
Tetrodotoxin-Producing Bacteria: Detection, Distribution and Migration of the Toxin in Aquatic Systems

Abstract
:1. Introduction
2. Methods for TTX Detection
3. Ecology of Marine TTX-Producing Bacteria
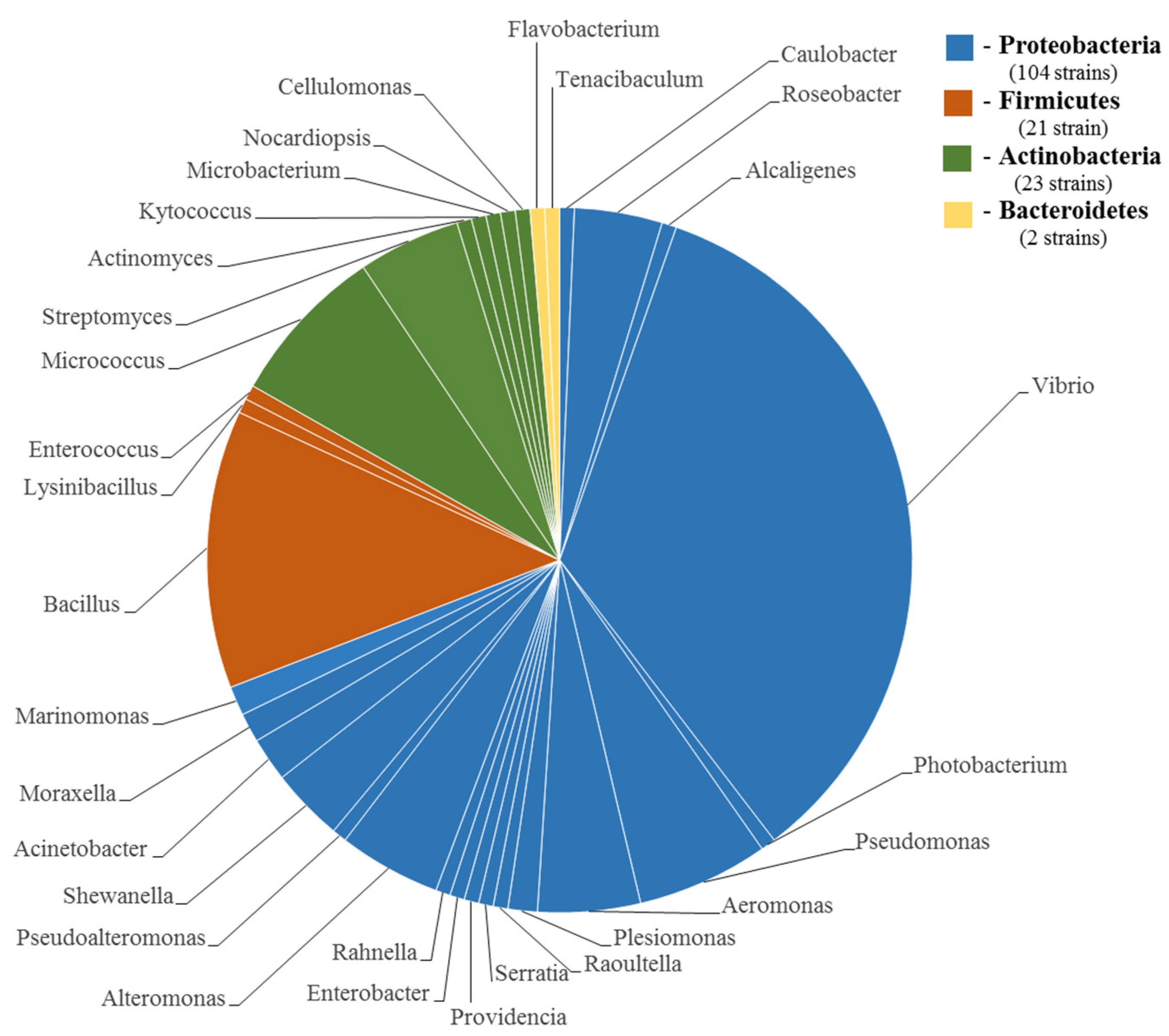
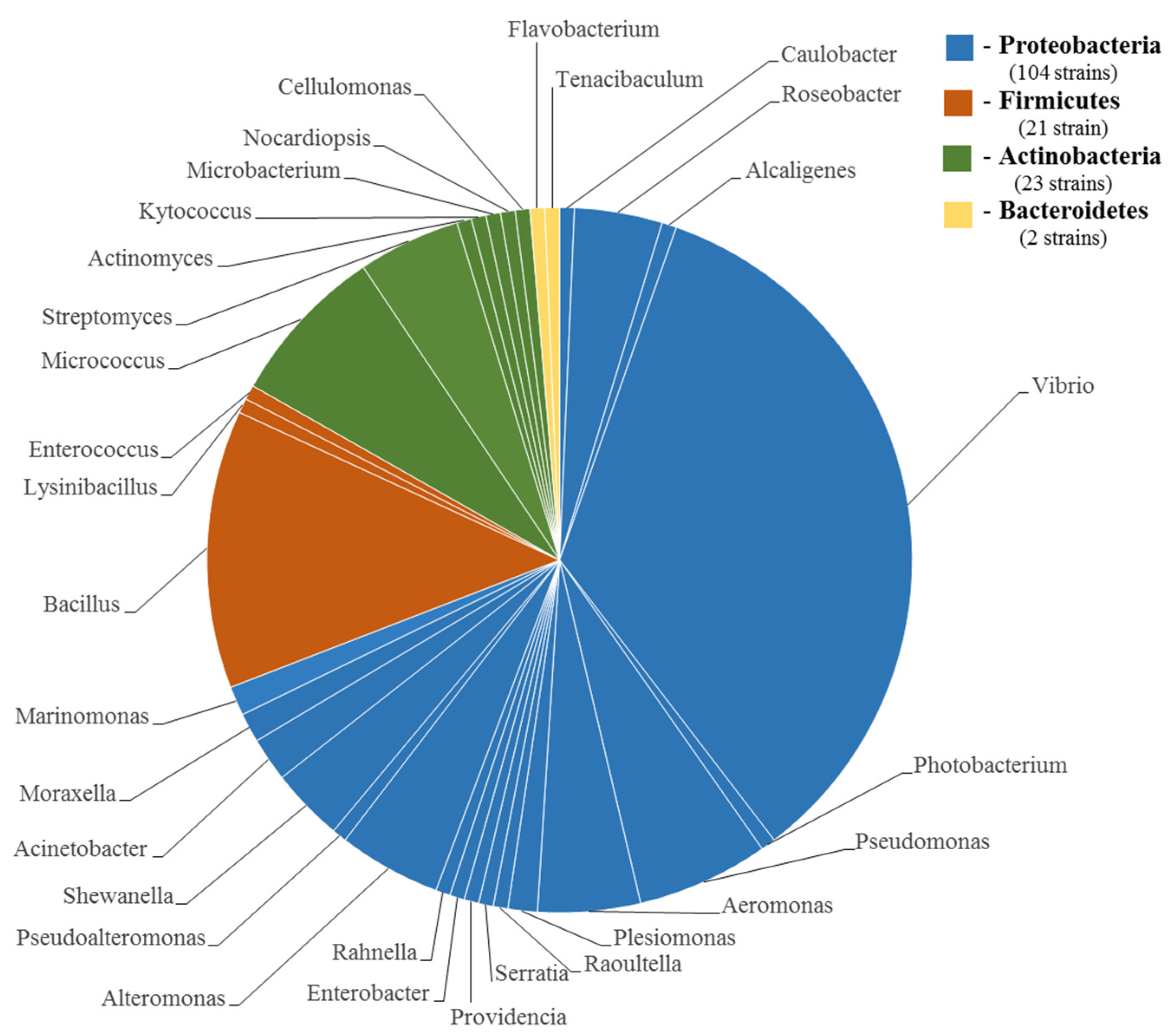
4. Taxonomic Diversity of the TTX-Producing Bacteria
5. TTX-Producing Bacteria as a Promising Source of the Toxin for the Pharmaceutical Industry
6. Discussion
7. Conclusions and Future Perspective
Acknowledgments
Conflicts of Interest
References
- Jal, S.; Khora, S.S. An overview on the origin and production of tetrodotoxin, a potent neurotoxin. J. Appl. Microbiol. 2015, 119, 907–916. [Google Scholar] [CrossRef] [PubMed]
- Bane, V.; Lehane, M.; Dikshit, M.; O’Riordan, A.; Furey, A. Tetrodotoxin: Chemistry, toxicity, source, distribution and detection. Toxins 2014, 6, 693–755. [Google Scholar] [CrossRef] [PubMed]
- Yokoo, A. Chemical studies on pufferfish toxin (3)—Separation of spheroidine. Nippon Kagaku Zasshi 1950, 71, 590–592. [Google Scholar] [CrossRef]
- Brown, M.S.; Mosher, H.S. Tarichatoxin: Isolation and purification. Science 1963, 140, 295–296. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.H.; Brown, G.B.; Mosher, F.A. Tetrodotoxin: Occurrence in atelopid frogs of Costa Rica. Science 1975, 189, 151–152. [Google Scholar] [CrossRef] [PubMed]
- Yasumoto, T.; Yotsu, M.; Murata, M.; Naoki, H. New tetrodotoxin analogs from the newt Cynops ensicauda. J. Am. Chem. Soc. 1988, 110, 2344–2345. [Google Scholar] [CrossRef]
- Sui, L.M.; Chen, K.; Hwang, P.A.; Hwang, D.F. Identification of tetrodotoxin in marine gastropods implicated in food poisoning. J. Nat. Toxins 2002, 11, 213–220. [Google Scholar] [PubMed]
- Huang, H.N.; Lin, J.; Lin, H.L. Identification and quantification of tetradotoxin in the marine gastropod Nassarius by LC-MS. Toxicon 2008, 51, 774–779. [Google Scholar] [CrossRef] [PubMed]
- Noguchi, T.; Arakawa, O. Tetrodotoxin—Distribution and accumulation in aquatic organisms, and cases of human intoxication. Mar. Drugs 2008, 6, 220–242. [Google Scholar] [CrossRef] [PubMed]
- Pratheepa, V.; Vasconcelos, V. Microbial diversity associated with tetrodotoxin production in marine organisms. Environ. Toxicol. Pharmacol. 2013, 36, 1046–1054. [Google Scholar] [CrossRef] [PubMed]
- Noguchi, T.; Mahmud, Y. Current methodologies for detection of tetrodotoxin. J. Toxicol. 2001, 20, 35–50. [Google Scholar] [CrossRef]
- Kogure, K.; Do, H.K.; Thuesen, E.V.; Nanba, K.; Ohwada, K.; Simidu, U. Accumulation of tetrodotoxin in marine sediment. Mar. Ecol. Prog. Ser. 1988, 45, 303–305. [Google Scholar] [CrossRef]
- Watabe, S.; Sato, Y.; Nakaya, M.; Hashimoto, K.; Enomoto, A.; Kaminogawa, S.; Yamauchi, K. Monoclonal antibody raised against tetrodonic acid, a derivative of tetrodotoxin. Toxicon 1989, 27, 265–268. [Google Scholar] [CrossRef]
- Kaufman, B.; Wright, D.C.; Ballou, W.R.; Monheit, D. Protection against tetrodotoxin and saxitoxin intoxication by a cross-protective rabbit anti-tetrodotoxin antiserum. Toxicon 1991, 29, 581–587. [Google Scholar] [CrossRef]
- Stokes, A.N.; Williams, B.L.; French, S.S. An improved competitive inhibition enzymatic immunoassay method for tetrodotoxin quantification. Biol. Proced. Online 2012, 14, 3. [Google Scholar] [CrossRef] [PubMed]
- Kao, C.Y. Structure-activity relations of tetrodotoxin, saxitoxin, and analogues. Ann. N. Y. Acad. Sci. 1986, 479, 52–67. [Google Scholar] [CrossRef] [PubMed]
- Huot, R.I.; Armstrong, D.L.; Chanh, T.C. Protection against nerve toxicity by monoclonal antibodies to the sodium channel blocker tetrodotoxin. J. Clin. Investig. 1989, 83, 1821–1826. [Google Scholar] [CrossRef] [PubMed]
- Turner, A.D.; Higgins, C.; Higman, W.; Hungerford, J. Potential threats posed by tetrodotoxins in UK waters: Examination of detection methodology used in their control. Mar. Drugs 2015, 13, 7357–7376. [Google Scholar] [CrossRef] [PubMed]
- Yasumoto, T.; Michishita, T. Fluorometric determination of tetrodotoxin by high performance liquid chromatography. Agric. Biol. Chem. 1985, 49, 3077–3080. [Google Scholar] [CrossRef]
