Modes of Action of Microbially-Produced Phytotoxins


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
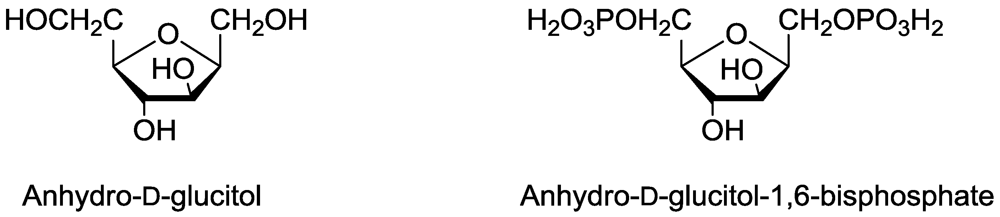{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Amino Acid Metabolism
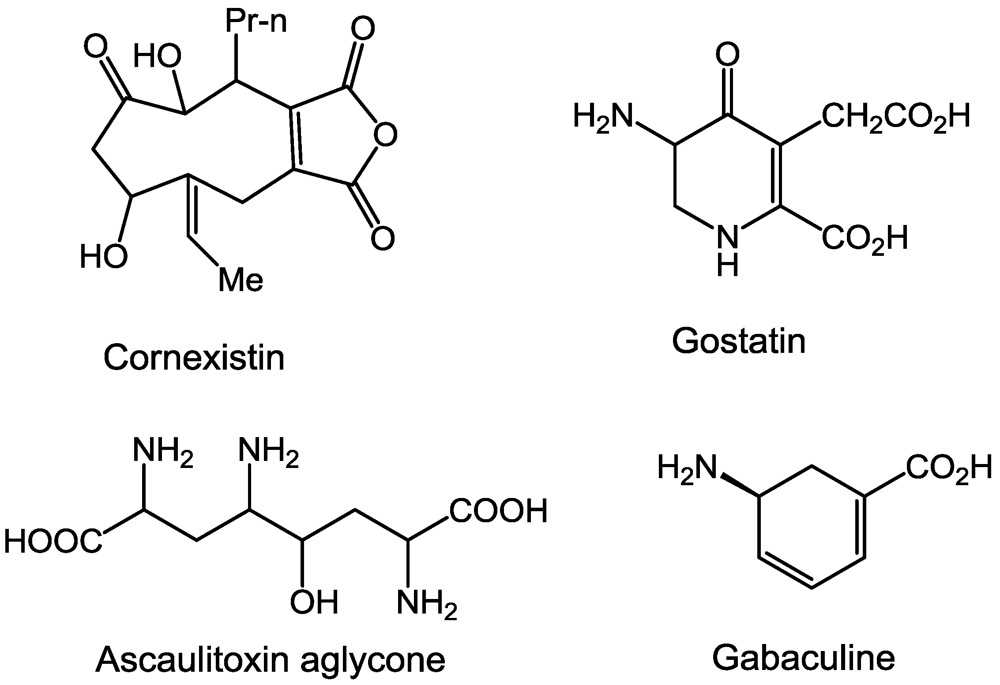
