Is Increased Susceptibility to Balkan Endemic Nephropathy in Carriers of Common GSTA1 (*A/*B) Polymorphism Linked with the Catalytic Role of GSTA1 in Ochratoxin A Biotransformation? Serbian Case Control Study and In Silico Analysis

 and
and 
Abstract
:1. Introduction

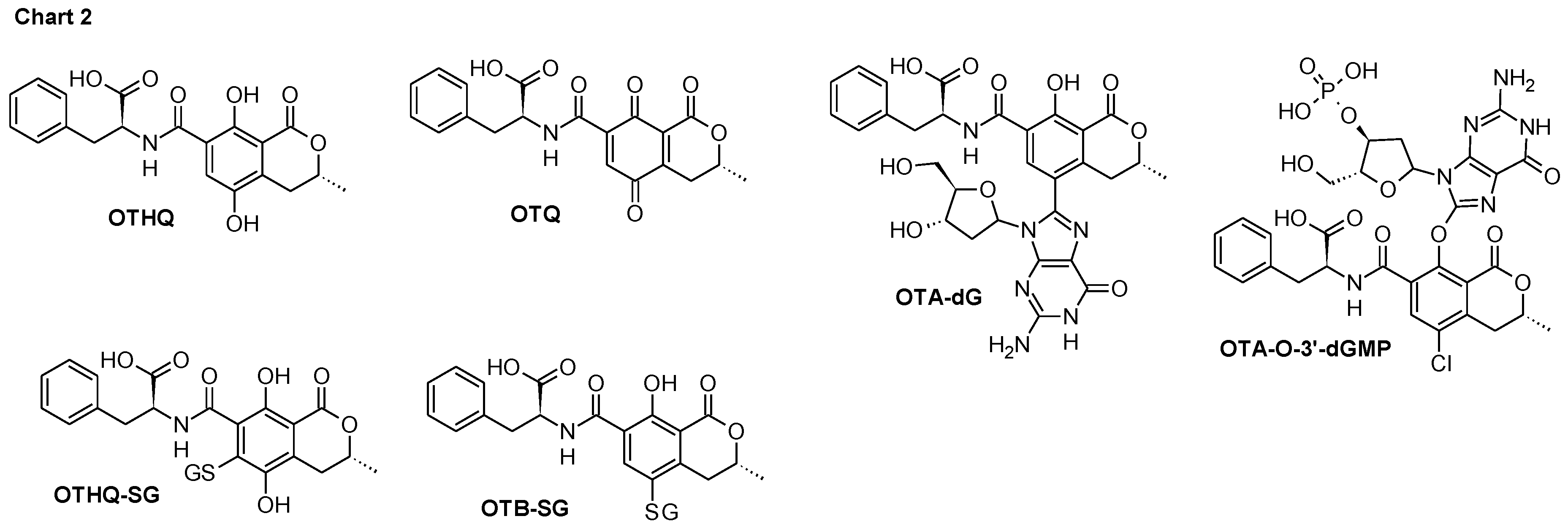
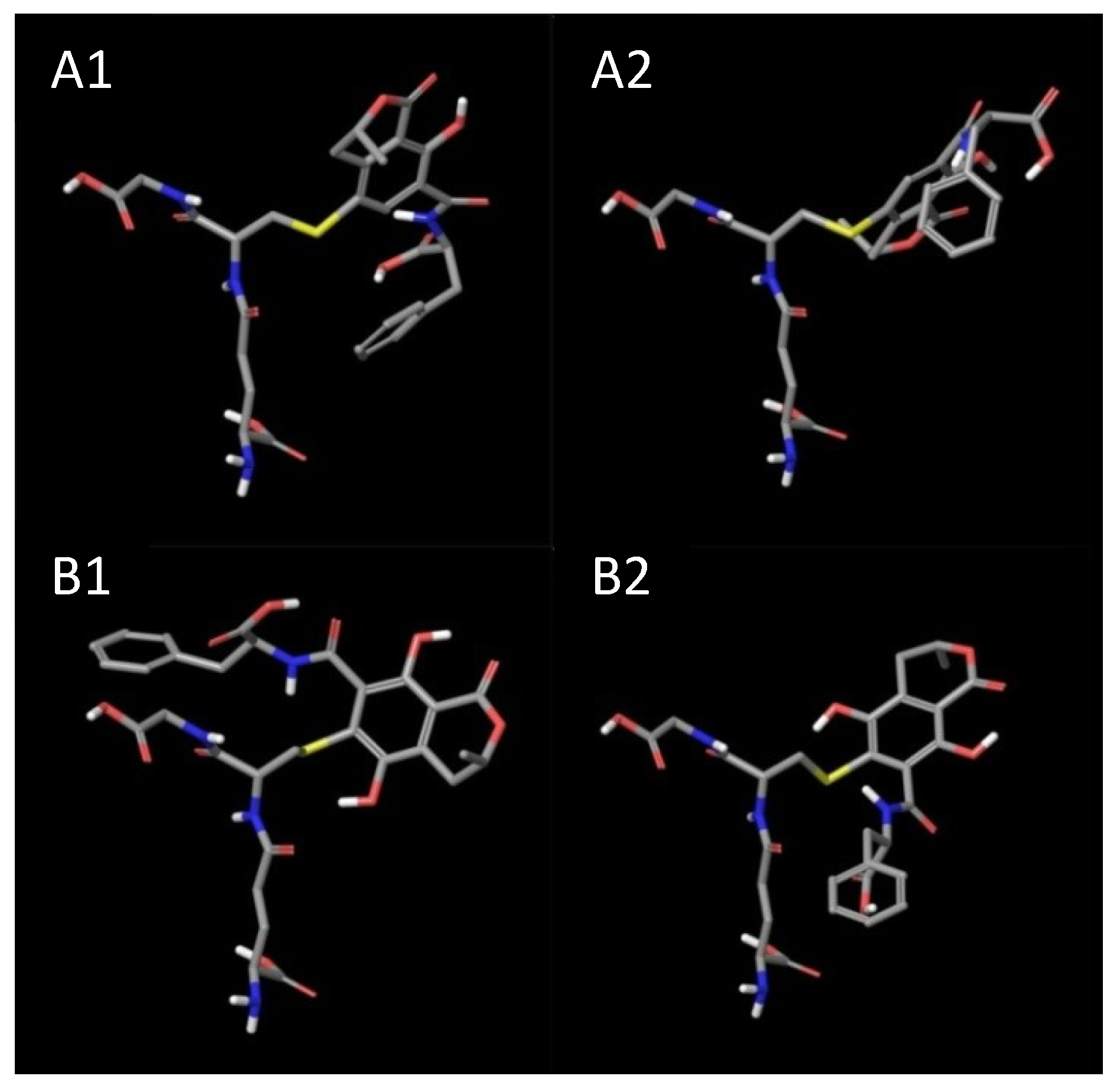
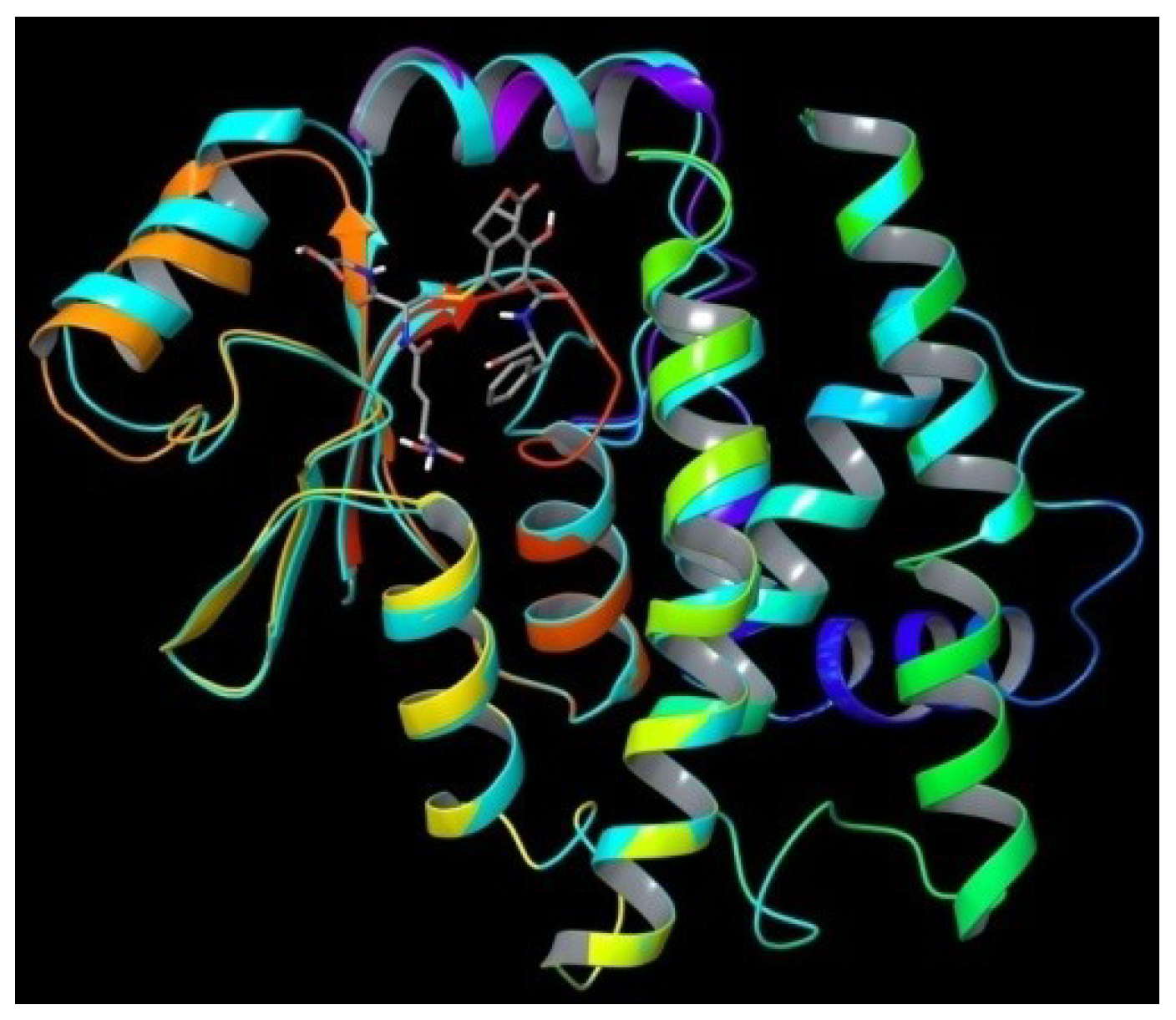
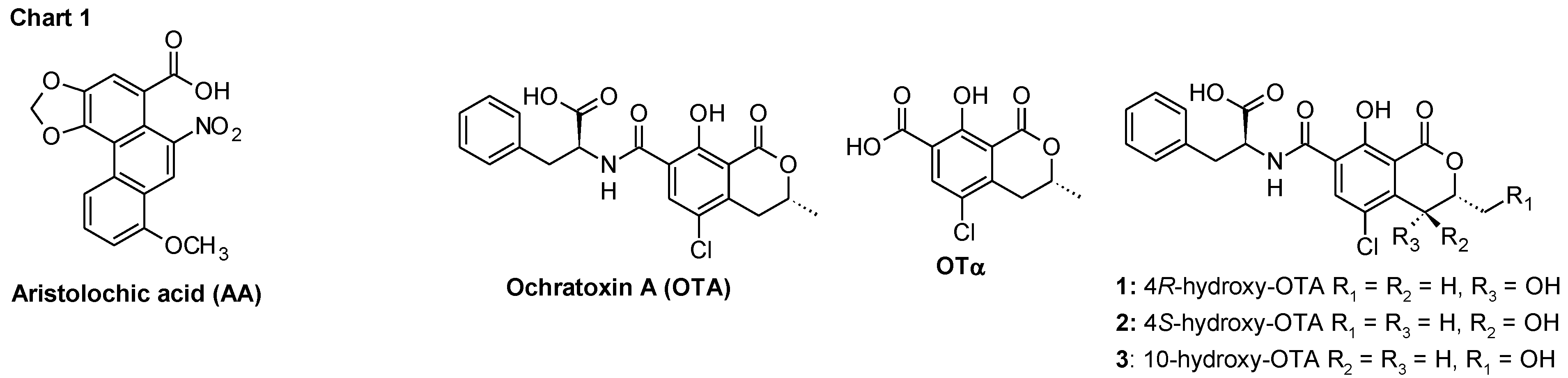
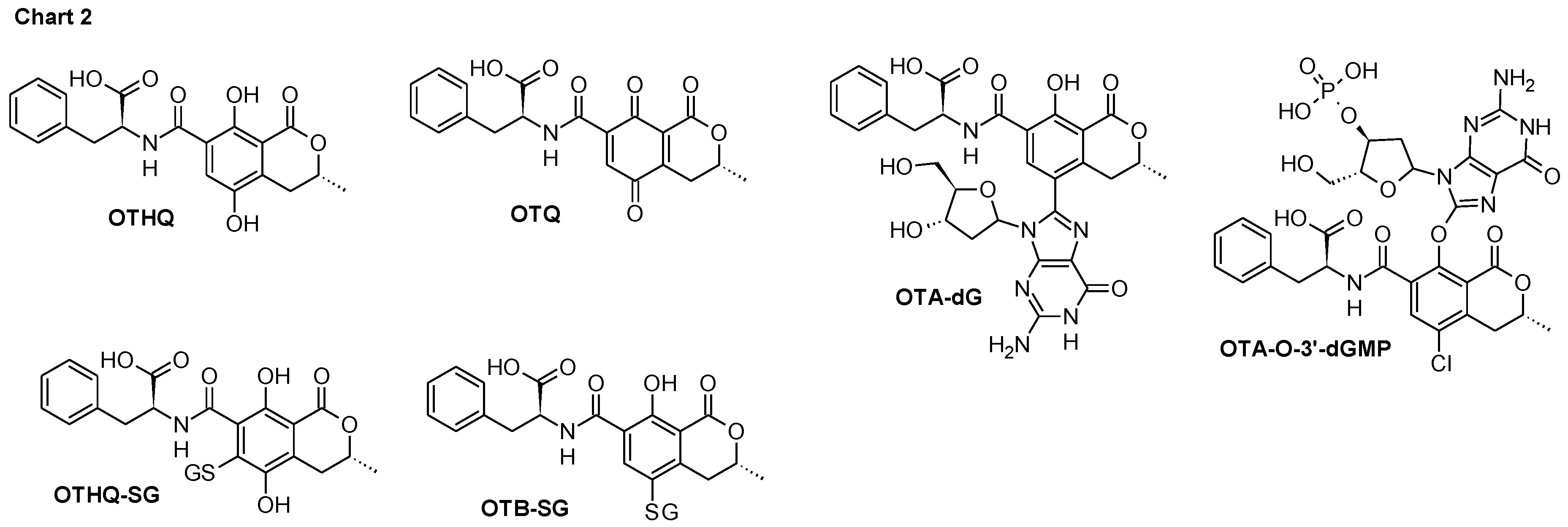
2. Results and Discussion

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Demographic characteristics | Cases | Controls | p value |
|---|---|---|---|
| Sex | 0.134 | ||
| Male n(%) | 116 (56) | 66 (48) | |
| Female n(%) | 91 (44) | 72 (52) | |
| Age | |||
| Mean ± SD | 70.60 ± 6.54 | 69.33 ± 9.98 | 0.153 |
| Genotypes of GST | Cases n (%) | Controls n (%) | OR a with 95% CI | p value |
|---|---|---|---|---|
| GSTA1 C-69T | ||||
| *A/*A | 58 (28) | 53 (39) | 1.0 b | |
| *A/*B | 110 (53) | 57 (41) | 1.8 (1.1–2.9) | 0.021 |
| *B/*B | 39 (19) | 28 (20) | 1.3 (0.7–2.4) | 0.377 |
| *A/*B + *B/*B | 149 (72) | 85 (61) | 1.6 (1.0–2.6) | 0.037 |
| GSTM1 | ||||
| active c | 115 (56) | 75 (54) | 1.0 b | |
| null d | 92 (44) | 63 (46) | 0.9 (0.6–1.5) | 0.790 |
| GSTT1 | ||||
| active c | 168 (81) | 100 (72) | 1.5 (0.9–2.5) | 0.126 |
| null d | 39 (19) | 38 (28) | 1.0 b | |
| GSTP1 A1578G | ||||
| Ile/Ile | 97 (47) | 62 (45) | 1.0 b | |
| Ile/Val | 92 (45) | 57 (42) | 1.0 (0.6–1.6) | 0.953 |
| Val/Val | 16 (8) | 18 (13) | 0.6 (0.3–1.2) | 0.142 |
| Ile/Val + Val/Val | 108 (53) | 75 (55) | 0.9 (0.6–1.4) | 0.666 |
| Genotypes | GSTA1 | |
|---|---|---|
| *A/*A | *A/*B+*B/*B | |
| GSTT1 activea | ||
| Ca/Co | 46/38 | 122/62 |
| OR b (95% CI) | 1.4 (0.6–3.4) | 2.3 (1.0–5.3) c |
| GSTT1 nulld | ||
| Ca/Co | 12/15 | 27/23 |
| OR b (95% CI) | 1.0 e | 1.5 (0.6–3.9) |
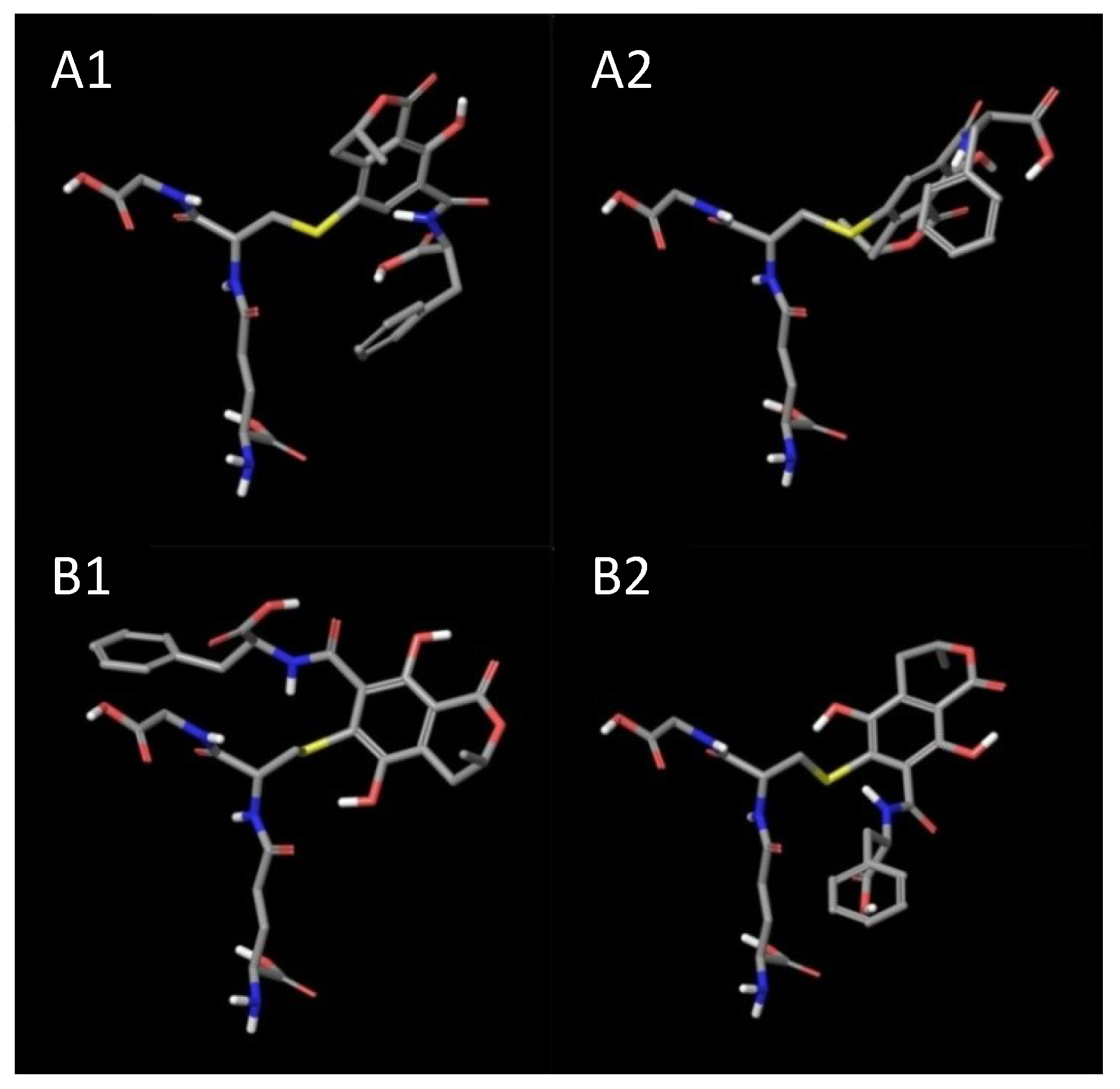
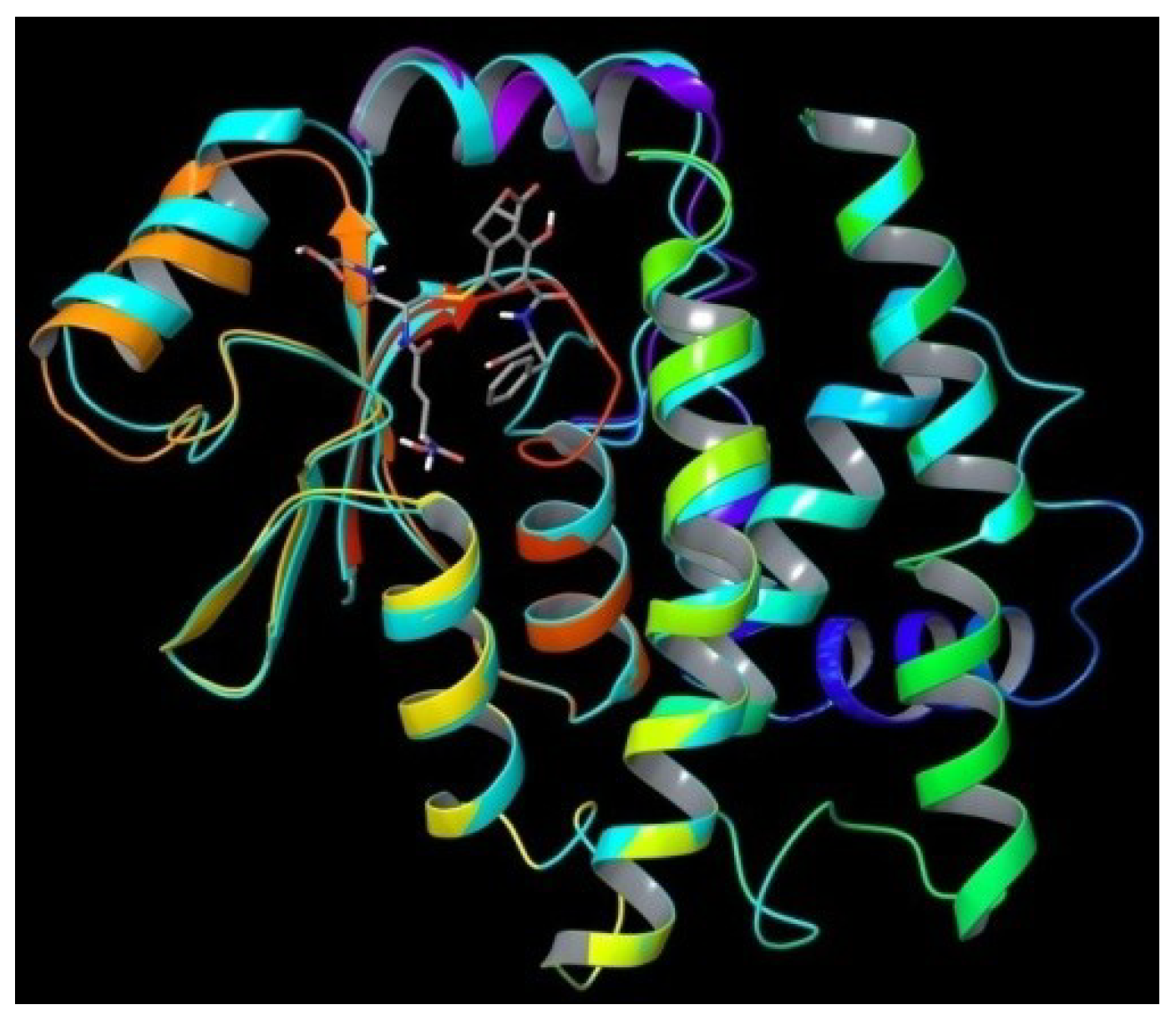
3. Experimental Section
3.1. Study Participants
3.2. GST Genotyping
3.3. Statistical Analysis
