
No Evidence for a Culturable Bacterial Tetrodotoxin Producer in Pleurobranchaea maculata (Gastropoda: Pleurobranchidae) and Stylochoplana sp. (Platyhelminthes: Polycladida)

Abstract
:1. Introduction

{kind=link}
{kind=link}
| Ref | Source | Toxicity of Host Species/Tissue * | Bacteria | Toxicity (TTX or Related Substances) ** | Detection Method * |
|---|---|---|---|---|---|
| [10] | Takifugu niphobles (pufferfish) | intestines: N/A | Raoultella terrigena | 4.3 μg·L−1 | ELISA |
| [22] | Fugu obscurus (pufferfish) | liver: 80 MU·g−1 | Lysinibacillus fusiformis | 23.9 MU in 200 mL broth | mouse bioassay |
| [14] | Fugu obscurus (pufferfish) | ovary: 125 MU·g−1 | Bacillus sp. | + | HPLC, EMI-MS |
| [16] | Takifugu obscurus (pufferfish) | ovary: N/A | Aeromonas sp. | 1.88 μg·L−1 cultured bacteria | ELISA |
| [21] | Arothron hispidus (pufferfish) | 1 μg·g−1 | Vibrio harveyi | 0.05–1.57 μg·mL−1 | LC-MS |
| [15] | Nassarius semiplicatus (sea snail) | 2 × 102 MU·g−1 tissue (3.6 mg in 100 g tissue) | Vibrio spp. | 11–184 ng·g−1 | competitive ELISA |
| Marinomonas spp. | 85–98 ng·g−1 | competitive ELISA | |||
| Tenacibaculum spp. | 54 ng·g−1 | competitive ELISA | |||
| [8] | Pseudocaligus fugu (copepod) | N/A | Roseobacter sp. | + | HPLC, GC-MS, LC-MS |
| [23] | Chelonodon patoca (pufferfish) | skin: N/A | Serratia marcescens | + | HPLC |
| [18,19] | Fugu rubripes (pufferfish) | ovary: 120 ± 6.2 MU·g−1 | Bacillus spp. | 0.1–1.6 MU·g−1 cells | mouse bioassay |
| Nocardiopsis dassonvillei | 0.5 MU·g−1 cells | mouse bioassay | |||
| Actinomycete spp. | 0.1–1.6 MU·g−1 cells | mouse bioassay | |||
