
Enzymatic and Pro-Inflammatory Activities of Bothrops lanceolatus Venom: Relevance for Envenomation

Abstract
:1. Introduction
2. Results
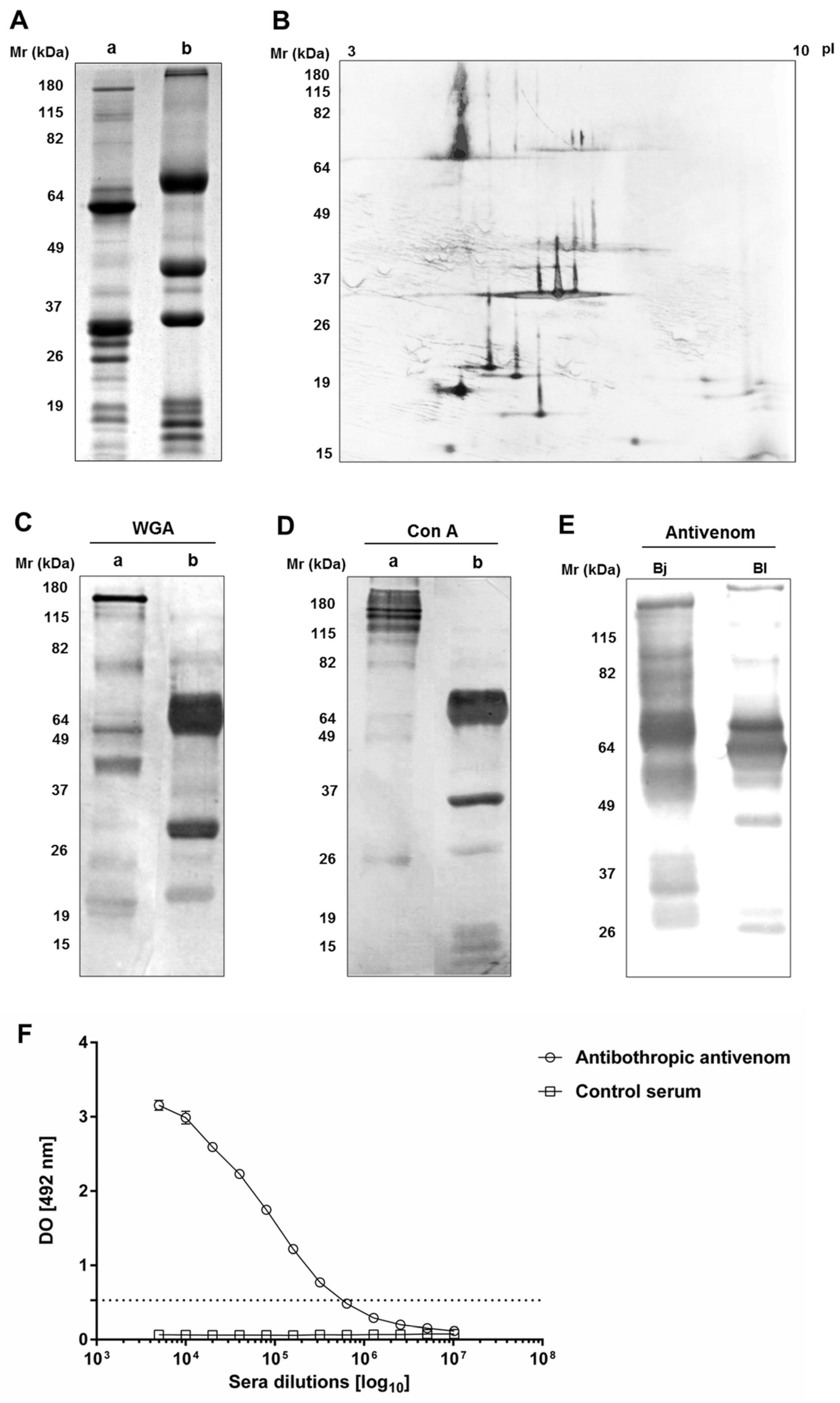
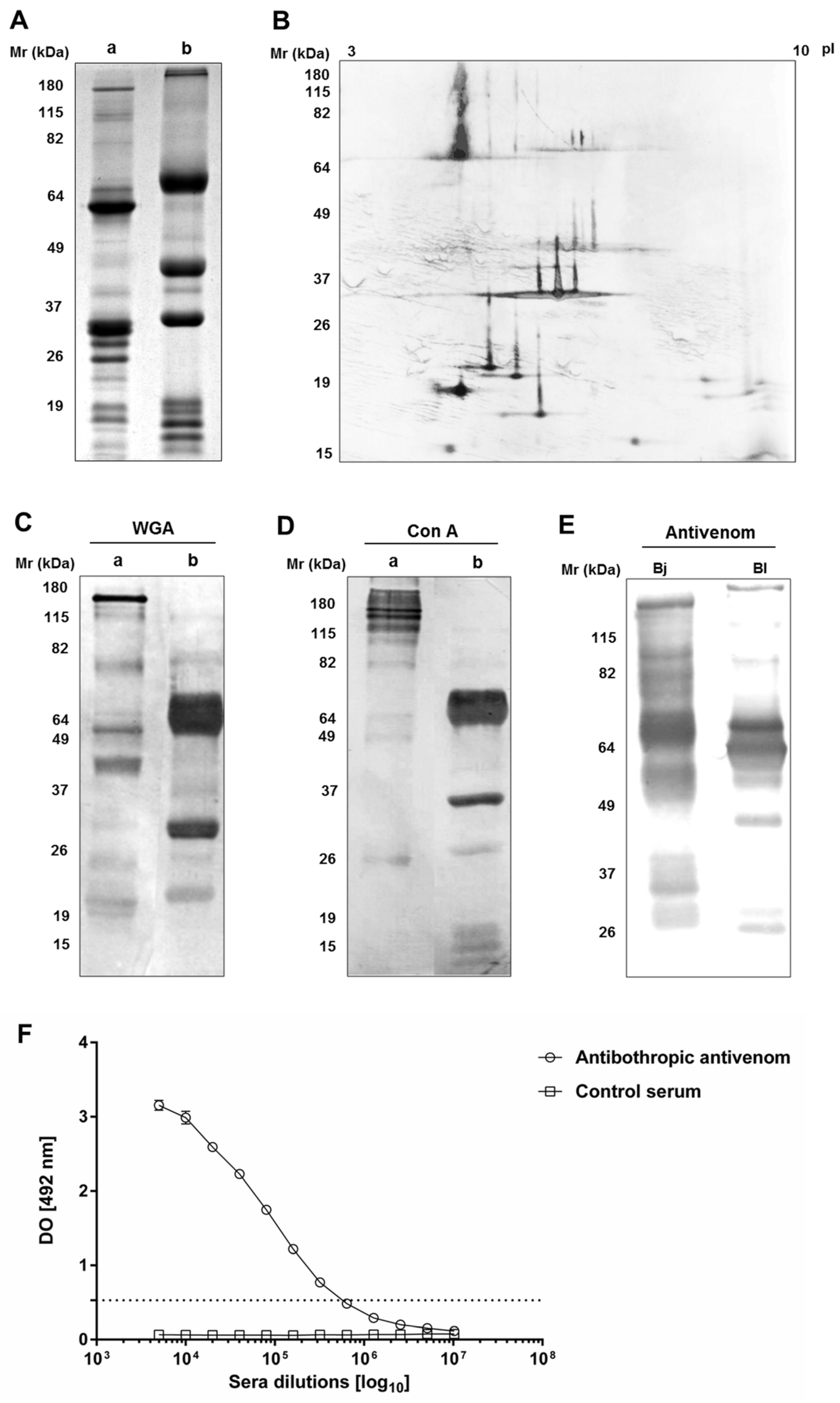
2.1. Immunochemical Characterization and Cross-Reactivity of B. lanceolatus Venom
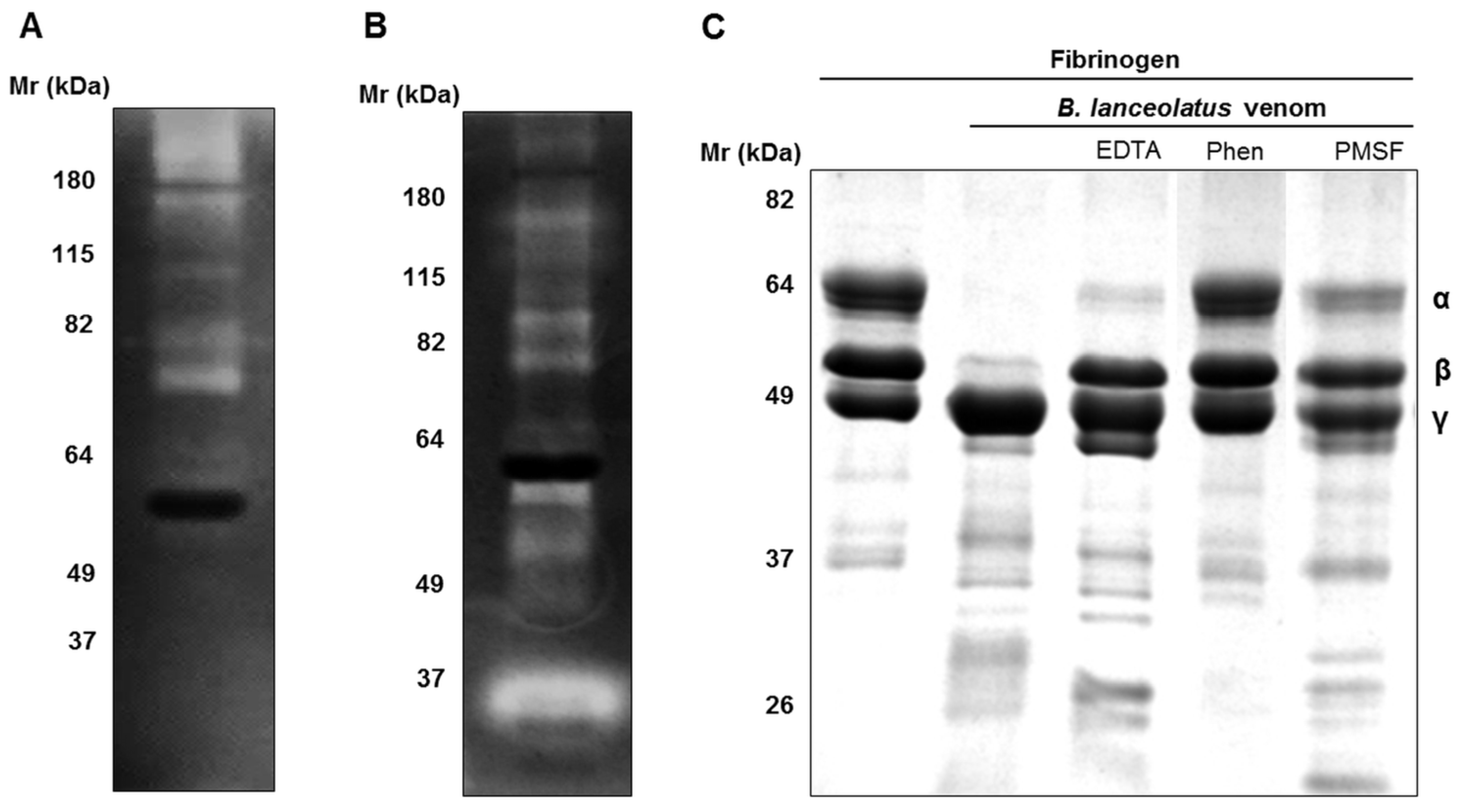
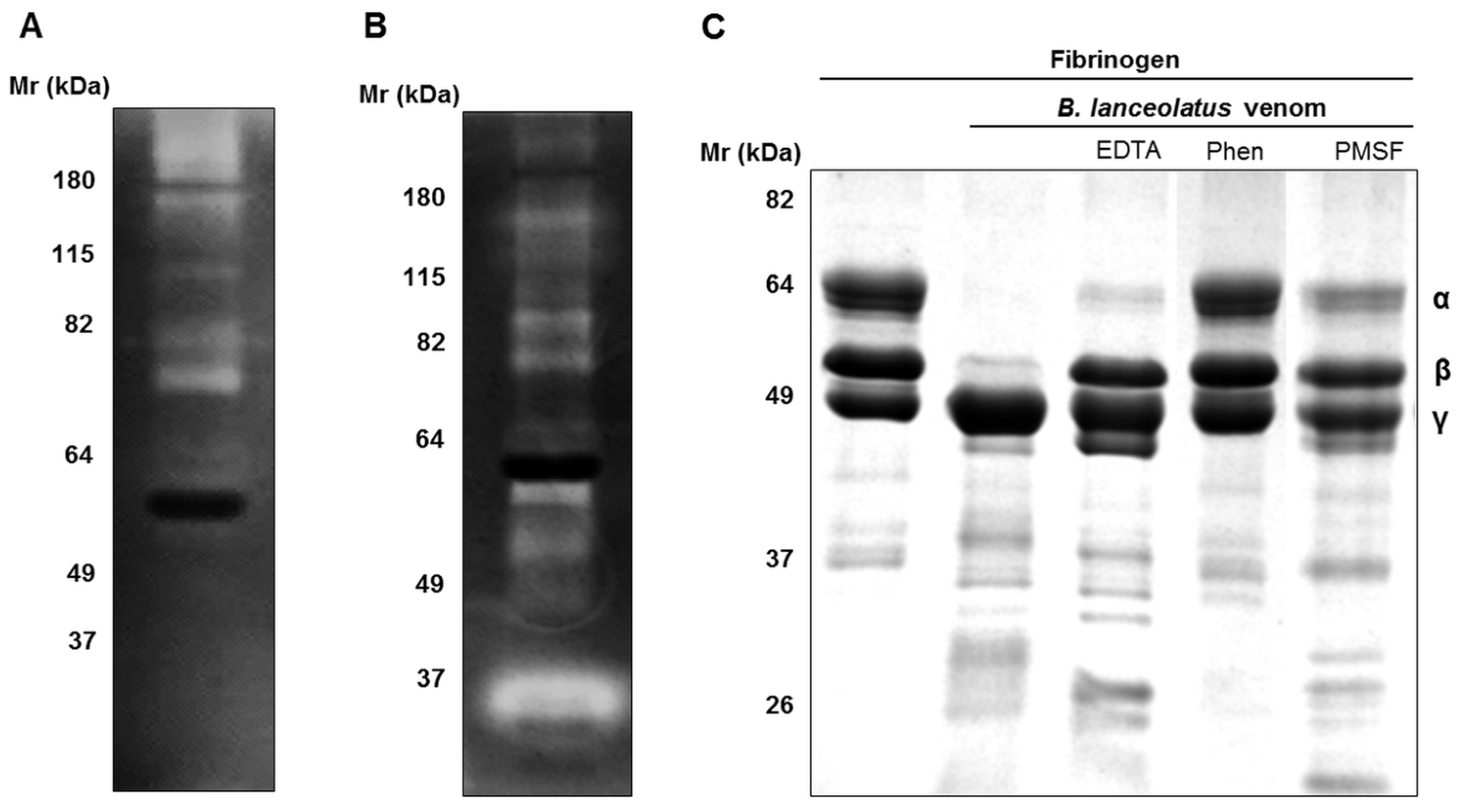
2.2. Enzymatic Activities of B. lanceolatus Venom
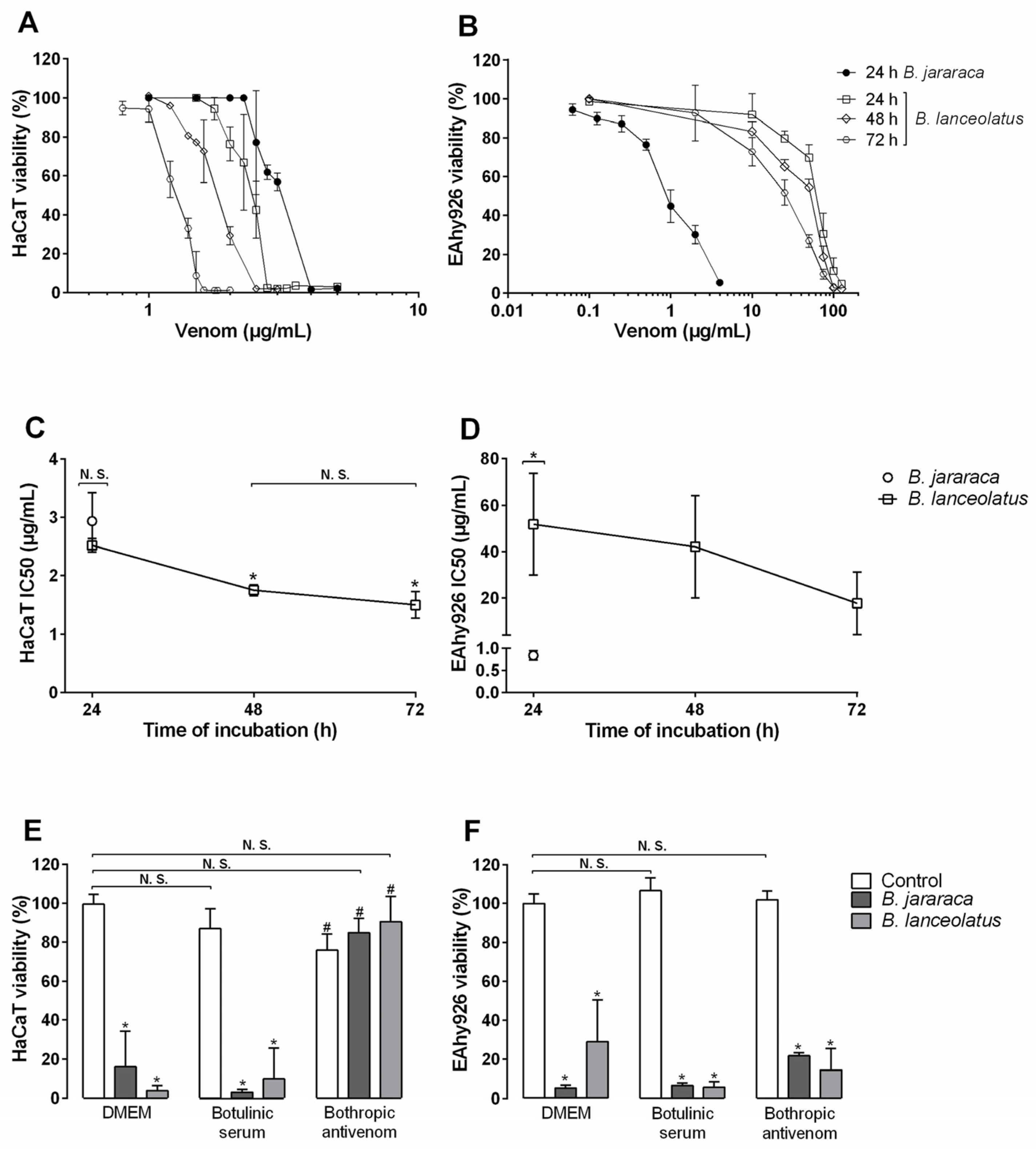
2.3. Cytotoxicity of the Venom for Human Cells
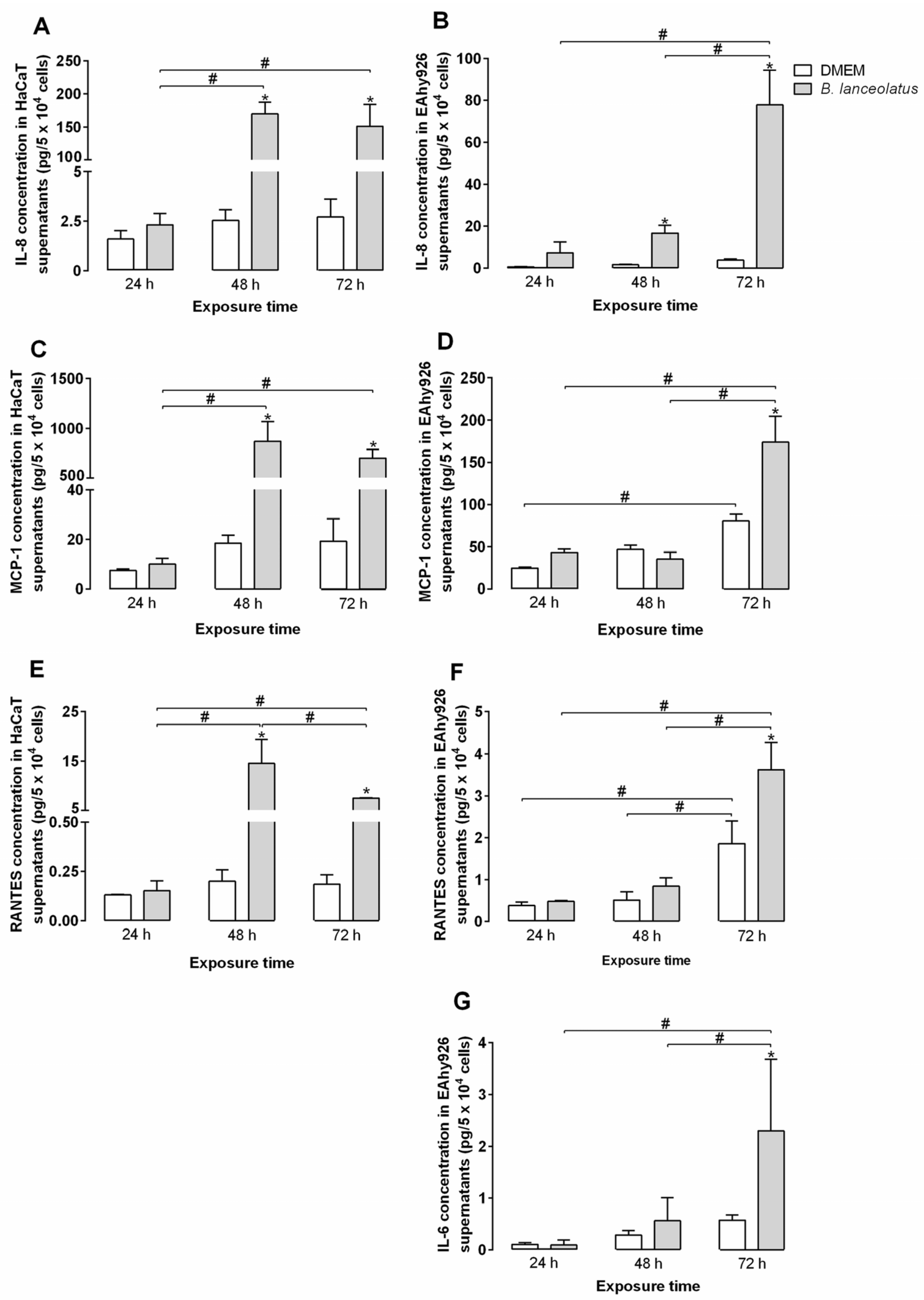
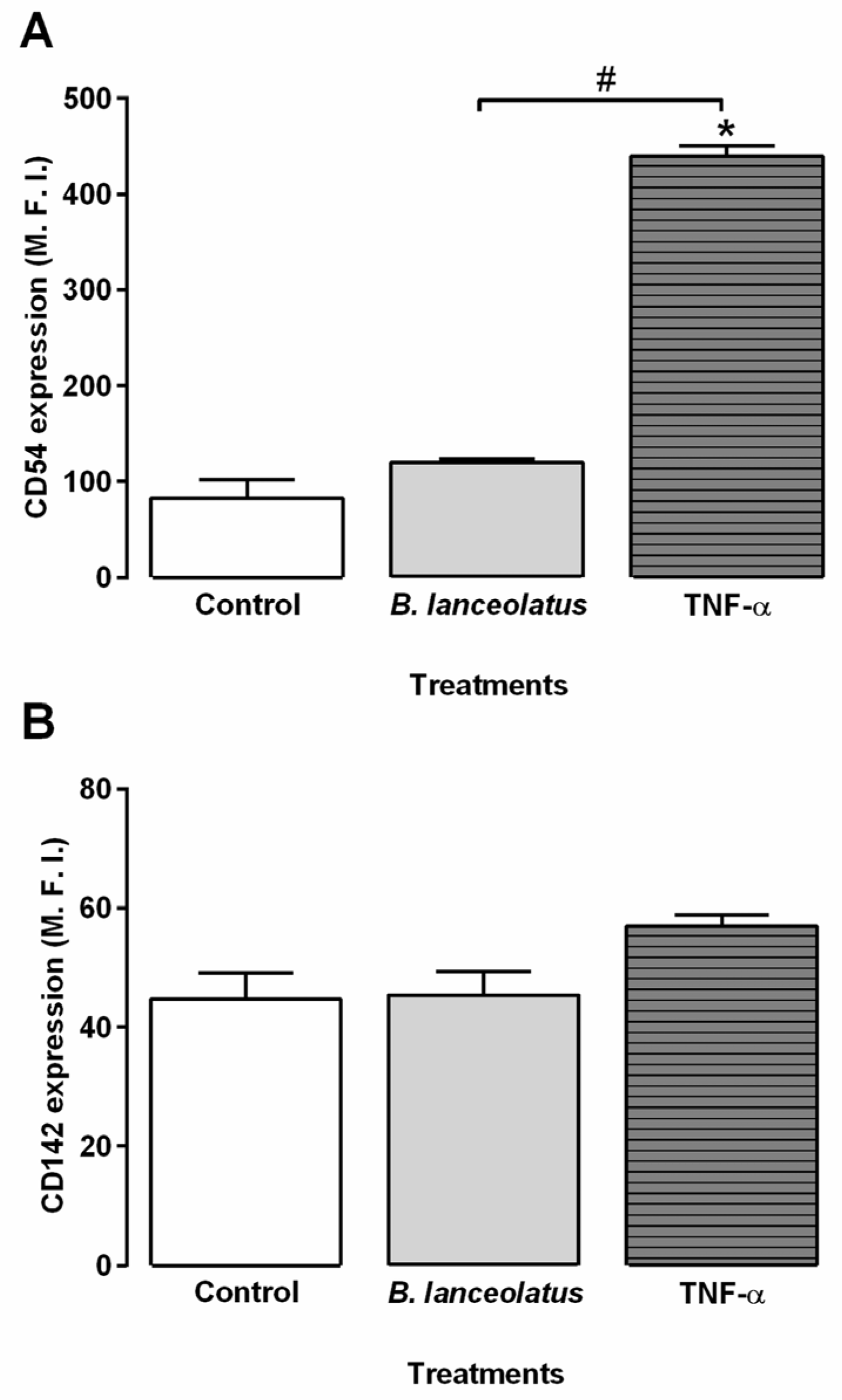
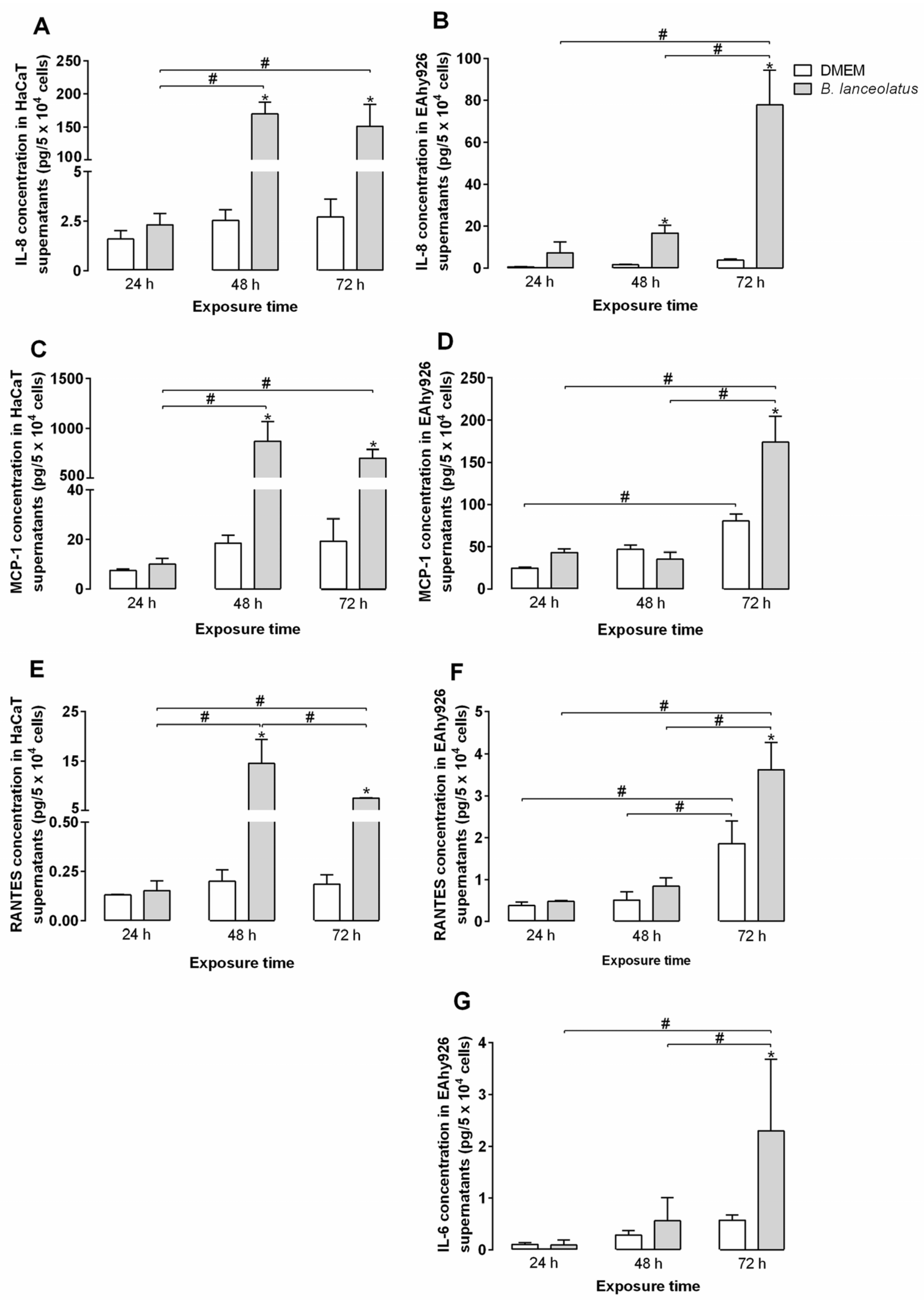
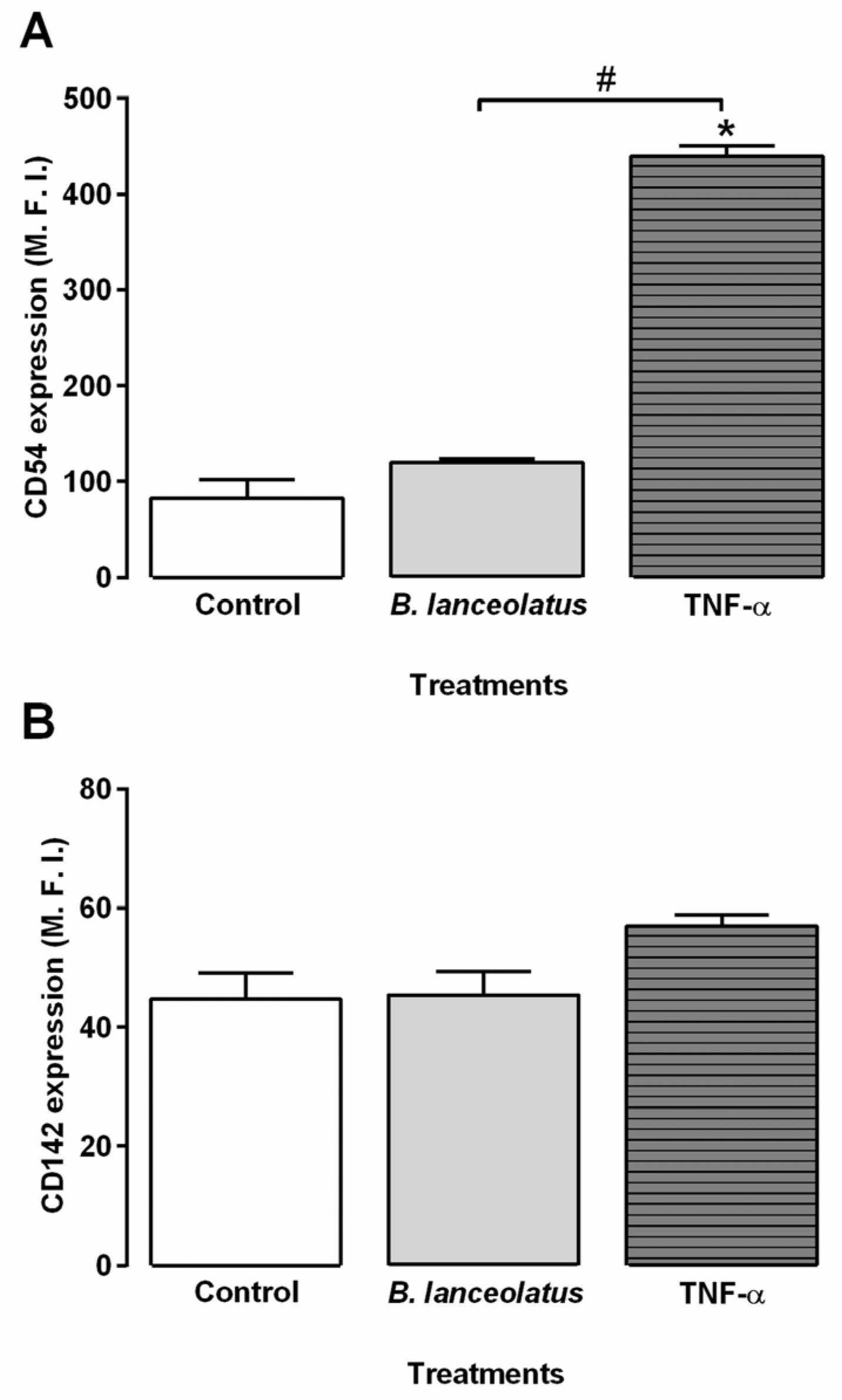
2.4. Inflammatory Potential of the Venom for Human Keratinocytes and Vascular Endothelial Cells
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Chemicals and Reagents
5.2. Venoms
5.3. Sera
5.4. Unidimensional Electrophoresis and Western Blotting
5.5. Bi-Dimensional Electrophoresis
5.6. Enzyme Linked Immunosorbent Assay (ELISA)
5.7. Phospholipase A2 Activity
5.8. Hyaluronidase Activity
5.9. Cleavage of Fibrinogen
5.10. Zymography
5.11. Fluorescent Peptidic Substrates
5.12. Neutralization of B. lanceolatus Venom Enzymatic Activities
5.13. Cytoxicity in Human Keratinocytes and Vascular Endothelial Cells
5.14. Neutralization of Venom Cytotoxicity by Bothropic Antivenom
5.15. Cytokine and Chemokine Release by Venom-Treated Cells
5.16. Expression of Membrane Proteins by Vascular Endothelial Cells Exposed to the Venom
5.17. Statistics
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
References
- França, F.O.S.; Málaque, C.M.S. Acidente botrópico. In Animais Peçonhentos no Brasil: Biologia, Clínica e Terapêutica dos Acidentes; Cardoso, J.L.C., França, F.O.S., Wen, F.H., Málaque, C.M.S., Haddad, V., Jr., Eds.; Sarvier: São Paulo, Brazil, 2003; pp. 72–86. [Google Scholar]
- Warrell, D.A. Snakebites in Central and South America: Epidemiology, clinical features, and clinical management. In The Venomous Reptiles of the Western Hemisphere; Campbell, J.A., Lamar, W.W., Eds.; Comstock Publishers/Cornell University Press: Ithaca, NY, USA, 2004; Volume 2, pp. 709–761. [Google Scholar]