- Asakawa, M.; Shida, Y.; Miyazawa, K.; Noguchi, T. Instrumental Analysis of Tetrodotoxin. In Chromatography—The Most Versatile Method of Chemical Analysis; Calderon, L.A., Ed.; InTech: Rijeka, Croatia, 2012; pp. 245–270. [Google Scholar]
- Shiomi, K.; Inaoka, H.; Yamanaka, H.; Kikuchi, T. Detection of tetrodotoxin-like compounds in two species of puffer fishes (Lagocephalus lunaris lunaris and Fugu niphobles). Toxicon 1985, 23, 331–336. [Google Scholar] [CrossRef]
- Pratheepa, V.; Alex, A.; Silva, M.; Vasconcelos, V. Bacterial diversity and tetrodotoxin analysis in the viscera of the gastropods from Portuguese coast. Toxicon 2016, 119, 186–193. [Google Scholar] [CrossRef] [PubMed]
- Matsumura, K. Reexamination of tetrodotoxin production by bacteria. Appl. Environ. Microbiol. 1995, 61, 3468–3470. [Google Scholar] [PubMed]
- Matsumura, K. No ability to produce tetrodotoxin in bacteria. Appl. Environ. Microbiol. 2001, 67, 2393–2394. [Google Scholar] [CrossRef] [PubMed]
- Strand, M.; Hedström, M.; Seth, H.; McEvoy, E.G.; Jacobsson, E.; Göransson, U.; Andersson, H.S.; Sundberg, P. The bacterial (Vibrio alginolyticus) production of tetrodotoxin in the ribbon worm Lineus longissimus—Just a False Positive? Mar. Drugs 2016, 14, 63. [Google Scholar] [CrossRef] [PubMed]
- Noguchi, T.; Jeon, J.K.; Arakawa, O.; Sugita, H.; Deguchi, Y.; Shida, Y.; Hashimoto, K. Occurrence of tetrodotoxin and anhydrotetrodotoxin in Vibrio sp. isolated from the intestines of a xanthid crab, Atergatis floridus. J. Biochem. 1986, 99, 311–314. [Google Scholar] [CrossRef] [PubMed]
- Yasumoto, T.; Yasumura, D.; Yotsu, M.; Michishita, T.; Endo, A.; Kotak, Y. Bacterial production of tetrodotoxin and anhydrotetrodotoxin. Agric. Biol. Chem. 1986, 50, 793–795. [Google Scholar] [CrossRef]
- Narita, H.; Matsubara, S.; Miwa, N.; Akahane, S.; Murakami, M.; Goto, T.; Nara, M.; Noguchi, T.; Saito, T.; Shida, Y.; et al. Vibrio alginolyticus, a TTX-producing bacterium isolated from the starfish Astropecten polyacanthus. Nippon Suisan Gakkaishi 1987, 53, 617–621. [Google Scholar] [CrossRef]
- Noguchi, T.; Hwang, D.F.; Arakawa, O.; Sugita, H.; Deguchi, Y.; Shida, Y.; Hashimoto, K. Vibrio alginolyticus, a tetrodotoxin-producing bacterium, in the intestines of the fish Fugu vermicularis vermicularis. Mar. Biol. 1987, 94, 625–630. [Google Scholar] [CrossRef]
- Simidu, U.; Noguchi, T.; Hwang, D.F.; Shida, Y.; Hashimoto, K. Marine bacteria which produce tetrodotoxin. Appl. Environ. Microbiol. 1987, 53, 1714–1715. [Google Scholar] [PubMed]
- Yotsu, M.; Yamazaki, T.; Meguro, Y.; Endo, A.; Murata, M.; Naoki, H.; Yasumoto, T. Production of tetrodotoxin and its derivatives by Pseudomonas sp. isolated from the skin of a pufferfish. Toxicon 1987, 25, 225–228. [Google Scholar] [CrossRef]
- Kungsuwan, A.; Noguchi, T.; Arakawa, O.; Simidu, U.; Tsukamoto, K.; Shida, Y.; Hashimoto, K. Tetrodotoxin producing bacteria from the horseshoe crab Carcinoscorpius rotundicauda. Nippon Suisan Gakkaishi 1988, 54, 1799–1802. [Google Scholar] [CrossRef]
- Hwang, D.F.; Arakawa, O.; Saito, T.; Noguchi, T.; Simidu, U.; Tsukamoto, K.; Shida, Y.; Hashimoto, K. Tetrodotoxin-producing bacteria from the blue-ringed octopus Octopus maculosus. Mar. Biol. 1989, 100, 327–332. [Google Scholar] [CrossRef]
- Matsui, T.; Taketsugu, S.; Kodama, K.; Shiba-Ishii, A.; Yamamori, K.; Shimizu, C.; Shira-Ishi, A. Production of tetrodotoxin by the intestinal bacteria of a puffer fish Takifugu niphobles. Nippon Suisan Gakkaishi 1989, 55, 2199–2203. [Google Scholar] [CrossRef]
- Thuesen, E.V.; Kogure, K. Bacterial production of tetrodotoxin in four species of Chaetognatha. Biol. Bull. 1989, 176, 191–194. [Google Scholar] [CrossRef]
- Do, H.K.; Kogure, K.; Simidu, U. Identification of deep-sea-sediment bacteria which produce tetrodotoxin. Appl. Environ. Microbiol. 1990, 56, 1162–1163. [Google Scholar] [PubMed]
- Do, H.K.; Kogure, K.; Imada, C.; Noguchi, T.; Ohwada, K.; Simidu, U. Tetrodotoxin production of actinomycetes isolated from marine sediment. J. Appl. Microbiol. 1991, 70, 464–468. [Google Scholar] [CrossRef]