| [17] | Takifugu alboplumbeus (pufferfish) | intestines: 24.9 ± 24.2 MU·g−1 [24] | Vibrio spp. | 78.3 MU in 500 mL broth (4 × 107 cells) | mouse bioassay |
| Takifugu niphobles (pufferfish) | ovary—100–1000 MU·g−1 [24] | Microbacterium arabinogalactanolyticum | 105.3 MU in 500 mL broth (4 × 107 cells) | mouse bioassay | |
| [25] | Seven species of nemertean worms | N/A | Vibrio spp. | + | HPLC |
| [13] | Fugu vermicularis radialis (pufferfish) | 70 ± 8 MU·g−1 | Vibrio spp. | + | HPLC |
| [26] | Meoma ventricosa (sea urchin) | N/A | Pseudoalteromonas spp. | + | immunoassay |
| [11] | Niotha clathrata (marine gastropod) | 2–50 MU·g−1 | Vibrio spp. | + | HPLC |
| Pseudomonas spp. | + | HPLC | |||
| Aeromonas spp. | + | HPLC | |||
| Plesiomonas spp. | + | HPLC | |||
| [27] | Freshwater sediment | +HPLC, GC-MS | Micrococcus spp. | + | HPLC |
| Bacillus spp. | + | HPLC | |||
| Caulobacter spp. | + | HPLC | |||
| Flavobacterium spp. | + | HPLC | |||
| [28] | Marine sediment | +HPLC, GC-MS | Streptomyces spp. | + | HPLC |
| [29] | Deep sea sediment | 25–90 ng TTX equivalents g−1 of mud [30] | Vibrio spp. | + | HPLC |
| Bacillus spp. | + | HPLC | |||
| Acinetobacter spp. | + | HPLC | |||
| Alteromonas spp. | + | HPLC | |||
| Aeromonas spp. | + | HPLC | |||
| Micrococcus spp. | + | HPLC | |||
| [31] | Four species of Chaetognaths (arrowworms) | 320 pg individual−1 [32] | Vibrio spp. | 280–790 pg·μL−1 culture medium | cell culture bioassay |
| [12] | Hapalochlaena maculosa (blue-ringed octopus) | 140–174 MU idividual−1 | Vibrio spp. | + | HPLC, GC-MS |