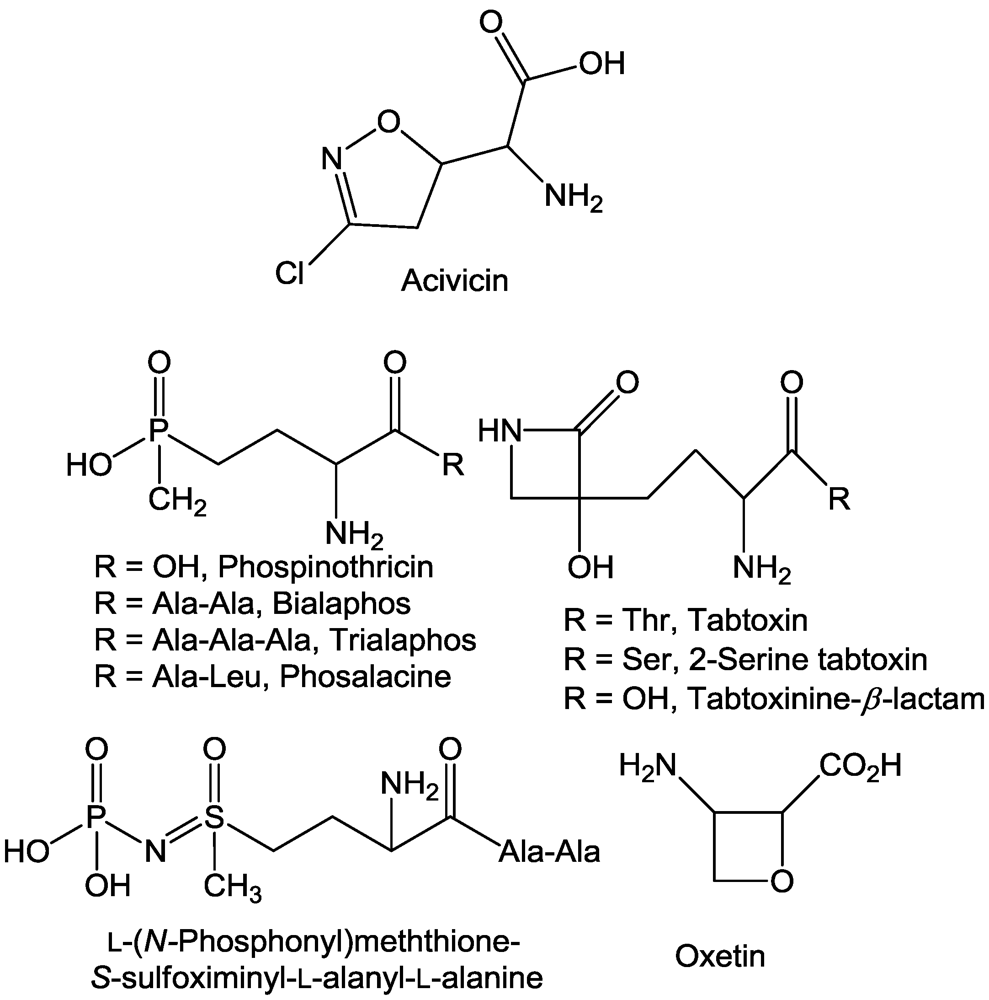
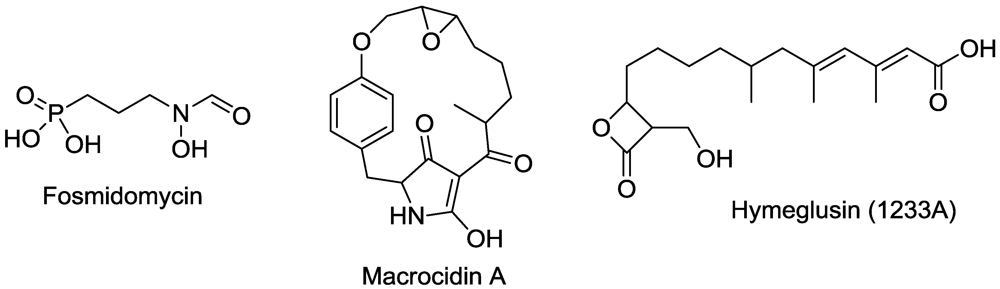
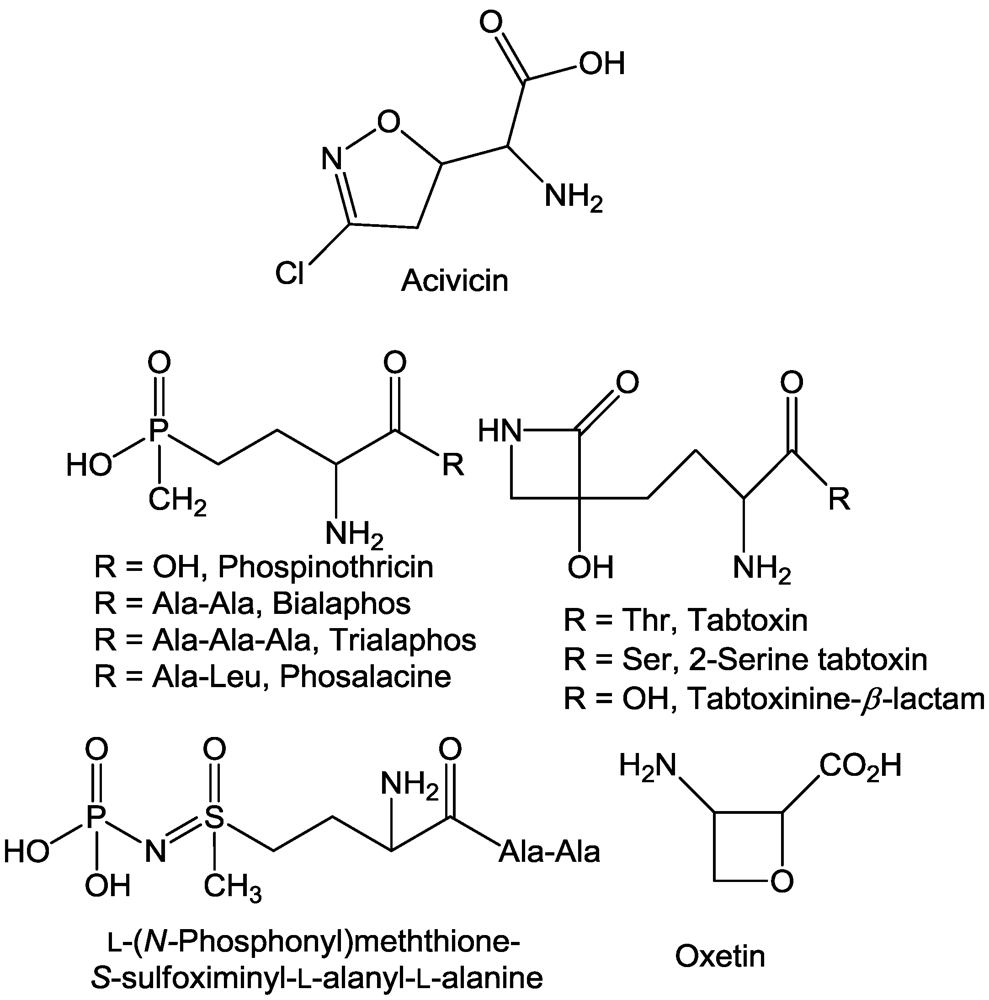
2.1. Aminotransferases
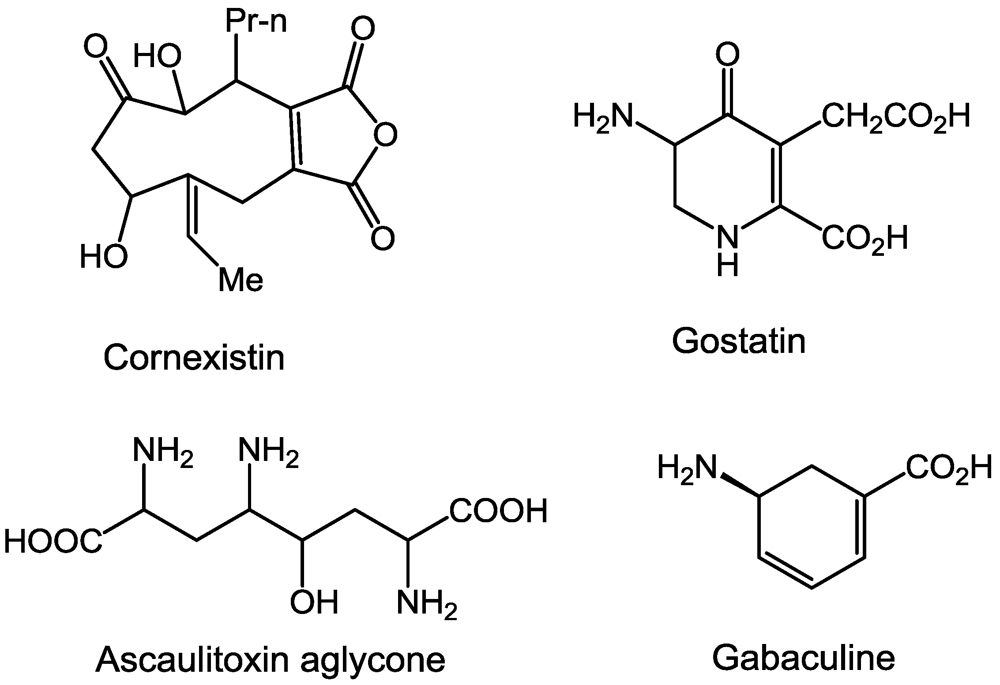
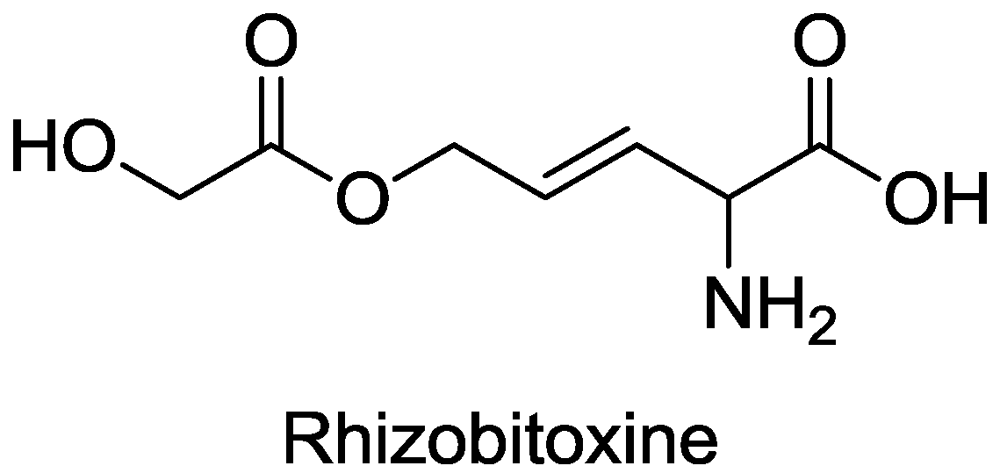
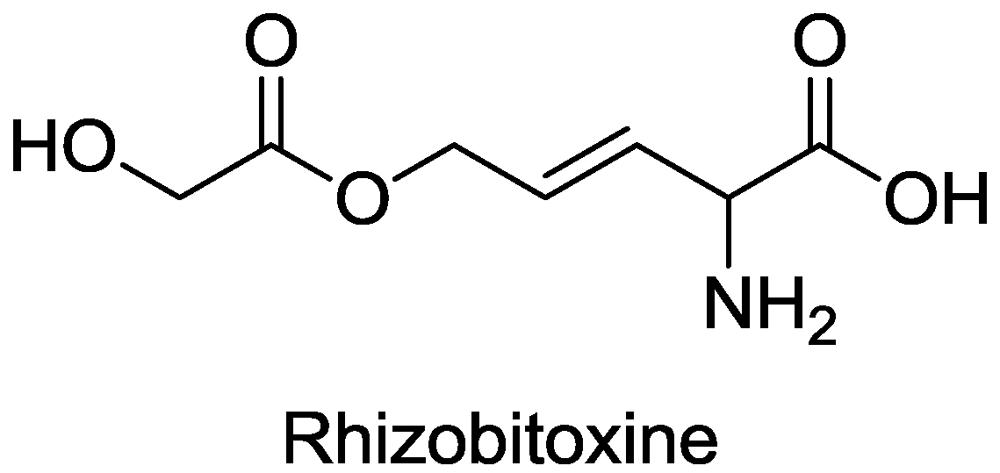
2.2. β-Cystathionase