- Do, H.K.; Hamasaki, K.; Ohwada, K.; Simidu, U.; Noguchi, T.; Shida, Y.; Kogure, K. Presence of tetrodotoxin and tetrodotoxin-producing bacteria in freshwater sediments. Appl. Environ. Microbiol. 1993, 59, 3934–3937. [Google Scholar] [PubMed]
- Hwang, D.F.; Cheng, C.A.; Chen, H.C.; Jeng, S.S.; Noguchi, T.; Ohwada, K.; Hashimoto, K. Microflora and tetrodotoxin-producing bacteria in the lined moon shell Natica lineata. Fish. Sci. 1994, 60, 567–571. [Google Scholar]
- Cheng, C.A.; Hwang, D.F.; Tsai, Y.H.; Chen, H.C.; Jeng, S.S.; Noguchi, T.; Ohwada, K.; Hasimoto, K. Microflora and tetrodotoxin-producing bacteria in a gastropod, Niotha clathrata. Food Chem. Toxicol. 1995, 33, 929–934. [Google Scholar] [CrossRef]
- Lee, M.J.; Jeong, D.Y.; Kim, W.S.; Kim, H.D.; Kim, C.H.; Park, W.W.; Park, Y.H.; Kim, K.S.; Kim, H.M.; Kim, D.S. A tetrodotoxin-producing Vibrio strain, LM-1, from the puffer fish Fugu vermicularis radiatus. Appl. Environ. Microbiol. 2000, 66, 1698–1701. [Google Scholar] [CrossRef] [PubMed]
- Ritchie, K.B.; Nagelkerken, I.; James, S.; Smith, G.W. A tetrodotoxin-producing marine pathogen. Nature 2000, 404, 354. [Google Scholar] [CrossRef] [PubMed]
- Carroll, S.; McEvoy, E.G.; Gibson, R. The production of tetrodotoxin-like substances by nemertean worms in conjunction with bacteria. J. Exp. Mar. Biol. Ecol. 2003, 288, 51–63. [Google Scholar] [CrossRef]
- Yu, C.F.; Yu, P.H.; Chan, P.L.; Yan, Q.; Wong, P.K. Two novel species of tetrodotoxin-producing bacteria isolated from toxic marine puffer fishes. Toxicon 2004, 44, 641–647. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Yang, Y.; Xie, L.; Xia, G.; Hu, J.; Wang, S.; Zhang, R. Toxicity and distribution of tetrodotoxin-producing bacteria in puffer fish Fugu rubripes collected from the Bohai Sea of China. Toxicon 2005, 46, 471–476. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Xie, L.; Xia, G.; Zhang, J.; Nie, Y.; Hu, J.; Wang, S.; Zhang, R. A new tetrodotoxin-producing actinomycete, Nocardiopsis dassonvillei, isolated from the ovaries of puffer fish Fugu rubripes. Toxicon 2005, 45, 851–859. [Google Scholar] [CrossRef] [PubMed]
- Maran, B.A.; Iwamoto, E.; Okuda, J.; Matsuda, S.; Taniyama, S.; Shida, Y.; Asakawa, M.; Ohtsuka, S.; Nakai, T.; Boxshall, G.A. Isolation and characterization of bacteria from the copepod Pseudocaligus fugu ectoparasitic on the panther puffer Takifugu pardalis with the emphasis on TTX. Toxicon 2007, 50, 779–790. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.J.; Yu, R.C.; Luo, X.; Zhou, M.J.; Lin, X.T. Toxin-screening and identification of bacteria isolated from highly toxic marine gastropod Nassarius semiplicatus. Toxicon 2008, 52, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Yi, R. Bacillus horikoshii, a tetrodotoxin-producing bacterium isolated from the liver of puffer fish. Ann. Microbiol. 2009, 59, 453–458. [Google Scholar] [CrossRef]
- Campbell, S.; Harada, R.M.; DeFelice, S.V.; Bienfang, P.K.; Li, Q.X. Bacterial production of tetrodotoxin in the pufferfish Arothron hispidus. Nat. Prod. Res. 2009, 23, 1630–1640. [Google Scholar] [CrossRef] [PubMed]
- Bragadeeswaran, S.; Therasa, D.; Prabhu, K.; Kathiresan, K. Biomedical and pharmacological potential of tetrodotoxin-producing bacteria isolated from marine pufferfish Arothron hispidus (Muller, 1841). J. Venom. Anim. Toxins Trop. Dis. 2010, 16, 421–431. [Google Scholar] [CrossRef]
- Wang, J.; Fan, Y. Isolation and characterization of a Bacillus species capable of producing tetrodotoxin from the puffer fish Fugu obscurus. World J. Microbiol. Biotechnol. 2010, 26, 1755–1760. [Google Scholar] [CrossRef]
- Wang, J.; Fan, Y.; Yao, Z. Isolation of a Lysinibacillus fusiformis strain with tetrodotoxin-producing ability from puffer fish Fugu obscurus and the characterization of this strain. Toxicon 2010, 56, 640–643. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Xu, J.; Liang, S.; Ren, D.; Yan, X.; Bao, B. A novel TTX-producing Aeromonas isolated from the ovary of Takifugu obscurus. Toxicon 2010, 56, 324–329. [Google Scholar] [CrossRef] [PubMed]
- Yu, V.C.; Yu, P.H.; Ho, K.C.; Lee, F.W. Isolation and identification of a new tetrodotoxin-producing bacterial species, Raoultella terrigena, from Hong Kong marine puffer fish Takifugu niphobles. Mar. Drugs 2011, 9, 2384–2396. [Google Scholar] [CrossRef] [PubMed]
