1. Historical Perspective of Botulinum Neurotoxin Serotypes
Botulinum neurotoxins (BoNTs) are the most potent naturally-occurring substances, with as little as 50 ng of neurotoxin sufficient to cause human botulism. This minimal lethal dose is estimated from data on the amount of neurotoxin consumed in cases of foodborne botulism and from animal experiments [
1,
2,
3]. Botulinum neurotoxins are 150-kDa proteins that are comprised of a heavy chain (HC-100 kDa) and a light chain (LC-50 kDa). The heavy chains have two functional domains, with the C-terminal domain (H
C) involved in neurotoxin binding to specific receptors in peripheral nerve terminals (
Table 1) and the N-terminal domain (H
N) involved in translocation of the light chain into the nerve cell cytoplasm [
4,
5,
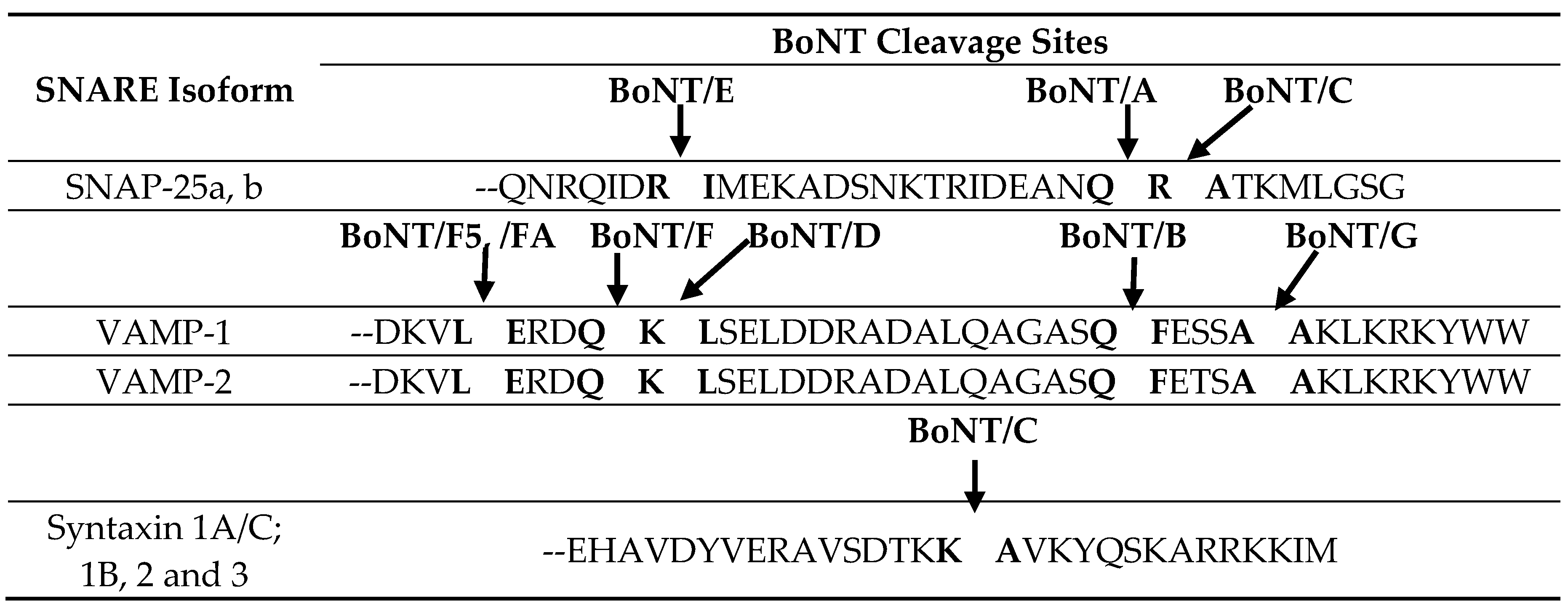
6]. The light chains are zinc metalloproteases that block the release of the neurotransmitter, acetylcholine, in cholinergic nerves by specific cleavage of SNARE (soluble
N-ethylmaleimide-sensitive factor (NSF) attachment receptor) proteins (
Table 2), leading to flaccid paralysis and botulism [
7,
8,
9].
Serological methods were first used to distinguish botulinum neurotoxins more than a century ago. Leuchs [
24] showed that botulinum neurotoxins formed by strains of
Clostridium botulinum isolated following outbreaks of foodborne botulism in Ellezelles (Belgium) and Darmstadt (Germany) were antigenically distinct, with antitoxin raised against one neurotoxin not cross-neutralizing neurotoxin formed by the other strain. Using a similar approach, Burke [
25] also recognized two antigenically-distinct botulinum neurotoxins and designated these as serotypes A and B. Strains that formed type A neurotoxin were reported to dominate in the western USA, and strains that formed type B neurotoxin dominated in the eastern USA [
26,
27]. These pioneering studies established the use of serological methods based on type-specific antitoxins to define and distinguish botulinum neurotoxin serotypes using small animal models. In the decades since Leuchs’ and Burke’s work, the application of the neurotoxin neutralization assay using serotype-specific antisera led to the recognition of seven confirmed botulinum neurotoxin serotypes (types A–G). A potential eighth type (“type H”) was described in 2013. Recent reports have variously described this novel neurotoxin as BoNT/H, BoNT/FA or BoNT/HA [
10,
28,
29,
30,
31,
32,
33].