2.3. Glutamate Synthase
2.4. Glutamine Synthetase
2.5. Ornithine Transcarboxylase
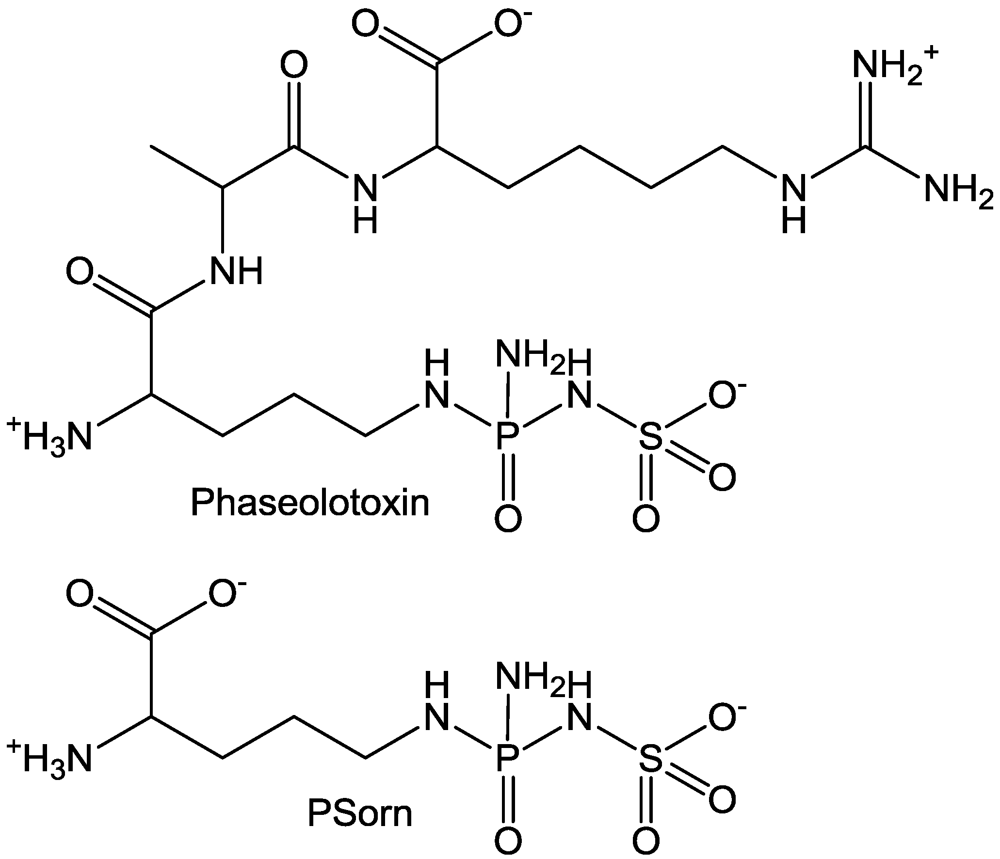
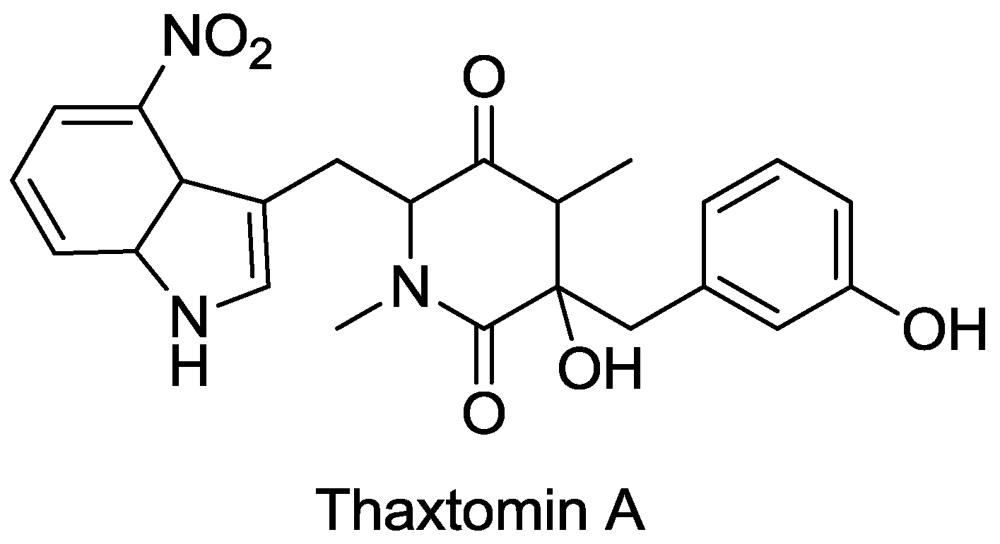
3. Cellulose Synthesis
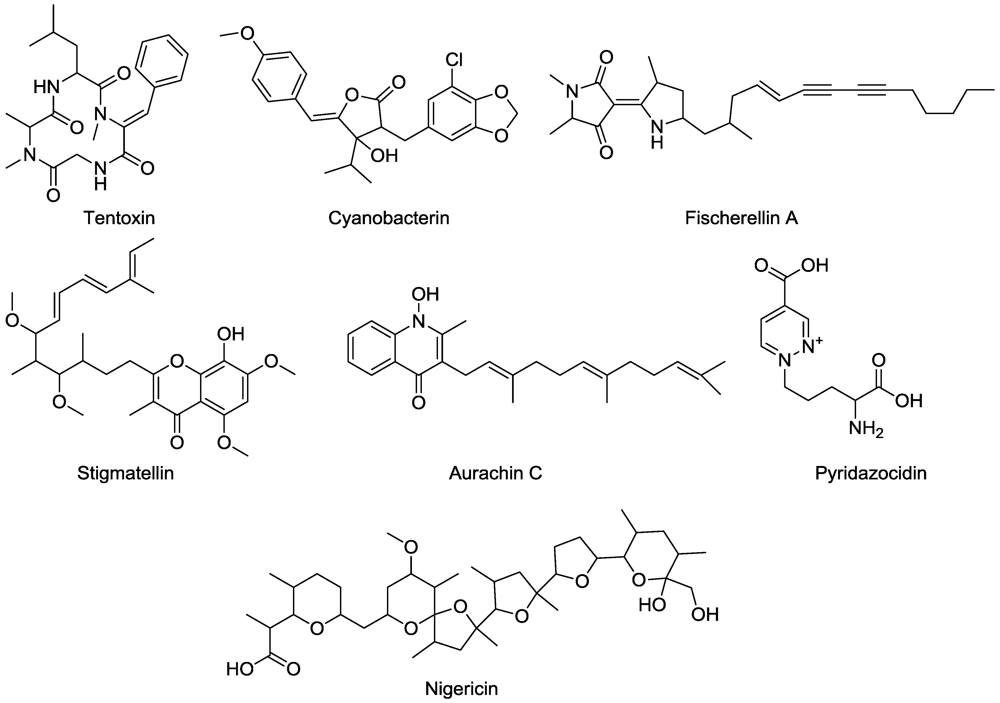
4. Energy Transfer
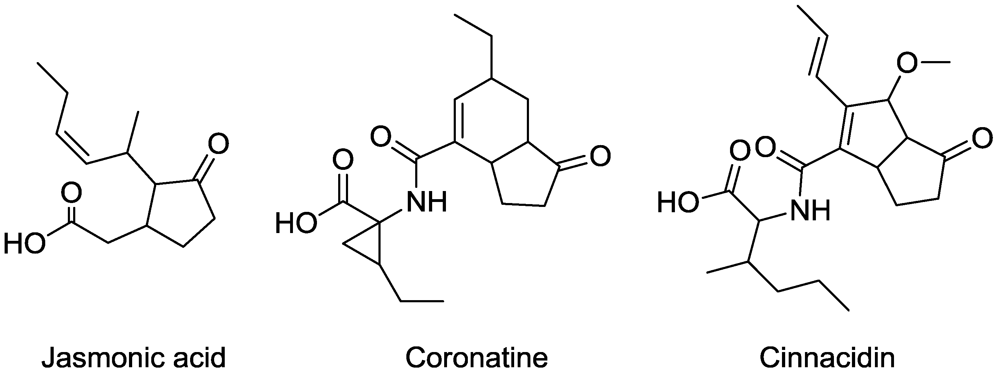
