
Presence of the Cyanotoxin Microcystin in Arctic Lakes of Southwestern Greenland

Abstract
:
{kind=link}
{kind=link}
1. Introduction
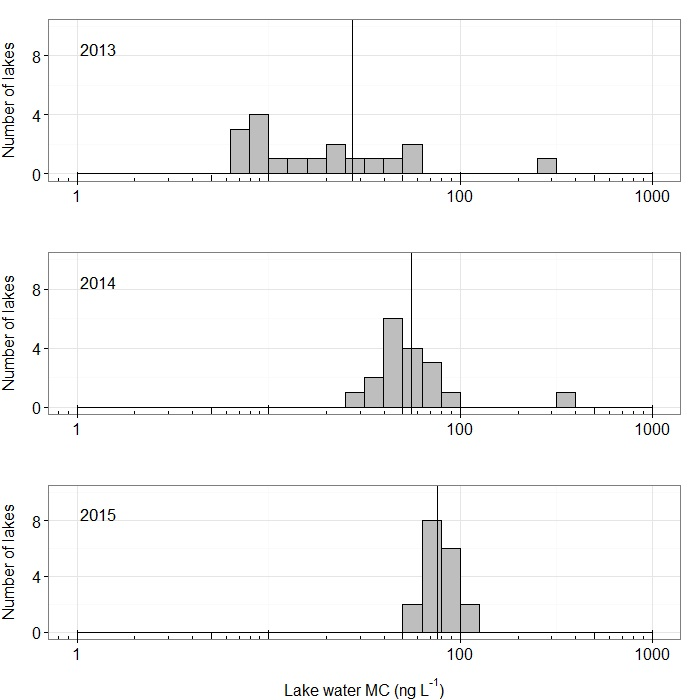
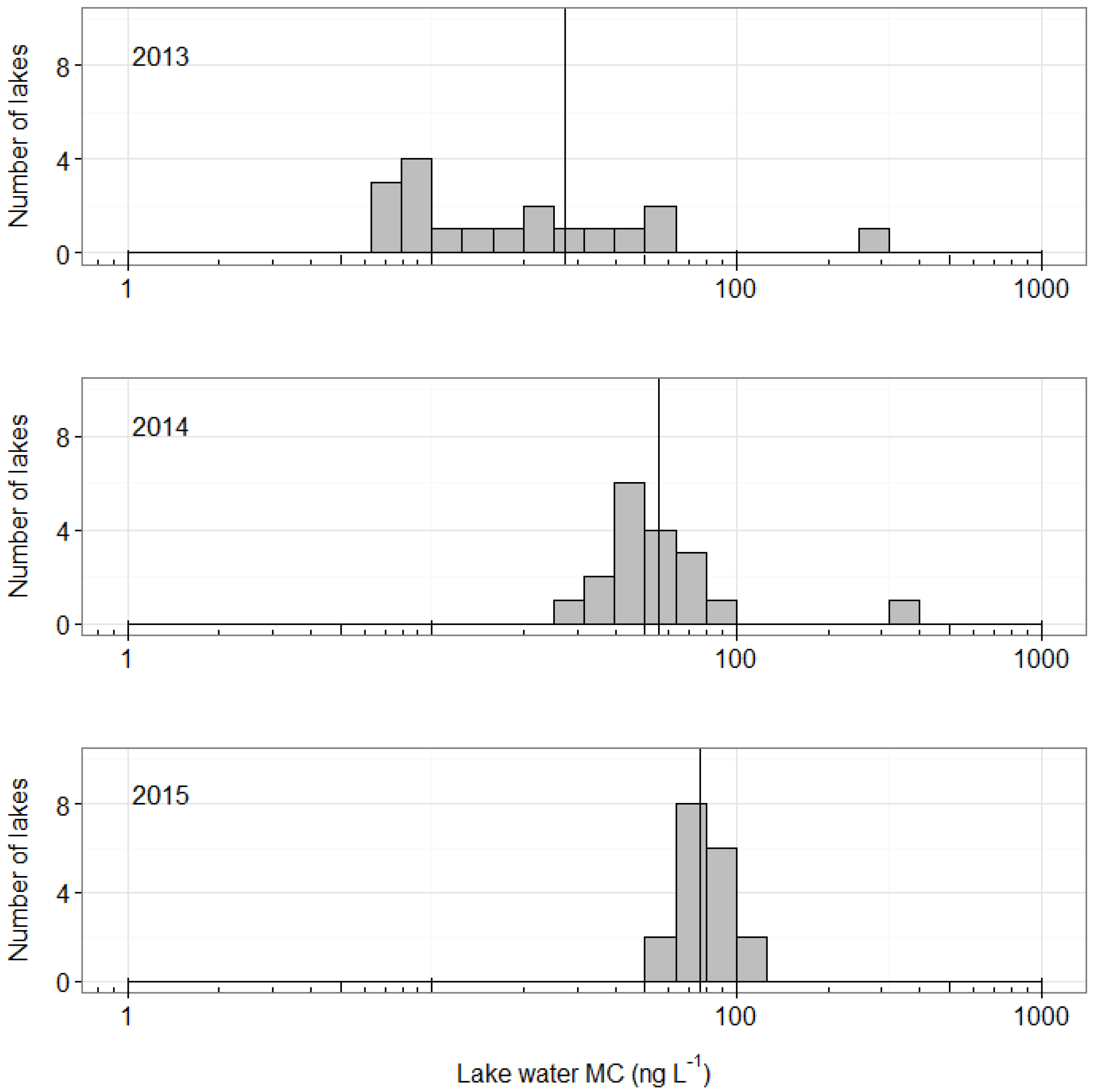
2. Results
3. Discussion
4. Materials and Methods
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| MC | Microcystin, a potent hepatotoxic cyanotoxin produced by a wide range of cyanobacterial taxa and frequently detected. MC refers to four possible microcystin variants (MC-LR, MC-LA, MC-RR, MC-YR) and nodularin toxin (this ELISA does not distinguish between microcystin toxin variants). |
References
- Sivonen, K.; Jones, G. Chapter 3: Cyanobacterial toxins. In Toxic Cyanobaceria in Water: A Guide to their Public Health Consequences, Monitoring and Management; Chorus, I., Bartram, J., Eds.; E. & F.N. Spon: London, UK, 1999. [Google Scholar]
- Paerl, H.W.; Paul, V.J. Climate change: Links to global expansion of harmful cyanobacteria. Water Res. 2012, 46, 1349–1363. [Google Scholar] [PubMed]
- Hilborn, E.D.; Beasley, V.R. One health and cyanobacteria in freshwater systems: Animal illnesses and deaths are sentinel events for human health risks. Toxins 2015, 7, 1374–1395. [Google Scholar] [PubMed]
- Sinha, R.; Pearson, L.A.; Davis, T.W.; Burford, M.A.; Orr, P.T.; Neilan, B.A. Increased incidence of Cylindrospermopsis raciborskii in temperate zones—Is climate change responsible? Water Res. 2012, 46, 1408–1419. [Google Scholar] [PubMed]
- Metcalf, J.S.; Codd, G.A. Cyanobacterial toxins (cyanotoxins) in water. Found. Water Res. 2014, 1–47. [Google Scholar]
- Chrapusta, E.; Węgrzyn, M.; Zabaglo, K.; Kaminski, A.; Adamski, M.; Wietrzyk, P.; Bialczyk, J. Microcystins and anatoxin-a in Arctic biocrust cyanobacterial communities. Toxicon 2015, 101, 35–40. [Google Scholar] [CrossRef] [PubMed]