- Resiere, D.; Mégarbane, B.; Valentino, R.; Mehdaoui, H.; Thomas, L. Bothrops lanceolatus bites: Guidelines for severity assessment and emergent management. Toxins 2010, 2, 163–173. [Google Scholar] [CrossRef] [PubMed]
- Thomas, L.; Chausson, N.; Uzan, J.; Kaidomar, S.; Vignes, R.; Plumelle, Y.; Bucher, B.; Smadja, D. Thrombotic stroke following snake bites by the ‘Fer-de-Lance’ Bothrops lanceolatus in Martinique despite antivenom treatment: A report of three recent cases. Toxicon 2006, 48, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Malbranque, S.; Piercecchi-Marti, M.D.; Thomas, L.; Barbey, C.; Courcier, D.; Bucher, B.; Ridarch, A.; Smadja, D.; Warrell, D.A. Fatal diffuse thrombotic microangiopathy after a bite by the ‘Fer-de-Lance’ pit viper (Bothrops lanceolatus) of Martinique. Am. J. Trop. Med. Hyg. 2008, 78, 856–861. [Google Scholar] [PubMed]
- Gutiérrez, J.M.; Sanz, L.; Escolano, J.; Fernández, J.; Lomonte, B.; Angulo, Y.; Rucavado, A.; Warrell, D.A.; Calvete, J.J. Snake venomics of the Lesser Antillean pit vipers Bothrops caribbaeus and Bothrops lanceolatus: Correlation with toxicological activities and immunoreactivity of a heterologous antivenom. J. Proteome Res. 2008, 7, 4396–4408. [Google Scholar] [CrossRef] [PubMed]
- Terra, R.M.S.; Pinto, A.F.M.; Guimarães, J.A.; Fox, J.W. Proteomic profiling of snake venom metalloproteinases (SVMPs): Insights into venom induced pathology. Toxicon 2009, 54, 836–844. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, K.C.; Da Silva, M.J.; Costa, G.G.L.; Torres, T.T.; Del Bem, L.E.V.; Vidal, R.O.; Menossi, M.; Hyslop, S. A transcriptomic analysis of gene expression in the venom gland of the snake Bothrops alternatus (urutu). BMC Genom. 2010, 11, 605. [Google Scholar] [CrossRef] [PubMed]
- Ramos, O.H.P.; Selistre-de-Araujo, H.S. Snake venom metalloproteases—Structure and function of catalytic and disintegrin domains. Comp. Biochem. Physiol. Toxicol. Pharmacol. 2006, 142, 328–346. [Google Scholar] [CrossRef] [PubMed]
- Markland, F.S. Snake venoms and the hemostatic system. Toxicon 1998, 36, 1749–1800. [Google Scholar] [CrossRef]