3.4. Molecular Modeling (in Silico)
4. Conclusions
Supplementary Files
Supplementary File 1Acknowledgments
Author Contributions
Conflict of Interest
References
- Jelakovic, B.; Nikolic, J.; Radovanovic, Z.; Nortier, J.; Cosyns, J.P.; Grollman, A.P.; Basic-Jukic, N.; Belicza, M.; Bukvic, D.; Cavaljuga, S.; et al. Consensus statement on screening, diagnosis, classification and treatment of endemic (Balkan) nephropathy. Nephrol. Dial. Transplant. 2013, 1–9. [Google Scholar] [CrossRef]
- Pfohl-Leszkowicz, A.; Tozlovanu, M.; Manderville, R.; Peraica, M.; Castegnaro, M.; Stefanovic, V. New molecular and field evidences for the implication of mycotoxins but not aristolochic acid in human nephropathy and urinary tract tumor. Mol. Nutr. Food Res. 2007, 51, 1131–1146. [Google Scholar] [CrossRef]
- Pfohl-Leszkowicz, A.; Grosse, Y.; Castegnaro, M.; Nicolov, I.G.; Chernozemsky, I.N.; Bartsch, H.; Betbeder, A.M.; Creppy, E.E.; Dirheimer, G. Ochratoxin A-related DNA adducts in urinary tract tumours of Bulgarian subjects. IARC Sci. Publ. 1993, 124, 141–148. [Google Scholar]
- Pfohl-Leszkowicz, A. Ochratoxin A and aristolochic acid involvement in nephropathies and associated urothelial tract tumours. Arh. Hig. Rada. Toksikol. 2009, 60, 465–483. [Google Scholar] [CrossRef]
- Daly, A.K.; Cholerton, S.; Armstrong, M.; Idle, J.R. Genotyping for polymorphisms in xenobiotic metabolism as a predictor of disease susceptibility. Environ. Health Perspect. 1994, 102, 55–61. [Google Scholar] [CrossRef]
- Stander, M.A.; Steyn, P.S.; van der Westhuizen, F.H.; Payne, B.E. A kinetic study into the hydrolysis of the ochratoxins and analogues by carboxypeptidase A. Chem. Res. Toxicol. 2001, 14, 302–304. [Google Scholar] [CrossRef]
- Dai, J.; Park, G.; Wright, M.W.; Adams, M.; Akman, S.A.; Manderville, R.A. Detection and characterization of a glutathione conjugate of Ochratoxin A. Chem. Res. Toxicol. 2002, 15, 1581–1588. [Google Scholar] [CrossRef]
