
Molecular Mechanisms of Lipoic Acid Protection against Aflatoxin B1-Induced Liver Oxidative Damage and Inflammatory Responses in Broilers




Abstract
:1. Introduction
2. Results
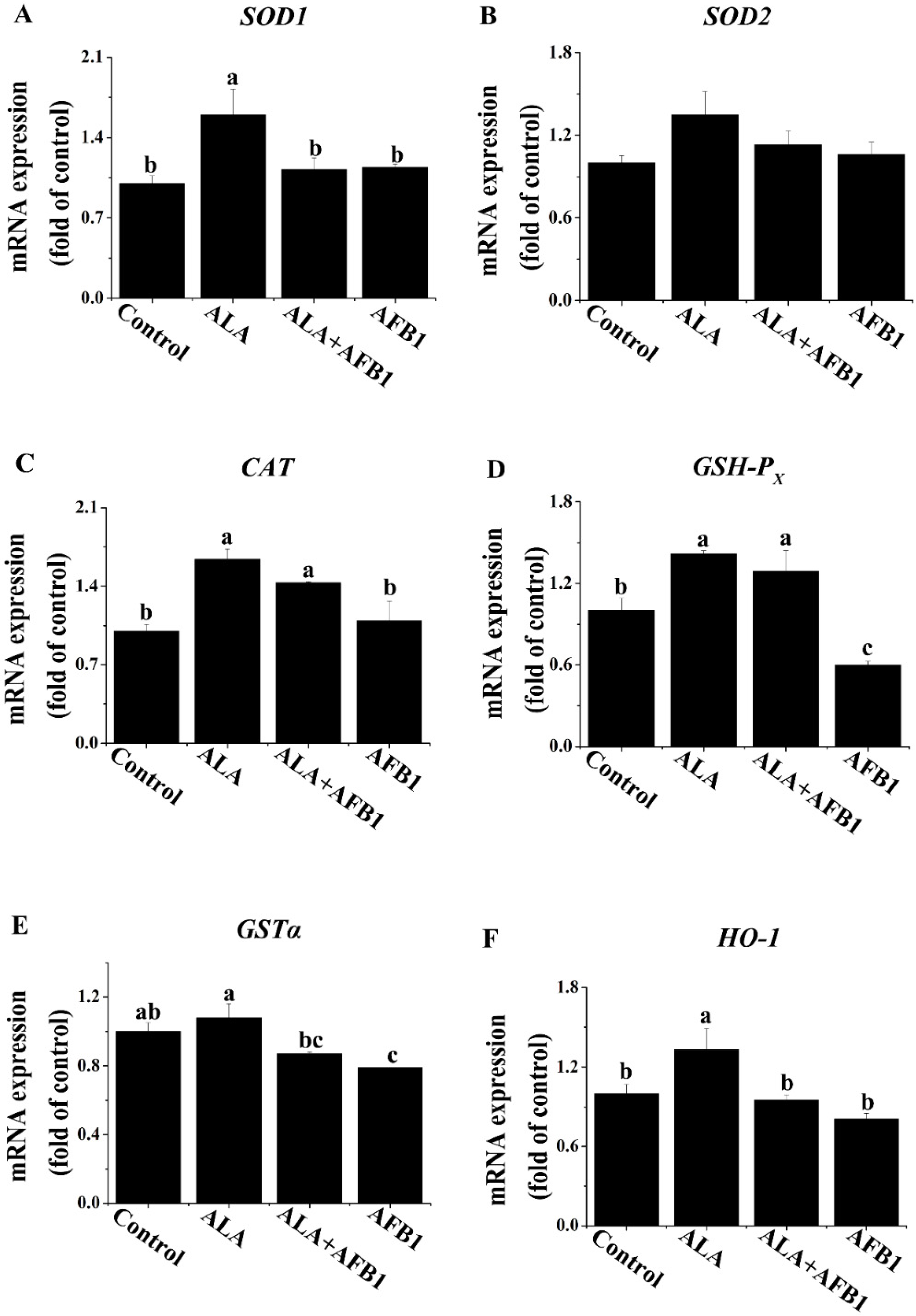
2.1. Effects of α-LA on the mRNA Levels of Antioxidant Genes in Livers of Chickens Exposed to AFB1
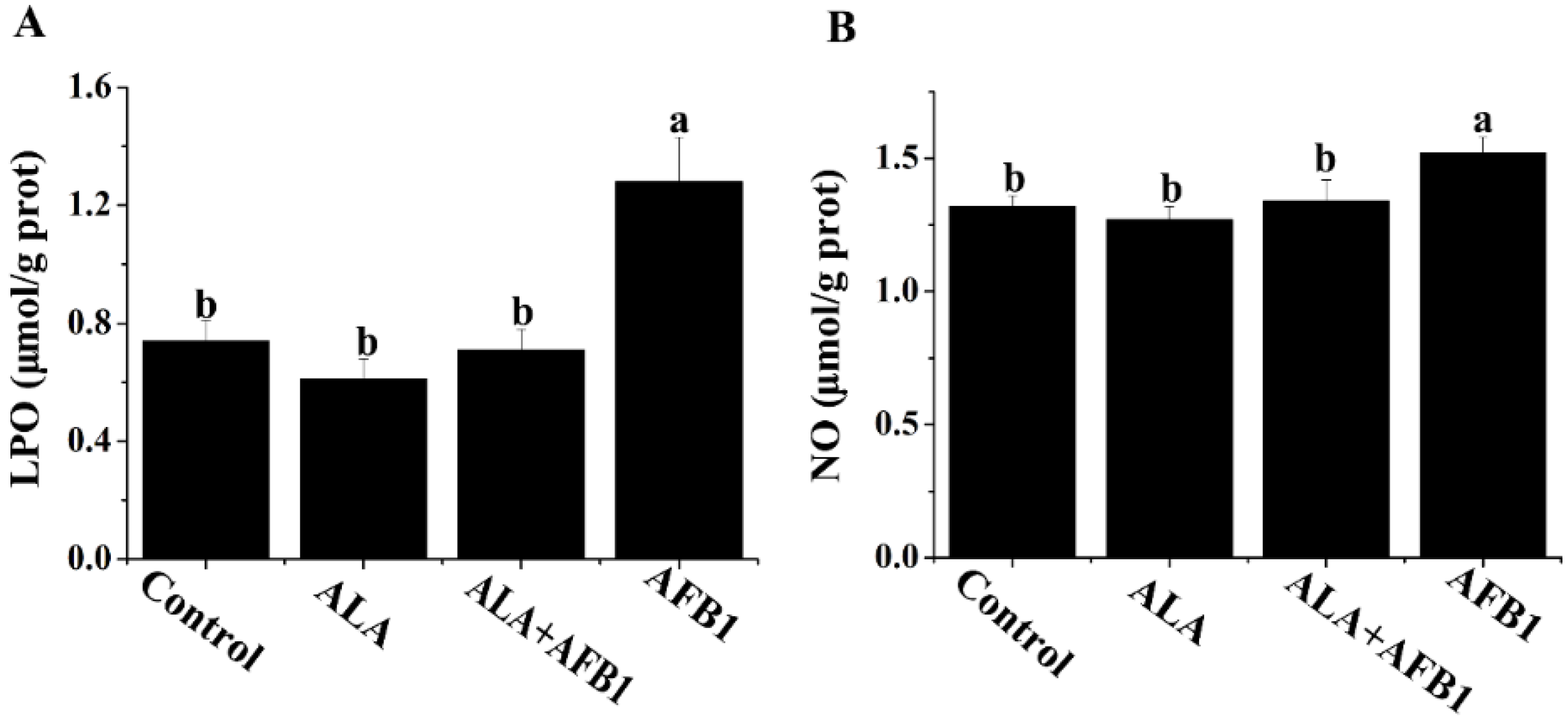
2.2. Effects of α-LA on the Levels of LPO and NO in Livers of Chickens Exposed to AFB1
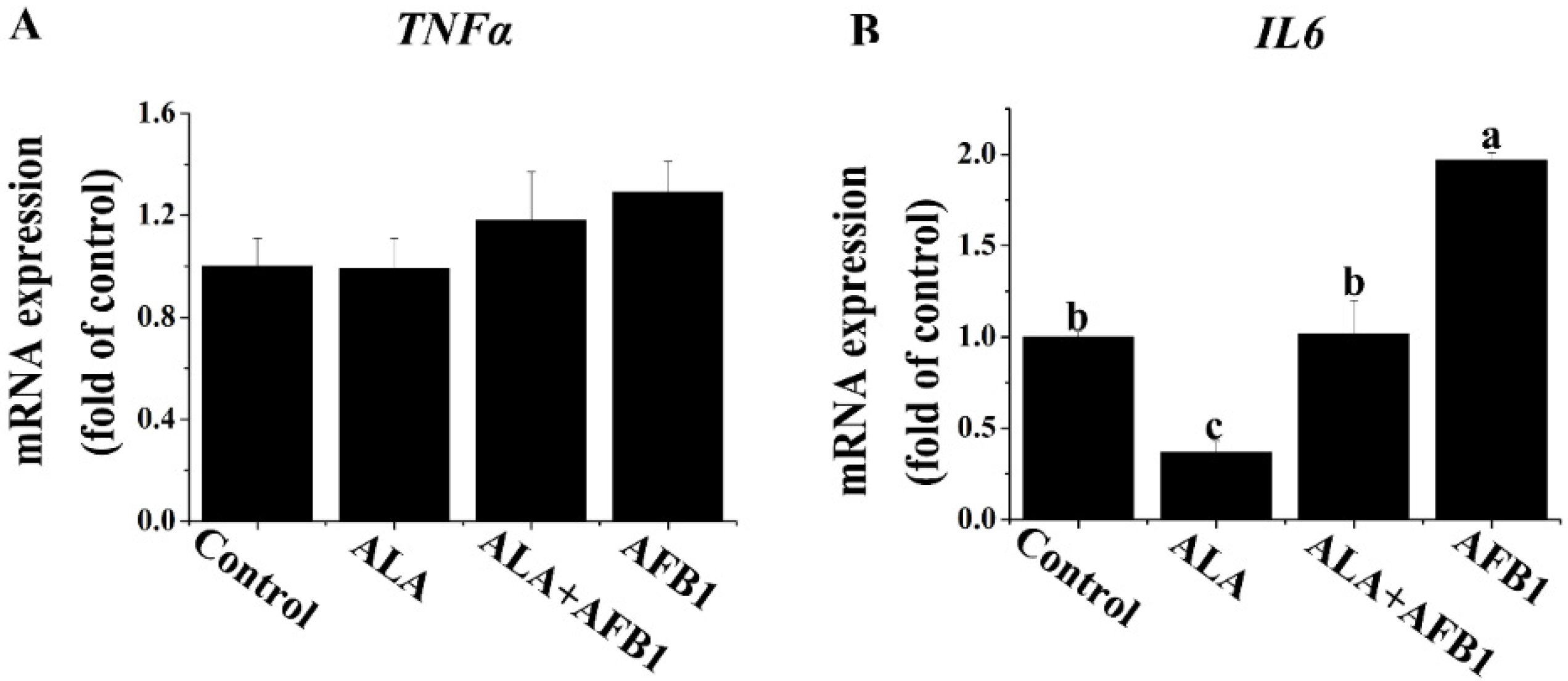
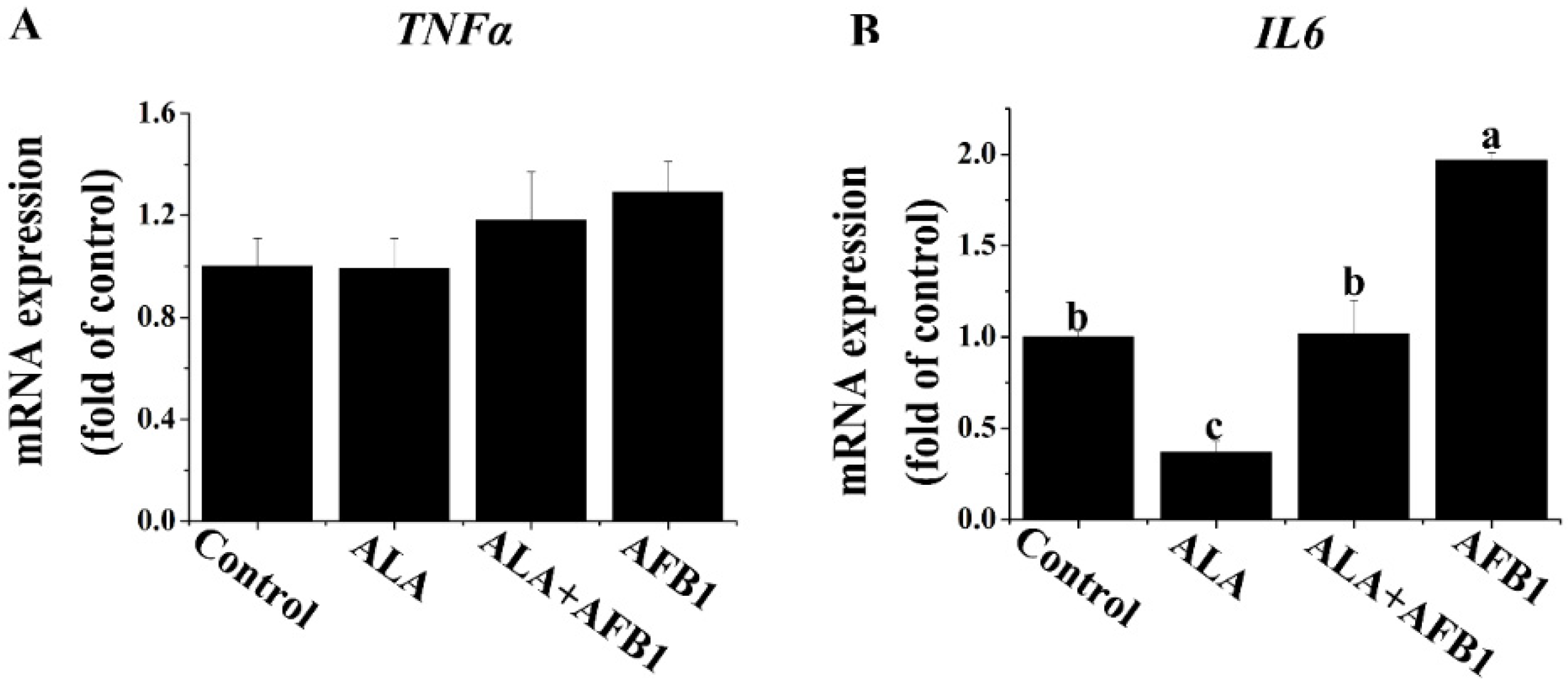
2.3. Effects of α-LA on the mRNA Levels of Proinflammatory Genes (TNF-α and IL6) in Livers of Chickens Exposed to AFB1
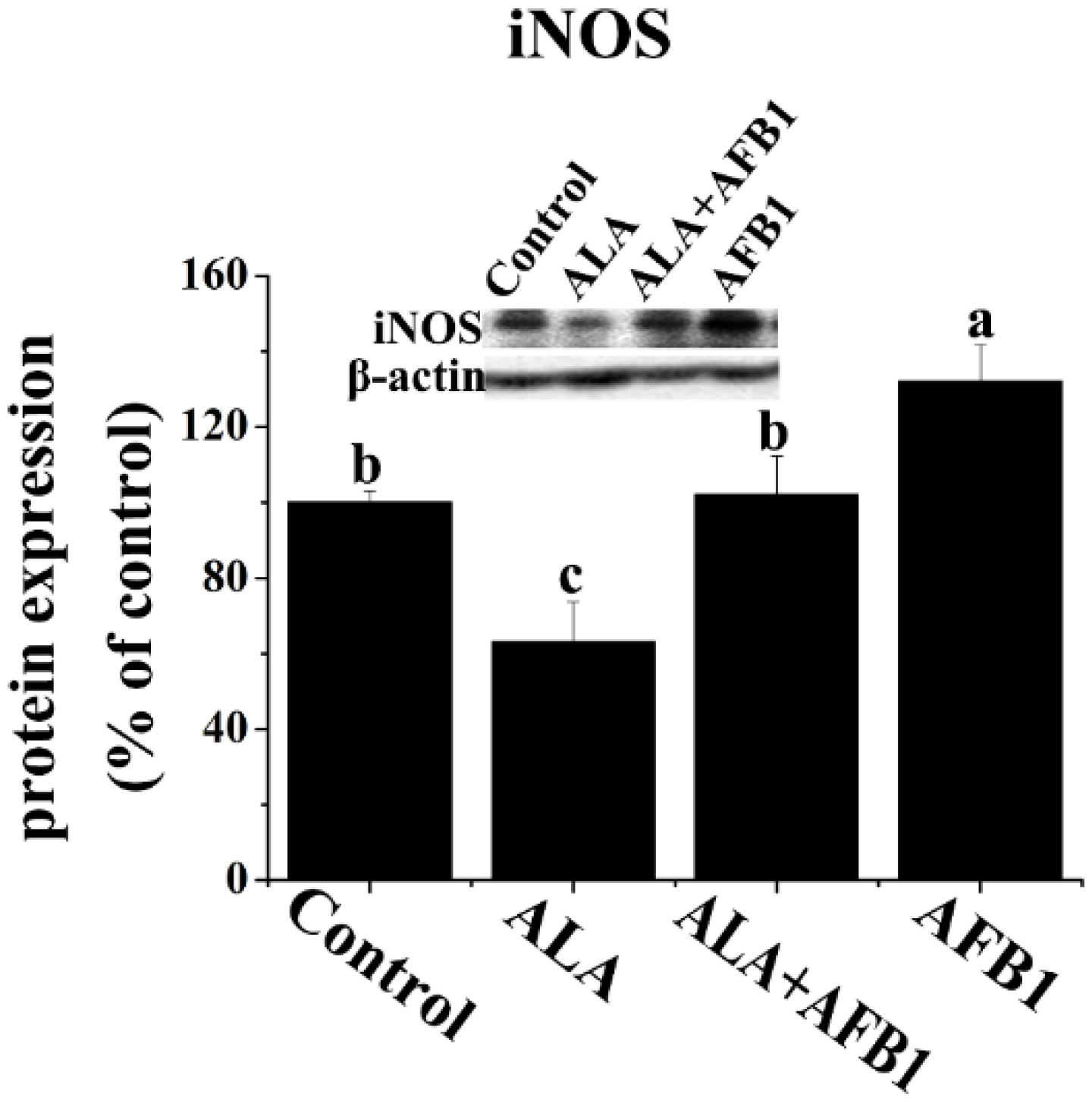
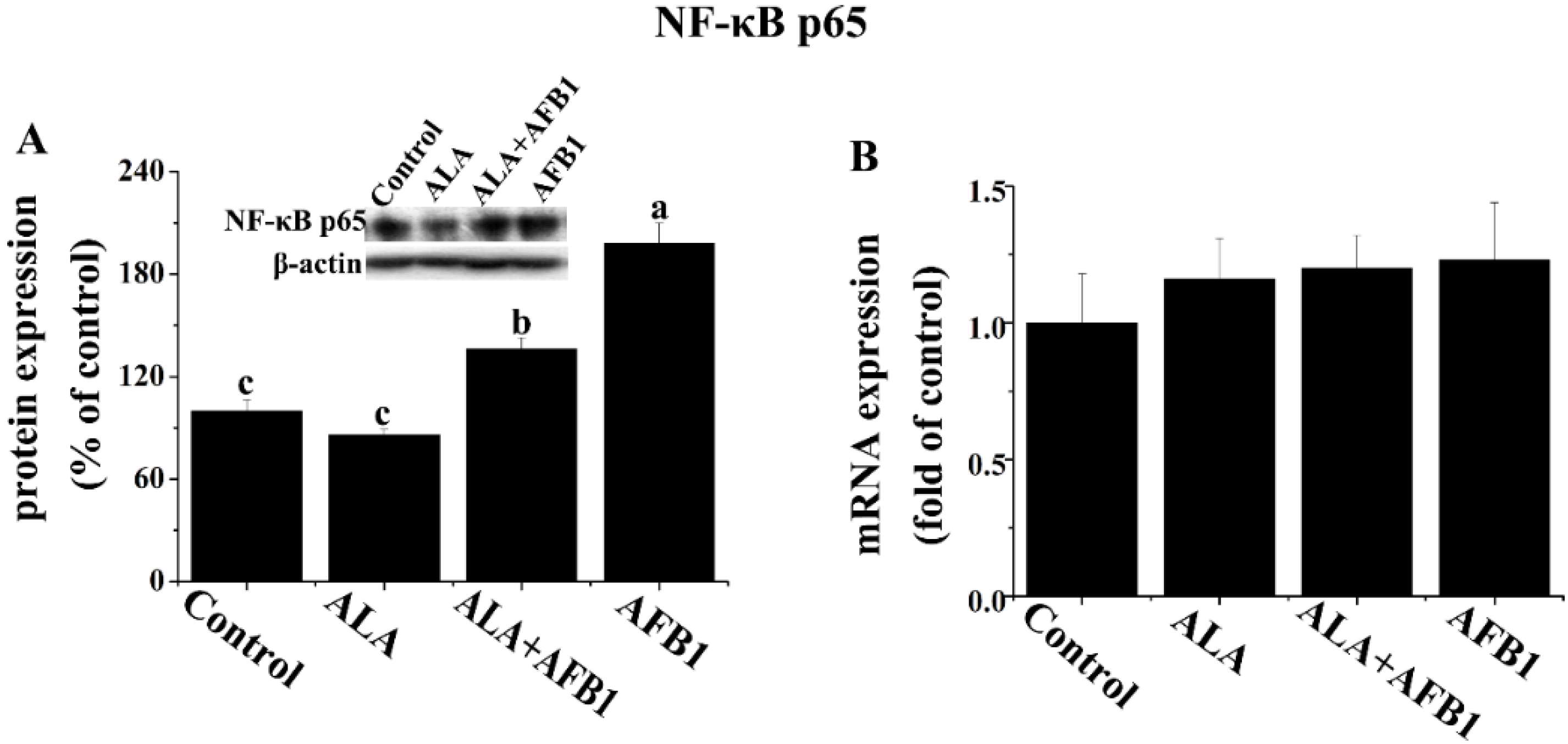
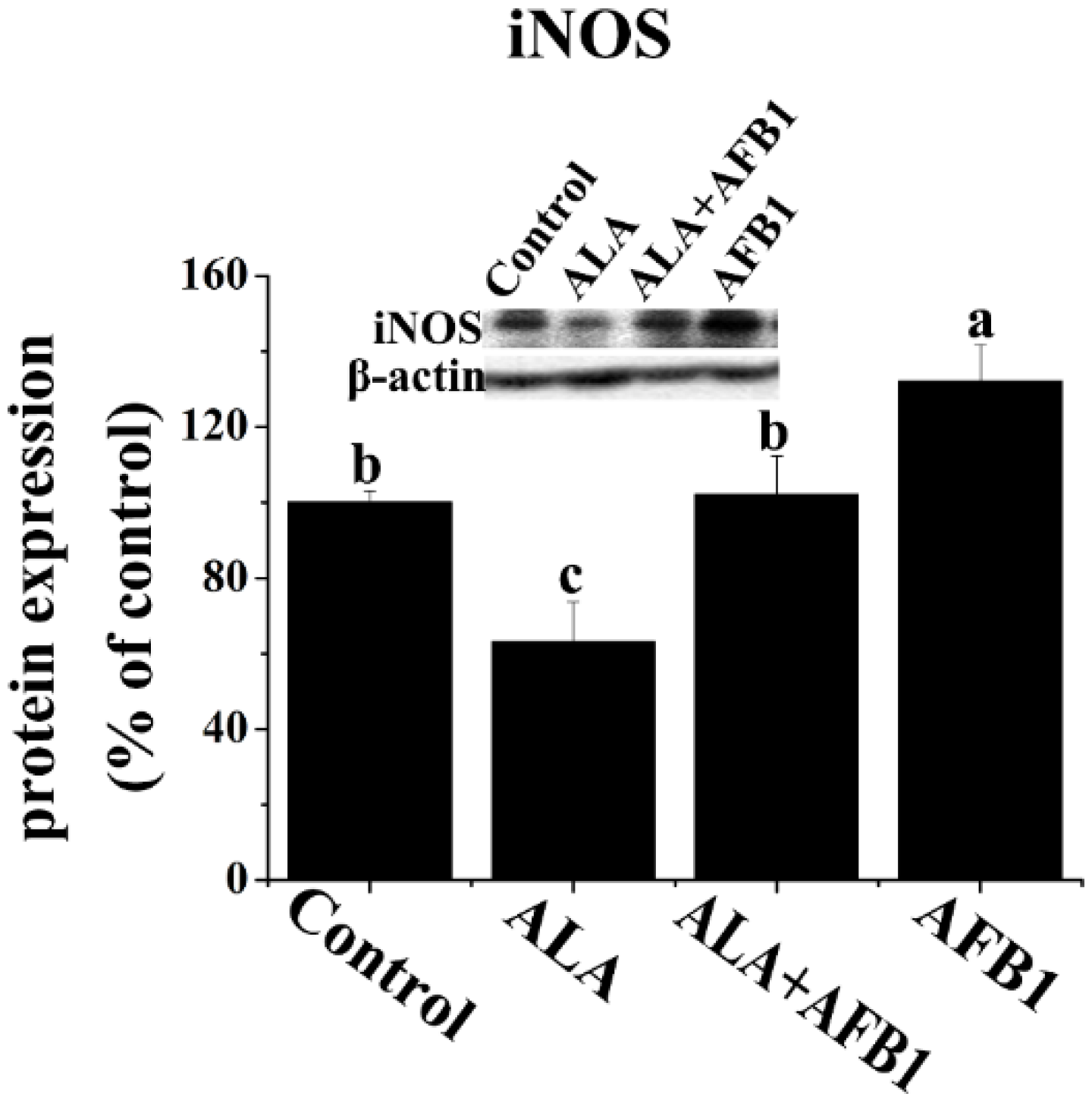
2.4. Effect of α-LA on the Expression of Hepatic NF-κB p65 and iNOS in Chickens Exposed to AFB1

3. Discussion
4. Experimental Section
4.1. Collection of Feed Ingredients Contaminated with AFB1
4.2. Animals
4.3. Experimental Design

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient | % | Nutrition Component | Content |
|---|---|---|---|
| Corn | 57.70 | Metabolizable energy (MJ/kg) | 12.55 |
| Expanded soybean | 6.00 | Crude protein (%) | 21.50 |
| Soybean meal | 8.20 | Calcium (%) | 0.99 |
| Peanut meal | 21.00 1 | Total Phosphorus (%) | 0.65 |
| Limestone | 1.30 | Nonphytate Phosphorus (%) | 0.43 |
| Dicalcium phosphate | 1.80 | Methionine (%) | 0.62 |
| Salt | 0.30 | Methionine + Cystine (%) | 0.91 |
| Corn oil | 2.00 | Lysine | 1.17 |
| Lysine [98.5%] | 0.47 | Tryptophan | 0.21 |
| DL-Methionine | 0.36 | Threonine | 0.82 |
| Threonine | 0.19 | Aflatoxin B1 (μg/kg) | 0/0/74.36/73.44 4 |
