.; Kozaki, S.; Morokuma, K.; Horikawa, Y.; Ginnaga, A.; Yamamoto, A.; Takahashi, M. Clinical Study of New Tetravalent (Type A, B, E, and F) Botulinum Toxoid Vaccine Derived from M Toxin in Japan. Jpn. J. Infect. Dis. 2017, 70, 423–429. [Google Scholar] [CrossRef] [PubMed]
- LaPenotiere, H.F.; Clayton, M.A.; Middlebrook, J.L. Expression of a large, nontoxic fragment of botulinum neurotoxin serotype A and its use as an immunogen. Toxicon Off. J. Int. Soc. Toxinol. 1995, 33, 1383–1386. [Google Scholar] [CrossRef]
- Byrne, M.P.; Smith, L.A. Development of vaccines for prevention of botulism. Biochimie 2000, 82, 955–966. [Google Scholar] [CrossRef]
- Webb, R.P.; Smith, L.A. What next for botulism vaccine development? Expert Rev. Vaccines 2013, 12, 481–492. [Google Scholar] [CrossRef] [PubMed]
- Baldwin, M.R.; Tepp, W.H.; Przedpelski, A.; Pier, C.L.; Bradshaw, M.; Johnson, E.A.; Barbieri, J.T. Subunit vaccine against the seven serotypes of botulism. Infect. Immun. 2008, 76, 1314–1318. [Google Scholar] [CrossRef] [PubMed]
- Przedpelski, A.; Tepp, W.H.; Kroken, A.R.; Fu, Z.; Kim, J.J.; Johnson, E.A.; Barbieri, J.T. Enhancing the protective immune response against botulism. Infect. Immun. 2013, 81, 2638–2644. [Google Scholar] [CrossRef] [PubMed]
- Henderson, I.I.; Shearer, J.; Swiderski, W.; Metcalfe, M.; Clagett, K.; Niemuth, N.; Smith, L.A. Protection against botulinum neurotoxin exposure in guinea pigs passively immunized with immune globulin purified from human volunteers vaccinated with recombinant botulinum vaccine (rBV A/B). In Proceedings of the Ninth Annual Conference on Vaccine Research, Baltimore, MD, USA, 21 April 2006. [Google Scholar]
- Shone, C.; Agostini, H.; Clancy, J.; Gu, M.; Yang, H.H.; Chu, Y.; Johnson, V.; Taal, M.; McGlashan, J.; Brehm, J.; et al. Bivalent recombinant vaccine for botulinum neurotoxin types A and B based on a polypeptide comprising their effector and translocation domains that is protective against the predominant A and B subtypes. Infect. Immun. 2009, 77, 2795–2801. [Google Scholar] [CrossRef] [PubMed]
- Binz, T.; Bade, S.; Rummel, A.; Kollewe, A.; Alves, J. Arg(362) and Tyr(365) of the botulinum neurotoxin type a light chain are involved in transition state stabilization. Biochemistry 2002, 41, 1717–1723. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, R.; Binz, T.; Swaminathan, S. Analysis of active site residues of botulinum neurotoxin E by mutational, functional, and structural studies: Glu335Gln is an apoenzyme. Biochemistry 2005, 44, 8291–8302. [Google Scholar] [CrossRef] [PubMed]
- Pier, C.L.; Tepp, W.H.; Bradshaw, M.; Johnson, E.A.; Barbieri, J.T.; Baldwin, M.R. Recombinant holotoxoid vaccine against botulism. Infect. Immun. 2008, 76, 437–442. [Google Scholar] [CrossRef] [PubMed]
- Ravichandran, E.; Janardhanan, P.; Patel, K.; Riding, S.; Cai, S.; Singh, B.R. In Vivo Toxicity and Immunological Characterization of Detoxified Recombinant Botulinum Neurotoxin Type A. Pharm. Res. 2016, 33, 639–652. [Google Scholar] [CrossRef] [PubMed]
- Webb, R.P.; Smith, T.J.; Wright, P.; Brown, J.; Smith, L.A. Production of catalytically inactive BoNT/A1 holoprotein and comparison with BoNT/A1 subunit vaccines against toxin subtypes A1, A2, and A3. Vaccine 2009, 27, 4490–4497. [Google Scholar] [CrossRef] [PubMed]
- Gu, S.; Rumpel, S.; Zhou, J.; Strotmeier, J.; Bigalke, H.; Perry, K.; Shoemaker, C.B.; Rummel, A.; Jin, R. Botulinum neurotoxin is shielded by NTNHA in an interlocked complex. Science 2012, 335, 977–981. [Google Scholar] [CrossRef] [PubMed]
- Skarin, H.; Tevell Aberg, A.; Woudstra, C.; Hansen, T.; Lofstrom, C.; Koene, M.; Bano, L.; Hedeland, M.; Anniballi, F.; De Medici, D.; et al. The workshop on animal botulism in Europe. Biosecur. Bioterror. 2013, 11 (Suppl 1), S183–S190. [Google Scholar] [CrossRef]