Historically, the use of serological methods to identify and characterize botulinum neurotoxins has not been without problems. In 1924 (only a few years after the work of Leuchs and Burke), problems were encountered when serotyping neurotoxin formed by newly-identified BoNT/C strains that were tested using specific antisera. It was found that antisera produced from type C strains that were isolated from fly larvae and chickens were able to neutralize type C neurotoxin from several strains isolated in the USA, as well as the “Seddon” type C strain that was isolated from cattle with botulism in Australia, but that antisera produced from the “Seddon” strain neutralized only its homologous toxin [
34]. It is now known that these “type C” toxins are BoNT/CD chimeras composed of the 2/3 type C and 1/3 type D sequence, whereas true type C1 toxins are produced by Seddon-like strains [
35].
Similar issues arose in the serotyping of toxins from multiple type A and type F strains in Argentina [
36]. Significant differences in the efficiency of neutralization were noted, particularly among the BoNT/F toxins. These observations led the researchers to conclude, “there is a general tendency to accept the antigenic homogeneity of the botulinum toxins within each type, and from the year 1924 in which the serological relations among the type C strains were described, later denominated type Cα and Cβ (currently CD and C1), up to this date it was never known with precision what to do with these strains, from the point of view of their classification. But the biologic reality is that there are serologic variations of differing magnitude in strains within each type, evidenced by significant differences in antitoxin consumption in cross neutralization, being the most obvious cases those observed in the strains of type C and F” [
37].
These observations pointed to a level of intratypic serological diversity that underlies the “serotype” designations. It was recognized that these differences may have an impact on the effectiveness of botulism treatment, as antitoxins have been raised against a single toxin subtype per serotype. For example, all currently-produced commercial botulinum antitoxins were produced following vaccination with BoNT/A1, BoNT/B1 and either BoNT/E1 or BoNT/E3 toxoids. The few research studies that have been published evaluating the effectiveness of such antisera have shown differential protection against the spectrum of toxins within a single serotype [
38,
39]. In addition, the impact of intratypic serological diversity on the effectiveness of current antitoxin treatments remains largely unknown.
2. Historical Perspective of Botulinum Neurotoxin Subtypes
Early work had suggested that each strain of
C. botulinum formed a single type of botulinum neurotoxin. However, using mouse tests and specific antisera, Giménez and Ciccarelli described a strain that formed two distinct types of neurotoxin, with a major amount of type A toxin activity and a minor amount of type F toxin activity [
40]. This strain was designated type A, subtype Af [
40]. A number of strains are now described that form more than one type of botulinum neurotoxin [
37,
41,
42]. For more than a decade now, however, the term subtype has been used in a different way, that is to describe intratypic neurotoxin variation based on the amino acid sequence of the neurotoxin (derived following sequencing of the neurotoxin gene). A numerical notation has also been introduced, so that the subtypes are designated BoNT/A1, BoNT/A2, BoNT/A3, etc. [
43,
44,
45,
46,
47,
48,
49] (
Table 3).
The development of techniques to enable the sequencing of individual genes has substantially increased our understanding of botulinum neurotoxin diversity. Within a three-year time span (1990–1993), sequences representing one member from each of the seven neurotoxin serotypes became available [
50,
51,
52,
53,
54,
55,
56,
57]. Five years after initial sequences for each serotype were made public, sequences for eight alternative neurotoxin subtypes had been published [
35,
58,
59,
60,
61,
62,
63]. Five of these subtypes were known to differ in some way from the “reference” toxins for each serotype prior to sequencing. This included a strain that produced a type A1 toxin and contained a nonfunctional BoNT/B gene (an A1(B) strain) [
64].
The first
C. botulinum whole genome sequence was published in 2007 [
47], and many full genomes in addition to individual neurotoxin-encoding genes have now been sequenced. Sequencing has confirmed the distinctiveness of the seven botulinum neurotoxin serotypes (types A–G), with amino acid differences between the seven neurotoxin serotypes ranging from 37.2%–69.6% [
65] (
Table 4). Furthermore, studies of the functionality of the botulinum neurotoxins also support the classification of seven serotypes. Botulinum neurotoxin light chains possess endopeptidase activity and selectively cleave proteins of the neurotransmitter vesicle docking/fusion complex, preventing the formation of a stable complex [
8,
9]. BoNT/A, C and E cleave SNAP-25 at distinct sites; BoNT/B, D, F and G cleave VAMP-1/2/3 at distinct sites; BoNT/C can also cleave syntaxin 1A/C (
Table 2). Each neurotoxin subtype within a serotype cleaves its target substrate at the same single conserved peptide bond, except for BoNT/F5 [
23] (
Table 2).