| Salinomycin | 0.07 | - | - |
| Choline chloride | 0.10 | - | - |
| 15% Chlortetracycline | 0.07 | - | - |
| Mordenzeo | 0.11 | - | - |
| Vitamin premix 2 | 0.03 | - | - |
| Mineral premix 3 | 0.30 | - | - |
| Total | 100.00 | - | - |
4.4. Preparation of Liver Homogenate
4.5. Assay of Oxidant Indices in Liver
4.6. Gene Expression Analysis
| Gene | Genebank Number | Primers Position | Primers Sequences (5′→ 3′) | Product Size |
|---|---|---|---|---|
| β-actin | AW05994 | Forward | tgcgtgacatcaaggagaag | 300 bp |
| Reverse | tgccagggtacattgtggta | |||
| SOD1 | NM_205064.155 | Forward | attaccggcttgtctgatgg | 173 bp |
| Reverse | cctccctttgcagtcacatt | |||
| SOD2 | NM_204211.1 | Forward | gccacctacgtgaacaacct | 208 bp |
| Reverse | agtcacgtttgatggcttcc | |||
| CAT | NM_001031215.1 | Forward | ccacgtggacctcttcttgt | 169 bp |
| Reverse | aaacactttcgccttgcagt | |||
| GSH-PX | NM_001163245.1 | Forward | cagcaagaaccagacaccaa | 156 bp |
| Reverse | ccaggttggttcttctccag | |||
| GSTα | NM_001001776.1 | Forward | gagtcaattcggtggctgtt | 157 bp |
| Reverse | tgctctgcaccatcttcatc | |||
| HMOX1 * | NM_205344.1 | Forward | ggtcccgaatgaatgcccttg | 137 bp |
| Reverse | accgttctcctggctcttgg | |||
| TNFα # | AY765397.1 | Forward | tgtgtatgtgcagcaacccgtagt | 229 bp |
| Reverse | ggcattgcaatttggacagaagt | |||
| IL6 | NM_204628.1 | Forward | agatgtgcaagaagttcacc | 286 bp |
| Reverse | accacttcatcgggatttat | |||
| NF-κB p65 | D13719.1 | Forward | ttgctgctggagttgatgtc | 167 bp |
| Reverse | tgctatgtgaagaggcgttg |
4.7. Western Blotting Analysis
4.8. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Hussein, H.S.; Brasel, J.M. Toxicity, metabolism, and impact of mycotoxins on humans and animals. Toxicology 2001, 167, 101–134. [Google Scholar] [CrossRef]