- Auawithoothij, W.; Noomhorm, A. Shewanella putrefaciens, a major microbial species related to tetrodotoxin (TTX)-accumulation of puffer fish Lagocephalus lunaris. J. Appl. Microbiol. 2012, 113, 459–465. [Google Scholar] [CrossRef] [PubMed]
- Magarlamov, T.Y.; Beleneva, I.A.; Chernyshev, A.V.; Kuhlevsky, A.D. Tetrodotoxin-producing Bacillus sp. from the ribbon worm (Nemertea) Cephalothrix simula (Iwata, 1952). Toxicon 2014, 85, 46–51. [Google Scholar] [CrossRef] [PubMed]
- Tu, N.; Tu, Q.; Tung, H.; Hieu, D.; Romero-Jovel, S. Detection of tetrodotoxin-producing Providencia rettgeri T892 in Lagocephalus pufferfish. World J. Microbiol. Biotechnol. 2014, 30, 1829–1835. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.H.; Nguyen, H.N.; Nghe, D.V.; Nguyen, K.H. Biological activities of tetrodotoxin-producing Enterococcus faecium AD1 isolated from puffer fishes. BioMed Res. Int. 2015, 2015, 973235. [Google Scholar] [CrossRef] [PubMed]
- Wei, F.; Ma, T.; Gong, X.; Zhang, N.; Bao, B. Identification of tetrodotoxin-producing bacteria from goby Yongeichthys criniger. Toxicon 2015, 104, 46–51. [Google Scholar] [CrossRef] [PubMed]
- Turner, A.D.; Powell, A.; Schofield, A.; Lees, D.N.; Baker-Austin, C. Detection of the pufferfish toxin tetrodotoxin in European bivalves, England, 2013 to 2014. Euro Surveill. 2015, 20, 1–33. [Google Scholar] [CrossRef]
- Rodríguez, I.; Alfonso, A.; Alonso, E.; Rubiolo, J.A.; Roel, M.; Vlamis, A.; Katikou, P.; Jackson, S.J.; Menon, M.S.; Dobson, A.; et al. The association of bacterial C9-based TTX-like compounds with Prorocentrum minimum opens new uncertainties about shellfish seafood safety. Sci. Rep. 2017, 7, 40880. [Google Scholar] [CrossRef] [PubMed]
- Simidu, U.; Kita-Tsukamoto, K.; Yasumoto, T.; Yotsu, M. Taxonomy of four marine bacterial strains that produce tetrodotoxin. Int. J. Syst. Bacteriol. 1990, 40, 331–336. [Google Scholar] [CrossRef] [PubMed]
- Ivanova, E.P.; Romanenko, L.A.; Matté, M.H.; Matté, G.R.; Lysenko, A.M.; Simidu, U.; Kita-Tsukamoto, K.; Sawabe, T.; Vysotskii, M.V.; Frolova, G.M.; et al. Retrieval of the species Alteromonas tetraodonis Simidu et al. 1990 as Pseudoalteromonas tetraodonis comb. nov. and emendation of description. Int. J. Syst. Evol. Microbiol. 2001, 51, 1071–1078. [Google Scholar] [CrossRef] [PubMed]
- Simmons, T.L.; Coates, R.C.; Clark, B.R.; Engene, N.; Gonzalez, D.; Esquenazi, E.; Dorrestein, P.C.; Gerwick, W.H. Biosynthetic origin of natural products isolated from marine microorganism–invertebrate assemblages. Proc. Natl. Acad. Sci. USA 2008, 105, 4587–4594. [Google Scholar] [CrossRef] [PubMed]
- Okita, K.; Takatani, T.; Nakayasu, J.; Yamazaki, H.; Sakiyama, K.; Ikeda, K.; Arakawa, O.; Sakakura, Y. Comparison of the localization of tetrodotoxin between wild pufferfish Takifugu rubripes juveniles and hatchery-reared juveniles with tetrodotoxin administration. Toxicon 2013, 71, 128–133. [Google Scholar] [CrossRef] [PubMed]
- Honda, S.; Arakawa, O.; Takatani, T.; Tachibana, K.; Yagi, M.; Tanigawa, A.; Noguchi, T. Toxification of cultured puffer fish Takifugu rubripes by feeding on tetrodotoxin-containing diet. Nippon Suisan Gakkaishi 2005, 71, 815–820. [Google Scholar] [CrossRef]
- Hwang, D.F.; Chueh, C.H.; Jeng, S.S. Tetrodotoxin secretion from the lined moon shell Natica Lineata in response to external stimulation. Toxicon 1990, 28, 1133–1136. [Google Scholar] [CrossRef]
- Hwang, D.F.; Lin, L.C.; Jeng, S.S. Occurrence of tetrodotoxin related toxins in the gastropod mollusk Niotha clathrata from Taiwan. Nippon Suisan Gakkaishi 1992, 55, 63–67. [Google Scholar] [CrossRef]
- Williams, B.L. Behavioral and chemical ecology of marine organisms with respect to tetrodotoxin. Mar. Drugs 2010, 8, 381–398. [Google Scholar] [CrossRef] [PubMed]
- Matsumura, K. Tetrodotoxin as a pheromone. Nature 1995, 378, 563–564. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, P.; Alfonso, A.; Otero, P.; Katikou, P.; Georgantelis, D.; Botana, L.M. Liquid chromatography–mass spectrometry method to detect tetrodotoxin and its analogues in the puffer fish Lagocephalus scelerat (Gmelin, 1789) from European waters. Food Chem. 2012, 132, 1103–1111. [Google Scholar] [CrossRef]