- Fox, J.W.; Serrano, S.M.T. Insights into and speculations about snake venom metalloproteinase (SVMP) synthesis, folding and disulfide bond formation and their contribution to venom complexity. FEBS J. 2008, 275, 3016–3030. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, J.M.; Rucavado, A.; Escalante, T.; Díaz, C. Hemorrhage induced by snake venom metalloproteinases: Biochemical and biophysical mechanisms involved in microvessel damage. Toxicon 2005, 45, 997–1011. [Google Scholar] [CrossRef] [PubMed]
- Serrano, S.M.T.; Maroun, R.C. Snake venom serine proteinases: Sequence homology vs. substrate specificity, a paradox to be solved. Toxicon 2005, 45, 1115–1132. [Google Scholar] [CrossRef] [PubMed]
- Kini, R.M. Excitement ahead: Structure, function and mechanism of snake venom phospholipase A2 enzymes. Toxicon 2003, 42, 827–840. [Google Scholar] [CrossRef] [PubMed]
- De Araujo, A.L.; Radvanyi, F.; Bon, C. Purification of an acidic phospholipase A2 from Bothrops lanceolatus (fer de lance) venom: Molecular and enzymatic properties. Toxicon 1994, 32, 1069–1081. [Google Scholar] [CrossRef]
- Lôbo de Araújo, A.; Donato, J.L.; Bon, C. Purification from Bothrops lanceolatus (fer de lance) venom of a fibrino(geno)lytic enzyme with esterolytic activity. Toxicon 1998, 36, 745–758. [Google Scholar] [CrossRef]
- Stroka, A.; Donato, J.L.; Bon, C.; Hyslop, S.; de Araújo, A.L. Purification and characterization of a hemorrhagic metalloproteinase from Bothrops lanceolatus (Fer-de-lance) snake venom. Toxicon 2005, 45, 411–420. [Google Scholar] [CrossRef] [PubMed]
- De Araújo, A.L.; de Souza, A.O.; da Cruz-Höfling, M.A.; Flores, C.A.; Bon, C. Bothrops lanceolatus (Fer de lance) venom induces edema formation and increases vascular permeability in the mouse hind paw. Toxicon 2000, 38, 209–221. [Google Scholar] [CrossRef]
- De Faria, L.; Antunes, E.; Bon, C.; de Araújo, A.L. Pharmacological characterization of the rat paw edema induced by Bothrops lanceolatus (Fer de lance) venom. Toxicon 2001, 39, 825–830. [Google Scholar] [CrossRef]
- Arruda, V.A.; de Queiroz Guimarães, A.; Hyslop, S.; de Araújo, P.M.F.; Bon, C.; de Araújo, A.L. Bothrops lanceolatus (Fer de lance) venom stimulates leukocyte migration into the peritoneal cavity of mice. Toxicon 2003, 41, 99–107. [Google Scholar] [CrossRef]