- Magnoli, A.P.; Monge, M.P.; Miazzo, R.D.; Cavaglieri, L.R.; Magnoli, C.E.; Merkis, C.I.; Cristofolini, A.L.; Dalcero, A.M.; Chiacchiera, S.M. Effect of low levels of aflatoxin B1 on performance, biochemical parameters, and aflatoxin B1 in broiler liver tissues in the presence of monensin and sodium bentonite. Poult. Sci. 2011, 90, 48–58. [Google Scholar] [CrossRef] [PubMed]
- Yunus, A.W.; Razzazi-Fazeli, E.; Bohm, J. Aflatoxin B1 in affecting broiler’s performance, immunity, and gastrointestinal tract: A review of history and contemporary issues. Toxins 2011, 3, 566–590. [Google Scholar] [CrossRef] [PubMed]
- Matur, E.; Ergul, E.; Akyazi, I.; Eraslan, E.; Inal, G.; Bilgic, S.; Demircan, H. Effects of Saccharomyces cerevisiae extract on haematological parameters, immune function and the antioxidant defence system in breeder hens fed aflatoxin contaminated diets. Br. Poult. Sci. 2011, 52, 541–550. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Wahhab, M.A.; Aly, S.E. Antioxidants and radical scavenging properties of vegetable extracts in rats fed aflatoxin-contaminated diet. J. Agric. Food Chem. 2003, 51, 2409–2414. [Google Scholar] [CrossRef] [PubMed]
- Packer, L.; Witt, E.H.; Tritschler, H.J. α-Lipoic acid as a biological antioxidant. Free Radic. Biol. Med. 1995, 19, 227–250. [Google Scholar] [CrossRef]
- Müller, C.; Dunschede, F.; Koch, E.; Vollmar, A.M.; Kiemer, A.K. α-Lipoic acid preconditioning reduces ischemia-reperfusion injury of the rat liver via the PI3-kinase/Akt pathway. Am. J. Physiol. Gastrointest. Liver Physiol. 2003, 285, G769–G778. [Google Scholar] [CrossRef] [PubMed]