- Katikou, P.; Georgantelis, D.; Sinouris, N.; Petsi, A.; Fotaras, T. First report on toxicity assessment of the Lessepsian migrant pufferfish Lagocephalus sceleratus (Gmelin, 1789) from European waters (Aegean Sea, Greece). Toxicon 2009, 54, 50–55. [Google Scholar] [CrossRef] [PubMed]
- Cassiday, L. First report of TTX in a European trumpet shell. Anal. Chem. 2008, 80, 5675. [Google Scholar] [CrossRef]
- Rodriguez, P.; Alfonso, A.; Vale, C.; Alfonso, C.; Vale, P.; Tellez, A.; Botana, L.M. First toxicity report of tetrodotoxin and 5,6,11-trideoxyTTX in the trumpet shell Charonia lampas lampas in Europe. Anal. Chem. 2008, 80, 5622–5629. [Google Scholar] [CrossRef] [PubMed]
- Silva, M.; Azevedo, J.; Rodriguez, P.; Alfonso, A.; Botana, L.M.; Vasconcelos, V. New gastropod vectors and tetrodotoxin potential expansion in temperate waters of the Atlantic Ocean. Mar. Drugs 2012, 10, 712–726. [Google Scholar] [CrossRef] [PubMed]
- Nzoughet, J.K.; Campbell, K.; Barnes, P.; Cooper, K.M.; Chevallier, O.P.; Elliott, C.T. Comparison of sample preparation methods, validation of an UPLC–MS/MS procedure for the quantification of tetrodotoxin present in marine gastropods and analysis of pufferfish. Food Chem. 2013, 136, 1584–1589. [Google Scholar] [CrossRef] [PubMed]
- Vlamis, A.; Katikou, P.; Rodriguez, I.; Rey, V.; Alfonso, A.; Papazachariou, A.; Zacharaki, T.; Botana, A.M.; Botana, L.M. First detection of tetrodotoxin in Greek shellfish by UPLC-MS/MS potentially linked to the presence of the dinoflagellate Prorocentrum minimum. Toxins 2015, 7, 1779–1807. [Google Scholar] [CrossRef] [PubMed]
- Sugita, H.; Iwata, J.; Miyajima, C.; Kubo, T.; Noguchi, T.; Hashimoto, K.; Deguchi, Y. Changes in microflora of a puffer fish Fugu niphobles, with different water temperatures. Mar. Biol. 1989, 101, 299–304. [Google Scholar] [CrossRef]
- Akopian, A.N.; Souslova, V.; England, S.; Okuse, K.; Ogata, N.; Ure, J.; Smith, A.; Kerr, B.J.; McMahon, S.B.; Boyce, S.; et al. The tetrodotoxin-resistant sodium channel SNS has a specialized function in pain pathways. Nat. Neurosci. 1999, 2, 541–548. [Google Scholar] [CrossRef] [PubMed]
- Wood, J.N.; Boorman, J.P.; Okuse, K.; Baker, M.D. Voltage-gated sodium channels and pain pathways. J. Neurobiol. 2004, 61, 55–71. [Google Scholar] [CrossRef] [PubMed]
- Lago, J.; Rodríguez, L.P.; Blanco, L.; Vieites, J.M.; Cabado, A.G. Tetrodotoxin, an extremely potent marine neurotoxin: Distribution, toxicity, origin and therapeutical uses. Mar. Drugs 2015, 13, 6384–6406. [Google Scholar] [CrossRef] [PubMed]
- Berde, C.B.; Athiraman, U.; Yahalom, B.; Zurakowski, D.; Corfas, G.; Bognet, C. Tetrodotoxin-bupivacaine-epinephrine combinations for prolonged local anesthesia. Mar. Drugs. 2011, 9, 2717–2728. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Shum, F. United States Patent: Method of Extracting Tetrodotoxin. US Patent 6552191, 22 April 2003. [Google Scholar]
- Chau, R.; Kalaitzis, J.A.; Neilan, B.A. On the origins and biosynthesis of tetrodotoxin. Aquat. Toxicol. 2011, 104, 61–72. [Google Scholar] [CrossRef] [PubMed]
- Magarlamov, T.Y.; Melnikova, D.I.; Shokur, O.A.; Gorobets, E.A. Rapid production of tetrodotoxin-like compounds during sporulation in a marine isolate Bacillus sp. 1839. Microbiology 2017, 86, 192–196. [Google Scholar] [CrossRef]
- Liu, J.; Wei, F.; Lu, Y.; Ma, T.; Zhao, J.; Gong, X.; Bao, B. Production level of tetrodotoxin in Aeromonas is associated with the copy number of a plasmid. Toxicon 2015, 101, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Chung, SH. Mechanism of tetrodotoxin block and resistance in sodium channels. Biochem. Biophys. Res. Commun. 2014, 446, 370–374. [Google Scholar] [CrossRef] [PubMed]
- Payandeh, J.; Minor, D.L., Jr. Bacterial voltage-gated Sodium Channels (BacNaVs) from the soil, sea, and salt lakes enlighten molecular mechanisms of electrical signaling and pharmacology in the brain and heart. J. Mol. Biol. 2015, 427, 3–30. [Google Scholar] [CrossRef]