5. Jasmonic Acid Analogues
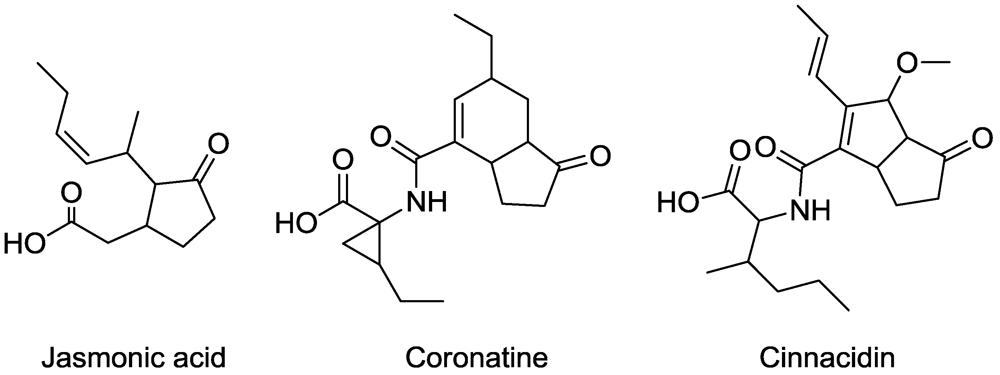
6. Lipid Metabolism
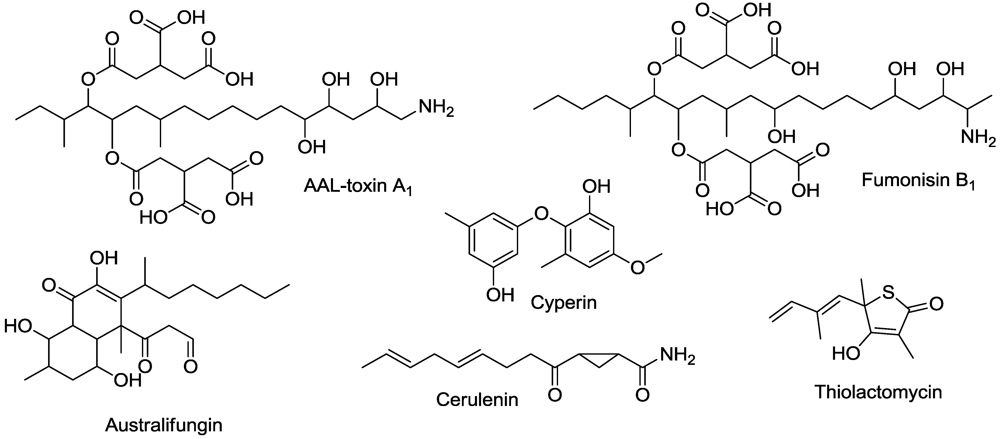
7. Membrane Function
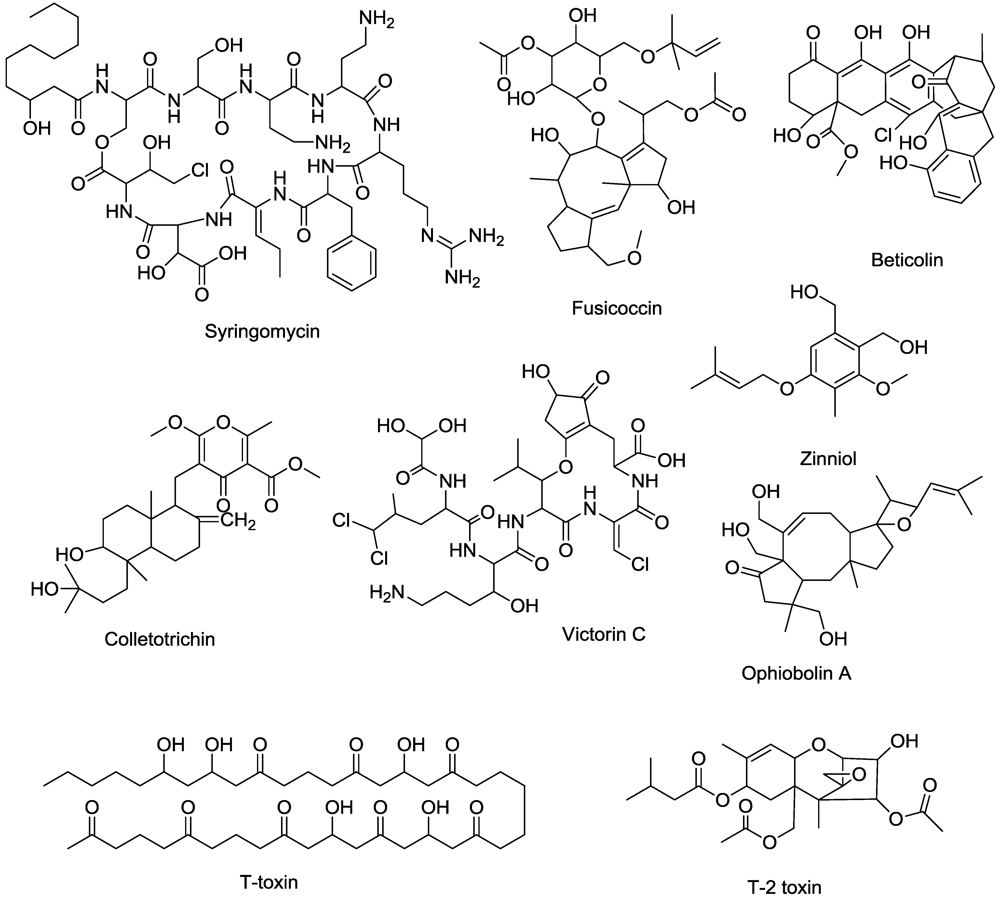
8. Mitotic Disruptors
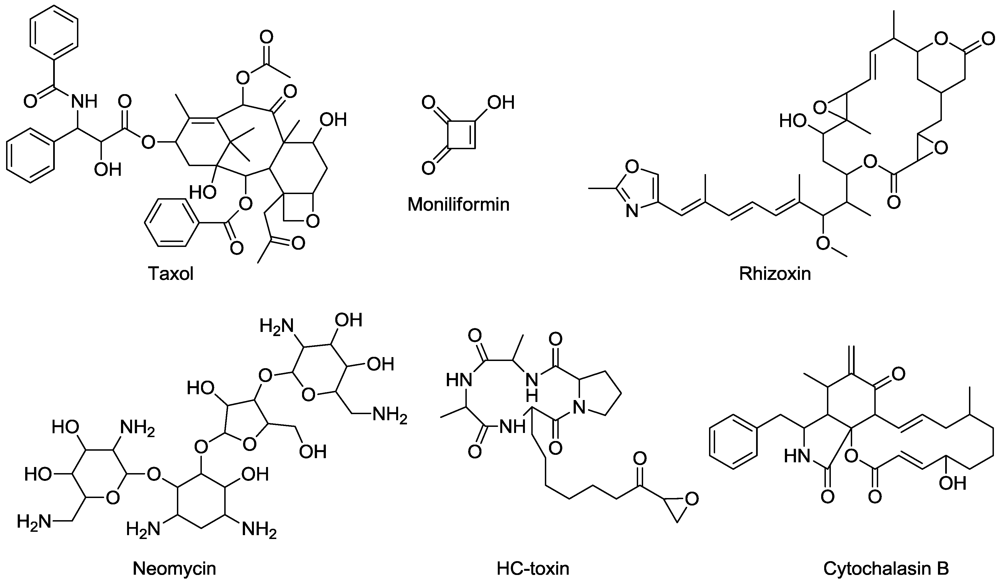
9. Nucleic Acid Synthesis