The technology resulting in the production of monoclonal antibodies provided a new way of evaluating these toxins serologically. Monoclonal antibodies developed against BoNT/A [
66], BoNT/B [
67], BoNT/C1 [
68] and BoNT/E [
69] were used to develop new toxin detection assays and also to discover new aspects of toxin structure and activity. However, these antibodies have been of limited use as predictors of subtype-level differences, since monoclonal antibody epitopes are, at most, limited to 5–7 continuous or discontinuous amino acids. Many toxin subtypes are very closely related, so that most monoclonal antibodies will bind multiple toxin subtypes, severely limiting their discriminating power and making them unsuitable for toxin subtype determinations.
Initial studies using monoclonal antibodies, however, often reported that the antibodies would neutralize some, but not all, subtypes of a specific serotype, suggesting variability within each neurotoxin serotype [
45,
69,
70]. The extent of this variability is now being revealed through sequencing of whole genomes and individual neurotoxin-encoding genes. A landmark article on sequence variation among botulinum neurotoxin serotypes published in 2005 described within-serotype variations among neurotoxin sequences as being of two types: those that were virtually identical and those that were more variable and differed by at least 2.6% in amino acid sequence. This observation was based on a study of 49 neurotoxin sequences (each serotype, with the exception of type G, was represented by 4–17 sequences) [
45]. Subsequent studies sorted these and additional toxin gene sequences into differing phylogenetic clades, which were then identified as new subtypes [
71,
72]. The identification of novel subtypes has thereby been based primarily on the sequence of the botulinum neurotoxin gene and/or derived amino acid sequence. There are more than forty neurotoxin subtypes presently described in the literature (
Table 3;
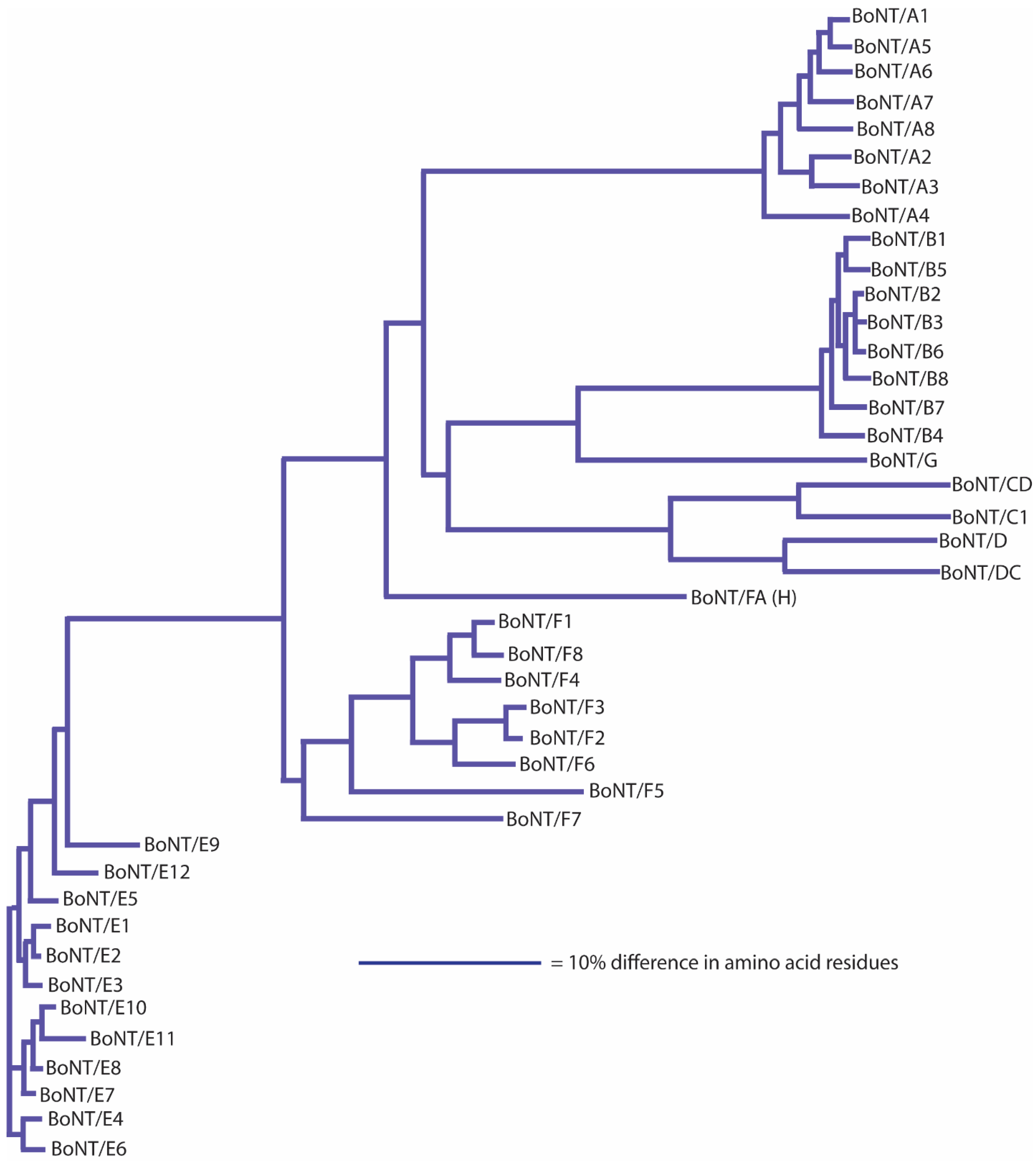
Figure 1).