- Ren, D.; Navarro, B.; Xu, H.; Yue, L.; Shi, Q.; Clapham, D.E. A prokaryotic voltage-gated sodium channel. Science 2001, 294, 2372–2375. [Google Scholar] [CrossRef] [PubMed]
- Kodama, M.; Sato, S.; Ogata, T.; Suzuki, Y.; Kaneko, T.; Aida, K. Tetrodotoxin secreting glands in the skin of puffer fishes. Toxicon 1986, 24, 819–829. [Google Scholar] [CrossRef]
- Williams, B.L.; Caldwell, R.L. Intra-organismal distribution of tetrodotoxin in two species of blue-ringed octopuses (Hapalochlaena fasciata and H. lunulata). Toxicon 2009, 54, 345–353. [Google Scholar] [CrossRef] [PubMed]
- Salvitti, L.R.; Wood, S.A.; McNabb, P.; Cary, S.C. No evidence for a culturable bacterial tetrodotoxin producer in Pleurobranchaea maculata (Gastropoda: Pleurobranchidae) and Stylochoplana sp. (Platyhelminthes: Polycladida). Toxins 2015, 7, 255–273. [Google Scholar] [CrossRef] [PubMed]
- Tanu, M.B.; Mahmud, Y.; Takatani, T.; Kawatsu, K.; Hamano, Y.; Arakawa, O.; Noguchi, T. Localization of tetrodotoxin in the skin of a brackishwater puffer Tetraodon steindachneri on the basis of immunohistological study. Toxicon 2002, 40, 103–106. [Google Scholar] [CrossRef]
- Tsuruda, K.; Arakawa, O.; Kawatsu, K.; Hamano, Y.; Takatani, T.; Noguchi, T. Secretory glands of tetrodotoxin in the skin of the Japanese newt Cynops pyrrhogaster. Toxicon 2002, 40, 131–136. [Google Scholar] [CrossRef]
- Tanu, M.B.; Mahmud, Y.; Arakawa, O.; Takatani, T.; Kajihara, H.; Kawatsu, K.; Hamano, Y.; Asakawa, M.; Miyazawa, K.; Noguchi, T. Immunoenzymatic visualization of tetrodotoxin (TTX) in Cephalothrix species (Nemertea: Anopla: Palaeonemertea: Cephalotrichidae) and Planocera reticulata (Platyhelminthes: Turbellaria: Polycladida: Planoceridae). Toxicon 2004, 44, 515–520. [Google Scholar] [CrossRef] [PubMed]
- Magarlamov, T.Y.; Shokur, O.A.; Chernyshev, A.V. Distribution of tetrodotoxin in the ribbon worm Lineus alborostratus (Takakura, 1898) (nemertea): Immunoelectron and immunofluorescence studies. Toxicon 2016, 112, 29–34. [Google Scholar] [CrossRef]
- Matsumoto, T.; Nagashima, Y.; Kusuhara, H.; Ishizaki, S.; Shimakura, K.; Shiomi, K. Pharmacokinetics of tetrodotoxin in puffer fish Takifugu rubripes by a single administration technique. Toxicon 2008, 51, 1051–1059. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, T.; Nagashima, Y.; Kusuhara, H.; Ishizaki, S.; Shimakura, K.; Shiomi, K. Evaluation of hepatic uptake clearance of tetrodotoxin in the puffer fish Takifugu rubripes. Toxicon 2008, 52, 369–374. [Google Scholar] [CrossRef] [PubMed]
- Tatsuno, R.; Shikina, M.; Shirai, Y.; Wang, J.; Soyano, K.; Nishihara, G.N.; Takatani, T.; Arakawa, O. Change in the transfer profile of orally administered tetrodotoxin to non-toxic cultured pufferfish Takifugu rubripes depending of its development stage. Toxicon 2013, 65, 76–80. [Google Scholar] [CrossRef] [PubMed]
- Noguchi, T.; Arakawa, O.; Takatani, T. TTX accumulation in pufferfish. Comp. Biochem. Physiol. Part D Genom. Proteom. 2006, 1, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Kogure, K.; Do, H.K.; Kim, D.S.; Shirayama, Y. High concentrations of neurotoxin in free-living marine nematodes. Mar. Ecol. Prog. Ser. 1996, 136, 147–151. [Google Scholar] [CrossRef]
- Kajihara, H.; Sun, S.C.; Chernyshev, A.V.; Chen, H.X.; Ito, K.; Asakawa, M.; Maslakova, S.A.; Norenburg, J.L.; Strand, M.; Sundberg, P.; et al. Taxonomic identity of a tetrodotoxin-accumulating ribbon-worm Cephalothrix simula (Nemertea: Palaeonemertea): A species artificially introduced from the Pacific to Europe. Zool. Sci. 2013, 30, 985–997. [Google Scholar] [CrossRef] [PubMed]
- Gallach, S.; Birkbeck, T.H. Effect of Phosphate Concentration on Production of Tetrodotoxin by Alteromonas tetraodonis. Appl. Environ. Microbiol. 1993, 59, 3981–3983. [Google Scholar]
- Bevins, C.L.; Salzman, N.H. The potter’s wheel: The host’s role in sculpting its microbiota. Cell. Mol. Life Sci. 2011, 68, 3675–3685. [Google Scholar] [CrossRef] [PubMed]
- Duarte, S.; Silva, F.C.; Zauli, D.A.; Nicoli, J.R.; Araújo, F.G. Gram-negative intestinal indigenous microbiota from two Siluriform fishes in a tropical reservoir. Braz. J. Microbiol. 2014, 45, 1283–1292. [Google Scholar] [CrossRef] [PubMed]