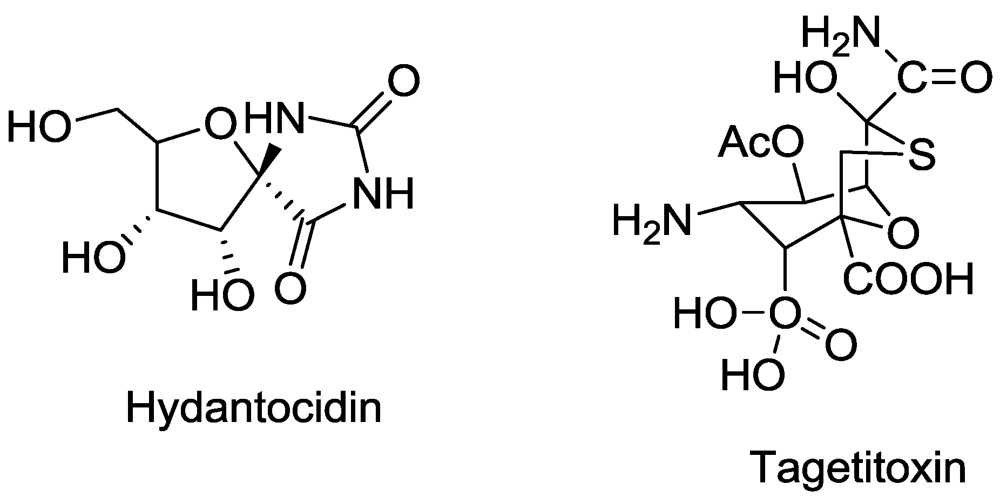
10. Photodynamic Compounds

11. Porphyrin Synthesis
12. Protein Synthesis
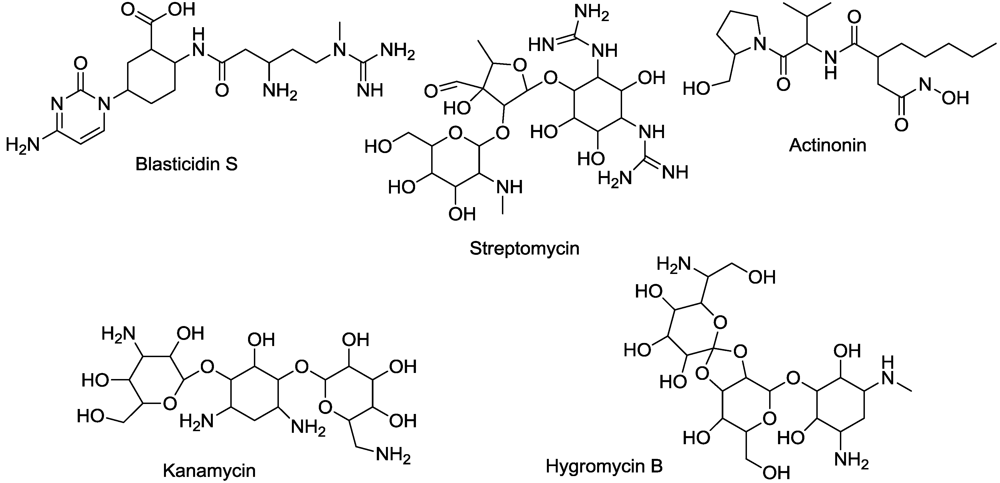
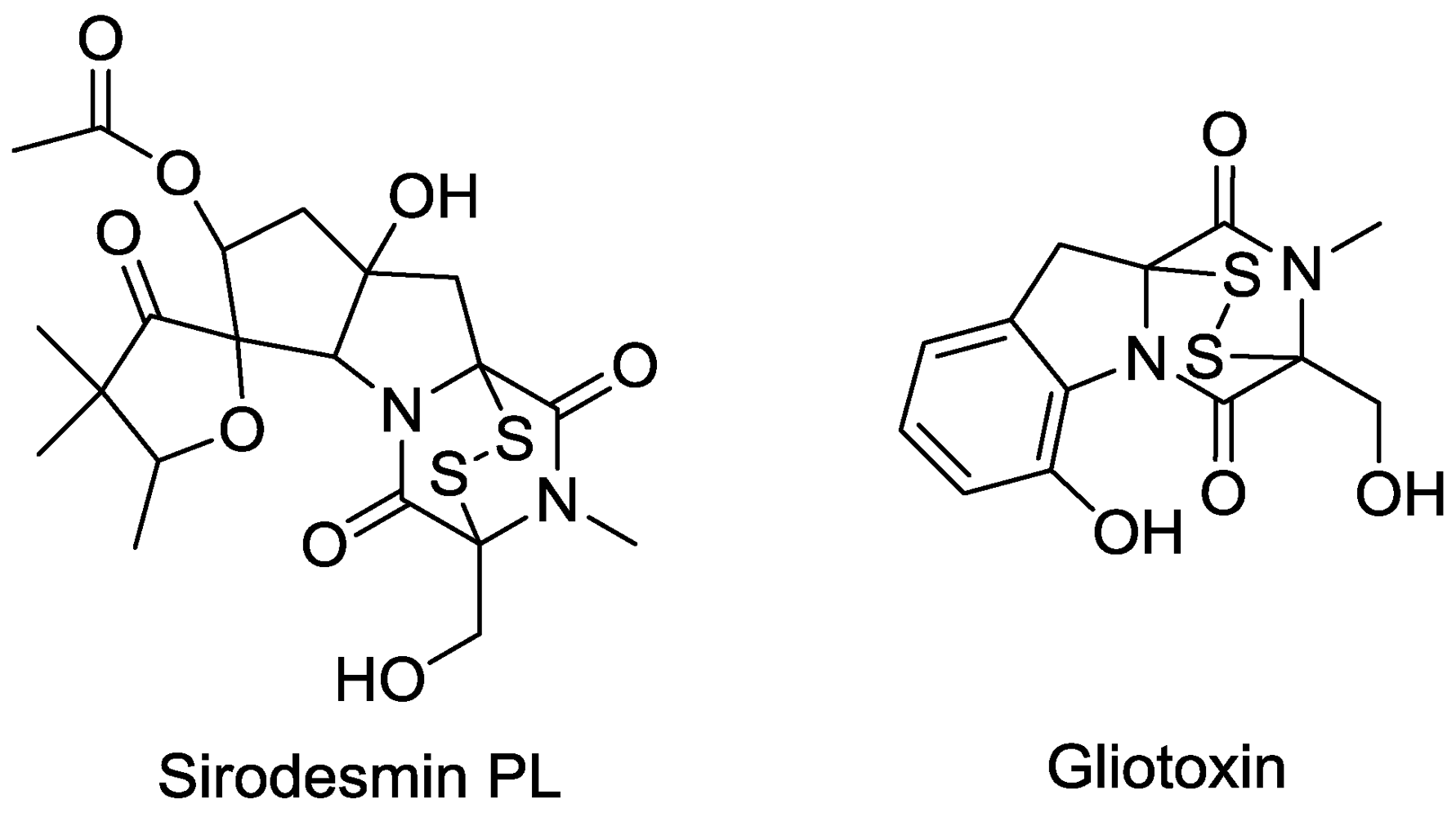
13. Protein Binding
14. Sugar Metabolism
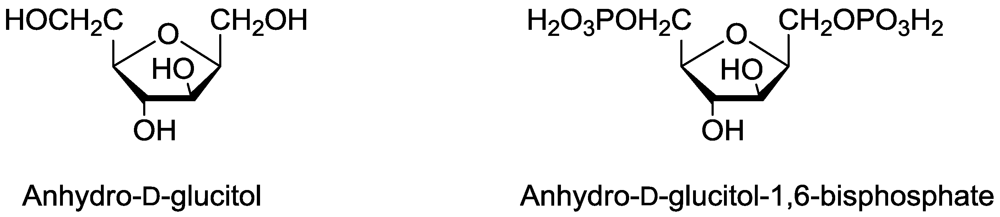
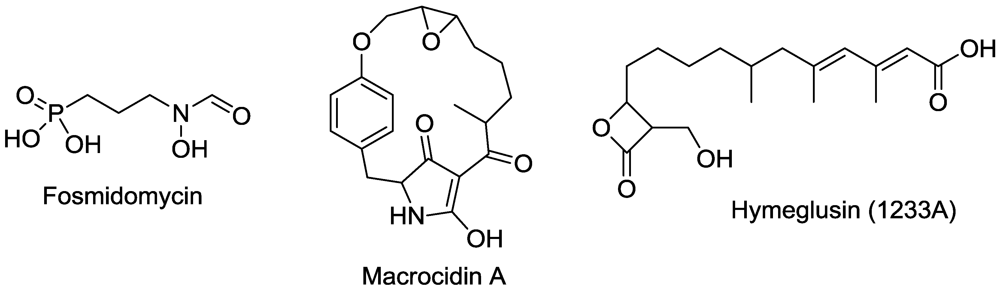
15. Terpenoid Synthesis
16. Conclusions
References
- Duke, S.O.; Abbas, H.K.; Amagasa, T.; Tanaka, T. Phytotoxins of microbial origin with potential for use as herbicides. In Crop Protection Agents from Nature: Natural Products and Analogues.