- Karaman, M.; Özen, H.; Tuzcu, M.; Çİğremİş, Y.; Önder, F.; Özcan, K. Pathological, biochemical and haematological investigations on the protective effect of α-lipoic acid in experimental aflatoxin toxicosis in chicks. Brit. Poult. Sci. 2010, 51, 132–141. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Ma, Q.G.; Ji, C.; Zhang, J.Y.; Zhao, L.H.; Zhang, Y.; Jie, Y.Z. Dietary lipoic acid influences antioxidant capability and oxidative status of broilers. Int. J. Mol. Sci. 2011, 12, 8476–8488. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Hongtrakul, K.; Ji, C.; Ma, Q.G.; Liu, L.T.; Hu, X.X. Effects of dietary alpha-lipoic acid on anti-oxidative ability and meat quality in Arbor Acres broilers. Asian Australas. J. Anim. Sci. 2009, 22, 1195–1201. [Google Scholar] [CrossRef]
- Bai, X.M.; Ma, Q.G.; Zhao, L.H.; Xi, L.; Ji, C. Effects of α-lipoic acid supplementation on antioxidative ability and performance of sows and nursing piglets. J. Anim. Physiol. Anim. Nutr. 2012, 96, 955–961. [Google Scholar] [CrossRef] [PubMed]
- Racanelli, V.; Rehermann, B. The liver as an immunological organ. Hepatology 2006, 43, S54–S62. [Google Scholar] [CrossRef] [PubMed]
- Odabasoglu, F.; Halici, Z.; Aygun, H.; Halici, M.; Atalay, F.; Cakir, A.; Cadirci, E.; Bayir, Y.; Suleyman, H. α-Lipoic acid has anti-inflammatory and anti-oxidative properties: An experimental study in rats with carrageenan-induced acute and cotton pellet-induced chronic inflammations. Br. J. Nutr. 2011, 105, 31–43. [Google Scholar] [CrossRef] [PubMed]
- Baldwin, A.S. The NF-κB and IκB proteins: New discoveries and insights. Annu. Rev. Immunol. 1996, 14, 649–681. [Google Scholar] [CrossRef] [PubMed]