It has been suggested that a
C. botulinum neurotoxin could be defined as a distinct subtype if it encoded a protein sequence that differed from the prototype neurotoxin by at least 2.6% [
46,
72,
73,
74,
75,
76,
77,
78,
79,
80,
81,
82,
83]. A comparison of neurotoxin sequences from 127 BoNT/A-, 91 BoNT/B-, 23 BoNT/C- and BoNT/D-, 235 BoNT/E- and 50 BoNT/F-producing strains obtained from various sources has been conducted; the results are reported in
Table 3,
Table 4,
Table 5,
Table 6,
Table 7,
Table 8 and
Table 9. This includes published and unpublished neurotoxin sequences, many of which were identical. While no particular efforts to publicly post the redundant sequences were made, examples of each subtype, including strain name, source and GenBank accession number, are listed in
Table 3.
Figure 1,
Figure 2,
Figure 3,
Figure 4,
Figure 5 and
Figure 6 illustrate the range of neurotoxin sequence diversity among these strains. While most presently-described neurotoxin subtypes differ from each other by more than 2.6% at the amino acid level, some BoNT/B subtypes and BoNT/E subtypes do not meet this criterion (
Table 6 and
Table 8). Additionally, it is more common for strains of
C. botulinum Group III to form a chimeric or hybrid protein that combines domains of BoNT/C and BoNT/D neurotoxin, rather than a distinct BoNT/C or BoNT/D neurotoxin (
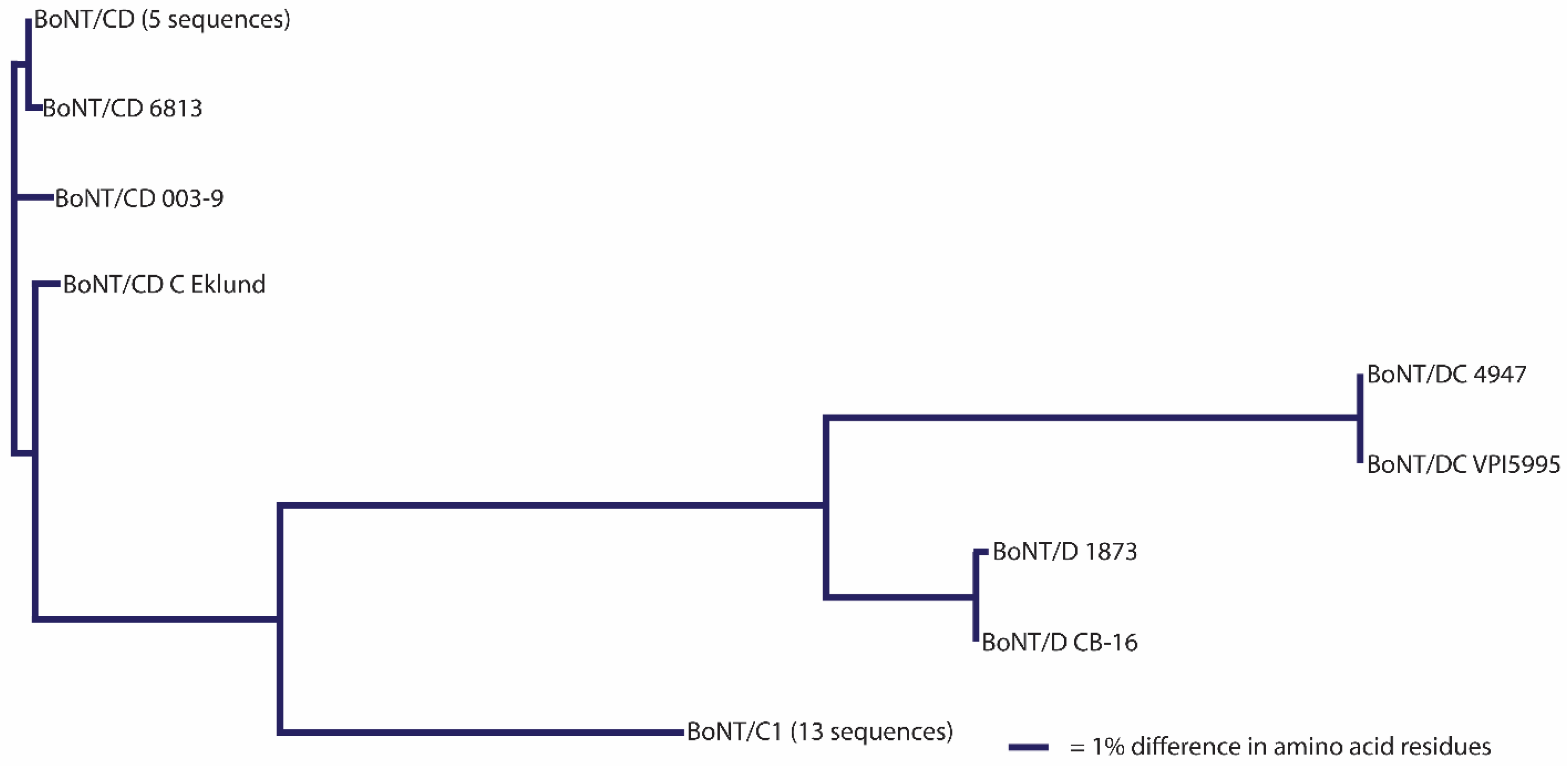
Table 7;
Figure 4).