- Hitzfeld, B.C.; Lampert, C.S.; Spaeth, N.; Mountfort, D.; Kaspar, H.; Dietrich, D.R. Toxin production in cyanobacterial mats from ponds on the McMurdo Ice Shelf, Antarctica. Toxicon 2000, 38, 1731–1748. [Google Scholar] [CrossRef]
- Jungblut, A.-D.; Hoeger, S.J.; Mountfort, D.; Hitzfeld, B.C.; Dietrich, D.R.; Neilan, B.A. Characterization of microcystin production in an Antarctic cyanobacterial mat community. Toxicon 2006, 47, 271–278. [Google Scholar] [CrossRef] [PubMed]
- Kleinteich, J.; Wood, S.A.; Küpper, F.C.; Camacho, A.; Quesada, A.; Frickey, T.; Dietrich, D.R. Temperature-related changes in polar cyanobacterial mat diversity and toxin production. Nat. Clim. Chang. 2012, 2, 356–360. [Google Scholar] [CrossRef]
- Kleinteich, J.; Wood, S.A.; Puddick, J.; Schleheck, D.; Küpper, F.C.; Dietrich, D. Potent toxins in Arctic environments—Presence of saxitoxins and an unusual microcystin variant in Arctic freshwater ecosystems. Chem. Biol. Interact. 2013, 206, 423–431. [Google Scholar] [CrossRef] [PubMed]
- Kleinteich, J.; Hildebrand, F.; Wood, S.A.; Cirés, S.; Agha, R.; Quesada, A.; Pearce, D.A.; Convey, P.; Küpper, F.C.; Dietrich, D.R. Diversity of toxin and non-toxin containing cyanobacterial mats of meltwater ponds on the Antarctic Peninsula: A pyrosequencing approach. Antarct. Sci. 2014, 26, 521–532. [Google Scholar] [CrossRef]
- Quiblier, C.; Wood, S.; Echenique-Subiabre, I.; Heath, M.; Villeneuve, A.; Humbert, J.-F. A review of current knowledge on toxic benthic freshwater cyanobacteria – Ecology, toxin production and risk management. Water Res. 2013, 47, 5464–5479. [Google Scholar]
- Wood, S.A.; Mountfort, D.; Selwood, A.I.; Holland, P.T.; Puddick, J.; Cary, S.C. Widespread distribution and identification of eight novel microcystins in Antarctic cyanobacterial mats. Appl. Environ. Microbiol. 2008, 74, 7243–7251. [Google Scholar] [CrossRef] [PubMed]