- Ames, B.N. Dietary carcinogens and anticarcinogens oxygen radicals and degenerative diseases. Science 1983, 221, 1256–1264. [Google Scholar] [CrossRef] [PubMed]
- Shen, H.M.; Shi, C.Y.; Shen, Y.; Ong, C.N. Detection of elevated reactive oxygen species level in cultured rat hepatocytes treated with aflatoxin B1. Free Radic. Biol. Med. 1996, 21, 139–146. [Google Scholar] [CrossRef]
- Rastogi, R.; Srivastava, A.K.; Rastogi, A.K. Long term effect of aflatoxin B1 on lipid peroxidation in rat liver and kidney: Effect of picroliv and silymarin. Phytother. Res. 2001, 15, 307–310. [Google Scholar] [CrossRef] [PubMed]
- Ho, Y.; Ho, S.; Lai, C.; Liu, H.; Tai, C.; Pan, M.; Wang, Y. Dihydrolipoic acid inhibits skin tumor promotion through anti-inflammation and anti-oxidation. Biochem. Pharmacol. 2007, 73, 1786–1795. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Ma, Q.G.; Zhao, L.H.; Guo, Y.Q.; Duan, G.X.; Zhang, J.Y.; Ji, C. Protective efficacy of alpha-lipoic acid against aflatoxin B1-induced oxidative damage in the liver. Asian Australas. J. Anim. Sci. 2014, 27, 907–915. [Google Scholar] [CrossRef] [PubMed]
- Kodama, M.; Inoue, F.; Akao, M. Enzymatic and non-enzymatic formation of free radicals from aflatoxin B1. Free Radic. Res. Commun. 1990, 10, 137–142. [Google Scholar] [CrossRef] [PubMed]
- Kanbur, M.; Eraslan, G.; Sarica, Z.S.; Aslan, Ö. The effects of evening primrose oil lipid peroxidation induced by subacute aflatoxin exposure in mice. Food Chem. Toxicol. 2011, 49, 1960–1964. [Google Scholar] [CrossRef] [PubMed]
- Naaz, F.; Javed, S.; Abdin, M.Z. Hepatoprotective effect of ethanolic extract of Phyllanthus amarus Schum. et Thonn. on aflatoxin B1-induced liver damage in mice. J. Ethnopharmacol. 2007, 113, 503–509. [Google Scholar] [CrossRef] [PubMed]