- Ringo, E.; Zhou, Z.; Vecino, J.L.G.; Wadsworth, S.; Romero, J.; Krogdahl, Å.; Olsen, R.E.; Dimitroglou, A.; Foey, A.; Davies, S.; et al. Effect of dietary components on the gut microbiota of aquatic animals. A never-ending story? Aquac. Nutr. 2016, 22, 219–282. [Google Scholar] [CrossRef]
- Lehman, E.M.; Brodie, E.D., Jr.; Brodie, E.D., III. No evidence for an endosymbiotic bacterial origin of tetrodotoxin in the newt Taricha granulosa. Toxicon 2004, 44, 243–249. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
| Group of Methods | Method | Abbreviation | Sensitivity µg/mL | Limitations | |
|---|---|---|---|---|---|
| Bioassay | Mouse Bioassay | MBA | 0.2 |
| |
| Tissue Culture Bioassay | TCBA | 0.5 |
| ||
| Immunological methods | Enzyme-Linked Immunosorbent Assay | ELISA | <0.001 |
| |
| Immunohistochemistry | IHC | ||||
| Physicochemical methods | Thin-Layer Chromatography | TLC | 2 |
| |
| Liquid Chromatography | with Fluorescence Detection | LC–FLD | 0.001–0.002 |
| |
| with Mass Spectrometry | LC–MS |
| |||
| with Tandem Mass Spectrometry | LC–MS–MS | - | |||
| Gas Chromatography with Mass Spectrometry | GC–MS | 0.001–0.002 |
| ||
| Ultraviolet–Visible Spectroscopy | UV–Vis spectroscopy | 0.1 |
| ||
| Electrophoresis | - | 2 |
| ||
| TTX-Producing Microorganism | Number of Strains | Source of Isolation * | Place | TTX Detection Method | Toxins Detected | TTX Concentration (µg/mL) | References |
|---|---|---|---|---|---|---|---|
| Vibrio sp. | 1 | xanthid crab Atergatis floridus | Japan | HPLC–FLD GC–MS | TTX anhydro-TTX | NM | [26] |
| Pseudomonas sp. | 1 | red alga Jania sp. | Japan | HPLC–FLD HPLC–FAB–MS MBA | TTX anhydro-TTX | <10 | [27] |
| Vibrio alginolyticus | 2 | starfish Astropecten polycanthus | Japan | HPLC–MS UV–Vis GC–MS | TTX anhydro-TTX 4-epi-TTX | 0.08 | [28] |
| Vibrio alginolyticus | 2 | puffer fish Fugu vermicularis vermicularis | Japan | HPLC–FLD GC–MS | TTX anhydro-TTX | 0.0012 | [29] |
| Vibrio alginolyticus ATCC 17749 Vibrio alginolyticus NCMB 1903 | - | American Type Culture Collection (Rockville, MD, USA) National Collection of Marine Bacteria (Aberdeen, Scotland) | - | HPLC–FLD GC–MS | anhydro-TTX | NM | [30] |
| Vibrio parahaemolyticus NCMB 1902 Vibrio parahaemolyticus ATCC 17802 | - | ||||||
| Vibrio anguillarum NCMB 1291 | - | ||||||
| Photobacterium phosphoreum NCMB 844 | - | ||||||
| Aeromonas salmonicida ATCC 14174 | - | ||||||
| Plesiomonas shigelloides ATCC 14029 | - | ||||||
| Pseudomonas sp. | 1 | puffer fish Fugu poecilonotus | Japan | HPLC–FLD GC–MS | TTX anhydro-TTX 4-epi-TTX | NM | [31] |
| Vibrio alginolyticus | 1 | horseshoe crab Carcinoscorpius rotundicauda | Thailand | HPLC–MS UV–Vis GC–MS | TTX anhydro-TTX | NM | [32] |
| Alteromonas sp. | 2 | octopus Octopus maculosus | Philippines | MBA HPLC–FLD GC–MS | TTX anhydro-TTX | NM | [33] |
| Bacillus sp. | 2 | 0.0015 | |||||
| Pseudomonas sp. | 1 | 0.0025 | |||||
| Vibrio sp. | 1 | NM | |||||
| Shewanella putrefaciens | 1 | puffer fish Takifugu niphobles | Japan | MBA HPLC–FLD GC–MS | TTX anhydro-TTX | 0.012 | [34] |
| Vibrio alginolyticus | 1 | planktonic chaetognaths: Flussisagitta lyra, Parasagitta elegans, Zonosagitta nagae, Eukrohnia hamata | Japan | TCBA HPLC–FLD | TTX | 0.28–0.79 | [35] |
| Vibrio sp. | 1 | marine sediments | Japan | TCBA HPLC–FLD | TTX anhydro-TTX | NM | [36] |
| Bacillus sp. | 5 | ||||||
| Alteromonas sp. | 5 | ||||||
| Aeromonas sp. | 1 | ||||||
| Micrococcus sp. | 4 | ||||||
| Acinetobacter sp. | 3 | ||||||
| Moraxella sp. | 2 | ||||||
| Streptomyces sp. | 7 | marine sediments | Japan | TCBA HPLC–FLD GC–MS | TTX | NM | [37] |
| Bacillus sp. | 7 | freshwater sediment | Japan | TCBA HPLC–FLD GC–MS | TTX anhydro-TTX 4-epi-TTX | NM | [38] |
| Micrococcus sp. | 7 | ||||||
| Alcaligens sp. | 1 | ||||||
| Caulobacter sp. | 1 | ||||||
| Flavobacterium sp. | 1 | ||||||
| Vibrio alginolyticus | 5 | lined moon shell Natica lineata | Taiwan | HPLC–MS UV–Vis GC–MS | TTX anhydro-TTX 4-epi-TTX | NM | [39] |
| Vibrio parahaemolyticus | 1 | ||||||
| Aeromonas sp. | 2 | ||||||
| Pseudomonas sp. | 2 | ||||||
