
First Detection of Tetrodotoxin in Greek Shellfish by UPLC-MS/MS Potentially Linked to the Presence of the Dinoflagellate Prorocentrum minimum



Abstract
:1. Introduction
2. Results
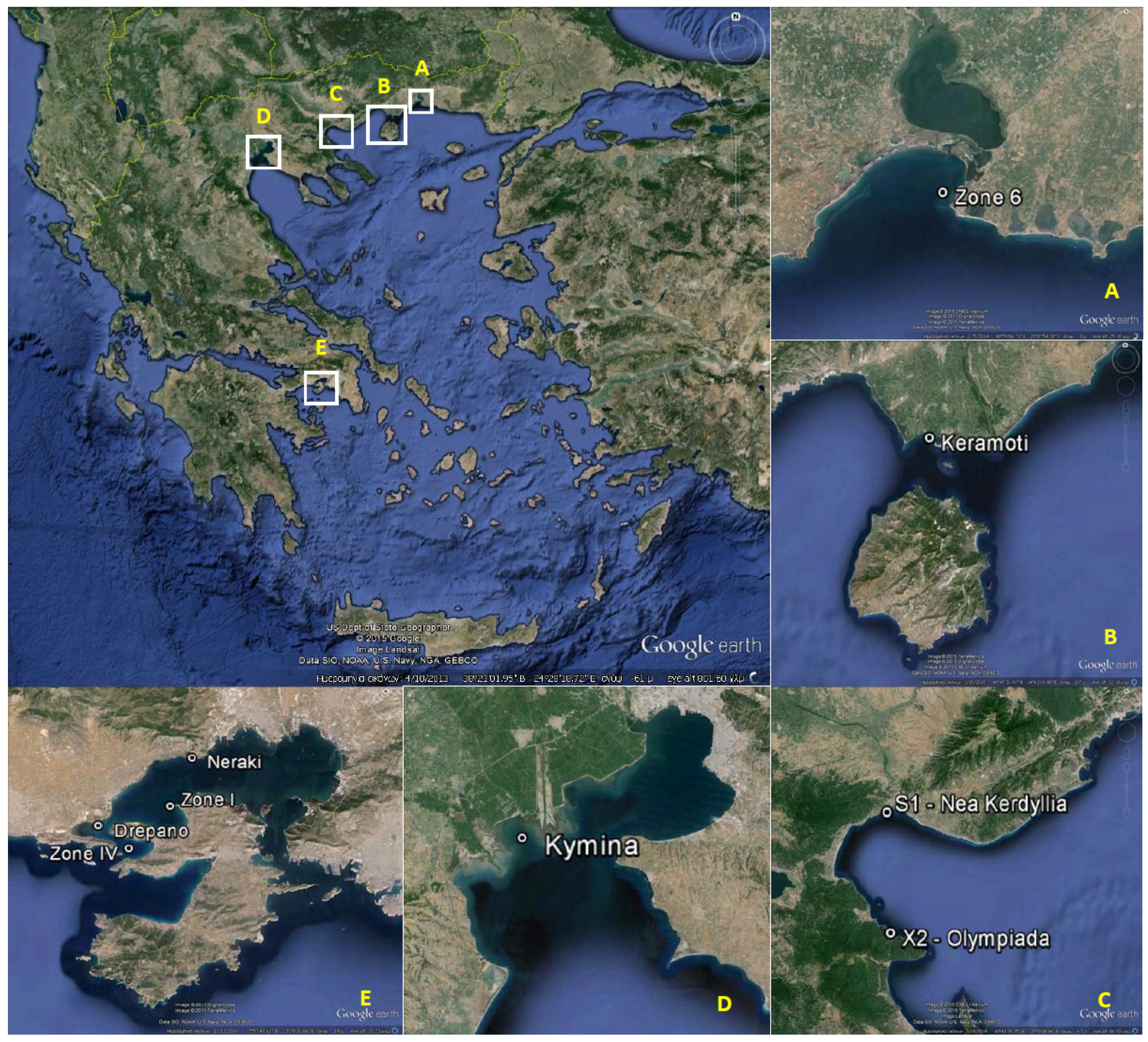
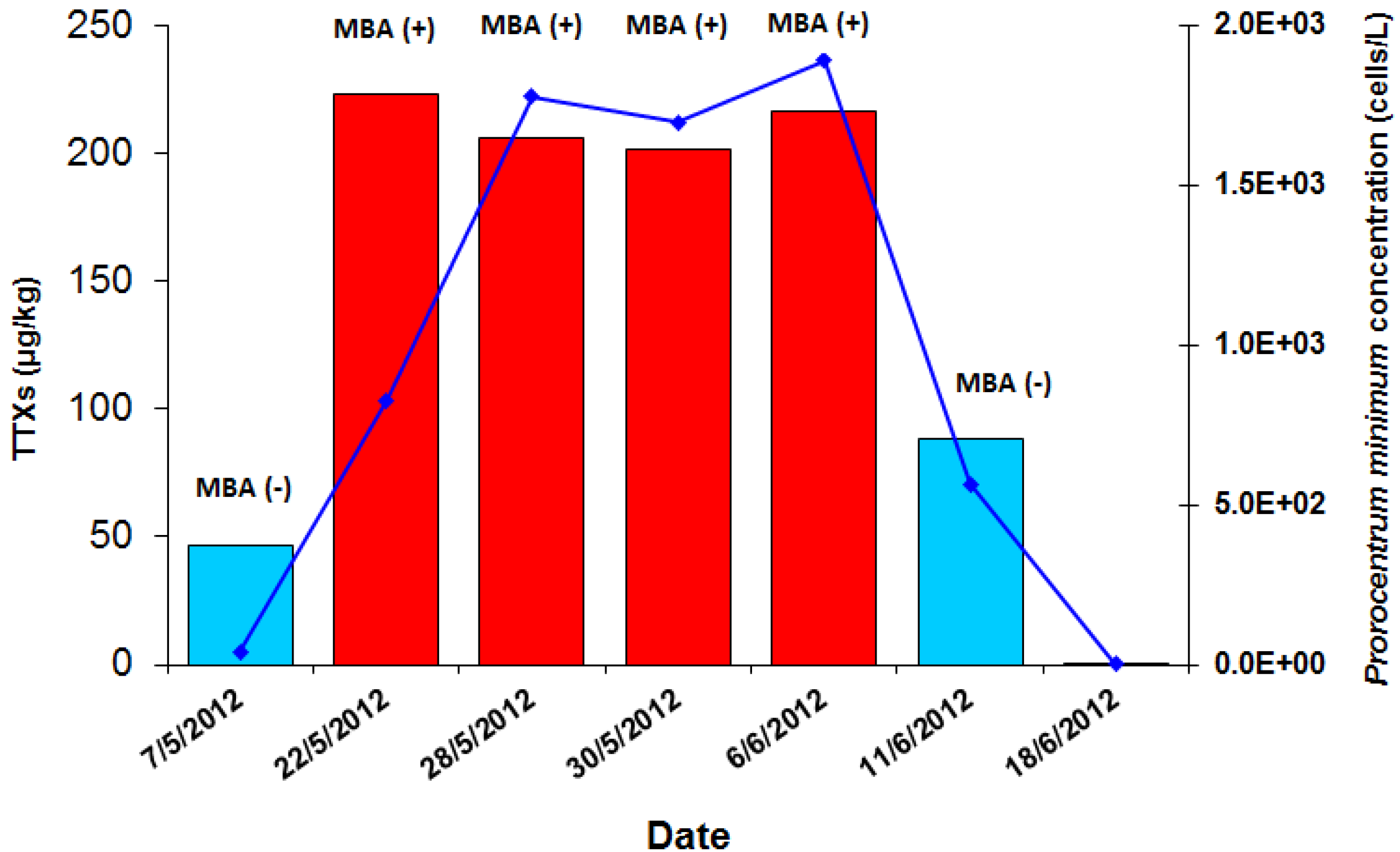
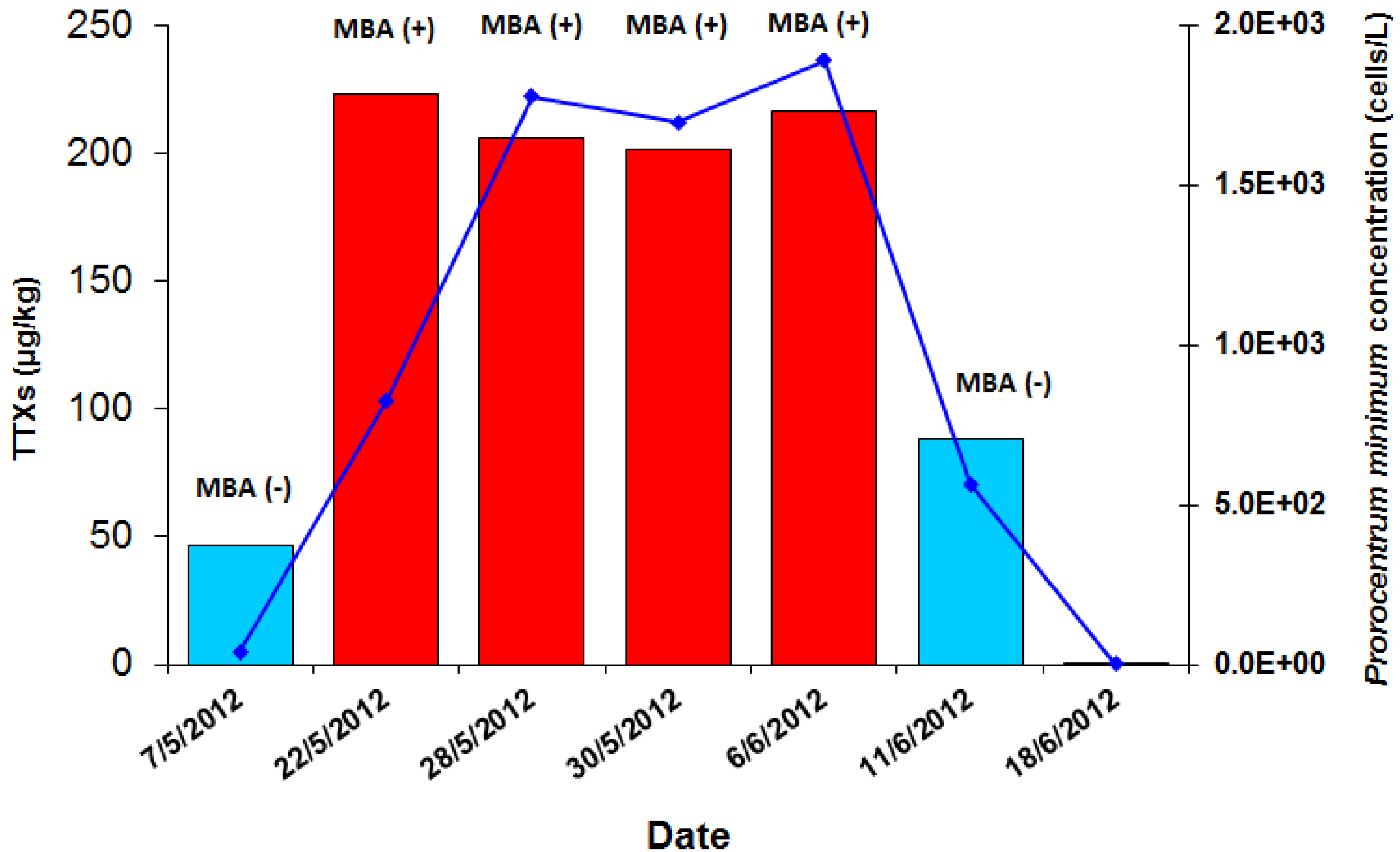
2.1. The 2012 P. minimum Episode in Vistonikos Bay–Lagos, Rodopi, Greece
2.1.1. Episode History and Preliminary Investigation

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Coastal Area | Regional Unit | Sampling Station | Latitude (N) | Longitude (E) |
|---|---|---|---|---|
| Vistonikos Bay–Lagos | Rodopi | Zone 6 | 40°58'23.73" | 25°07'06.56" |
| Keramoti Bay | Kavala | Keramoti | 40°52'00.70" | 24°40'34.60" |
| Strymonikos Bay | Serres | S1—Nea Kerdyllia | 40°46'46.49" | 23°49'50.00 |
| Chalkidiki | X2—Olympiada | 40°33'00.60" | 23°50'00.00" | |
| Thermaikos Gulf | Thessaloniki | Kymina | 40°30'00.22" | 22°41'00.06" |
| Saronikos Gulf | Attiki/Nisson | Zone I—Vasilika–Faneromeni | 37°59'42.29" | 23°27'50.06" |
| Zone IV—Agios Georgios | 37°58'09.89" | 23°25'53.92" | ||
| Dytiki Attiki/Megara | Neraki | 38°01'30.19" | 23°28'52.79" | |
| Drepano | 37°59'01.02" | 23°24'30.09" |

| Sampling data | Prorocentrum minimum (cells/L) | MBA (Yasumoto1978 [39]) | MBA (Other) | UPLC-MS/MS | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Sample code | Date of collection | Regional Unit | Site | Species | Result | Symptomatology | Protocol | Result | Symptomatology | Tissue tested | TTX (μg/kg) | 4-epi-TTX (μg/kg) | 4,9-anhydro-TTX (μg/kg) | Sum of TTXs (μg/kg) | |
| 1727/2012 | 7 May 2012 | Rodopi | Zone 6 | Mussels | Presence | Negative (− − −) | No symptomatology | N/A | N/A | N/A | DG | 46.2 | ND | ND | 46.2 |
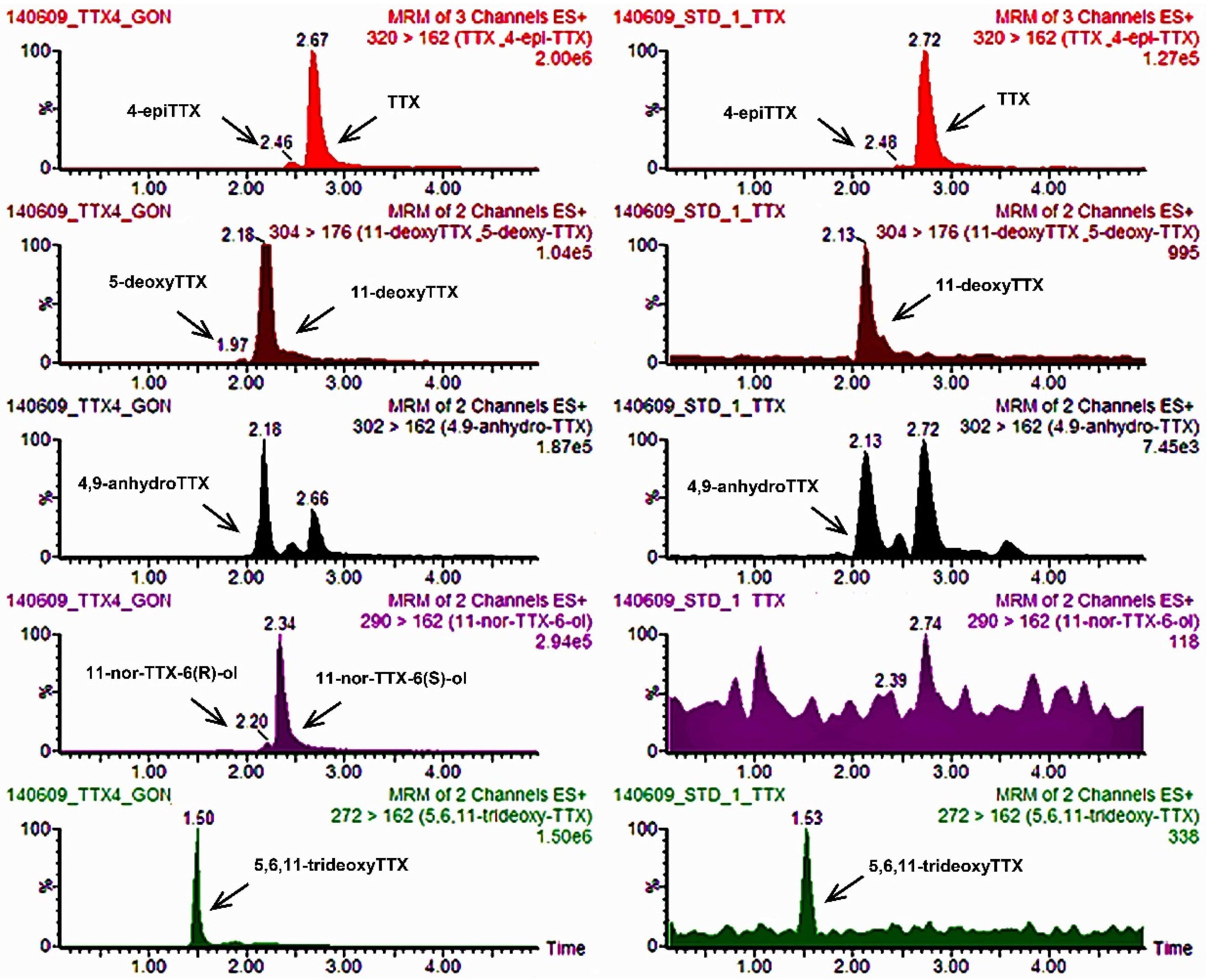
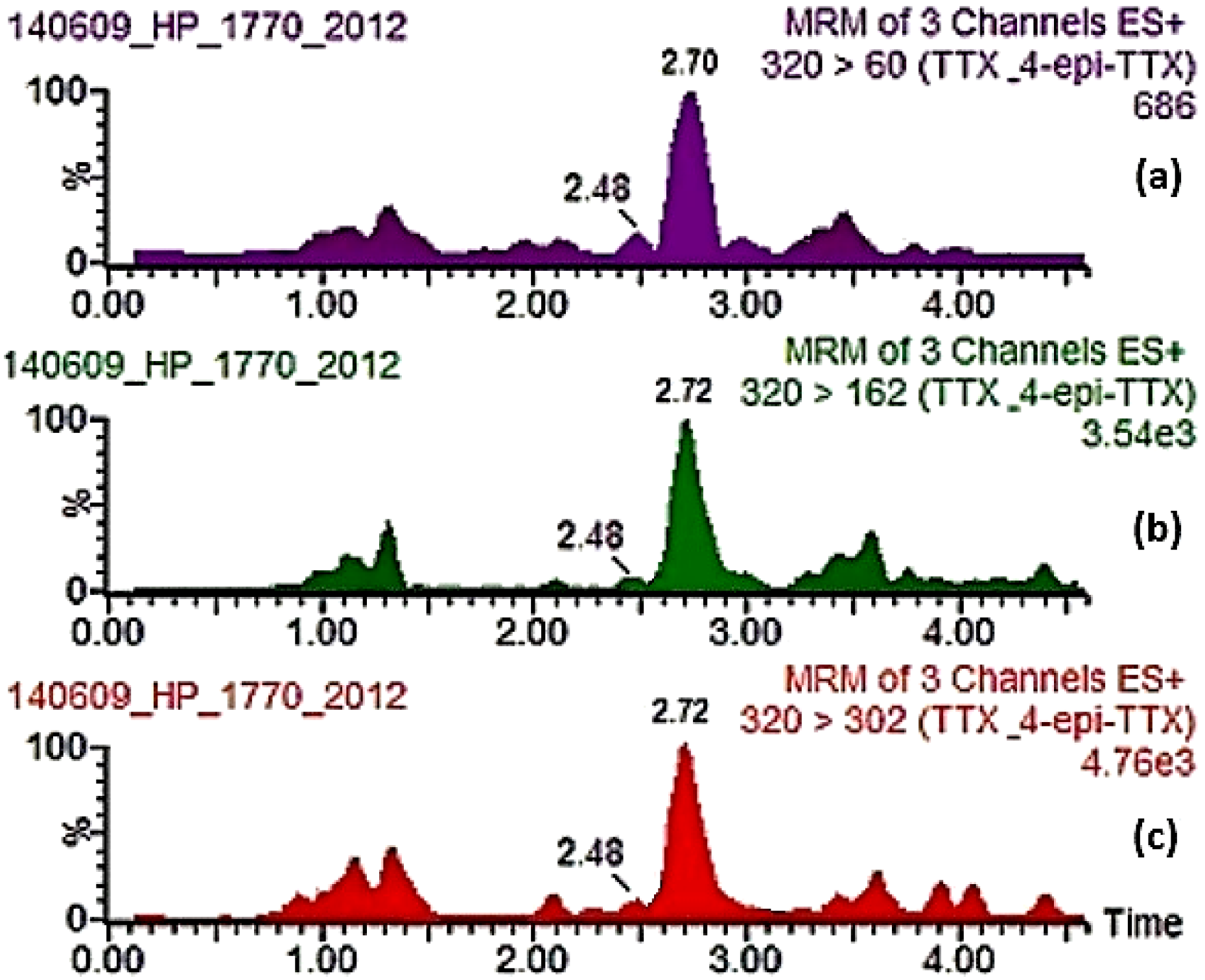
| 1770/2012 | 22 May 2012 | Rodopi | Zone 6 | Mussels | 8.26× 102 | Positive (+ + +) | 3/3 mice dead in 1:00–1:30 h | CRL-MBA-SOP v.5 (DEE fraction) | Negative (− − −) | None | DG | 202.9 | 7.6 | <LOQ (12.4) * | 222.9 |
| WF | 179.1 | <LOQ (4.2) * | <LOQ (10.3) * | 193.6 | |||||||||||
| 1774/2012 | 28 May 2012 | Rodopi | Zone 6 | Mussels | 1.78 × 103 | Positive (+ + +) | 3/3 mice dead in 1:30–2:00 h | CRL-MBA-SOP v.5 (DEE fraction) | Negative (− − −) | 2/3 mice with mild diarrhea | DG | 185.9 | <LOQ (4.5) * | <LOQ (15.9) * | 206.3 |
| 1786/2012 | 30 May 2012 | Rodopi | Zone 6 | Mussels | 1.70 × 103 | Positive (+ + +) | 3/3 mice dead (1/3 in 1:15–1:30 h, 1/3 in 1:48 h & 1/3 in 1:51 h) | CRL-MBA-SOP v.5 (DEE fraction) | Negative (− − −) | 1/3 mice with mild diarrhea | DG | 180.0 | <LOQ (6.8) * | <LOQ (14.5) * | 201.4 |
| CRL-MBA-SOP v.5 (aqueous fraction) | Positive (+ + +) | 3/3 mice dead in 7–10 min | |||||||||||||
| 1798/2012 | 6 Jun 2012 | Rodopi | Zone 6 | Mussels | 1.89 × 103 | Positive (+ + +) | 3/3 mice dead (1/3 in 1:31 h, 1/3 in 2:30 h & 1/3 in 2:31) | CRL-MBA-SOP v.5 (DEE fraction) | Negative (− − −) | None | DG | 186.2 | 15.6 | <LOQ (14.7) * | 216.6 |
| 1801/2012 | 11 Jun 2012 | Rodopi | Zone 6 | Mussels | 5.65 × 102 | Negative (− − −) | 2/3 mice with diarrhoea | CRL-MBA-SOP v.5 (ether fraction) | Negative (− − −) | 1/3 mice with diarrhea | DG | 56.5 | 8.8 | 22.9 | 88.2 |
2.1.2. Performance Parameters and Quality Control of the UPLC-MS/MS Method for TTXs
| TTX Concentration and Recovery | Tissue | TTX Fortification Level | ||||