Discussions as to the relationship between sequence differences and potential immunological or functional differences have led to studies comparing toxin characteristics versus sequence. The majority of these studies involved differential binding to antibodies. As expected, binding differences were more noticeable when monoclonal antibodies were used, but quantitative differences were also noted with assays involving polyclonal antibodies [
38,
45]. However, as noted above, these immunological differences cannot reliably identify distinctive toxin subtypes.
Biological and functional activities are presumed to be largely conserved within individual neurotoxin serotypes. Currently, the only naturally-occurring amino acid sequence changes that have led to functional differences were reported with subtype BoNT/F5, whose enzymatic domain differs from all other BoNT/F enzymatic domains by greater than 50% in amino acid sequence (
Table 2), leading to differences in the enzymatic target site [
23]. All other subtypes within a serotype utilize the same enzymatic cleavage target substrates and sites.
Even though studies of toxin:receptor interactions of subtypes are limited, current information indicates that all toxin subtypes within a serotype also interact with the same receptor targets (
Table 1), with one exception. BoNT/DC interacts with synaptotagmin 1 and 2; the putative receptor for BoNT/D is
N-glycosylated SV2A and B; and BoNT/C appears to interact solely with gangliosides [
15,
16,
17]. This difference may not be too surprising, as BoNT/DC is a hybrid toxin with a BoNT/C-like receptor-binding domain that differs in amino acid sequence from BoNT/D by 60.2% and from BoNT/C by 22.2%. These differences have directed different receptor interactions for each of the toxins. It should also be noted that subtype quantitative binding differences to receptors were seen with two BoNT/B subtypes. BoNT/B1 and BoNT/B2 both interact with synaptotagmin, but BoNT/B1 binds both synaptotagmin 1 and 2, with binding affinities of 3.4 and 0.52 nM, respectively, while BoNT/B2 binds only synaptotagmin 2, with an intermediate binding affinity of 2.4 nM [
44]. There is a significant amino acid sequence difference (8.1%) in the receptor binding domains of BoNT/B1 and BoNT/B2. BoNT/A8 has also been reported to have a reduced affinity to ganglioside receptors compared to BoNT/A1 [
81]. Additional differences in catalytic activity have also been described among BoNT/A subtypes [
84,
85,
86,
87,
88].
It is presently not feasible to use biological, structural, immunological or functional characteristics to subtype botulinum neurotoxins, as knowledge is limited. However, a sequencing-based approach can be used to rapidly categorize botulinum neurotoxin subtypes, to avoid confusion in the literature and to facilitate future research endeavors. One important benefit of this approach is that it allows for comparison of neurotoxins formed by strains located in different laboratories.