- Dwivedi, N.; Flora, G.; Kushwaha, P.; Flora, S. α-lipoic acid protects oxidative stress, changes in cholinergic system and tissue histopathology during co-exposure to arsenic-dichlorvos in rats. Environ. Toxicol. Pharmacol. 2014, 37, 7–23. [Google Scholar] [CrossRef] [PubMed]
- Hinton, D.M.; Myers, M.J.; Raybourne, R.A.; Francke-Carroll, S.; Sotomayor, R.E.; Shaddock, J.; Warbritton, A.; Chou, M.W. Immunotoxicity of aflatoxin B1 in rats: Effects on lymphocytes and the inflammatory response in a chronic intermittent dosing study. Toxicol. Sci. 2003, 73, 362–377. [Google Scholar] [CrossRef] [PubMed]
- Park, E.J.; Lee, J.H.; Yu, G.; He, G.; Ali, S.R.; Holzer, R.G.; Österreicher, C.H.; Takahashi, H.; Karin, M. Dietary and genetic obesity promote liver inflammation and tumorigenesis by enhancing IL-6 and TNF expression. Cell 2010, 140, 197–208. [Google Scholar] [CrossRef] [PubMed]
- Yarru, L.P.; Settivari, R.S.; Gowda, N.K.S.; Antoniou, E.; Ledoux, D.R.; Rottinghaus, G.E. Effects of turmeric (Curcuma longa) on the expression of hepatic genes associated with biotransformation, antioxidant, and immune systems in broiler chicks fed aflatoxin. Poult. Sci. 2009, 88, 2620–2627. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.Y.; Li, Y.; Han, P.; Wu, N.; He, B.; Lu, Y.; Li, S.W.; Liu, Y.; Zhao, S.; Liu, L.T. Amelioration of lipid abnormalities by α-Lipoic acid through antioxidative and anti-inflammatory effects. Obesity 2011, 19, 1647–1653. [Google Scholar] [CrossRef] [PubMed]