|---|---|---|---|---|---|---|
| 100 μg/kg | 200 μg/kg | 400 μg/kg | 800 μg/kg | 1200 μg/kg | ||
| Mean TTX detected (μg/kg) | DG | 90.1 | 211.2 | 431.5 | 833.9 | 1171.5 |
| Mean Recovery (%) | 90.1 | 105.6 | 107.9 | 104.2 | 97.6 | |
| Mean TTX detected (μg/kg) | WF | 94.3 | 209.3 | 384.3 | 754.6 | 1211.0 |
| Mean Recovery (%) | 94.3 | 104.6 | 96.1 | 94.3 | 100.9 | |
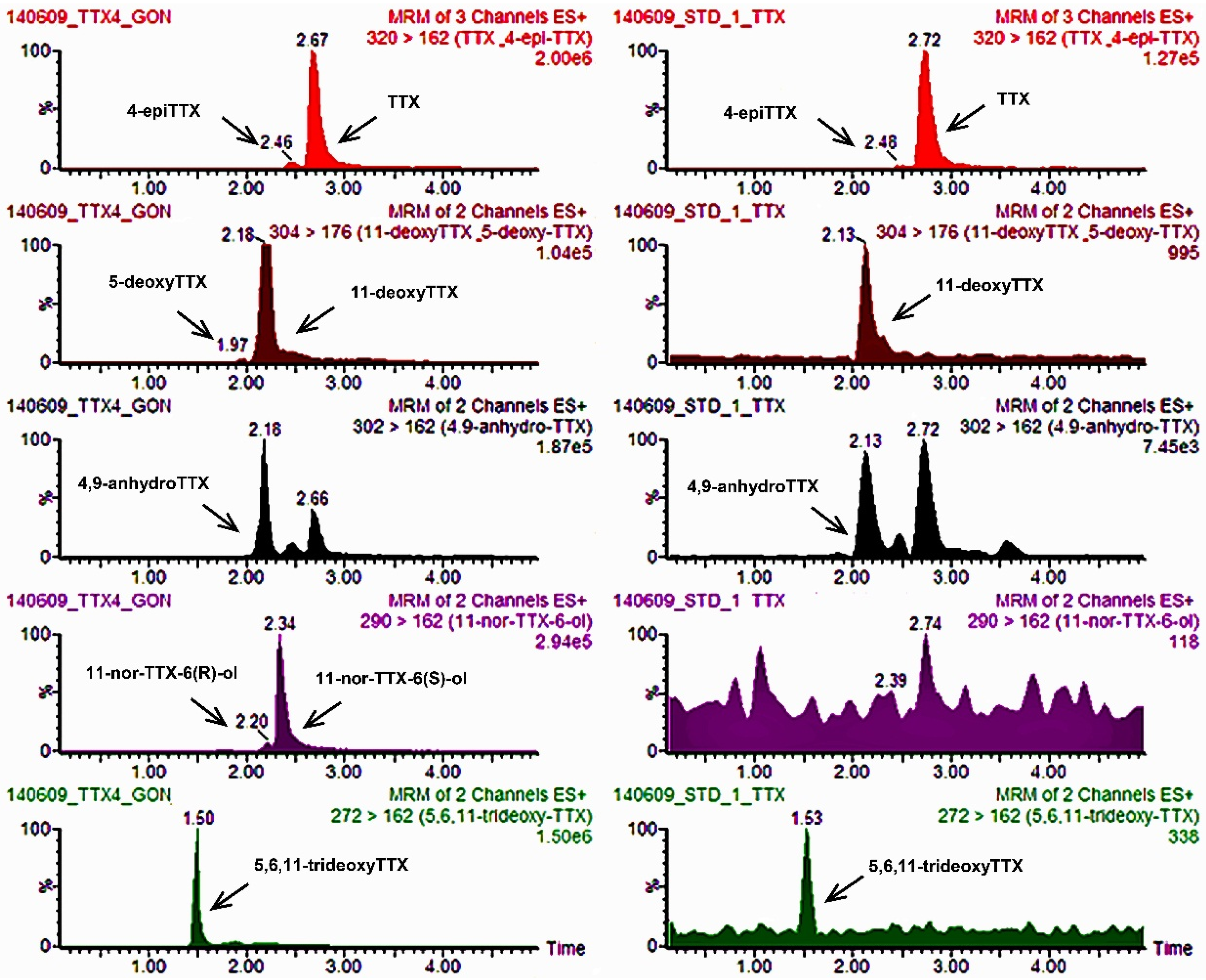
2.1.3. Application of the UPLC-MS/MS Method to the Mussel Samples of the P. minimum Episode under Investigation
2.1.4. Experimentally TTX-Contaminated Digestive Gland Tissue Tested by MBA
| TTX Fortification Level (μg/kg) | Mice Dead | Mouse Survival Time | |
|---|---|---|---|
| Median | Range | ||
| 0 | 0/6 | >24 h | >24 h |
| 50 | 0/6 | >24 h | >24 h |
| 100 | 4/6 | 310 min | 260 min→24 h |
| 200 | 6/6 | 85 min | 72–98 min |
| 360 | 6/6 | 33 min | 22–37 min |
| 1000 | 6/6 | 9 min | 7.5–10 min |
2.2. Monitoring of Vistonikos Bay–Lagos and Neighboring Coastal Areas in 2014
2.3. Investigation of Routine Shellfish Samples from 2006–2012 Obtained in Periods of P. minimum Increased Presence
| Sampling Data | MBA (Yasumoto1978) | UPLC-MS/MS | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Year | Sample Code | Date of Collection | Regional Unit | Site | Species | Result (24 h) | Symptomatology | Tissue Tested | TTX (μg/kg) | 4-epi-TTX (μg/kg) | 4,9-anhydro-TTX (μg/kg) | Sum of TTXs (μg/kg) |
| 2014 | 2084/2014 | 08 April 2014 | Rodopi | Zone 6 | Mussels | Negative (− − −) | None | WF | 43.4 | ND | ND | 43.4 |
| 2014 | 2126/2014 | 24 April 2014 | Rodopi | Zone 6 | Mussels | Negative (− − −) | None | WF | 38.9 | ND | ND | 38.9 |
| 2014 | 2141/2014 | 29 April 2014 | Rodopi | Zone 6 | Mussels | Negative (− − −) | None | WF | 34.9 | ND | ND | 34.9 |
| 2014 | 2167/2014 | 07 May 2014 | Rodopi | Zone 6 | Mussels | Negative (− − −) | None | WF | 37.7 | ND | ND | 37.7 |
| 2014 | 2172/2014 | 08 May 2014 | Rodopi | Zone 6 | Mussels | Negative (− − −) | None | WF | 34.8 | ND | ND | 34.8 |
| 2014 | 2203/2014 | 20 May 2014 | Rodopi | Zone 6 | Mussels | Negative (− − −) | None | WF | 30.1 | ND | ND | 30.1 |
| 2014 | 2239/2014 | 28 May 2014 | Rodopi | Zone 6 | Mussels | Negative (− − −) | 2/3 mice with mild diarrhea | WF | 25.8 | ND | ND | 25.8 |
| 2014 | 2232/2014 | 28 May 2014 | Serres | S1: Nea Kerdyllia | Mussels | Negative (− − −) | None | WF | 37.9 | 9.1 | ND | 47.0 |
| 2014 | 2234/2014 | 28 May 2014 | Chalkidiki | X2: South Strymonikos Gulf—Olympiada | Mussels | Negative (− − −) | None | WF | 30.6 | <LOQ (3.2) * | ND | 33.8 |
| 2014 | 2270/2014 | 11 June 2014 | Rodopi | Zone 6 | Mussels | Negative (− − −) | 3/3 mice with diarrhea | WF | 28.4 | ND | ND | 28.4 |
| 2014 | 2290/2014 | 19 June 2014 | Rodopi | Zone 6 | Mussels | Positive (− + +) | 1/3 mice dead in 3:30–19:30 h, 1/3 dead in 21:30–22:00 h | WF | 36.8 | ND | ND | 36.8 |
| 2014 | 2310/2014 | 25 June 2014 | Rodopi | Zone 6 | Mussels | Negative (− − −) | 3/3 mice with bad clinical condition | WF | 25.1 | ND | ND | 25.1 |
| 2014 | 2319/2014 | 02 July 2014 | Rodopi | Zone 6 | Mussels | Negative (− − −) | 3/3mice with bad clinical condition and diarrhea | WF | 33.3 | ND | ND | 33.3 |
| 2014 | 2336/2014 | 08 July 2014 | Rodopi | Zone 6 | Mussels | Negative (− − −) | 3/3 mice with diarrhea | WF | 25.0 | ND | ND | 25.0 |