| Pseudomonas spp. | 3 MU, + | mouse bioassay, HPLC, GC-MS | |||
| Bacillus spp. | 5 MU, + | mouse bioassay, HPLC, GC-MS | |||
| Alteromonas spp. | + | HPLC, GC-MS | |||
| [33] | Takifugu niphobles (pufferfish) | intestine 3890 MU·g−1 | Shewanella putrefaciens | 15 MU in 250 mL culture broth, + | mouse bioassay, HPLC, GC-MS |
| [34] | Fugu vermicularis vermicularis (pufferfish) | 178 MU·g−1 | Vibrio spp. | 3 MU, +, + | mouse bioassay, HPLC, GC-MS |
| [35] | Astropecten polyacanthus (comb seastar) | 32 MU·g−1 | Vibrio spp. | + | HPLC, GC-MS |
| [36] | Fugu poecilonotus (pufferfish) | N/A | Pseudomonas spp. | + | HPLC, GC-MS |
| [34] | Atergatis floridus (reef crab) | + TLC, eletrophoresis | Vibrio spp. | + | HPLC, GC-MS |
| [7] | Jania spp. (red alga) | N/A | Pseudomonas spp. | + | HPLC, GC-MS |
2. Results and Discussion
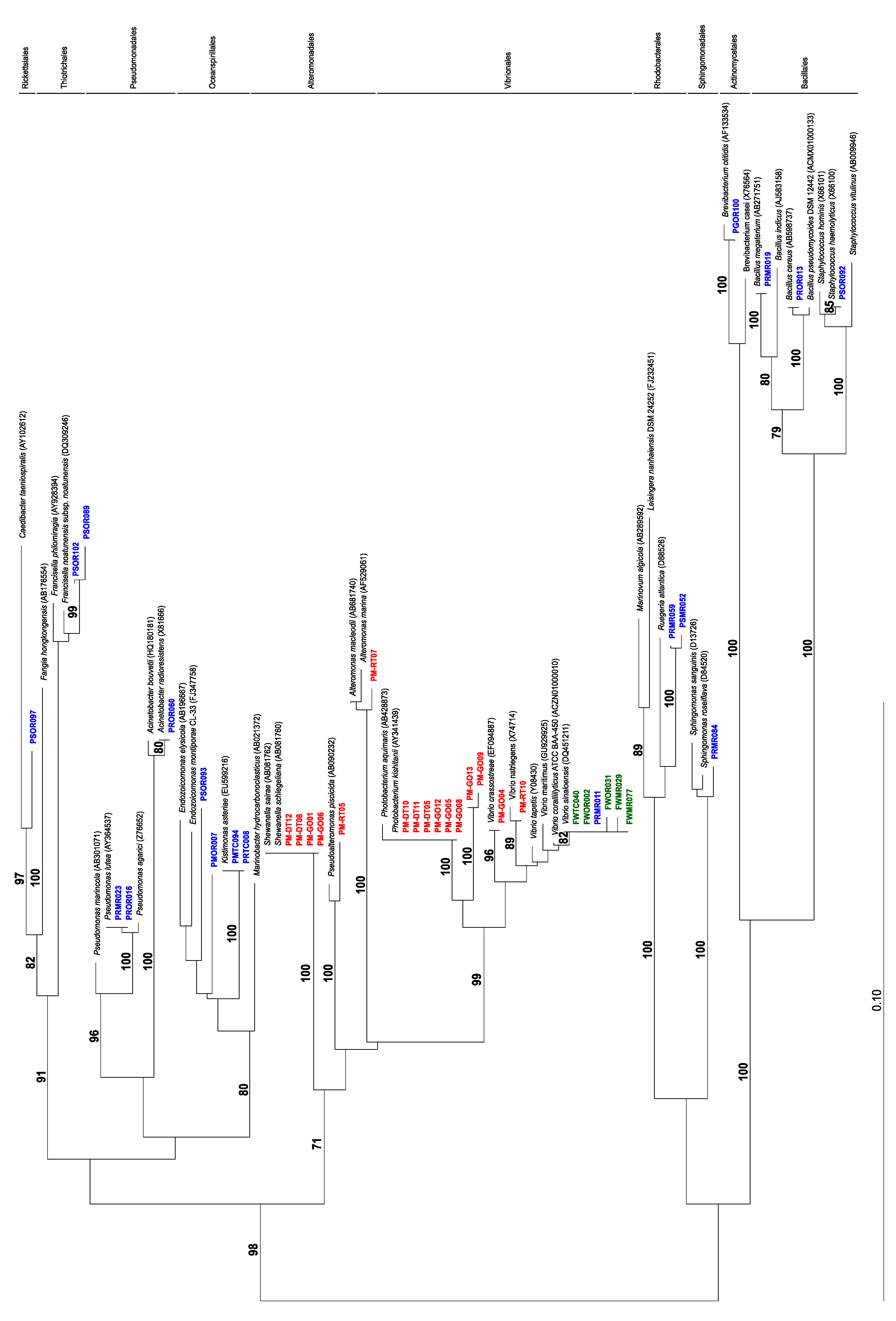
2.1. Bacterial Isolation and Toxin Analysis
| Sample | Location | TTX (mg·kg−1) | Bacterial Strains |
|---|---|---|---|
| P. maculata; digestive | M.I. | 2 | 16 |
| P. maculata; gonad | M.I. | 5 | 3 |
| P. maculata; mantle | M.I. | 7 | 21 |
| P. maculata;“rest” | M.I. | 8 | 23 |
| Stylochoplana sp. | M.I., P.B. | 174 | 39 |
| P. maculata; digestive | I.R. | 771 | - |
| P. maculata; gonad | I.R. | 136 | - |
| P. maculata; mantle | I.R. | 97 | - |
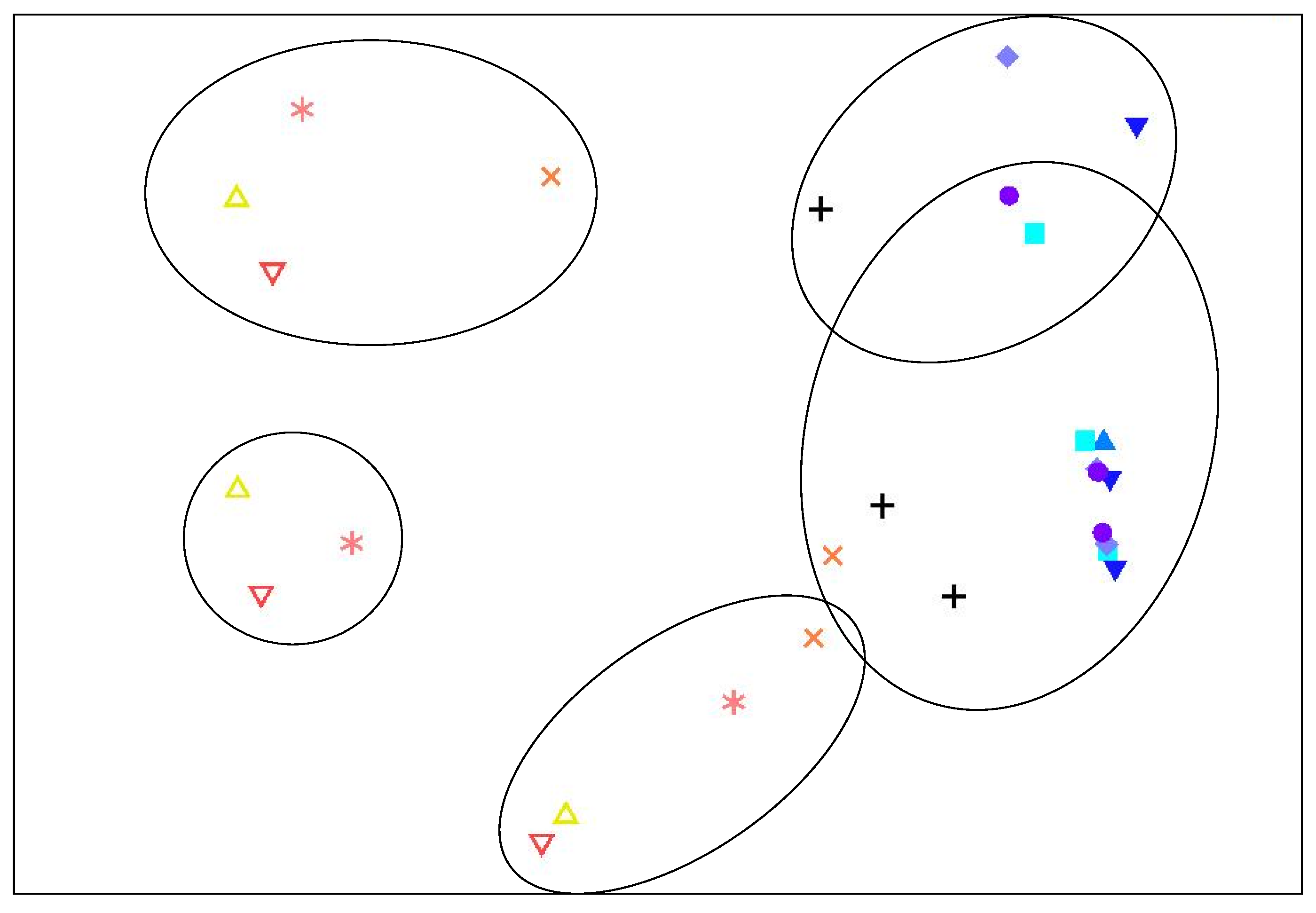
2.2. Bacterial Community Analysis
 0 days,
0 days,  day 1,
day 1,  day 2,
day 2,  day 3,
day 3,  day 4, + day 6,
day 4, + day 6,  day 8,
day 8,  day 10,
day 10,  day 12,
day 12,  day 14. Points enclosed by solid line cluster at 40% similarity.