When analyzing presently-published toxin subtypes, in which subtype categorization has been based on amino acid sequence differences, it is important to recognize that each serotype shows a unique pattern of between-subtype and within-subtype differences (
Table 5,
Table 6,
Table 7,
Table 8 and
Table 9). In the case of BoNT/A, inter-subtype differences range from 2.9%–15.6% (
Table 5;
Figure 2). Intra-subtype differences are much smaller (≤0.8%, with one exception), thereby providing a sufficient margin of discrimination in sequences that are within subtypes versus those between subtypes. The exception is strain CDC 2171, which differs from other BoNT/A2 by ~2.5%. Due to an apparent recombination event, this toxin shares 100% identity with BoNT/A2 Kyoto-F for approximately 2/3 of the sequence, but differs by 5.6% in the terminal H
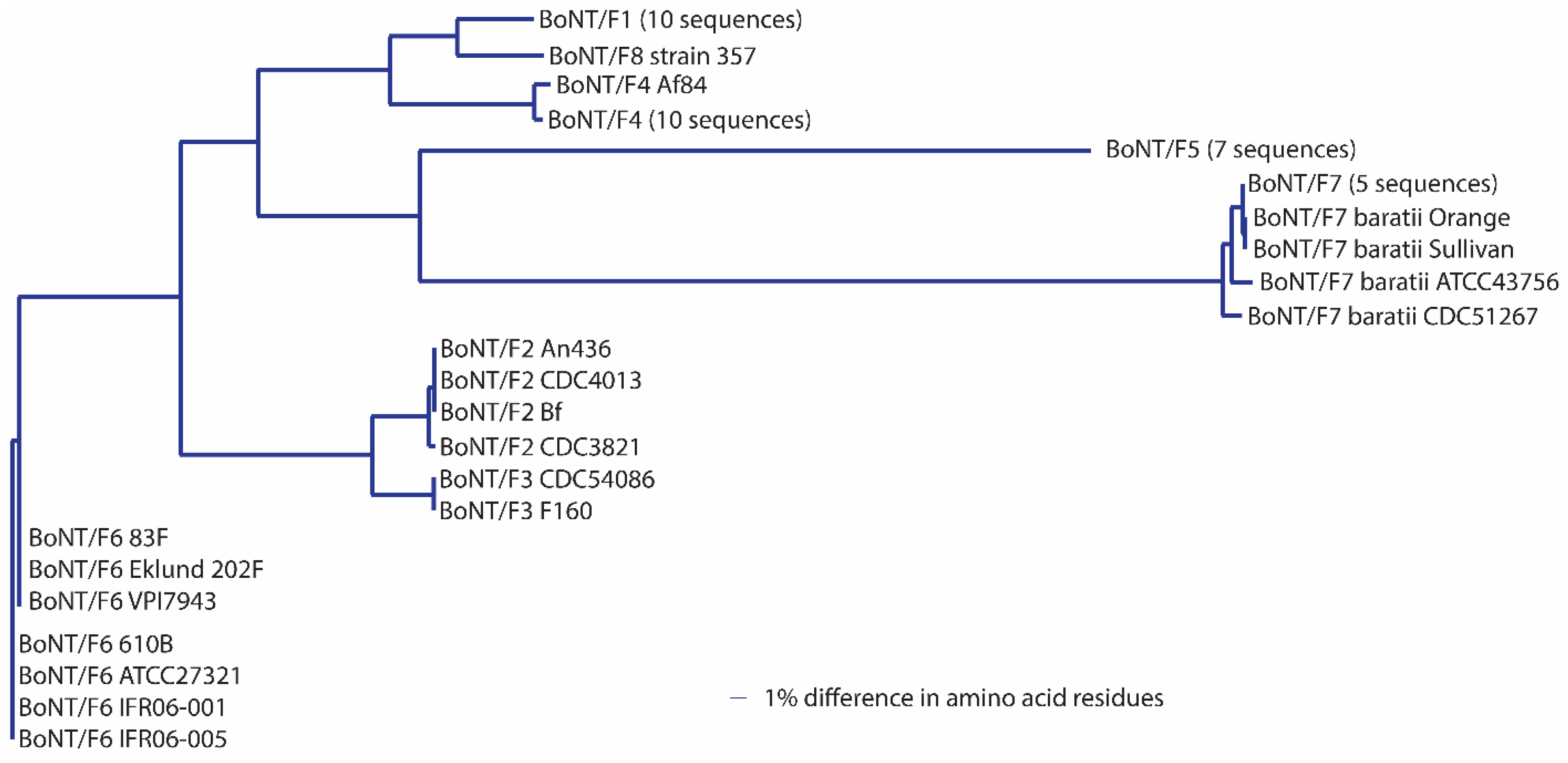
C region of the molecule. The margin of discrimination among BoNT/A subtypes is similar to the scenario with most BoNT/F subtypes, where inter-subtype differences range from 3.0%–36.2%. Intra-subtype differences are ≤0.6% with the exception of BoNT/F7, which shows an intra-subtype difference of 1.7% among the eight sequences that were analyzed (
Table 9). Removal of a single BoNT/F7 sequence, from strain ATCC 43756, results in a reduction in variability within this subtype from 1.7% to 0.8%.