- Guimarães, A.Q.; Cruz-Höfling, M.A.; Ferreira de Araújo, P.M.; Bon, C.; de Araújo, A.L. Pharmacological and histopathological characterization of Bothrops lanceolatus (Fer de lance) venom-induced edema. Inflamm. Res. 2004, 53, 284–291. [Google Scholar] [CrossRef] [PubMed]
- Bogarín, G.; Romero, M.; Rojas, G.; Lutsch, C.; Casadamont, M.; Lang, J.; Otero, R.; Gutiérrez, J.M. Neutralization, by a monospecific Bothrops lanceolatus antivenom, of toxic activities induced by homologous and heterologous Bothrops snake venoms. Toxicon 1999, 37, 551–557. [Google Scholar] [CrossRef]
- De Araújo, A.L.; Kamiguti, A.; Bon, C. Coagulant and anticoagulant activities of Bothrops lanceolatus (Fer de lance) venom. Toxicon 2001, 39, 371–375. [Google Scholar] [CrossRef]
- Trummal, K.; Tõnismägi, K.; Paalme, V.; Järvekülg, L.; Siigur, J.; Siigur, E. Molecular diversity of snake venom nerve growth factors. Toxicon 2011, 58, 363–368. [Google Scholar] [CrossRef] [PubMed]
- Lidington, E.A.; Moyes, D.L.; McCormack, A.M.; Rose, M.L. A comparison of primary endothelial cells and endothelial cell lines for studies of immune interactions. Transplant. Immunol. 1999, 7, 239–246. [Google Scholar] [CrossRef]
- Paes Leme, A.F.; Kitano, E.S.; Furtado, M.F.; Valente, R.H.; Camargo, A.C.M.; Ho, P.L.; Fox, J.W.; Serrano, S.M.T. Analysis of the subproteomes of proteinases and heparin-binding toxins of eight Bothrops venoms. Proteomics 2009, 9, 733–745. [Google Scholar] [CrossRef] [PubMed]
- Zelanis, A.; Serrano, S.M.T.; Reinhold, V.N. N-glycome profiling of Bothrops jararaca newborn and adult venoms. J. Proteom. 2012, 75, 774–782. [Google Scholar] [CrossRef] [PubMed]
- Zelanis, A.; Tashima, A.K.; Rocha, M.M.T.; Furtado, M.F.; Camargo, A.C.M.; Ho, P.L.; Serrano, S.M.T. Analysis of the ontogenetic variation in the venom proteome/peptidome of Bothrops jararaca reveals different strategies to deal with prey. J. Proteome Res. 2010, 9, 2278–2291. [Google Scholar] [CrossRef] [PubMed]
- Girish, K.S.; Kemparaju, K. The magic glue hyaluronan and its eraser hyaluronidase: A biological overview. Life Sci. 2007, 80, 1921–1943. [Google Scholar] [CrossRef] [PubMed]
- Queiroz, G.P.; Pessoa, L.A.; Portaro, F.C.V.; de Furtado, M.F.D.; Tambourgi, D.V. Interspecific variation in venom composition and toxicity of Brazilian snakes from Bothrops genus. Toxicon 2008, 52, 842–851. [Google Scholar] [CrossRef] [PubMed]