- Wu, Q.; Dohnal, V.; Huang, L.; Kuča, K.; Wang, X.; Chen, G.; Yuan, Z. Metabolic pathways of ochratoxin A. Curr. Drug Metab. 2011, 12, 1–10. [Google Scholar] [CrossRef]
- Atanasova, S.Y.; von Ahsen, N.; Toncheva, D.I.; Dimitrov, T.G.; Oellerich, M.; Armstrong, V.W. Genetic polymorphisms of cytochrome P450 among patients with Balkan endemic nephropathy (BEN). Clin. Biochem. 2005, 38, 223–228. [Google Scholar] [CrossRef]
- Tozlovanu, M.; Canadas, D.; Pfohl-Leszkowicz, A.; Frenette, C.; Paugh, R.J.; Manderville, R.A. Glutathione conjugates of ochratoxin A as biomarkers of exposure. Arh. Hig. Rada. Toksikol. 2012, 63, 417–427. [Google Scholar]
- Rowe, J.D.; Nieves, E.; Listowsky, I. Subunit diversity and tissue distribution of human glutathione S-transferases: Interpretations based on electrospray ionization-MS and peptide sequence-specific antisera. Biochem. J. 1997, 325, 481–486. [Google Scholar]
- Dirr, H.W.; Wallace, L.A. Role of the C-Terminal Helix 9 in the stability and ligandin function of class R glutathione transferase A1-1. Biochemistry 1999, 38, 15631–15640. [Google Scholar] [CrossRef]
- Seidegård, J.; Vorachek, W.R.; Pero, R.W.; Pearson, W.R. Hereditary differences in the expression of the human glutathione transferase active on trans-stilbene oxide are due to a gene deletion. Proc. Natl. Acad. Sci. USA 1988, 85, 7293–7297. [Google Scholar]
- Pemble, S.; Schroeder, K.R.; Spencer, S.R.; Meyer, D.J.; Hallier, E.; Bolt, H.M.; Ketterer, B.; Taylor, J.B. Human glutathione S-transferase theta (GSTT1): cDNA cloning and the characterization of a genetic polymorphism. Biochem. J. 1994, 300, 271–276. [Google Scholar]
- Zimniak, P.; Nanduri, B.; Pikuła, S.; Bandorowicz-Pikuła, J.; Singhal, S.S.; Srivastava, S.K.; Awasthi, S.; Awasthi, Y.C. Naturally occurring human glutathione S-transferase GSTP1-1 isoforms with isoleucine and valine in position 104 differ in enzymic properties. Eur. J. Biochem. 1994, 224, 893–899. [Google Scholar] [CrossRef]