- Jeong, W.; Kim, I.; Hu, R.; Kong, A.T. Modulatory properties of various natural chemopreventive agents on the activation of NF-κB signaling pathway. Pharm. Res. 2004, 21, 661–670. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Wei, H.; Hagen, T.; Frei, B. α-Lipoic acid attenuates LPS-induced inflammatory responses by activating the phosphoinositide 3-Kinase/Akt signaling pathway. Proc. Natl. Acad. Sci. USA 2007, 104, 4077–4082. [Google Scholar] [CrossRef] [PubMed]
- Sass, G.; Koerber, K.; Bang, R.; Guehring, H.; Tiegs, G. Inducible nitric oxide synthase is critical for immune-mediated liver injury in mice. J. Clin. Investig. 2001, 107, 439–447. [Google Scholar] [CrossRef] [PubMed]
- Graziani, F.; Pujol, A.; Nicoletti, C.; Pinton, P.; Armand, L.; di Pasquale, E.; Oswald, I.P.; Perrier, J.; Maresca, M. The Food-Associated Ribotoxin Deoxynivalenol Modulates Inducible NO Synthase in Human Intestinal Cell Model. Toxicol. Sci. 2015, 145, 372–382. [Google Scholar] [CrossRef] [PubMed]
- Razafimanjato, H.; Benzaria, A.; Taïeb, N.; Guo, X.J.; Vidal, N.; Di Scala, C.; Varini, K.; Maresca, M. The ribotoxin deoxynivalenol affects the viability and functions of glial cells. Glia 2011, 59, 1672–1683. [Google Scholar] [CrossRef] [PubMed]
- Trucksess, M.W.; Stack, M.E.; Nesheim, S.; Albert, R.H.; Romer, T.R. Multifunctional column coupled with liquid chromatography for determination of aflatoxins B1, B2, G1, and G2 in corn, almonds, brazil nuts, peanuts, and pistachio nuts: Collaborative study. J. AOAC Int. 1994, 77, 1512–1521. [Google Scholar] [PubMed]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem. 1979, 95, 351–358. [Google Scholar] [CrossRef]
- Gabor, G.; Allon, N. Spectrofluorometric method for NO determination. Anal. Biochem. 1994, 220, 16–19. [Google Scholar] [CrossRef] [PubMed]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [PubMed]