| Vibrio alginolyticus | 3 | gastropod Niotha clathrata | Taiwan | HPLC–MS UV–Vis GC–MS | TTX anhydro-TTX | NM | [40] |
| Vibrio parahaemolyticus | 1 | ||||||
| Pseudomonas sp. | 4 | ||||||
| Aeromonas sp. | 1 | ||||||
| Plesiomonas sp. | 1 | ||||||
| Vibrio sp. | 1 | puffer fish Fugu vermicularis radiates | Korea | MBA HPLC–FLD GC–MS TLC Electrophoresis | TTX anhydro-TTX TDA | NM | [41] |
| Pseudoalteromonas sp. | 1 | sea urchin Meoma ventricosa | Caribbean | IHC | TTX | NM | [42] |
| Vibrio sp. | - | ribbon worms: Cephalothrix rufifrons, Lineus longissimus, Lineus ruber, Lineus viridis, Ramphogordius sanguineus, Riseriellus occultus, Amphiporus lactifloreus | England | UV–Vis HPLC–FLD | TTX 4-epi-TTX TDA | NM | [43] |
| Microbacterium arabinogalactanolyticum | 1 | puffer fish Chelonodon patoca, Takifugu alboplumbeus, Takifugu niphobles | Hong Kong | MBA TLC LC–ESI–MS | TTX anhydro-TTX | 0.042 | [44] |
| Serratia marcescens | 1 | 0.04 | |||||
| Vibrio alginolyticus | 1 | 0.03 | |||||
| Bacillus sp. | 3 | puffer fish Fugu rubripes | China | MBA TCBA TLC HPLC–FLD LC–ESI–MS | TTX | 0.32 | [45] |
| Actinomyces sp. | 1 | ||||||
| Nocardiopsis dassonvillei | 1 | puffer fish Fugu rubripes | China | MBA TCBA HPLC–FLD TLC LC–ESI–MS UV–Vis | TTX anhydro-TTX | 0.1 | [46] |
| Roseobacter sp | 6 | Copepod Pseudocaligus fugu | Japan | MBA GC–MS HPLC–MS | TTX anhydro-TTX | NM | [47] |
| Vibrio sp. | 13 | gastropod Nassarius semiplicatus | China | ELISA | TTX | 0.184 | [48] |
| Shewanella sp. | 3 | ||||||
| Marinomonas sp. | 1 | ||||||
| Tenacibaculum sp. | 1 | ||||||
| Aeromonas sp. | 1 | ||||||
| Bacillus horikoshii | 1 | puffer fish | Taiwan | MBA HPLC–MS HPLC–FLD HPLC–MS–MS | TTX | NM | [49] |
| Vibrio harveyi | 1 | puffer fish Arothron hispidus | Hawaii | HPLC–MS TCBA | TTX anhydro-TTX | 0.5–15.7 | [50] |
| Kytococcus sedentarius | 1 | puffer fish Arothron hispidus | India | MBA | TTX | NM | [51] |
| Cellulomonas fimi | 1 | ||||||
| Bacillus lentimorbus | 1 | ||||||
| Bacillus sp. | 1 | puffer fish Fugu obscurus | China | MBA HPLC–FLD LC–ESI–MS | TTX 4-epi-TTX anhydro-TTX | NM | [52] |
| Lysinibacillus fusiformis | 1 | puffer fish Fugu obscurus | China | MBA LC–ESI–MS | TTX anhydro-TTX | 0.024 | [53] |
| Aeromonas sp. | 1 | puffer fish Takifugu obscurus | China | MBA ELISA HPLC–MS | TTX | 0.002 | [54] |
| Raoultella terrigena | 1 | puffer fish Takifugu niphobles | Hong Kong | MBA ELISA LC–MALDI–TOF MS | TTX | 0.008 | [55] |
| Shewanella putrefaciens | 1 | puffer fish Lagocephalus lunaris | Thailand | HPLC–MS–MS | TTX | 0.195–0.366 | [56] |
| Bacillus sp. | 1 | ribbon worm Cephalothrix simula | Russia | IHC | TTX | NM | [57] |
| Providencia rettgeri | 1 | puffer fish Lagocephalus sp. | Vietnam | MBA HPLC–FLD TLC | TTX | 0.015–0.021 | [58] |
| Enterococcus faecium | 1 | puffer fish | Vietnam | MBA HPLC–FLD TLC | TTX | 0.015–0.03 | [59] |
| Enterobacter cloaca | 1 | gobyfish Yongeichthys criniger | China | MBA ELISA HPLC–MS–MS | TTX | 0.96 | [60] |
| Rahnella aquatilis | 1 | 0.6 | |||||
| Vibrio cholerae Vibrio parahaemolyticus | 1 9 | mussel Mytilus edulis oyster Crassostrea gigas | England | UPLC–ESI–MS–MS | TTX | 0.000042–0.000718 | [61] |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Magarlamov, T.Y.; Melnikova, D.I.; Chernyshev, A.V. Tetrodotoxin-Producing Bacteria: Detection, Distribution and Migration of the Toxin in Aquatic Systems. Toxins 2017, 9, 166. https://doi.org/10.3390/toxins9050166
Magarlamov TY, Melnikova DI, Chernyshev AV. Tetrodotoxin-Producing Bacteria: Detection, Distribution and Migration of the Toxin in Aquatic Systems. Toxins. 2017; 9(5):166. https://doi.org/10.3390/toxins9050166
Chicago/Turabian StyleMagarlamov, Timur Yu., Daria I. Melnikova, and Alexey V. Chernyshev. 2017. "Tetrodotoxin-Producing Bacteria: Detection, Distribution and Migration of the Toxin in Aquatic Systems" Toxins 9, no. 5: 166. https://doi.org/10.3390/toxins9050166
APA StyleMagarlamov, T. Y., Melnikova, D. I., & Chernyshev, A. V. (2017). Tetrodotoxin-Producing Bacteria: Detection, Distribution and Migration of the Toxin in Aquatic Systems. Toxins, 9(5), 166. https://doi.org/10.3390/toxins9050166