- Clissa, P.B.; Laing, G.D.; Theakston, R.D.; Mota, I.; Taylor, M.J.; Moura-da-Silva, A.M. The effect of jararhagin, a metalloproteinase from Bothrops jararaca venom, on pro-inflammatory cytokines released by murine peritoneal adherent cells. Toxicon 2001, 39, 1567–1573. [Google Scholar] [CrossRef]
- Tensen, C.P.; Flier, J.; Van Der Raaij-Helmer, E.M.; Sampat-Sardjoepersad, S.; Van Der Schors, R.C.; Leurs, R.; Scheper, R.J.; Boorsma, D.M.; Willemze, R. Human IP-9: A keratinocyte-derived high affinity CXC-chemokine ligand for the IP-10/Mig Receptor (CXCR3). J. Investig. Dermatol. 1999, 112, 716–722. [Google Scholar] [CrossRef] [PubMed]
- Kuniyoshi, A.K.; Rocha, M.; Cajado Carvalho, D.; Juliano, M.A.; Juliano Neto, L.; Tambourgi, D.V.; Portaro, F.C.V. Angiotensin-degrading serine peptidase: A new chymotrypsin-like activity in the venom of Bothrops jararaca partially blocked by the commercial antivenom. Toxicon 2012, 59, 124–131. [Google Scholar] [CrossRef] [PubMed]
- Lomonte, B.; Lundgren, J.; Johansson, B.; Bagge, U. The dynamics of local tissue damage induced by Bothrops asper snake venom and myotoxin II on the mouse cremaster muscle: An intravital and electron microscopic study. Toxicon 1994, 32, 41–55. [Google Scholar] [CrossRef]
- Lomonte, B.; Gutiérrez, J.M.; Borkow, G.; Ovadia, M.; Tarkowski, A.; Hanson, L.A. Activity of hemorrhagic metalloproteinase BaH-1 and myotoxin II from Bothrops asper snake venom on capillary endothelial cells in vitro. Toxicon 1994, 32, 505–510. [Google Scholar] [CrossRef]
- Borkow, G.; Gutiérrez, J.M.; Ovadia, M. Inhibition of toxic activities of Bothrops asper venom and other crotalid snake venoms by a novel neutralizing mixture. Toxicol. Appl. Pharmacol. 1997, 147, 442–447. [Google Scholar] [CrossRef] [PubMed]
- Lomonte, B.; Angulo, Y.; Rufini, S.; Cho, W.; Giglio, J.R.; Ohno, M.; Daniele, J.J.; Geoghegan, P.; Gutiérrez, J.M. Comparative study of the cytolytic activity of myotoxic phospholipases A2 on mouse endothelial (tEnd) and skeletal muscle (C2C12) cells in vitro. Toxicon 1999, 37, 145–158. [Google Scholar] [CrossRef]