- Druyan, S.; Cahaner, A.; Ashwell, C. The expression patterns of hypoxia-inducing factor subunit α-1, heme oxygenase, hypoxia upregulated protein 1, and cardiac troponin T during development of the chicken heart. Poult. Sci. 2007, 86, 2384–2389. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.H.; Lillehoj, H.S.; Hyen Lee, S.; Woon Park, D.; Lillehoj, E.P. Molecular cloning and characterization of chicken lipopolysaccharide-induced TNF-α factor (LITAF). Dev. Comp. Immunol. 2006, 30, 919–929. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, Q.; Li, Y.; Fan, Y.; Zhao, L.; Wei, H.; Ji, C.; Zhang, J. Molecular Mechanisms of Lipoic Acid Protection against Aflatoxin B1-Induced Liver Oxidative Damage and Inflammatory Responses in Broilers. Toxins 2015, 7, 5435-5447. https://doi.org/10.3390/toxins7124879
Ma Q, Li Y, Fan Y, Zhao L, Wei H, Ji C, Zhang J. Molecular Mechanisms of Lipoic Acid Protection against Aflatoxin B1-Induced Liver Oxidative Damage and Inflammatory Responses in Broilers. Toxins. 2015; 7(12):5435-5447. https://doi.org/10.3390/toxins7124879
Chicago/Turabian StyleMa, Qiugang, Yan Li, Yu Fan, Lihong Zhao, Hua Wei, Cheng Ji, and Jianyun Zhang. 2015. "Molecular Mechanisms of Lipoic Acid Protection against Aflatoxin B1-Induced Liver Oxidative Damage and Inflammatory Responses in Broilers" Toxins 7, no. 12: 5435-5447. https://doi.org/10.3390/toxins7124879
APA StyleMa, Q., Li, Y., Fan, Y., Zhao, L., Wei, H., Ji, C., & Zhang, J. (2015). Molecular Mechanisms of Lipoic Acid Protection against Aflatoxin B1-Induced Liver Oxidative Damage and Inflammatory Responses in Broilers. Toxins, 7(12), 5435-5447. https://doi.org/10.3390/toxins7124879