0 days, day 1, day 2, day 3, day 4, + day 6, day 8, day 10, day 12, day 14. Points enclosed by solid line cluster at 40% similarity.
day 14. Points enclosed by solid line cluster at 40% similarity.
0 days, day 1, day 2, day 3, day 4, + day 6, day 8, day 10, day 12, day 14. Points enclosed by solid line cluster at 40% similarity.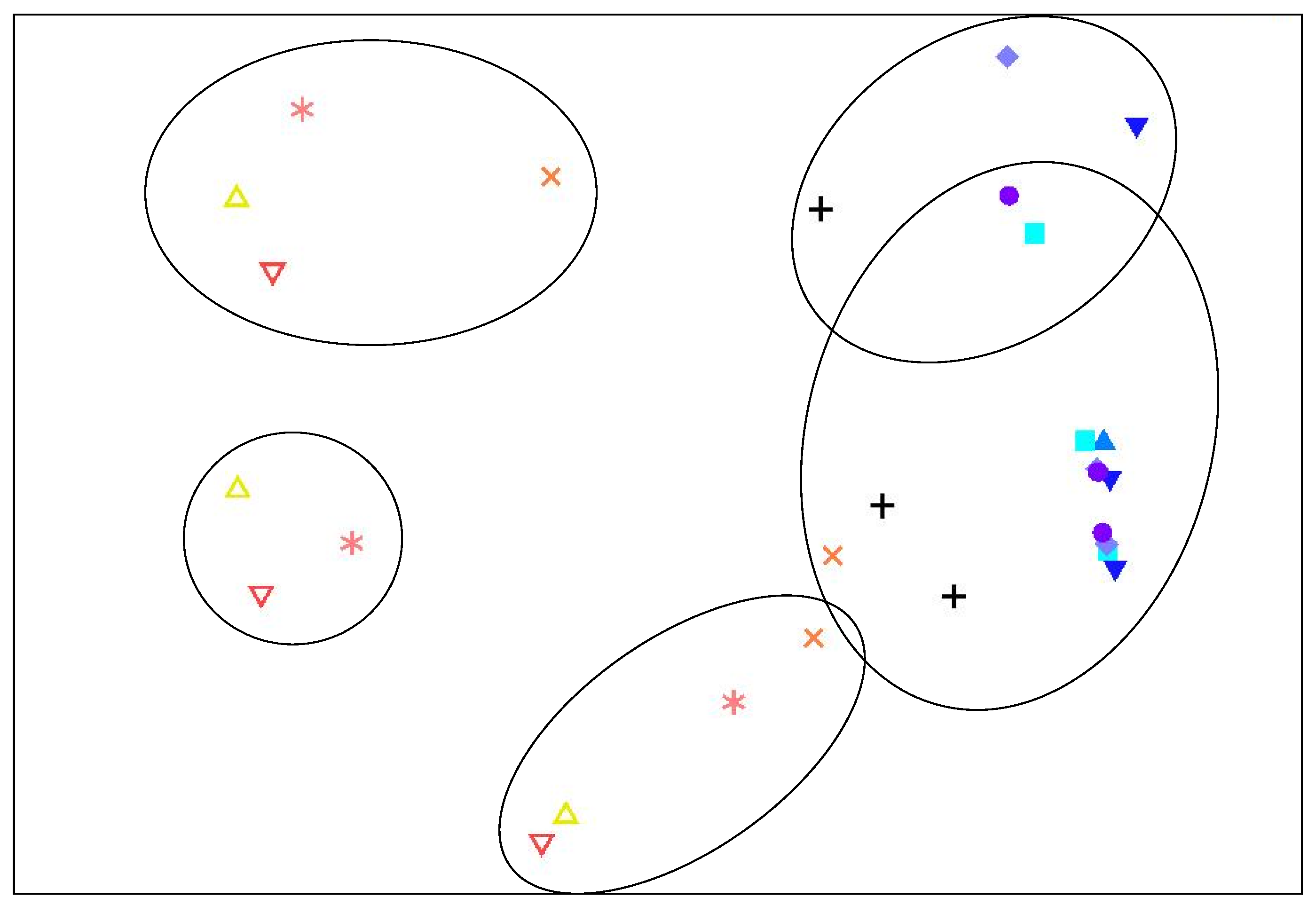
3. Experimental Section
3.1. Bacterial Strain Isolation
3.1.1. Collection and Strain Isolation
3.1.2. Molecular and Phylogenetic Analysis of Bacterial Strains
3.1.3. Bacterial Culturing
3.2. Bacterial Community Broth Experiments
3.2.1. Collection and Inoculation
3.2.2. Molecular Analysis
3.3. Tetrodotoxin and C9 Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Halstead, B.W. The microbial biogenesis of aquatic biotoxins. Toxicol. Mech. Methods 2002, 12, 135–153. [Google Scholar] [CrossRef] [PubMed]
- Mosher, H.S. The chemistry of tetrodotoxin. Ann. NY Acad. Sci. 1986, 479, 32–43. [Google Scholar] [CrossRef] [PubMed]
- Hwang, D.-F.; Noguchi, T. Tetrodotoxin poisoning. Adv. Food Nutr. Res. 2007, 52, 141–236. [Google Scholar] [PubMed]
- Noguchi, T.; Ebesu, J.S.M. Puffer poisoning: epidemiology and treatment. Toxin Rev. 2001, 20, 1–10. [Google Scholar] [CrossRef]
- Chau, R.; Kalaitzis, J.A.; Neilan, B.A. On the origins and biosynthesis of tetrodotoxin. Aquat Toxicol. 2011, 104, 61–72. [Google Scholar] [CrossRef] [PubMed]
- Zimmer, T. Effects of tetrodotoxin on the mammalian cardiovascular system. Mar. Drugs 2010, 8, 741–762. [Google Scholar] [CrossRef] [PubMed]
- Yasumoto, T.; Yasumura, D.; Yotsu, M.; Michishita, T.; Endo, A.; Kotaki, Y. Bacterial production of tetrodotoxin and anhydrotetrodotoxin. Agric. Biol. Chem. 1986, 50, 793–795. [Google Scholar] [CrossRef]
- Maran, B.A.V.; Iwamoto, E.; Okuda, J.; Matsuda, S.; Taniyama, S.; Shida, Y.; Asakawa, M.; Ohtsuka, S.; Nakai, T.; Boxshall, G.A. Isolation and characterization of bacteria from the copepod Pseudocaligus fugu ectoparasitic on the panther puffer Takifugu pardalis with the emphasis on TTX. Toxicon 2007, 50, 779–790. [Google Scholar] [CrossRef] [PubMed]
- Pratheepa, V.; Vasconcelos, V. Microbial diversity associated with tetrodotoxin production in marine organisms. Environ. Toxicol. Pharmacol. 2013, 36, 1046–1054. [Google Scholar] [CrossRef] [PubMed]
- Yu, V.C.-H.; Yu, P.H.-F.; Ho, K.-C.; Lee, F.W.-F. Isolation and identification of a new tetrodotoxin-producing bacterial species, Raoultella terrigena, from Hong Kong marine puffer fish Takifugu niphobles. Mar. Drugs 2011, 9, 2384–2396. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.A.; Hwang, D.F.; Tsai, Y.H.; Chen, H.C.; Jeng, S.S.; Noguchi, T.; Ohwada, K.; Hashimoto, K. Microflora and tetrodotoxin-producing bacteria in a gastropod, Niotha clathrata. Food Chem. Toxicol. 1995, 33, 929–934. [Google Scholar] [CrossRef] [PubMed]
- Hwang, D.F.; Arakawa, O.; Saito, T.; Noguchi, T.; Simidu, U.; Tsukamoto, K.; Shida, Y.; Hashimoto, K. Tetrodotoxin-producing bacteria from the blue-ringed octopus Octopus maculosus. Mar. Biol. 1989, 100, 327–332. [Google Scholar] [CrossRef]