- Sundberg, K.; Johansson, A.S.; Stenberg, G.; Widersten, M.; Seidel, A.; Mannervik, B.; Jernström, B. Differences in the catalytic efficiencies of allelic variants of glutathione transferase P1-1 towards carcinogenic diol epoxides of polycyclic aromatic hydrocarbons. Carcinogenesis 1998, 19, 433–436. [Google Scholar] [CrossRef]
- Coles, B.F.; Morel, F.; Rauch, C.; Huber, W.W.; Yang, M.; Teitel, C.H.; Green, B.; Lang, N.P.; Kadlubar, F.F. Effect of polymorphism in the human glutathione S-transferase A1 promoter on hepatic GSTA1 and GSTA2 expression. Pharmacogenetics 2001, 11, 663–669. [Google Scholar] [CrossRef]
- Suvakov, S.; Damjanovic, T.; Stefanovic, A.; Pekmezovic, T.; Savic-Radojevic, A.; Pljesa-Ercegovac, M.; Matic, M.; Djukic, T.; Coric, V.; Jakovljevic, J.; et al. Glutathione S-transferase A1, M1, P1 and T1 null or low-activity genotypes are associated with enhanced oxidative damage among haemodialysis patients. Nephrol. Dial. Transplant. 2013, 28, 202–212. [Google Scholar] [CrossRef]
- Matic, M.; Pekmezovic, T.; Djukic, T.; Mimic-Oka, J.; Dragicevic, D.; Krivic, B.; Suvakov, S.; Savic-Radojevic, A.; Pljesa-Ercegovac, M.; Tulic, C.; et al. GSTA1, GSTM1, GSTP1, and GSTT1 polymorphisms and susceptibility to smoking-related bladder cancer: A case-control study. Urol. Oncol. 2013, 31, 1184–1192. [Google Scholar] [CrossRef]
- Andonova, I.E.; Sarueva, R.B.; Horvath, A.D.; Simeonov, V.A.; Dimitrov, P.S.; Petropoulos, E.A.; Ganev, V.S. Balkan endemic nephropathy and genetic variants of glutathione S-transferases. J. Nephrol. 2004, 17, 390–398. [Google Scholar]
- Toncheva, D.I.; von Ahsen, N.; Atanasova, S.Y.; Dimitrov, T.G.; Armstrong, V.W.; Oellerich, M. Identification of NQO1 and GSTs genotype frequencies in Bulgarian patients with Balkan endemic nephropathy. J. Nephrol. 2004, 17, 384–389. [Google Scholar]
- Lebrun, S.; Golka, K.; Schulze, H.; Föllmann, W. Glutathione S-transferase polymorphisms and ochratoxin A toxicity in primary human urothelial cells. Toxicology 2006, 224, 81–90. [Google Scholar] [CrossRef]