| 2014 | 2362/2014 | 16 July 2014 | Rodopi | Zone 6 | Mussels | Negative (− − −) | None | WF | 37.6 | ND | ND | 37.6 |
| Sampling Data | Prorocentrum minimum (cells/L) | MBA (Yasumoto1978) | UPLC-MS/MS | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Year | Sample Code | Date of Collection | Regional Unit | Site | Species | Result (24 h) | Symptomatology | Tissue Tested | TTX (μg/kg) | 4-epi-TTX (μg/kg) | 4,9-anhydro-TTX (μg/kg) | Sum of TTXs (μg/kg) | |
| 2006 | 1357/2006 | 20 March 2006 | Dytiki Attiki/ Megara | Drepano | Mussels | No data provided | Positive (+ + +) | 3/3 mice dead in <15 h | DG | 70.7 | <LOQ (6.7) * | ND | 77.4 |
| 2006 | 1366/2006 | 11 April 2006 | Dytiki Attiki/ Megara | Neraki | Mussels | 3.50 × 104 | Positive (+ + −) | 2/3 mice dead in <15 h | DG | 68.4 | <LOQ (3.4) * | ND | 71.8 |
| 2006 | 1367/2006 | 11 April 2006 | Dytiki Attiki/ Megara | Drepano | Mussels | 7.50 × 104 | Negative (+ − −) | 1/3 mice dead in <15 h, 1/3 mild diarrhea | DG | 61.2 | <LOQ (7.5) * | ND | 68.8 |
| 2008 | 1547/2008 | 9 September 2008 | Dytiki Attiki/ Megara | Drepano | Mussels | 7.50 × 104 | Positive (+ + −) | 1/3 mice dead in 2:00–12:30 h, 1/3 in 23:00–23:45 h, 1/3 with bad clinical condition and diarrhea (and death in 26:00–36:30 h) | DG | 55.4 | <LOQ (4.7) * | <LOQ (12.4) * | 72.5 |
| 2008 | 1583/2008 | 6 October 2008 | Dytiki Attiki/ Megara | Neraki | Mussels | No data provided. Bloom in previous month | Negative (− − −) | 3/3 mice with bad clinical condition and diarrhea (and death 2/3 in 26:00–35:30 h and 1/3 in 42:30–44:00 h) | DG | 52.9 | <LOQ (4.3) * | <LOQ (10.8) * | 68.0 |
| 2008 | 1590/2008 | 13 October 2008 | Dytiki Attiki/ Megara | Neraki | Mussels | No data provided. Bloom in previous month | Positive (+ + +) | 1/3 mice dead in 2:00–2:30 h, 1/3 in 2:20–2:40 h, 1/3 in 2:40–3:30 h | DG | 65.6 | <LOQ (4.7) * | <LOQ (15.1) * | 85.4 |
| 2008 | 1594/2008 | 14 October 2008 | Attiki/Nisson | Salamina—Zone Ι—Vasilika–Faneromeni | Venus clams | Bloom in the wider area during the previous month (Neraki, Drepano) | Positive (+ + +) | 3/3 mice dead in 2:00–2:30 h | DG | 176.5 | <LOQ (3.9) * | <LOQ (14.3) * | 194.7 |
| 2008 | 1607/2008 | 23 October 2008 | Dytiki Attiki/ Megara | Neraki | Mussels | 3.68 × 105 | Negative (− − −) | 2/3 bad clinical condition and diarrhea, 1/3 bad clinical condition and mild diarrhea | DG | 62.6 | <LOQ (4.9) * | <LOQ (18.6) * | 86.2 |
| 2008 | 1625/2008 | 4 November 2008 | Attiki/Nisson | Salamina—Zone ΙV—Agios Georgios | Mussels | Simultaneous bloom in the wider area (Neraki: 1.11 × 104) | Negative (− − −) | 1/3 mice with bad clinical condition and diarrhea, 2/3 bad clinical condition and mild diarrhea (and death 1/3 in 27:35–37:10 h and 1/3 in 45:00–46:20 h) | DG | 57.2 | <LOQ (3.8) * | ND | 61.0 |
| 2009 | 1544/2009 | 16 June 2009 | Rodopi | Zone 6 | Mussels | Bloom in the previous month (6 May 2009: 1.13 × 105 & 12 May 2009: 8.24 × 103) | Positive (+ + +) | 2/3 mice dead in 13:00–21:00 h, 1/3 in 21:00–23:00 h | DG | 46.7 | <LOQ (5.4) * | <LOQ (19.2) * | 71.3 |
| 2009 | 1552/2009 | 22 June 2009 | Rodopi | Zone 6 | Mussels | No data provided | Negative (− − −) | 3/3 mice with bad clinical condition and diarrhea (and death 3/3 in 29:20–46:20 h) | DG | 48.1 | <LOQ (5.0) * | <LOQ (15.9) * | 69.0 |
| 2009 | 1556/2009 | 24 June 2009 | Rodopi | Zone 6 | Mussels | No data provided | Positive (+ + −) | 2/3 mice dead in 1:30–2:00 h, 1/3 with bad clinical condition and diarrhea (and death in 30:00–48:00 h) | DG | 52.6 | <LOQ (3.5) * | <LOQ (8.6) * | 64.8 |