- Wood, S.A.; Kuhajek, J.M.; de Winton, M.; Phillips, N.R. Species composition and cyanotoxin production in periphyton mats from three lakes of varying trophic status. FEMS Microbiol. Ecol. 2012, 79, 312–326. [Google Scholar] [CrossRef] [PubMed]
- Callieri, C.; Cronberg, G.; Stockner, J. Chapter 8: Freshwater picocyanobacteria: Single cells, microcolonies, and colonial forms. In Ecology of Cyanobacteria II: Their Diversity in Space and Time; Whitton, B.A., Ed.; Springer: Dordrecht, The Netherlands, 2012; pp. 229–261. [Google Scholar]
- Vincent, W.R.; Quesada, A. Chapter 13: Cyanobacteria in high latitude lakes, rivers, and seas. In Ecology of Cyanobacteria II: Their Diversity in Space and Time; Whitton, B.A., Ed.; Springer: Dordrecht, The Netherlands, 2012; pp. 229–261. [Google Scholar]
- Prepas, E.E.; Kotak, B.G. Accumulation and elimination of cyanobacterial hepatotoxins by the freshwater clam Anodonta grandis simpsoniana. Can. J. Fish. Aquat. Sci. 1997, 54, 41–46. [Google Scholar]
- Tang, E.P.Y.; Tremblay, R.; Vincent, W.F. Cyanobacterial dominance of polar freshwater ecosystems. J. Phycol. 1997, 33, 171–181. [Google Scholar] [CrossRef]
- Vincent, W.F.; Hobbie, J.E. Ecology of Arctic Lakes. In The Arctic: Environment, People, Policy; Nuttall, M., Callaghan, T., Eds.; Harwood Academic Publishers: Amsterdam, The Netherlands, 2000; pp. 197–232. [Google Scholar]
- Frank, C.A.P. Microcystin-producing cyanobacteria in recreational waters in southwestern Germany. Environ. Toxicol. 2002, 17, 361–366. [Google Scholar] [CrossRef] [PubMed]
- Haney, J.; Ikawa, M. A survey of 50 NH lakes for microcystins (MCs). UNH Cent. Freshw. Biol. Res. NH Water Resources Research Center (WRRC), 2000. Available online: http://wrrc.unh.edu/sites/wrrc.unh.edu/files/media/pdf/annualreports/Haney%201998.pdf (accessed on 12 March 2014).