- Moore, L.E.; Boffetta, P.; Karami, S.; Brennan, P.; Stewart, P.S.; Hung, R.; Zaridze, D.; Matveev, V.; Janout, V.; Kollarova, H.; et al. Occupational trichloroethylene exposure and renal carcinoma risk: Evidence of genetic susceptibility by reductive metabolism gene variants. Cancer Res. 2010, 70, 6527–6536. [Google Scholar] [CrossRef]
- Hong, Y.C.; Park, H.S.; Ha, E.H. Influence of genetic susceptibility on the urinary excretion of 8-hydroxydeoxyguanosine of firefighters. Occup. Environ. Med. 2000, 57, 370–375. [Google Scholar] [CrossRef]
- Buzio, L.; de Palma, G.; Mozzoni, P.; Tondel, M.; Buzio, C.; Franchini, I.; Axelson, O.; Mutti, A. Glutathione S-transferases M1-1 and T1-1 as risk modifiers for renal cell cancer associated with occupational exposure to chemicals. Occup. Environ. Med. 2003, 60, 789–793. [Google Scholar] [CrossRef]
- Shokeer, A.; Mannervik, B. Residue 234 is a master switch of the alternative-substrate activity profile of human and rodent theta class glutathione transferase T1-1. Biochim. Biophys. Acta 2010, 1800, 466–473. [Google Scholar] [CrossRef]
- Sundberg, A.; Appelkvist, E.L.; Dallner, G.; Nilsson, R. Glutathione transferases in the urine: Sensitive methods for detection of kidney damage induced by nephrotoxic agents in humans. Environ. Health Perspect. 1994, 102, 293–296. [Google Scholar] [CrossRef]
- Rahimtula, A.D.; Béréziat, J.C.; Bussacchini-Griot, V.; Bartsch, H. Lipid peroxidation as a possible cause of ochratoxin A toxicity. Biochem. Pharmacol. 1988, 37, 4469–4477. [Google Scholar] [CrossRef]
- Meki, A.R.; Hussein, A.A. Melatonin reduces oxidative stress induced by ochratoxin A in rat liver and kidney. Comp. Biochem. Physiol. C 2001, 130, 305–313. [Google Scholar]
- Palabiyik, S.S.; Erkekoglu, P.; Zeybek, N.D.; Kizilgun, M.; Baydar, D.E.; Sahin, G.; Giray, B.K. Protective effect of lycopene against ochratoxin A induced renal oxidative stress and apoptosis in rats. Exp. Toxicol. Pathol. 2013, 65, 853–861. [Google Scholar] [CrossRef]