| 2012 | 2057/2012 | 6 November 2012 | Rodopi | Zone 6 | Mussels | No data provided. Bloom reported in the following 2 weeks, max. 5.36 × 105 | Negative (− − −) | 2/3 mice with bad clinical condition and mild diarrhea, 1/3 mice with bad clinical condition | DG | 68.5 | <LOQ (3.4) * | <LOQ (12.0) * | 83.9 |
| 2012 | 1216/2012 | 22 November 2012 | Kavala | Keramoti | Mussels | No data provided. Bloom reported in the following weeks | Positive (+ − +) | 2/3 mice dead in 4:00–20:00 h, 1/3 mice with bad clinical condition | DG | 71.0 | <LOQ (3.4) * | <LOQ (20.7) * | 95.0 |
| 2012 | 1224/2012 | 10 December 2012 | Kavala | Keramoti | Mussels | 3.13 × 104 | Negative (− − +) | 1/3 mice dead in 2:00–18:00 h, 2/3 mice with bad clinical condition and mild diarrhea | DG | 71.2 | <LOQ (5.2) * | <LOQ (11.7) * | 88.2 |
| 2012 | 1227/2012 | 17 December 2012 | Kavala | Keramoti | Mussels | 1.56 × 104 | Negative (− − −) | 2/3 mice with bad clinical condition and mild diarrhea, 1/3 mice with bad clinical condition | DG | 91.8 | <LOQ (4.3) * | <LOQ (13.2) * | 109.2 |
| 2012 | 1241/2012 | 3 December 2012 | Thessaloniki | Kymina | Mussels | 174 (in wider area 1.75 × 105) | Negative (− − −) | None | WF | 79.5 | <LOQ (5.6) * | <LOQ (18.0) * | 103.1 |
| Sampling Data | MBA (Yasumoto1978) | LC-MS/MS | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Year | Sample Code | Date of Collection | Regional Unit | Site | Species | Result (24 h) | Symptomatology | Tissue Tested | TTX (μg/kg) | 4-epi-TTX (μg/kg) | 4,9-anhydro-TTX (μg/kg) |
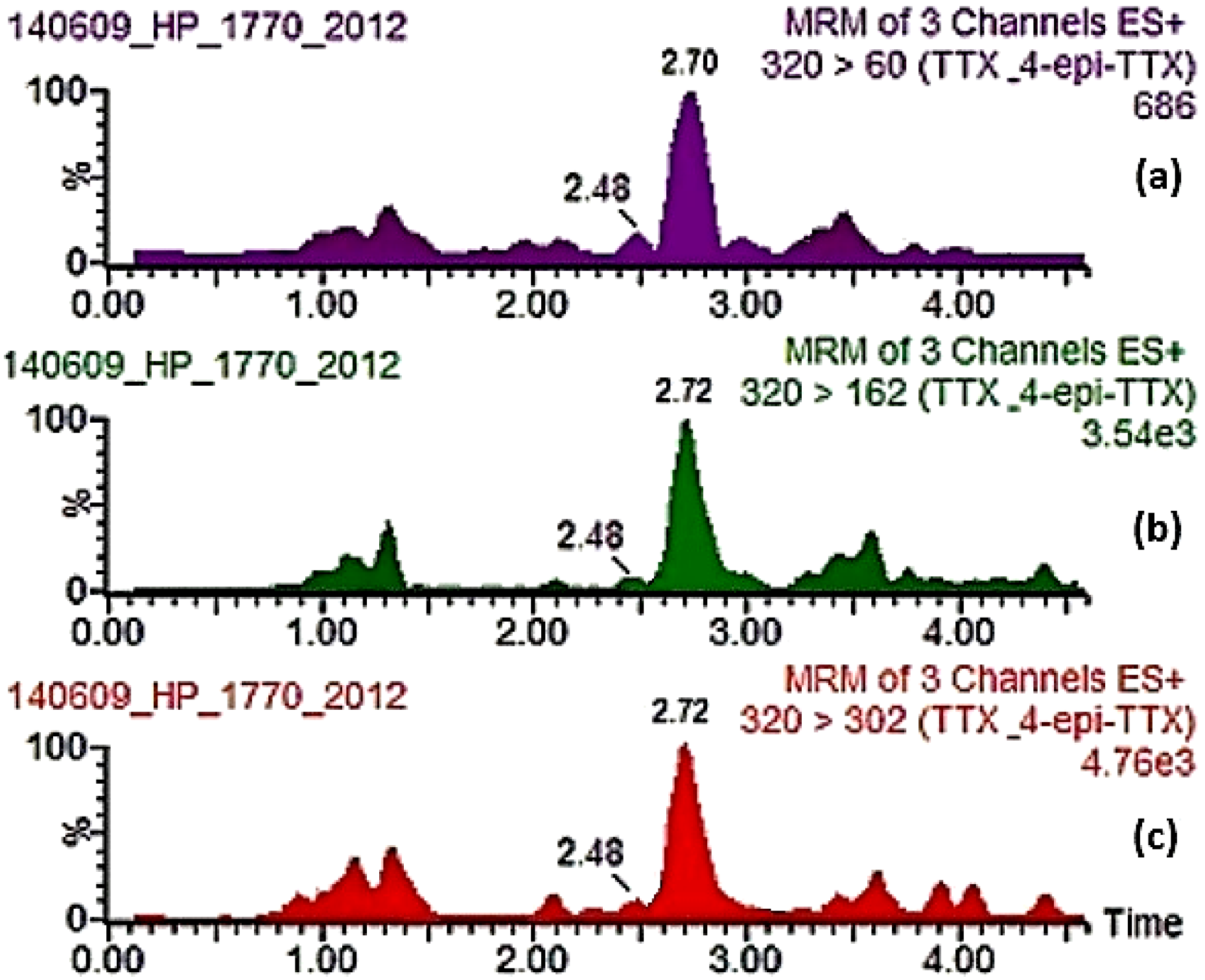
| 2014 | 2303/2014 | 23 June 2014 | Chalkidiki | X2: South Strymonikos Gulf—Olympiada | Mussels | Negative (− − −) | 3/3 mice with diarrhea | HP/WF | <LOQ | ND | ND |
| 2014 | 272/2014 | 30 June 2014 | Thessaloniki | Kymina | Mussels | Negative (− − −) | None | HP/WF | ND | ND | ND |
| 2014 | 2313/2014 | 30 June 2014 | Dytiki Attiki/ Megara | Drepano | Mussels | Negative (− − −) | 3/3 mice with bad clinical condition and diarrhea | HP | ND | ND | ND |
| 2014 | 2315/2014 | 30 June 2014 | Dytiki Attiki/ Megara | Neraki | Mussels | Negative (− − −) | None | HP/WF | ND | ND | ND |
| 2014 | 1303/2014 | 01 July 2014 | Kavala | Keramoti | Mussels | Negative (− − −) | None | HP/WF | <LOQ | ND | ND |
| 2014 | 2323/2014 | 01 July 2014 | Serres | S1: Nea Kerdyllia | Mussels | Negative (− − −) | None | HP/WF | <LOQ | ND | ND |
| 2014 | 283/2014 | 07 July 2014 | Thessaloniki | Kymina | Mussels | Negative (− − −) | 1/3 mice with mild diarrhea | HP/WF | ND | ND | ND |
| 2014 | 2332/2014 | 07 July 2014 | Dytiki Attiki/ Megara | Drepano | Mussels | Negative (− − −) | 3/3 mice with bad clinical condition | HP | ND | ND | ND |
| 2014 | 2334/2014 | 07 July 2014 | Dytiki Attiki/ Megara | Neraki | Mussels | Negative (− − −) | None | HP/WF | ND | ND | ND |
| 2014 | 2337/2014 | 07 July 2014 | Chalkidiki | X2: South Strymonikos Gulf—Olympiada | Mussels | Negative (− − −) | 1/3 mice with bad clinical condition | HP | <LOQ | ND | ND |
| 2014 | 1307/2014 | 08 July 2014 | Kavala | Keramoti | Mussels | Negative (− − −) | None | HP | ND | ND | ND |
| 2014 | 2342/2014 | 08 July 2014 | Serres | S1: Nea Kerdyllia | Mussels | Negative (− − −) | 1/3 mice with diarrhea | HP | ND | ND | ND |