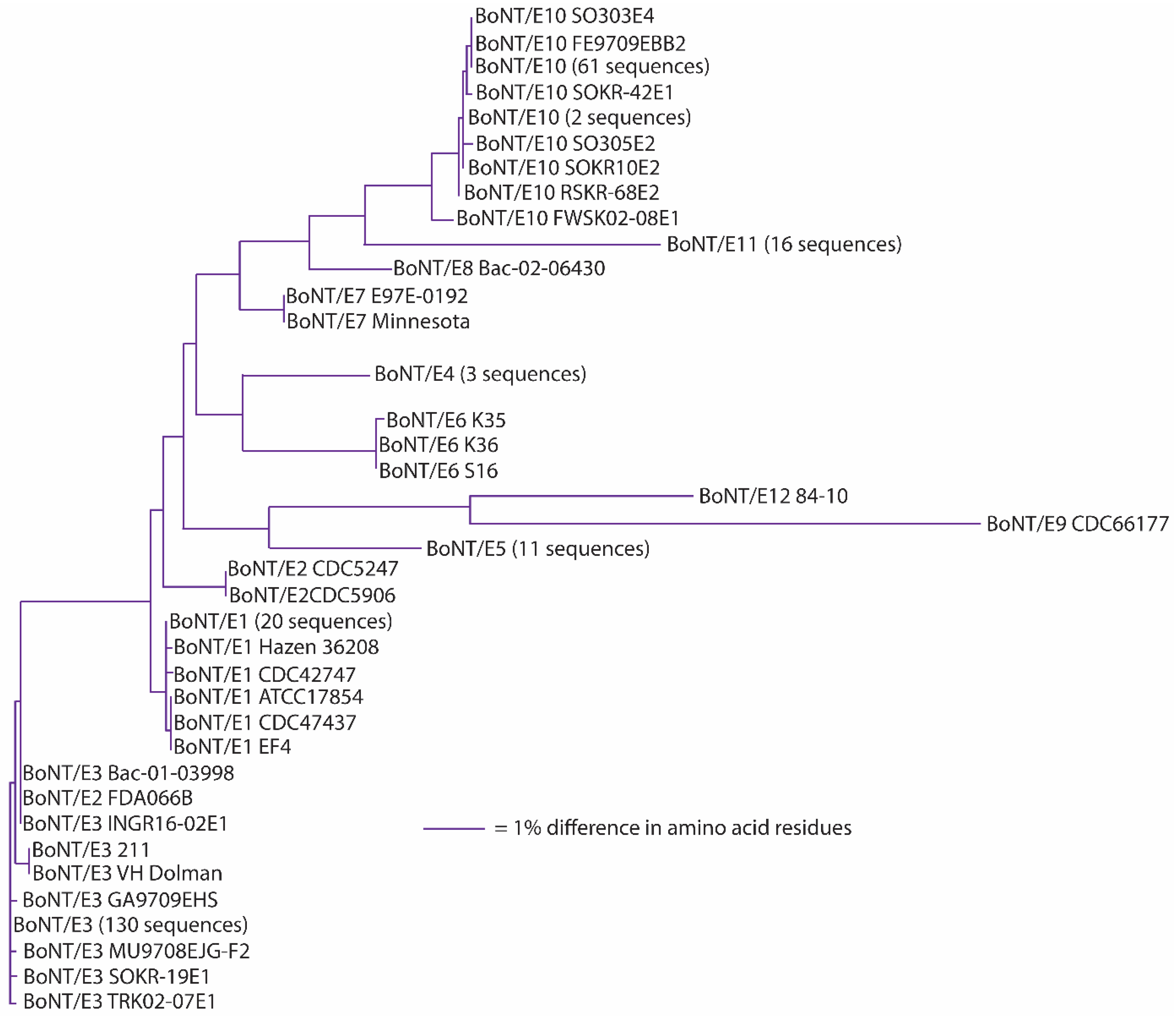
BoNT/E subtype sequences, with the exception of BoNT/E9 and BoNT/E12, are closely related (
Figure 5). Amino acid differences among BoNT/E1-BoNT/E8 range from 0.9%–5.9%, but intra-subtype differences of 0.1%–0.2% are seen within BoNT/E1, BoNT/E2, BoNT/E4-6 and BoNT/E11 subtypes (
Table 8). The lone BoNT/E9 strain sequence differs from BoNT/E1-8, BoNT/E10 and BoNT/E11 by 10.1%–11.8% in amino acid residues, making it distinctive among the BoNT/E subtypes [
78]. One issue that has arisen when distinguishing different subtypes having limited differences has been the use of phylogenetic clade analysis of neurotoxin nucleotide sequences, instead of comparisons of their amino acid differences, as the basis for discrimination [
72,
89]. While BoNT/E1, BoNT/E2 and BoNT/E3 clearly sort into distinct, but closely-related, phylogenetic clades [
89,
90], amino acid analysis of their sequences shows their differences range from 0.9%–1.8% when BoNT/E2 and BoNT/E3 are compared to BoNT/E1 (
Table 8). If the 2.6% amino acid difference guideline had been applied, this would be considered a single larger subtype. While it has been decided that historical subtype designations will continue to be used (see below), it is useful to understand that BoNT/E1, BoNT/E2 and BoNT/E3 could be considered as a single subtype entity. The effect of analyzing large numbers of sequences is seen with BoNT/E3 (toxin sequences from 143 strains) and BoNT/E10 (toxins sequences from 36 strains), where intra-subtype sequence differences were 0.6% and 0.8%, respectively. It is possible that as additional toxin sequences become available, intra-subtype differences may increase.