- Lee, M.-J.; Jeong, D.-Y.; Kim, W.-S.; Kim, H.-D.; Kim, C.-H.; Park, W.-W.; Park, Y.-H.; Kim, K.-S.; Kim, H.-M.; Kim, D.-S. A tetrodotoxin-producing Vibrio strain, LM-1, from the puffer fish Fugu vermicularis radiatus. Appl. Environ. Microbiol. 2000, 66, 1698–1701. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Fan, Y.H. Isolation and characterization of a Bacillus species capable of producing tetrodotoxin from the puffer fish Fugu obscurus. World J. Microbiol. Biotechnol. 2010, 26, 1755–1760. [Google Scholar] [CrossRef]
- Wang, X.-J.; Yu, R.-C.; Luo, X.; Zhou, M.-J.; Lin, X.-T. Toxin-screening and identification of bacteria isolated from highly toxic marine gastropod Nassarius semiplicatus. Toxicon 2008, 52, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Xu, J.; Liang, S.; Ren, D.; Yan, X.; Bao, B. A novel TTX-producing Aeromonas isolated from the ovary of Takifugu obscurus. Toxicon 2010, 56, 324–329. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.F.; Yu, P.H.F.; Chan, P.L.; Yan, Q.; Wong, P.K. Two novel species of tetrodotoxin-producing bacteria isolated from toxic marine puffer fishes. Toxicon 2004, 44, 641–647. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.L.; Xie, L.P.; Xia, G.L.; Zhang, J.F.; Nie, Y.C.; Hu, J.C.; Wang, S.J.; Zhang, R.Q. A new tetrodotoxin-producing actinomycete, Nocardiopsis dassonvillei, isolated from the ovaries of puffer fish Fugu rubripes. Toxicon 2005, 45, 851–859. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.L.; Yang, Y.; Xie, L.P.; Xia, G.L.; Hu, J.C.; Wang, S.J.; Zhang, R.Q. Toxicity and distribution of tetrodotoxin-producing bacteria in puffer fish Fugu rubripes collected from the Bohai Sea of China. Toxicon 2005, 46, 471–476. [Google Scholar] [CrossRef] [PubMed]
- Matsumura, K. Reexamination of tetrodotoxin production by bacteria. Appl. Environ. Microbiol. 1995, 61, 3468–3470. [Google Scholar] [PubMed]
- Campbell, S.; Harada, R.M.; DeFelice, S.V.; Bienfang, P.K.; Li, Q.X. Bacterial production of tetrodotoxin in the pufferfish Arothron hispidus. Nat. Prod. Res. 2009, 23, 1630–1640. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Fan, Y.; Yao, Z. Isolation of a Lysinibacillus fusiformis strain with tetrodotoxin-producing ability from puffer fish Fugu obscurus and the characterization of this strain. Toxicon 2010, 56, 640–643. [Google Scholar] [CrossRef] [PubMed]
- Yan, Q.; Yu, P.H.F.; Li, H.Z. Detection of tetrodotoxin and bacterial production by Serratia marcescens. World J. Microbiol. Biotechnol. 2005, 21, 1255–1258. [Google Scholar] [CrossRef]
- Yu, C.-F.; Yu, P.H.-F. The annual toxicological profiles of two common puffer fish, Takifugu niphobles (Jordan and Snyder) and Takifugu alboplumbeus (Richardson), collected along Hong Kong coastal waters. Toxicon 2002, 40, 313–316. [Google Scholar] [CrossRef] [PubMed]
- Carroll, S.; McEvoy, E.G.; Gibson, R. The production of tetrodotoxin-like substances by nemertean worms in conjunction with bacteria. J. Exp. Mar. Biol. Ecol. 2003, 288, 51–63. [Google Scholar] [CrossRef]
- Ritchie, K.B.; Nagelkerken, I.; James, S.; Smith, G.W. Environmental microbiology—A tetrodotoxin-producing marine pathogen. Nature 2000, 404. [Google Scholar] [CrossRef]