- Critical Reviews on Applied Chemistry; Copping, L.G. (Ed.) Society for Chemical Industries: Cambridge, UK, 1996; Volume 35, pp. 82–113.
- Berestetskiy, A.O. A review of fungal phytotoxins: From basic studies to practical use. Appl. Biochem. Microbiol. 2008, 44, 453–465. [Google Scholar]
- Möbius, N.; Hertwerk, C. Fungal phytotoxins as mediators of virulence. Curr. Opin. Plant Biol. 2009, 12, 390–398. [Google Scholar]
- Strange, R.N. Phytotoxins produced by plant pathogens. Nat. Prod. Rep. 2007, 24, 127–144. [Google Scholar]
- Mallik, M.A.B. Selective isolation and screening of soil microorganisms for metabolites with herbicidal potential. J. Crop Prot. 2001, 4, 219–236. [Google Scholar]
- Abbas, H.K.; Duke, S.O. Phytotoxins from plant pathogens as potential herbicides. Toxin Rev. 1995, 14, 523–543. [Google Scholar]
- Cutler, H.G. Microbial natural products that affect plants, phytopathogens, and certain other microorganisms. Crit. Rev. Plant Sci. 1995, 14, 413–444. [Google Scholar]
- Evidente, A. Chemical and biological characterization of toxins produced by weed pathogenic fungi as potential natural herbicides. ACS Symp. Ser. 2006, 927, 62–75. [Google Scholar]
- Strobel, G.; Kenfield, D.; Bunkers, G.; Sugawara, F.; Clardy, J. Phytotoxins as potential herbicides. Experientia 1991, 47, 819–826. [Google Scholar]
- Barazani, O.; Friedman, J. Allelopathic bacteria and their impact on higher plants. Crit. Rev. Microbiol. 2001, 27, 41–55. [Google Scholar]
- Schinko, E.; Schad, K.; Eys, S.; Keller, U.; Wohlleben, W. Phosphinothricin-tripeptide biosynthesis: An original version of bacterial secondary metabolism. Phytochemisry 2009, 70, 1787–1800. [Google Scholar]
- Cutler, H.G.; Cutler, S.J.; Matesic, D. Mode of action of phytotoxic fungal metabolites. In Allelopathy: Chemistry and Mode of Action of Allelochemicals; Macías, F.A., Galindo, J.C.G., Molinillo, J.M.G., Cutler, H.G., Eds.; CRC Press: Boca Raton, FL, USA, 2004; pp. 253–270. [Google Scholar]
- Amagasa, T.; Paul, R.N.; Heitholt, J.J.; Duke, S.O. Physiological effects of cornexistin on Lemna pauscicostata. Pestic. Biochem. Physiol. 1994, 49, 37–52. [Google Scholar] [CrossRef]
- Nishino, T.; Murao, S. Isolation and some properties of an aspartate amino transferase inhibitor. Agric. Biol. Chem. 1983, 47, 1961–1966. [Google Scholar]
- Nishino, T.; Murao, S.; Wada, H. Mechanism of inactivation of pyridoxal phosphate-linked aspartate transaminase by gostatin. J. Biochem. 1984, 95, 1283–1288. [Google Scholar]
- Rando, R.R. Mechanism of the irreversible inhibition of gamma-aminobutyric acid-alpha-ketoglutaric acid transaminase by the neurotoxin gabaculin. Biochemistry 1977, 16, 4604–4610. [Google Scholar]
- Rajaram, V.; Prasuna, P.R.; Savithri, H.S.; Murthy, M.R.N. Structure of biosynthetic N-acetylornithine aminotransferase from Salmonella typhimurium: studies on substrate specificity and inhibitor binding. Proteins 2008, 70, 429–441. [Google Scholar] [PubMed]
- Rosellini, D.; Capamaccio, S.; Ferradini, N.; Savo Sardaro, M.L.; Nicolia, A.; Veronesi, F. Non-antibiotic, efficient selection for alfalfa genetic engineering. Plant Cell Rep. 2007, 26, 1035–1044. [Google Scholar]
- Evidente, A.; Capasso, R.; Cutignano, A.; Taglialatela-Scafati, O.; Vurro, M.; Zonno, M.C.; Motta, A. Ascaulitoxin, a phytotoxic bis-amino acid N-glucoside from Ascochyta caulina. Phytochemistry 1998, 48, 1131–1137. [Google Scholar]
- Duke, S.O.; Evidente, A.; Fiore, M.; Rimando, A.M.; Vurro, M.; Chistiansen, N.; Looser, R.; Grossmann, K. Effects of the aglycone of ascaulitoxin on amino acid metabolism in Lemna paucicostata. Pestic. Biochem. Physiol. 2011, 100, 41–50. [Google Scholar] [CrossRef]
- Owens, L.D.; Guggenheim, S.; Hilton, J.L. Rhizobium-synthesized phytotoxin: And inhibitor of β-cystathionase in Salmonella typhimurium. Biochim. Biophys. Acta 1968, 158, 219–225. [Google Scholar] [PubMed]
- Giovanelli, J.; Owens, L.D.; Mudd, S.H. β-cystathionase. In vivo inactivation by rhizobitoxine and role of the enzyme in methionine biosynthesis in corn seedlings. Plant Physiol. 1973, 51, 492–503. [Google Scholar] [CrossRef] [PubMed]
- Owens, L.D. Herbicidal potential of rhizobitoxine. Weed Sci. 1973, 21, 63–66. [Google Scholar]
- Owens, L.D.; Lieberman, M.; Kunishi, A. Inhibition of ethylene production by rhizobitoxine. Plant Physiol. 1971, 48, 1–4. [Google Scholar]
- Yasuta, T.; Satoh, S.; Minamisawa, K. New assay for rhizobitoxine based on inhibition of 1-aminocyclopropane-1-carboxylate synthase. Appl. Environ. Microbiol. 1999, 65, 849–852. [Google Scholar]