BoNT/B shows the greatest degree of intra-subtype variability of any serotype (
Figure 3). Inter-subtype differences range from 1.6%–7.1%, and intra-subtype differences range from 0.8%–2.1%. A clear relationship can be seen among BoNT/B2, BoNT/B3 and BoNT/B6 (within-subtype amino acid differences of 1.6%–1.9%), which is similar to the situation with BoNT/E1, BoNT/E2 and BoNT/E3. BoNT/B2, BoNT/B3 and BoNT/B6 were also initially differentiated using phylogenetic clade analysis, not amino acid differences [
72,
73], with the result that BoNT/B2, BoNT/B3 and BoNT/B6, like the BoNT/E1/E2/E3 grouping, could be considered a single subtype. In addition, with BoNT/B, the within-subtype variability is higher overall than with other toxin types, ranging from 0.8%–1.9%. The BoNT/B2 and BoNT/B4 subtypes are particularly variable (
Table 6). This unique ranging of BoNT/B2 and related subtypes may indicate that horizontal genetic interactions between certain BoNT/B-producing strains show a higher level of activity than that seen among other serotypes.
These recombination events may be the major factor responsible for the proliferation of subtypes seen within this toxin. It is important to note these events as part of toxin characterizations; however, attempts to define the toxin subtypes on the basis of these events could become challenging. A prime example is the current dispute over the nomenclature for the newly-described novel toxin known as BoNT/H, BoNT/HA or BoNT/FA, depending on how it is characterized [
10,
28,
29,
30,
31,
32,
33].
An interesting finding is the identification of a novel homolog of BoNT in the genome of a non-
Clostridium species [
91]. The homolog, named BoNT/Wo to correspond with its bacterial host (
Weissella oryzae), was verified by the Montecucco laboratory to have BoNT-like enzymatic activity [
92]. However, at the classification level, more work is required to determine whether BoNT/Wo should be considered a new family altogether or a highly divergent member of the BoNT family.
3. Historical Perspective of Botulinum Neurotoxin Forming Clostridia
Historically, the production of a botulinum neurotoxin was the only criterion for the species nomenclature for these strains, so that all botulinum neurotoxin-producing clostridia were known as
Clostridium botulinum. Today, at least six physiologically- and genetically-distinct bacteria are known to form botulinum neurotoxins [
47,
65,
93,
94,
95,
96,
97]. Currently-recognized species include
C. botulinum Groups I–IV, some strains of
C. baratii,
C. butyricum [
98] and possibly also neurotoxin-producing
C. sporogenes.
C. botulinum comprises four discrete groups of bacteria.
C. botulinum Group I (proteolytic
C. botulinum) strains are mesophilic and form spores of high heat resistance [
94].
C. botulinum Group I strains produce BoNT/A, BoNT/B, many of which were identical, and/or BoNT/F. The number of neurotoxin genes located in the Group I genomes and the number of neurotoxins produced is variable, with strains possessing up to three neurotoxin genes, and producing one or, more rarely, two or three distinct neurotoxins [
93]. Nontoxic representatives have also been isolated.
C. botulinum Group I strains are primarily responsible for human botulism.
C. botulinum Group II (non-proteolytic
C. botulinum) strains are psychrotrophic and form spores of moderate heat resistance [
94].
C. botulinum Group II strains produce either BoNT/B4, BoNT/E or BoNT/F6. Group II strains are not known to produce multiple toxins, however sequencing of the genomes of
C. botulinum Group II BoNT/F6 strains revealed that they also contain fragments of a type B and a type E neurotoxin gene [
99]. Non-toxic strains have been described [
100].
C. botulinum Group II causes human botulism. Neurotoxin encoding genes of
C. botulinum Groups I and II are located on the chromosome or on a plasmid [
3,
42,
49,
65,
75,
94,
95,
101,
102,
103,
104].
C. botulinum Group III strains, also included in
C. novyi sensu lato [
96], are mesophiles and cause botulism in various animal species. Strains form BoNT/C or BoNT/D, although more frequently a hybrid BoNT/CD or BoNT/DC neurotoxin is produced [
35,
105].
C. botulinum Group IV (also known as
C. argentinense) strains form BoNT/G, which has not been definitively associated with human or animal botulism [
106]. Some strains of
C. baratii form type F7 neurotoxin, and some strains of
C. butyricum form type E4 or E5 neurotoxin; both bacteria are associated with human botulism [
98]. It has also recently been noted that some BoNT/B-producing strains formerly thought to be within
C. botulinum Group I may be neurotoxigenic strains of
C. sporogenes [
49,
107,
108].
The earliest botulinum neurotoxins described were a type B neurotoxin formed by a strain of
C. botulinum Group II and a type A neurotoxin formed by a
C. botulinum Group I strain [
109,
110]. However, one important difference between these neurotoxins was not due to characteristics of the neurotoxins, but rather to characteristics imparted by the bacteria themselves. The neurotoxin is formed as a progenitor toxin, a single 150-kDa polypeptide. Strains of
C. botulinum Group I produce proteolytic enzymes that are responsible for post-expression processing of the neurotoxin, leading to a more active di-chain structure with a 50-kDa light chain attached to a 100-kDa heavy chain by a disulfide bond [
111]. Strains of
C. botulinum Group II lack these enzymes, and the neurotoxin remains as a single polypeptide chain to be fully activated by host proteases [
112].


 ,
, 




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}