- Do, H.K.; Hamasaki, K.; Ohwada, K.; Simidu, U.; Noguchi, T.; Shida, Y.; Kogure, K. Presence of tetrodotoxin and tetrodotoxin-producing bacteria in fresh-water sediments. Appl. Environ. Microbiol. 1993, 59, 3934–3937. [Google Scholar] [PubMed]
- Do, H.K.; Kogure, K.; Imada, C.; Noguchi, T.; Ohwada, K.; Simidu, U. Tetrodotoxin production of actinomycetes isolated from marine sediment. J. Appl. Microbiol. 1991, 70, 464–468. [Google Scholar]
- Do, H.K.; Kogure, K.; Simidu, U. Identification of deep-sea-sediment bacteria which produce tetrodotoxin. Appl. Environ. Microbiol. 1990, 56, 1162–1163. [Google Scholar] [PubMed]
- Kogure, K.; Do, H.K.; Thuesen, E.V.; Nanba, K.; Ohwada, K.; Simidu, U. Accumulation of tetrodotoxin in marine sediment. Mar. Ecol. Prog. Ser. 1988, 45, 303–305. [Google Scholar] [CrossRef]
- Thuesen, E.V.; Kogure, K. Bacterial production of tetrodotoxin in four species of Chaetognatha. Biol. Bull. 1989, 176, 191–194. [Google Scholar] [CrossRef]
- Thuesen, E.V.; Kogure, K.; Hashimoto, K.; Nemoto, T. Poison arrowworms: A tetrodotoxin venom in the marine phylum Chaetognatha. J. Exp. Mar. Biol. Ecol. 1988, 116, 249–256. [Google Scholar] [CrossRef]
- Matsui, T.; Taketsugu, S.; Kodama, K.; Ishii, A.; Yamamori, K.; Shimizu, C. Production of tetrodotoxin by the intestinal bacteria of a puffer fish Takifugu-niphobles. Nippon Suisan Gakkaishi 1989, 55, 2199–2203. [Google Scholar] [CrossRef]
- Noguchi, T.; Hwang, D.F.; Arakawa, O.; Sugita, H.; Deguchi, Y.; Shida, Y.; Hashimoto, K. Vibrio alginolyticus, a tetrodotoxin-producing bacterium, in the intestines of the fish Fugu vermicularis vermicularis. Mar. Biol. 1987, 94, 625–630. [Google Scholar] [CrossRef]
- Narita, H.; Matsubara, S.; Miwa, N.; Akahane, S.; Murakami, M.; Goto, T.; Nara, M.; Noguchi, T.; Saito, T.; Shida, Y.; et al. Vibrio alginolyticus, a TTX-producing Bacterium Isolated from the Starfish Astropecten polyacanthus. Bull Jpn. Soc. Sci. Fish 1987, 53, 617–621. [Google Scholar]
- Yotsu, M.; Yamazaki, T.; Meguro, Y.; Endo, A.; Murata, M.; Naoki, H.; Yasumoto, T. Production of tetrodotoxin and its derivatives by Pseudomonas sp. isolated from the skin of a pufferfish. Toxicon 1987, 25, 225–228. [Google Scholar]
- Lehman, E.M.; Brodie, E.D., Jr.; Brodie, E.D., III. No evidence for an endosymbiotic bacterial origin of tetrodotoxin in the newt Taricha granulosa. Toxicon 2004, 44, 243–249. [Google Scholar] [CrossRef] [PubMed]
- Cardall, B.L.; Brodie, E.D.; Hanifin, C.T. Secretion and regeneration of tetrodotoxin in the rough-skin newt (Taricha granulosa). Toxicon 2004, 44, 933–938. [Google Scholar] [CrossRef] [PubMed]
- McNabb, P.; Selwood, A.I.; Munday, R.; Wood, S.A.; Taylor, D.I.; MacKenzie, L.A.; van Ginkel, R.; Rhodes, L.L.; Cornelisen, C.; Heasman, K.; et al. Detection of tetrodotoxin from the grey side-gilled sea slug—Pleurobranchaea maculata, and associated dog neurotoxicosis on beaches adjacent to the Hauraki Gulf, Auckland, New Zealand. Toxicon 2010, 56, 466–473. [Google Scholar]
- Willan, R.C. New Zealand side-gilled sea slugs (Opisthobranchia: Notaspidea: Pluerobranchidae). Malacologia 1983, 23, 221–270. [Google Scholar]