| 2014 | 297/2014 | 14 July 2014 | Thessaloniki | Kymina | Mussels | Negative (− − −) | 1/3 mice with diarrhea, 2/3 mild diarrhea | HP | ND | ND | ND |
| 2014 | 1311/2014 | 15 July 2014 | Kavala | Keramoti | Mussels | Negative (− − −) | 1/3 mice with bad clinical condition | HP | ND | ND | ND |
| 2014 | 2360/2014 | 15 July 2014 | Dytiki Attiki/ Megara | Drepano | Venus clams | Negative (− + −) | 1/3 mice dead in 1:05 h, 1/3 diarrhea | HP/WF | ND | ND | ND |
| 2014 | 2368/2014 | 15 July 2014 | Serres | S1: Nea Kerdyllia | Mussels | Negative (− − −) | None | HP/WF | ND | ND | ND |
2.4. Investigation of Random Routine Shellfish Samples Obtained in June–July 2014
3. Discussion
3.1. UPLC-MS/MS Method Performance
3.2. Presence of TTXs in Greek Shellfish
3.3. Comparison of Tetrodotoxin to Venerupin Shellfish Poisoning Toxin
4. Experimental Section
4.1. Standards and Reagents
4.2. Sampling and Test Materials
4.3. Seawater Analyses
4.4. Shellfish Preparation and Sub-Sampling
4.5. Mouse Bioassay Tests
4.6. UPLC-MS/MS Analysis (Ultra-Performance Liquid Chromatography Coupled to Tandem Mass Spectrometry)
4.6.1. Sample Treatment
4.6.2. UPLC-MS/MS Method
4.6.3. UPLC-MS/MS Method Quality Control
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Narahashi, T. Pharmacology of tetrodotoxin. J. Toxicol. Toxin Rev. 2001, 20, 67–84. [Google Scholar] [CrossRef]
- Field, J. Puffer fish poisoning. J. Accid. Emerg. Med. 1998, 15, 334–336. [Google Scholar] [CrossRef] [PubMed]
- Noguchi, T.; Ebesu, J.S.M. Puffer poisoning: Epidemiology and treatment. J. Toxicol. Toxin Rev. 2001, 20, 1–10. [Google Scholar] [CrossRef]
- Yin, H.L.; Lin, H.S.; Huang, C.C.; Hwang, D.F.; Liu, J.S.; Chen, W.H. Tetrodotoxication with Nassauris glans: A possibility of tetrodotoxin spreading in marine products near Pratas Island. Am. J. Trop. Med. Hyg. 2005, 73, 985–990. [Google Scholar] [PubMed]
- Noguchi, T.; Arakawa, O. Tetrodotoxin—Distribution and accumulation in aquatic organisms, and cases of human intoxication. Mar. Drugs 2008, 6, 220–242. [Google Scholar] [CrossRef] [PubMed]
- Mebs, D.; Yotsu-Yamashita, M.; Yasumoto, T.; Lotters, S.; Schluter, A. Further report of the occurrence of tetrodotoxin in Atelopus species (family: Bufonidae). Toxicon 1995, 33, 246–249. [Google Scholar] [CrossRef] [PubMed]
- Katikou, P.; Georgantelis, D.; Sinouris, N.; Petsi, A.; Fotaras, T. First report on toxicity assessment of the Lessepsian migrant pufferish Lagocephalus sceleratus (Gmelin, 1789) from European waters (Aegean Sea, Greece). Toxicon 2009, 54, 50–55. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, P.; Alfonso, A.; Otero, P.; Katikou, P.; Georgantelis, D.; Botana, L.M. Liquid chromatography–mass spectrometry method to detect Tetrodotoxin and its analogues in the puffer fish Lagocephalus sceleratus (Gmelin, 1789) from European waters. Food Chem. 2012, 132, 1103–1111. [Google Scholar] [CrossRef]
- Bane, V.; Lehane, M.; Dikshit, M.; O’Riordan, A.; Furey, A. Tetrodotoxin: Chemistry, toxicity, source, distribution and detection. Toxins 2014, 6, 693–755. [Google Scholar] [CrossRef] [PubMed]
- McNabb, P.S.; Taylor, D.I.; Ogilvie, S.C.; Wilkinson, L.; Anderson, A.; Hamon, D.; Wood, S.A.; Peake, B.M. First detection of tetrodotoxin in the bivalve Paphies australis by liquid chromatography coupled to triple quadrupole mass spectrometry with and without precolumn reaction. J. AOAC Int. 2014, 97, 325–333. [Google Scholar] [CrossRef] [PubMed]