- De Carvalho, D.D.; Schmitmeier, S.; Novello, J.C.; Markland, F.S. Effect of BJcuL (a lectin from the venom of the snake Bothrops jararacussu) on adhesion and growth of tumor and endothelial cells. Toxicon 2001, 39, 1471–1476. [Google Scholar] [CrossRef]
- Díaz, C.; Valverde, L.; Brenes, O.; Rucavado, A.; Gutiérrez, J.M. Characterization of events associated with apoptosis/anoikis induced by snake venom metalloproteinase BaP1 on human endothelial cells. J. Cell. Biochem. 2005, 94, 520–528. [Google Scholar] [CrossRef] [PubMed]
- Schattner, M.; Fritzen, M.; de Ventura, J.S.; de Albuquerque Modesto, J.C.; Pozner, R.G.; Moura-da-Silva, A.M.; Chudzinski-Tavassi, A.M. The snake venom metalloproteases berythractivase and jararhagin activate endothelial cells. Biol. Chem. 2005, 386, 369–374. [Google Scholar] [CrossRef] [PubMed]
- Pereira, A.L.M.; Fritzen, M.; Faria, F.; da Motta, G.; Chudzinski-Tavassi, A.M. Releasing or expression modulating mediator involved in hemostasis by berythractivase and jararhagin (SVMPs). Toxicon 2006, 47, 788–796. [Google Scholar] [CrossRef] [PubMed]
- Brenes, O.; Muñóz, E.; Roldán-Rodríguez, R.; Díaz, C. Cell death induced by Bothrops asper snake venom metalloproteinase on endothelial and other cell lines. Exp. Mol. Pathol. 2010, 88, 424–432. [Google Scholar] [CrossRef] [PubMed]
- Lopes, D.S.; Faquim-Mauro, E.; Magalhães, G.S.; Lima, I.C.; Baldo, C.; Fox, J.W.; Moura-da-Silva, A.M.; Clissa, P.B. Gene expression of inflammatory mediators induced by jararhagin on endothelial cells. Toxicon 2012, 60, 1072–1084. [Google Scholar] [CrossRef] [PubMed]
- Ache, D.C.; Gomes, M.S.R.; de Souza, D.L.N.; Silva, M.A.; Brandeburgo, M.I.H.; Yoneyama, K.A.G.; Rodrigues, R.S.; Borges, M.H.; Lopes, D.S.; de Rodrigues, V.M. Biochemical properties of a new PI SVMP from Bothrops pauloensis: Inhibition of cell adhesion and angiogenesis. Int. J. Biol. Macromol. 2015, 72, 445–453. [Google Scholar] [CrossRef] [PubMed]
- Bustillo, S.; Garcia-Denegri, M.E.; Gay, C.; Van de Velde, A.C.; Acosta, O.; Angulo, Y.; Lomonte, B.; Gutiérrez, J.M.; Leiva, L. Phospholipase A(2) enhances the endothelial cell detachment effect of a snake venom metalloproteinase in the absence of catalysis. Chem. Biol. Interact. 2015, 240, 30–36. [Google Scholar] [CrossRef] [PubMed]