- Marin-Kuan, M.; Ehrlich, V.; Delatour, T.; Cavin, C.; Schilter, B. Evidence for a role of oxidative stress in the carcinogenicity of ochratoxin A. J. Toxicol. 2011. [Google Scholar] [CrossRef]
- Guilford, F.T.; Hope, J. Deficient glutathione in the pathophysiology of mycotoxin-related illness. Toxins 2014, 6, 608–623. [Google Scholar] [CrossRef]
- Arbillaga, L.; Azqueta, A.; Ezpeleta, O.; López de Cerain, A. Oxidative DNA damage induced by Ochratoxin A in the HK-2 human kidney cell line: Evidence of the relationship with cytotoxicity. Mutagenesis 2007, 22, 35–42. [Google Scholar]
- Cavin, C.; Delatour, T.; Marin-Kuan, M.; Holzhäuser, D.; Higgins, L.; Bezençon, C.; Guignard, G.; Junod, S.; Richoz-Payot, J.; Gremaud, E. Reduction in antioxidant defenses may contribute to ochratoxin A toxicity and carcinogenicity. Toxicol. Sci. 2007, 96, 30–39. [Google Scholar]
- Boesch-Saadatmandi, C.; Loboda, A.; Jozkowicz, A.; Huebbe, P.; Blank, R.; Wolffram, S.; Dulak, J.; Rimbach, G. Effect of ochratoxin A on redox-regulated transcription factors, antioxidant enzymes and glutathione-S-transferase in cultured kidney tubulus cells. Food Chem. Toxicol. 2008, 46, 2665–2671. [Google Scholar] [CrossRef]
- Limonciel, A.; Jennings, P. A review of the evidence that ochratoxin A is an Nrf2 inhibitor: Implications for nephrotoxicity and renal carcinogenicity. Toxins 2014, 6, 371–379. [Google Scholar] [CrossRef]
- Coles, B.F.; Kadlubar, F.F. Human alpha class glutathione S-transferases: Genetic polymorphism, expression, and susceptibility to disease. Methods Enzymol. 2005, 401, 9–42. [Google Scholar] [CrossRef]
- Savin, M.; Bumbasirevic, V.; Djukanovic, L.J.; Petronić, V. The significance of apoptosis for early diagnosis of Balkan nephropathy. Nephrol. Dial. Transplant. 2001, 16, 30–32. [Google Scholar] [CrossRef]
- Belicza, M.; Dzombeta, T.; Stanić, G.; Tomić, K.; Lenicek, T.; Perić-Balja, M.; Vukelić, M.; Jakovina. T., *REPLACE*; Jakovina, K.; Kruslin, B. Higher apoptotic cell rate in Balkan endemic nephropathy—stereologic analysis. Acta Clin. Croat. 2011, 50, 45–50. [Google Scholar]