- Wood, S.A.; Taylor, D.I.; McNabb, P.; Walker, J.; Adamson, J.; Cary, S.C. Tetrodotoxin concentrations in Pleurobranchaea maculata: temporal, spatial and individual variability from New Zealand populations. Mar. Drugs 2012, 10, 163–176. [Google Scholar] [CrossRef] [PubMed]
- Salvitti, L.R.; Wood, S.A.; Taylor, D.I.; McNabb, P.; Cary, S.C. First identification of tetrodotoxin (TTX) in the flatworm Stylochoplana sp.; a source of TTX for the sea slug Pleurobranchaea maculata. Toxicon 2015, 95, 23–29. [Google Scholar]
- Khor, S.; Wood, S.A.; Salvitti, L.; Taylor, D.I.; Adamson, J.; McNabb, P.; Cary, S.C. Investigating diet as the source of tetrodotoxin in Pleurobranchaea maculata. Mar. Drugs 2013, 12, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Chau, R.; Kalaitzis, J.A.; Wood, S.A.; Neilan, B.A. Diversity and biosynthetic potential of culturable microbes associated with toxic marine animals. Mar. Drugs 2013, 11, 2695–2712. [Google Scholar] [CrossRef] [PubMed]
- McEvoy, E.G.; Rogers, A.; Gibson, R. Preliminary investigation of Vibrio alginolyticus-like bacteria associated with marine nemerteans. Hydrobiologia 1997, 365, 287–291. [Google Scholar] [CrossRef]
- McNabb, P.S.; Taylor, D.I.; Ogilvie, S.C.; Wilkinson, L.; Anderson, A.; Hamon, D.; Wood, S.A.; Peake, B.M. First detection of tetrodotoxin in the bivalve Paphies australis by liquid chromatography coupled to triple quadrupole mass spectrometry with and without precolumn reaction. J. AOAC Int. 2014, 97, 325–333. [Google Scholar] [PubMed]
- Kudo, Y.; Yamashita, Y.; Mebs, D.; Cho, Y.; Konoki, K.; Yasumoto, T.; Yotsu-Yamashita, M. C5-C10 directly bonded tetrodotoxin analogues: Possible biosynthetic precursors of tetrodotoxin from newts. Angew. Chem. Int. Ed. Engl. 2014, 53, 14546–14549. [Google Scholar] [CrossRef] [PubMed]
- Auawithoothij, W.; Noomhorm, A. Shewanella putrefaciens, a major microbial species related to tetrodotoxin (TTX)-accumulation of puffer fish Lagocephalus lunaris. J. Appl. Microbiol. 2012, 113, 459–465. [Google Scholar] [CrossRef] [PubMed]
- Proksch, P.; Edrada, R.A.; Ebel, R. Drugs from the seas—Current status and microbiological implications. Appl. Microbiol. Biotechnol. 2002, 59, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Noguchi, T.; Jeon, J.-K.; Arakawa, O.; Sugita, H.; Deguchi, Y.; Shida, Y.; Hashimoto, K. Occurrence of tetrodotoxin and anhydrotetrodotoxin in Vibrio sp. isolated from the intestines of a xanthid crab, Atergatis floridus. J. Biochem. 1986, 99, 311–314. [Google Scholar]
- Beleneva, I.A.; Magarlamov, T.Y.; Kukhlevsky, A.D. Characterization, identification, and screening for tetrodotoxin production by bacteria associated with the ribbon worm (Nemertea) Cephalotrix simula (Ivata, 1952). Microbiology 2014, 83, 220–226. [Google Scholar] [CrossRef]
- Hugenholtz, P.; Goebel, B.M.; Pace, N.R. Impact of culture-independent studies on the emerging phylogenetic view of bacterial diversity. J. Bacteriol. 1998, 180, 4765–4774. [Google Scholar] [PubMed]
- Simidu, U.; Tsukamoto, K. Habitat segregation and biochemical activities of marine members of the family Vibrionaceae. Appl. Environ. Microbiol. 1985, 50, 781–790. [Google Scholar] [PubMed]