- Kodama, M.; Sato, S.; Ogata, T. Alexandrium tamarense as a source of Tetrodotoxin in the scallop Patinopecten yessoensis. In Toxic Phytoplankton Blooms in the Sea; Smayda, T.J., Shimizu, Y., Eds.; Elsevier Science Publishers: Amsterdam, The Netherlands, 1993; pp. 401–406. [Google Scholar]
- Rodriguez, P.; Alfonso, A.; Vale, C.; Alfonso, C.; Vale, P.; Tellez, A.; Botana, L.M. First toxicity report of tetrodotoxin and 5,6,11-trideoxyTTX in the trumpet shell Charonia lampas lampas in Europe. Anal. Chem. 2008, 80, 5622–5629. [Google Scholar] [CrossRef] [PubMed]
- Turner, A.D.; Powell, A.; Schofield, A.; Lees, D.N.; Baker-Austin, C. Detection of the pufferfish toxin tetrodotoxin in European bivalves, England, 2013 to 2014. Euro Surveill. 2015, 20. Available online: http://www.eurosurveillance.org/ViewArticle.aspx?ArticleId=21009 (accessed on 25 February 2015). [Google Scholar]
- McNabb, P.; Selwood, A.I.; Munday, R.; Wood, S.A.; Taylor, D.I.; MacKenzie, L.A.; van Ginkel, R.; Rhodes, L.L.; Cornelisen, C.; Heasman, K.; et al. Detection of tetrodotoxin from the grey side-gilled sea slug—Pleurobranchaea maculata, and associated dog neurotoxicosis on beaches adjacent to the Hauraki Gulf, Auckland, New Zealand. Toxicon 2010, 3, 466–473. [Google Scholar] [CrossRef]
- Yotsu-Yamashita, M.; Mebs, D.; Flachsenberger, W. Distribution of tetrodotoxin in the body of the blue-ringed octopus (Hapalochlaena maculosa). Toxicon 2007, 49, 410–412. [Google Scholar] [CrossRef] [PubMed]
- Noguchi, T.; Arakawa, O.; Takatani, T. TTX accumulation in pufferfish. Comp. Biochem. Physiol. 2006, 1, 145–152. [Google Scholar]
- Croci, L.; Cozzi, L.; Suffredini, E.; Ciccaglioni, G.; Toti, L.; Milandri, A. Characterization of microalgae and associated bacteria collected from shellfish harvesting areas. Harmful Algae 2006, 5, 266–274. [Google Scholar] [CrossRef]
- Wu, Z.; Xie, L.; Xia, G.; Zhang, J.; Nie, Y.; Hu, J.; Wang, S.; Zhang, R. A new tetrodotoxin-producing actinomycete, Nocardiopsis dassonvillei, isolated from the ovaries of puffer fish Fugu rubripes. Toxicon 2005, 45, 851–859. [Google Scholar] [CrossRef] [PubMed]
- Noguchi, T.; Onuki, K.; Arakawa, O. Tetrodotoxin poisoning due to pufferfish and gastropods, and their intoxication mechanism. ISRN Toxicol. 2011. Available online: http://www.hindawi.com/journals/isrn/2011/276939/ (accessed on 25 February 2015). [Google Scholar] [CrossRef]
- Nakazima, M. Studies on the source of shellfish poison in Lake Hamana. I. Relation of the abundance of a species of dinoflagellate, Prorocentrum sp. to shellfish toxicity. Bull. Jpn. Soc. Sci. Fish. 1965, 31, 198–203. [Google Scholar] [CrossRef]
- Heil, C.A.; Glibert, P.M.; Fan, C. Prorocentrum minimum (Pavillard) Schiller: A review of a harmful algal bloom species of growing worldwide importance. Harmful Algae 2005, 4, 449–470. [Google Scholar] [CrossRef]
- Nakazima, M. Studies on the source of shellfish poison in Lake Hamana. II. Shellfish toxicity during the “red-tide”. Bull. Jpn. Soc. Sci. Fish. 1965, 31, 204–207. [Google Scholar] [CrossRef]
- Nakazima, M. Studies on the source of shellfish poison in Lake Hamana. III. Poisonous effects of shellfish feeding on Prorocentrum sp. Bull. Jpn. Soc. Sci. Fish. 1965, 31, 281–285. [Google Scholar] [CrossRef]