- Jiménez, N.; Escalante, T.; Gutierrez, J.M.; Rucavado, A. Skin pathology induced by snake venom metalloproteinase: Acute damage, revascularization, and re-epithelization in a mouse ear model. J. Investig. Dermatol. 2008, 128, 2421–2428. [Google Scholar] [CrossRef] [PubMed]
- Aranda-Souza, M.A.; Rossato, F.A.; Costa, R.A.P.; Figueira, T.R.; Castilho, R.F.; Guarniere, M.C.; Nunes, E.S.; Coelho, L.C.B.B.; Correia, M.T.S.; Vercesi, A.E. A lectin from Bothrops leucurus snake venom raises cytosolic calcium levels and promotes B16-F10 melanoma necrotic cell death via mitochondrial permeability transition. Toxicon 2014, 82, 97–103. [Google Scholar] [CrossRef] [PubMed]
- Edgell, C.J.; McDonald, C.C.; Graham, J.B. Permanent cell line expressing human factor VIII-related antigen established by hybridization. Proc. Natl. Acad. Sci. USA 1983, 80, 3734–3737. [Google Scholar] [CrossRef] [PubMed]
- Barker, J.; Mitra, R.; Griffiths, C.; Dixit, V.; Nickoloff, B. Keratinocytes as initiators of inflammation. Lancet 1991, 337, 211–214. [Google Scholar] [CrossRef]
- Zamuner, S.R.; Zuliani, J.P.; Fernandes, C.M.; Gutiérrez, J.M.; de Fatima Pereira Teixeira, C. Inflammation induced by Bothrops asper venom: Release of proinflammatory cytokines and eicosanoids, and role of adhesion molecules in leukocyte infiltration. Toxicon 2005, 46, 806–813. [Google Scholar] [CrossRef] [PubMed]
- Gallagher, P.G.; Bao, Y.; Serrano, S.M.T.; Kamiguti, A.S.; Theakston, R.D.G.; Fox, J.W. Use of microarrays for investigating the subtoxic effects of snake venoms: Insights into venom-induced apoptosis in human umbilical vein endothelial cells. Toxicon 2003, 41, 429–440. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Towbin, H.; Staehelin, T.; Gordon, J. Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: Procedure and some applications. Proc. Natl. Acad. Sci. USA 1979, 76, 4350–4354. [Google Scholar] [CrossRef] [PubMed]
- Pukrittayakamee, S.; Warrell, D.A.; Desakorn, V.; McMichael, A.J.; White, N.J.; Bunnag, D. The hyaluronidase activities of some Southeast Asian snake venoms. Toxicon 1988, 26, 629–637. [Google Scholar] [CrossRef]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enzymatic Activities | C. durissus terrificus | B. jararaca | B. lanceolatus |