- Adnan, H.; Antenos, M.; Kirby, G.M. The effect of menadione on glutathione S-transferase A1 (GSTA1): c-Jun N-terminal kinase (JNK) complex dissociation in human colonic adenocarcinoma Caco-2 cells. Toxicol. Lett. 2012, 214, 53–62. [Google Scholar] [CrossRef]
- Romero, L.; Andrews, K.; Ng, L.; O’Rourke, K.; Maslen, A.; Kirby, G. Human GSTA1-1 reduces c-Jun N-terminal kinase signalling and apoptosis in Caco-2 cells. Biochem. J. 2006, 400, 135–141. [Google Scholar] [CrossRef]
- Stefanovic, V.; Djukanović, L.; Cukuranović, R.; Bukvić, D.; Ležaić, V.; Marić, I.; Ogrizovic, S.S.; Jovanović, I.; Vlahovic, P.; Pešić, I.; et al. β2-microglobulin and α1-microglobulin as markers of Balkan endemic nephropathy, a worldwide disease. Ren. Fail. 2011, 33, 176–183. [Google Scholar] [CrossRef]
- Ping, J.; Wang, H.; Huang, M.; Liu, Z.S. Genetic analysis of glutathione S-transferase A1 polymorphism in the Chinese population and the influence of genotype on enzymatic properties. Toxicol. Sci. 2006, 89, 438–443. [Google Scholar] [CrossRef]
- Abdel-Rahman, S.Z.; El-Zein, R.A.; Anwar, W.A.; Au, W.W. A muptiplex PCR procedure for polymorphic analysis of GSTM1 and GSTT1 genes in population studies. Cancer Lett. 1996, 107, 229–233. [Google Scholar] [CrossRef]
- Davies, M.H.; Elias, E.; Acharya, S.; Cotton, W.; Faulder, G.C.; Fryer, A.A.; Strange, R.C. GSTM1 null polymorphism at the glutathione S-transferase M1 locus: Phenotype and genotype studies in patients with primary biliary cirrhosis. Gut 1993, 34, 549–553. [Google Scholar] [CrossRef]
- Harries, L.W.; Stubbins, M.J.; Forman, D.; Howard, G.C.; Wolf, C.R. Identification of genetic polymorphisms at the glutathione S-transferase Pi locus and association with susceptibility to bladder, testicular and prostate cancer. Carcinogenesis 1997, 18, 641–644. [Google Scholar] [CrossRef]