- Loftin, K.A.; Graham, J.L.; Hilborn, E.D.; Lehmann, S.C.; Meyer, M.T.; Dietze, J.E.; Griffith, C.B. Cyanotoxins in inland lakes of the United States: Occurrence and potential recreational health risks in the EPA National Lakes Assessment 2007. Harmful Algae 2016, 56, 77–90. [Google Scholar] [CrossRef]
- Ueno, Y.; Nagata, S.; Tsutsumi, T.; Hasegawa, A.; Yoshida, F.; Suttajit, M.; Mebs, D.; Pütsch, M.; Vasconcelos, V. Survey of microcystins in environmental water by a highly sensitive immunoassay based on monoclonal antibody. Nat. Toxins 1996, 4, 271–276. [Google Scholar] [CrossRef]
- World Health Organization Guidelines for Drinking-water Quality. Guidelines for Drinking Water Quality; Sheffer, M., Ed.; World Health Organization: Geneva, IL, USA, 2011; p. 178. Available online: http://www.who.int/water_sanitation_health/publications/2011/dwq_guidelines/en/ (accessed on 19 February 2016).
- Carmichael, W.W. Cyanobacteria secondary metabolites—The cyanotoxins. J. Appl. Bacteriol. 1992, 72, 445–459. [Google Scholar] [CrossRef] [PubMed]
- Intergovernmental Panel on Climate Change (IPCC). Contributions of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. In Climate Change 2014: Synthesis Report; Pachauri, R.K., Meyer, L.A., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2014; p. 151. [Google Scholar]
- Stendel, M.; Christensen, J.H.; Petersen, D. High-Arctic ecosystem dynamics in a changing climate. In Advances in Ecological Research; Academic Press, Elsevier Ltd.: Oxford, UK, 2008; Volume 40, pp. 13–43. [Google Scholar]
- Paerl, H.W.; Huisman, J. Climate. Blooms like it hot. Science 2008, 320, 57–58. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2015. The R Project for Statistical Computing. Available online: https://www.r-project.org/ (accessed on 20 April 2015).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer-Verlag: New York, NY, USA, 2009. [Google Scholar]
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Trout-Haney, J.V.; Wood, Z.T.; Cottingham, K.L. Presence of the Cyanotoxin Microcystin in Arctic Lakes of Southwestern Greenland. Toxins 2016, 8, 256. https://doi.org/10.3390/toxins8090256
Trout-Haney JV, Wood ZT, Cottingham KL. Presence of the Cyanotoxin Microcystin in Arctic Lakes of Southwestern Greenland. Toxins. 2016; 8(9):256. https://doi.org/10.3390/toxins8090256
Chicago/Turabian StyleTrout-Haney, Jessica V., Zachary T. Wood, and Kathryn L. Cottingham. 2016. "Presence of the Cyanotoxin Microcystin in Arctic Lakes of Southwestern Greenland" Toxins 8, no. 9: 256. https://doi.org/10.3390/toxins8090256
APA StyleTrout-Haney, J. V., Wood, Z. T., & Cottingham, K. L. (2016). Presence of the Cyanotoxin Microcystin in Arctic Lakes of Southwestern Greenland. Toxins, 8(9), 256. https://doi.org/10.3390/toxins8090256