|---|---|---|---|
| Hyaluronidase | |||
| Specific activity (TRU/mg) | NT | 116.6 ± 8.6 | 20.5 ± 2.4 * |
| Amount of bothropic antivenom (mL/mg of venom) required to inhibit activity by 95% | NT | 0.056 | ≤0.056 |
| Phospholipase | |||
| Specific activity (UF/min/µg) | 461.4 ± 11.7 | 121.9 ± 6.9 # | 231.3 ± 6.4 * |
| Amount of bothropic antivenom (mL/mg of venom) required to inhibit activity by 95% | NT | 0.32 | 2.29 |
| Fluorescent Peptides | |||
|---|---|---|---|
| Abz-FRSSRQ | Abz-RPPGFSPFRQ | ||
| Venom activity (UF/min/µg) | 111.0 ± 4.7 | 78.3 ± 9.6 | |
| Inhibition (%) | EDTA (100 mM) | 0.8 ± 1.7 | 94.1 ± 6.8 ** |
| 1,10-phenanthroline (5 mM) | 4.9 ± 4.6 | 100 ± 0 * | |
| PMSF (5 mM) | 93.5 ± 4.5 * | 79.5 ± 13.8 * | |
| Bothropic antivenom | 33.5 ± 33.6 * | 60.8 ± 21.2 * | |
| Cytokine/Chemokine | |||
|---|---|---|---|
| IL-12p70 | MIG | ||
| Venom activity (pg/µg of venom) | 16.9 ± 9.2 | 3.0 ± 0.2 | |
| Inhibition (%) | EDTA (20 mM) | 97.0 ± 5.9 * | 74.3 ± 7.6 * |
| 1,10-phenanthroline (20 mM) | 86.9 ± 16.1 * | 98.1 ± 7.9 * | |
| PMSF (20 mM) | 62.3 ± 32.2 * | 0.0 ± 0.0 | |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Delafontaine, M.; Villas-Boas, I.M.; Mathieu, L.; Josset, P.; Blomet, J.; Tambourgi, D.V. Enzymatic and Pro-Inflammatory Activities of Bothrops lanceolatus Venom: Relevance for Envenomation. Toxins 2017, 9, 244. https://doi.org/10.3390/toxins9080244
Delafontaine M, Villas-Boas IM, Mathieu L, Josset P, Blomet J, Tambourgi DV. Enzymatic and Pro-Inflammatory Activities of Bothrops lanceolatus Venom: Relevance for Envenomation. Toxins. 2017; 9(8):244. https://doi.org/10.3390/toxins9080244
Chicago/Turabian StyleDelafontaine, Marie, Isadora Maria Villas-Boas, Laurence Mathieu, Patrice Josset, Joël Blomet, and Denise V. Tambourgi. 2017. "Enzymatic and Pro-Inflammatory Activities of Bothrops lanceolatus Venom: Relevance for Envenomation" Toxins 9, no. 8: 244. https://doi.org/10.3390/toxins9080244
APA StyleDelafontaine, M., Villas-Boas, I. M., Mathieu, L., Josset, P., Blomet, J., & Tambourgi, D. V. (2017). Enzymatic and Pro-Inflammatory Activities of Bothrops lanceolatus Venom: Relevance for Envenomation. Toxins, 9(8), 244. https://doi.org/10.3390/toxins9080244