- Kruger, M.; Skau, M.; Shehata, A.A.; Schrodl, W. Efficacy of Clostridium botulinum types C and D toxoid vaccination in Danish cows. Anaerobe 2013, 23, 97–101. [Google Scholar] [CrossRef] [PubMed]
- Ireland, J.L.; McGorum, B.C.; Proudman, C.J.; Newton, J.R. Designing a field trial of an equine grass sickness vaccine: A questionnaire-based feasibility study. Vet. J. 2016, 213, 64–71. [Google Scholar] [CrossRef] [PubMed]
- Takeda, M.; Kasai, H.; Torii, Y.; Mukamoto, M.; Kohda, T.; Tsukamoto, K.; Kozaki, S. Protective effect of botulinum C/D mosaic toxoid against avian botulism. J. Vet. Med. Sci. 2006, 68, 325–330. [Google Scholar] [CrossRef] [PubMed]
- Johnson, A.L.; Sweeney, R.W.; McAdams, S.C.; Whitlock, R.H. Quantitative real-time PCR for detection of the neurotoxin gene of Clostridium botulinum type B in equine and bovine samples. Vet. J. 2012, 194, 118–120. [Google Scholar] [CrossRef] [PubMed]
- Johnson, A.L.; McAdams-Gallagher, S.C.; Sweeney, R.W. Quantitative real-time PCR for detection of neurotoxin genes of Clostridium botulinum types A, B and C in equine samples. Vet. J. 2014, 199, 157–161. [Google Scholar] [CrossRef] [PubMed]
- Steinman, A.; Galon, N.; Arazi, A.; Bar-Giora, Y.; Shpigel, N.Y. Cattle immune response to botulinum type D toxoid: Results of a vaccination study. Vaccine 2007, 25, 7636–7640. [Google Scholar] [CrossRef] [PubMed]
- Cunha, C.E.; Moreira, G.M.; Salvarani, F.M.; Neves, M.S.; Lobato, F.C.; Dellagostin, O.A.; Conceicao, F.R. Vaccination of cattle with a recombinant bivalent toxoid against botulism serotypes C and D. Vaccine 2014, 32, 214–216. [Google Scholar] [CrossRef] [PubMed]
- Killeen, K.P.; Escuyer, V.; Mekalanos, J.J.; Collier, R.J. Reversion of recombinant toxoids: Mutations in diphtheria toxin that partially compensate for active-site deletions. Proc. Natl. Acad. Sci. USA 1992, 89, 6207–6209. [Google Scholar] [CrossRef] [PubMed]
- Bruno, L.; Cortese, M.; Rappuoli, R.; Merola, M. Lessons from Reverse Vaccinology for viral vaccine design. Curr. Opin. Virol. 2015, 11, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Fieldhouse, R.J.; Turgeon, Z.; White, D.; Merrill, A.R. Cholera- and anthrax-like toxins are among several new ADP-ribosyltransferases. PLoS Comput. Biol. 2010, 6, e1001029. [Google Scholar] [CrossRef] [PubMed]
- Mansfield, M.J.; Adams, J.B.; Doxey, A.C. Botulinum neurotoxin homologs in non-Clostridium species. FEBS Lett. 2015, 589, 342–348. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
| Material | Vaccine Vector | Vaccine Properties | Comments | ||
|---|---|---|---|---|---|
| Nucleic acid | Plasmid | HC-based vaccine |
| ||
| Plasmid DNA replicon | Semliki Forest Virus genome with HC replacing the structural proteins |
| |||
| Viral Particle | Virus | Genome | Attenuated/Inactivation | ||
| Adenovirus | Linear dsDNA | Inactivated—lacks the E1 gene |
| ||
| Influenza | Segmented -ssRNA | Attenuated |
| ||
| Rabies | -ssRNA | Attenuated and inactivated by β-Propiolactone |
| ||
| Alphaviruses | Semliki Forest | +ssRNA | Inactivated—lacks the genes for structural proteins |
| |
| Venezuelan Equine Encephalitis |
| ||||
| Protein | Toxoid | Chemical inactivation |
| ||
| Recombinant HC | Receptor binding domain |
| |||
| Recombinant LHN | Light chain and Translocation domain |
| |||
| Recombinant BoNT | Light chain and Heavy chain |
| |||
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sundeen, G.; Barbieri, J.T. Vaccines against Botulism. Toxins 2017, 9, 268. https://doi.org/10.3390/toxins9090268
Sundeen G, Barbieri JT. Vaccines against Botulism. Toxins. 2017; 9(9):268. https://doi.org/10.3390/toxins9090268
Chicago/Turabian StyleSundeen, Grace, and Joseph T. Barbieri. 2017. "Vaccines against Botulism" Toxins 9, no. 9: 268. https://doi.org/10.3390/toxins9090268
APA StyleSundeen, G., & Barbieri, J. T. (2017). Vaccines against Botulism. Toxins, 9(9), 268. https://doi.org/10.3390/toxins9090268