- Karpusas, M.; Axarli, I.; Chiniadis, L.; Papakyriakou, A.; Bethanis, K.; Scopelitou, K.; Clonis, Y.D.; Labrou, N.E. The Interaction of the Chemotherapeutic Drug Chlorambucil with Human Glutathione Transferase A1-1: Kinetic and Structural Analysis. PLoS One 2013, 8, e56337. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Reljic, Z.; Zlatovic, M.; Savic-Radojevic, A.; Pekmezovic, T.; Djukanovic, L.; Matic, M.; Pljesa-Ercegovac, M.; Mimic-Oka, J.; Opsenica, D.; Simic, T. Is Increased Susceptibility to Balkan Endemic Nephropathy in Carriers of Common GSTA1 (*A/*B) Polymorphism Linked with the Catalytic Role of GSTA1 in Ochratoxin A Biotransformation? Serbian Case Control Study and In Silico Analysis. Toxins 2014, 6, 2348-2362. https://doi.org/10.3390/toxins6082348
Reljic Z, Zlatovic M, Savic-Radojevic A, Pekmezovic T, Djukanovic L, Matic M, Pljesa-Ercegovac M, Mimic-Oka J, Opsenica D, Simic T. Is Increased Susceptibility to Balkan Endemic Nephropathy in Carriers of Common GSTA1 (*A/*B) Polymorphism Linked with the Catalytic Role of GSTA1 in Ochratoxin A Biotransformation? Serbian Case Control Study and In Silico Analysis. Toxins. 2014; 6(8):2348-2362. https://doi.org/10.3390/toxins6082348
Chicago/Turabian StyleReljic, Zorica, Mario Zlatovic, Ana Savic-Radojevic, Tatjana Pekmezovic, Ljubica Djukanovic, Marija Matic, Marija Pljesa-Ercegovac, Jasmina Mimic-Oka, Dejan Opsenica, and Tatjana Simic. 2014. "Is Increased Susceptibility to Balkan Endemic Nephropathy in Carriers of Common GSTA1 (*A/*B) Polymorphism Linked with the Catalytic Role of GSTA1 in Ochratoxin A Biotransformation? Serbian Case Control Study and In Silico Analysis" Toxins 6, no. 8: 2348-2362. https://doi.org/10.3390/toxins6082348
APA StyleReljic, Z., Zlatovic, M., Savic-Radojevic, A., Pekmezovic, T., Djukanovic, L., Matic, M., Pljesa-Ercegovac, M., Mimic-Oka, J., Opsenica, D., & Simic, T. (2014). Is Increased Susceptibility to Balkan Endemic Nephropathy in Carriers of Common GSTA1 (*A/*B) Polymorphism Linked with the Catalytic Role of GSTA1 in Ochratoxin A Biotransformation? Serbian Case Control Study and In Silico Analysis. Toxins, 6(8), 2348-2362. https://doi.org/10.3390/toxins6082348