- Nakajima, M. Studies on the source of shellfish poison in Lake Hamana. IV. Identification and collection of the noxious dinoflagellates. Bull. Jpn. Soc. Sci. Fish. 1968, 34, 130–131. [Google Scholar] [CrossRef]
- Akiba, T.; Hattori, Y. Food poisoning caused by eating asari and oyster-toxic substance, venerupin. Jpn. J. Exp. Med. 1949, 20, 271–284. [Google Scholar]
- Silva, E.S. Les “red waters” à la lagune d’ Obidos. Ses causes probables et ses rapports avec la toxicité des bivalves. Notas Est. Inst. Biol. Mar. 1963, 27, 265–275. (In French) [Google Scholar]
- Silva, E.S. As grandes populacòes de dinoflagelados tóxicos na lagoa de óbidos. Arq. Inst. Nac. Saude 1980, 4, 253–262. [Google Scholar]
- Silva, E.S.; Sousa, I. Experimental Work on the Dinoflagellate Toxin Production. Arq. Inst. Nac. Saude 1981, 6, 381–387. [Google Scholar]
- Chen, Y.Q.; Gu, X.G. An ecological study of red tides in the East China Sea. In Toxic Phytoplankton Blooms in the Sea; Smayda, T.J., Shimizu, Y., Eds.; Elsevier: Amsterdam, The Netherlands, 1993; pp. 217–221. [Google Scholar]
- Tangen, K. Brunt vann i Oslofjordi September 1979, forarsaket av den toksiske Prorocentrum minimum og andre dinoflagellater. Blyttia 1980, 38, 145–158. (In Norwegian) [Google Scholar]
- Tangen, K. Shellfish poisoning and the occurrence of potentially toxic dinoflagellates in Norwegian waters. Sarsia 1983, 68, 1–7. [Google Scholar]
- Kimor, B.; Moigis, A.G.; Dohms, V.; Stienen, C. A case of mass occurrence of Prorocentrum minimum in the Kiel Fjord. Mar. Ecol. Prog. Ser. 1985, 27, 209–215. [Google Scholar] [CrossRef]
- Moncheva, S.P. On the toxicity of Exuviaella cordata Ost. Blooming in Black Sea. Rev. Int. Océanogr. Méd. 1991, 101/104, 124–126. [Google Scholar]
- Rabbani, M.M.; Rehman, A.U.; Harms, C.E. Mass mortality of fishes caused by dinoflagellate bloom in Gwdar Bay, Southwestern Pakistan. In Toxic Marine Phytoplankton; Granéli, E., Sundstrom, B., Edler, L., Anderson, D.M., Eds.; Elsevier: New York, NY, USA, 1990; pp. 209–214. [Google Scholar]
- Tseng, C.K.; Zhou, M.J.; Zou, J.Z. Toxic phytoplankton studies in China. In Toxic Phytoplankton Blooms in the Sea; Smayda, T.J., Shimizu, Y., Eds.; Elsevier: New York, NY, USA, 1993; pp. 347–352. [Google Scholar]
- Shumway, S. A review of the effects of algal blooms on shellfish and aquaculture. J. World Aquacult. Soc. 1990, 21, 65–104. [Google Scholar] [CrossRef]
- Grzebyk, D.; Denardou, A.; Berland, B.; Pouchus, Y.F. Evidence of a new toxin in the red-tide dinoflagellate Prorocentrum minimum. J. Plankton Res. 1997, 19, 1111–1124. [Google Scholar] [CrossRef]
- Denardou-Queneherve, D.; Grzebyk, D.; Pouchus, Y.F.; Sauviat, M.P.; Alliot, E.; Biard, J.F.; Berland, B.; Verbist, J.F. Toxicity of French strains of the dinoflagellate Prorocentrum minimum experimental and natural contaminations of mussels. Toxicon 1999, 37, 1711–1719. [Google Scholar] [CrossRef] [PubMed]
- Yasumoto, T.; Oshima, Y.; Yamaguchi, M. Occurrence of a new type of shellfish poisoning in the Tohoku district. Bull. Jpn. Soc. Sci. Fish. 1978, 44, 1249–1255. [Google Scholar] [CrossRef]
- Moestrup, Ø.; Akselman, R.; Cronberg, G.; Elbraechter, M.; Fraga, S.; Halim, Y.; Hansen, G.; Hoppenrath, M.; Larsen, J.; Lundholm, N.; et al. 2014 IOC-UNESCO Taxonomic Reference List of Harmful Micro Algae. Available online: http://www.marinespecies.org/hab/aphia.php?p=taxdetails&id=232376 (accessed on 25 February 2015).