- DeSantis, T.Z.; Hugenholtz, P.; Larsen, N.; Rojas, M.; Brodie, E.L.; Keller, K.; Huber, T.; Dalevi, D.; Hu, P.; Andersen, G.L. Greengenes, a chimera-checked 16S rRNA gene database and workbench compatible with ARB. Appl. Environ. Microbiol. 2006, 72, 5069–5072. [Google Scholar] [CrossRef] [PubMed]
- Ludwig, W.; Strunk, O.; Westram, R.; Richter, L.; Meier, H.; Buchner, A.; Lai, T.; Steppi, S.; Jobb, G.; Förster, W. ARB: A software environment for sequence data. Nucleic Acids Res. 2004, 32, 1363–1371. [Google Scholar] [CrossRef] [PubMed]
- Felsenstein, J. PHYLIP—Phylogeny Inference Package. Cladistics 1989, 5, 164–166. [Google Scholar]
- Jukes, T.H.; Cantor, C.R. Evolution of protein molecules. In Mammalian protein metabolism II; Munro, H.N., Ed.; Academic Press: New York, NY, USA, 1969; pp. 21–132. [Google Scholar]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [PubMed]
- Barrett, J.E.; Virginia, R.A.; Wall, D.H.; Cary, S.C.; Adams, B.J.; Hacker, A.L.; Aislabie, J.M. Co-variation in soil biodiversity and biogeochemistry in northern and southern Victoria Land, Antarctica. Antarct. Sci. 2006, 18, 535–548. [Google Scholar] [CrossRef]
- Cardinale, M.; Brusetti, L.; Quatrini, P.; Borin, S.; Puglia, A.M.; Rizzi, A.; Zanardini, E.; Sorlini, C.; Corselli, C.; Daffonchio, D. Comparison of different primer sets for use in automated ribosomal intergenic spacer analysis of complex bacterial communities. Appl. Environ. Microbiol. 2004, 70, 6147–6156. [Google Scholar] [CrossRef] [PubMed]
- Abdo, Z.; Schuette, U.M.E.; Bent, S.J.; Williams, C.J.; Forney, L.J.; Joyce, P. Statistical methods for characterizing diversity of microbial communities by analysis of terminal restriction fragment length polymorphisms of 16S rRNA genes. Environ. Microbiol. 2006, 8, 929–938. [Google Scholar] [CrossRef] [PubMed]
- R: A Language and Environment for Statistical Computing. Available online: http://www.R-project.org/ (accessed on 26 January 2015).
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salvitti, L.R.; Wood, S.A.; McNabb, P.; Cary, S.C. No Evidence for a Culturable Bacterial Tetrodotoxin Producer in Pleurobranchaea maculata (Gastropoda: Pleurobranchidae) and Stylochoplana sp. (Platyhelminthes: Polycladida). Toxins 2015, 7, 255-273. https://doi.org/10.3390/toxins7020255
Salvitti LR, Wood SA, McNabb P, Cary SC. No Evidence for a Culturable Bacterial Tetrodotoxin Producer in Pleurobranchaea maculata (Gastropoda: Pleurobranchidae) and Stylochoplana sp. (Platyhelminthes: Polycladida). Toxins. 2015; 7(2):255-273. https://doi.org/10.3390/toxins7020255
Chicago/Turabian StyleSalvitti, Lauren R., Susanna A. Wood, Paul McNabb, and Stephen Craig Cary. 2015. "No Evidence for a Culturable Bacterial Tetrodotoxin Producer in Pleurobranchaea maculata (Gastropoda: Pleurobranchidae) and Stylochoplana sp. (Platyhelminthes: Polycladida)" Toxins 7, no. 2: 255-273. https://doi.org/10.3390/toxins7020255
APA StyleSalvitti, L. R., Wood, S. A., McNabb, P., & Cary, S. C. (2015). No Evidence for a Culturable Bacterial Tetrodotoxin Producer in Pleurobranchaea maculata (Gastropoda: Pleurobranchidae) and Stylochoplana sp. (Platyhelminthes: Polycladida). Toxins, 7(2), 255-273. https://doi.org/10.3390/toxins7020255