- EU Regulation 15/2011. Commission Regulation (EU) No 15/2011 of 10 January 2011 amending Regulation (EC) No 2074/2005 as regards recognized testing methods for detecting marine biotoxins in live bivalve molluscs. Off. J. Eur. Union 2011, L6, 3–6. [Google Scholar]
- Community Reference Laboratory for Marine Biotoxins. EU-HarmonisedStandard Operating Procedure for Determination of Lipophilic Toxins by Mouse Bioassay, Version 5; Community Reference Laboratory for Marine Biotoxins (CRLMB): Vigo, Spain, 2009. [Google Scholar]
- EU Decision 2002/657/EC. Commission Decision of 12 August 2002 implementing Council Directive 96/23/EC concerning the performance of analytical methods and the interpretation of results. Off. J. Eur. Commun. 2002, L221, 8–36. [Google Scholar]
- Nzoughet, J.K.; Campbell, K.; Barnes, P.; Cooper, K.M.; Chevallier, O.P.; Elliott, C.T. Comparison of sample preparation methods, validation of an UPLC–MS/MS procedure for the quantification of tetrodotoxin present in marine gastropods and analysis of pufferfish. Food Chem. 2013, 136, 1584–1589. [Google Scholar] [CrossRef] [PubMed]
- Silva, M.; Azevedo, J.; Rodriguez, P.; Alfonso, A.; Botana, L.M.; Vasconcelos, V. New gastropod vectors and tetrodotoxin potential expansion in temperate waters of the Atlantic Ocean. Mar. Drugs 2012, 10, 712–726. [Google Scholar] [CrossRef] [PubMed]
- Arakawa, O.; Hwang, D.-F.; Taniyama, S.; Takatani, T. Toxins of pufferfish that cause human intoxications. In Coastal Environmental and Ecosystem Issues of the East China Sea; Ishimatsu, A., Lie, H.-J., Eds.; Nagasaki University/Terrapub: Tokyo, Japan, 2010; pp. 227–244. [Google Scholar]
- European Food Safety Authority. Scientific Opinion: Marine Biotoxins in Shellfish—Saxitoxin group. EFSA J. 2009, 1019, 1–76. [Google Scholar]
- Ministry of Rural Development and Food. MRDF Decision 313153/20-12-2002: National Program for Monitoring of Bivalve Molluscs’ Production Areas for the Presence of Marine Biotoxins; Ministry of Rural Development and Food: Athens, Greece, 2002; p. 16. (In Greek) [Google Scholar]
- Ministry of Rural Development and Food. MRDF Circular 1558/45842/12-04-2012: Guidelines for Categorization and Monitoring of Production/Relay Areas of Live Bivalve Molluscs; Ministry of Rural Development and Food: Athens, Greece, 2012; p. 16. (In Greek) [Google Scholar]
- Utermöhl, H. Zur Vervollkommnung der quantitativen Phytoplankton-Methodik. Mitt. Int. Verein. Limnol. 1958, 9, 1–38. (In German) [Google Scholar]
- Yotsu-Yamashita, M.; Jang, J.H.; Cho, Y.; Konoki, K. Optimization for simultaneous analysis of tetrodotoxin, 4-epitetrodotoxin, 4,9-anhydrotetrodotoxin and 5,6,11-trideoxytetrodotoxin by hydrophilic interaction liquid chromatography-tandem mass spectrometry. Forensic Toxicol. 2011, 29, 61–64. [Google Scholar] [CrossRef]
- Deeds, J.R.; White, K.D.; Etheridge, S.M.; Landsberg, J.H. Concentrations of saxitoxin and tetrodotoxin in three species of puffers from the Indian river lagoon, Florida, the location for multiple cases of Saxitoxin Puffer Poisoning from 2002 to 2004. Trans. Am. Fish. Soc. 2008, 137, 1317–1326. [Google Scholar] [CrossRef]
- Huang, H.N.; Lin, J.; Lin, H.L. Identification and quantification of tetrodotoxin in the marine gastropod Nassarius by LC–MS. Toxicon 2008, 51, 774–779. [Google Scholar] [CrossRef] [PubMed]
- Jang, J.H.; Lee, J.S.; Yotsu-Yamashita, M. LC/MS analysis of tetrodotoxin and its deoxy analogs in the marine Puffer Fish Fugu niphobles from the Southern Coast of Korea, and in the Brackish water Puffer Fishes Tetraodon nigroviridis and Tetraodon biocellatus from Southeast Asia. Mar. Drugs 2010, 8, 1049–1058. [Google Scholar] [CrossRef] [PubMed]
- EU Regulation 853/2004. Regulation (EC) No 853/2004 of the European Parliament and of the Council of 29 April 2004 laying down specific hygiene rules for food of animal origin. Off. J. Eur. Union 2004, 226, 22–82. [Google Scholar]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vlamis, A.; Katikou, P.; Rodriguez, I.; Rey, V.; Alfonso, A.; Papazachariou, A.; Zacharaki, T.; Botana, A.M.; Botana, L.M. First Detection of Tetrodotoxin in Greek Shellfish by UPLC-MS/MS Potentially Linked to the Presence of the Dinoflagellate Prorocentrum minimum. Toxins 2015, 7, 1779-1807. https://doi.org/10.3390/toxins7051779
Vlamis A, Katikou P, Rodriguez I, Rey V, Alfonso A, Papazachariou A, Zacharaki T, Botana AM, Botana LM. First Detection of Tetrodotoxin in Greek Shellfish by UPLC-MS/MS Potentially Linked to the Presence of the Dinoflagellate Prorocentrum minimum. Toxins. 2015; 7(5):1779-1807. https://doi.org/10.3390/toxins7051779
Chicago/Turabian StyleVlamis, Aristidis, Panagiota Katikou, Ines Rodriguez, Verónica Rey, Amparo Alfonso, Angelos Papazachariou, Thetis Zacharaki, Ana M. Botana, and Luis M. Botana. 2015. "First Detection of Tetrodotoxin in Greek Shellfish by UPLC-MS/MS Potentially Linked to the Presence of the Dinoflagellate Prorocentrum minimum" Toxins 7, no. 5: 1779-1807. https://doi.org/10.3390/toxins7051779
APA StyleVlamis, A., Katikou, P., Rodriguez, I., Rey, V., Alfonso, A., Papazachariou, A., Zacharaki, T., Botana, A. M., & Botana, L. M. (2015). First Detection of Tetrodotoxin in Greek Shellfish by UPLC-MS/MS Potentially Linked to the Presence of the Dinoflagellate Prorocentrum minimum. Toxins, 7(5), 1779-1807. https://doi.org/10.3390/toxins7051779