- Gould, S.J.; Ju, S. The biosynthesis of acividin and 4-hydroxyacivicin from Nδ-hydroxyornithine. J. Am. Chem. Soc. 1989, 111, 2329–2331. [Google Scholar]
- Brunner, H.G.; Chemla, P.; Dobler, M.R.; O’Sullivan, A.C.; Pachlatko, P.; Pillonel, C.; Stierli, D. Fungicidal properties of acivicin and its derivatives. Am. Chem. Soc. Ser. 2007, 948, 121–135. [Google Scholar]
- Conti, P.; Roda, G.; Stabile, H.; Vanoni, M.A.; Curti, B.; De Amici, M. Synthesis and biological evaluation of new amino acids structurally related to the antitumor agent acivicin. Farmaco 2003, 58, 683–690. [Google Scholar]
- Natsumeda, Y.; Ikegami, T.; Olah, E.; Weber, G. Significance of purine salvage in circumventing the action of antimetabolites in rat hepatoma cells. Cancer Res. 1989, 49, 88–92. [Google Scholar]
- Rodriguez-Suerez, R.; Xu, D.; Veillette, K.; Davison, J.; Sillaots, S.; Kauffman, S.; Hu, W.; Bowman, H.; Martel, N.; Trosok, S.; et al. Mechanism-of-action determination of GMP synthase inhibitors and target validation in Candida albicans and Aspergillis fumigates. Chem. Biol. 2007, 14, 1163–1175. [Google Scholar] [CrossRef] [PubMed]
- Williams, K.; Cullati, S.; Sand, A.; Biterova, E.I.; Barycki, J.J. Crystal Structure of acivicin-inhibited γ-glutamyltranspeptidase reveals critical roles for its C-terminus in autoprocessing and catalysis. Biochemistry 2009, 48, 2459–2467. [Google Scholar] [PubMed]
- Lydon, J.; Duke, S.O. Inhibitors of Glutamine Biosynthesis. In Plant Amino Acids: Biochemistry and Biotechnology; Singh, B.K., Ed.; Marcel Dekker: New York, NY, USA, 1998; pp. 445–464. [Google Scholar]
- Wild, A.; Ziegler, C. The effect of bialaphos and ammonium on ammonium-assimilation and photosynthesis. I. Effect on the enzymes of ammonium-assimilation. Z. Naturforsch. 1989, 44, 97–102. [Google Scholar]
- Ziegler, C.; Wild, A. The effect of bialaphos and ammonium on ammonium-assimilation and photosynthesis. II. Effect on photosynthesis and photorespiration. Z. Naturforsch. 1989, 44, 103–108. [Google Scholar]
- Kato, H.; Nagayama, K.; Abe, H.; Kobayashi, R.; Ishihara, E. Isolation, structure and biological activity of trialaphos. Agric. Biol. Chem. 1991, 55, 1133–1134. [Google Scholar]
- Ōmura, S.; Murata, M.; Hanaki, H.; Hinotozawa, T.; Ōiwa, R.; Tanaka, H. Phosalacine, a new herbicidal antibiotic containing phosphinothricin. Fermentation, isolation, biological activity and mechanism of action. J. Antibiot. 1984, 37, 829–835. [Google Scholar] [PubMed]
- Copping, L.; Duke, S.O. Natural products that have been used commercially as crop protection agents. A review. Pest Manag. Sci. 2007, 63, 524–554. [Google Scholar] [CrossRef] [PubMed]
- Ōmura, S.; Murata, M.; Imamura, N.; Iwai, Y.; Tanaka, H.; Furusaki, A.; Matsumoto, T. Oxetin, a new antimetabolite from and ascomycete. Fermentation, isolation, structure and biological activity. J. Antibiot. 1984, 37, 1324–1332. [Google Scholar] [PubMed]
- Preuss, D.L.; Scannell, J.P.; Ax, H.A.; Kellett, M.; Weiss, F.; Demny, T.C.; Stempel, A. Antimetabolites produced by microorganisms. VII. L-(N5-phosphono)methionine-S-sulfoximinyl-L-alanyl-L-alanine. J. Antibiot. 1973, 26, 261–266. [Google Scholar] [PubMed]
- Uchytil, T.F.; Durbin, R.D. Hydrolysis of tabtoxins by plant and bacterial enzymes. Experientia 1980, 36, 301–302. [Google Scholar]
- Lanston-Unkefer, P.L.; Robinson, A.C.; Knight, T.J.; Durbin, R.D. Inactivation of pea seed glutamine synthetase by tabtoxinine-β-lactam. J. Biol. Chem. 1987, 262, 1608–1613. [Google Scholar]
- Taylor, P.A.; Schnoes, H.K.; Durbin, R.D. Characterization of chlorosis-inducing toxins from a plant pathogenic Pseudomonas species. Biochim. Biophys. Acta 1972, 286, 107–117. [Google Scholar] [PubMed]
- Sato, Y.; Watabe, H.O.; Ueno, S.; Miyadoh, S.; Koyama, M. SF2448 A, B, and C, new herbicidal peptide antibiotics related to tabtoxin. Sci. Rep. Meiji Seika Kaisha 1990, 29, 14–20. [Google Scholar]
- Templeton, M.D.; Reinhardt, L.A.; Collyer, C.A.; Mitchell, R.E.; Cleland, W.W. Kinetic analysis of the L-ornithine transcarbamolylase from Pseudomonas savastanoi pv. phaseolicola that is resistant to the transition state analogue (R)-Nδ-(N'-sulfodiaminophosphinyl)-L-ornithine. Biochemistry 2005, 44, 4408–4415. [Google Scholar] [PubMed]
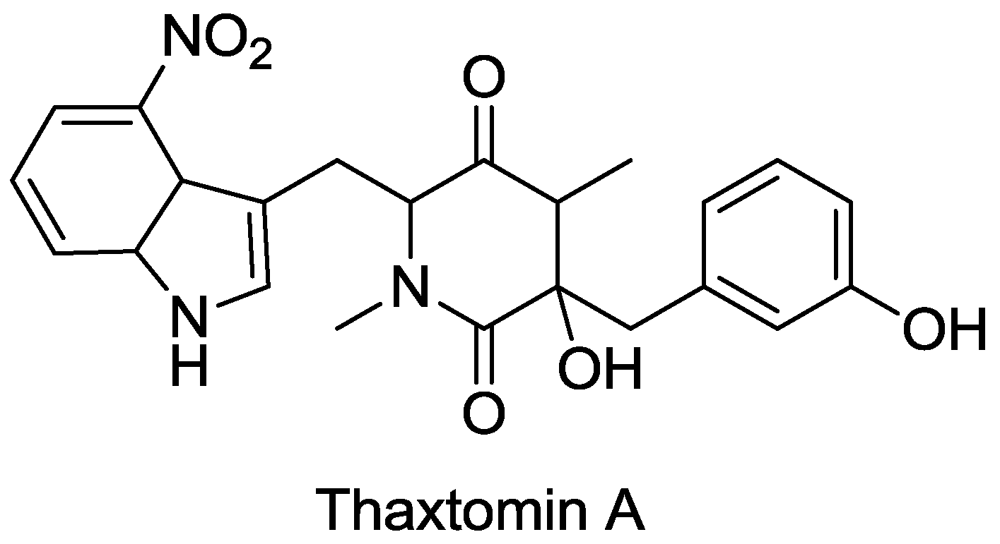
- King, R.R.; Calhoun, L.A. The thaxtomin phytotoxins: sources, synthesis, biosynthesis, biotransformation and biological activity. Phytochemistry 2009, 70, 833–841. [Google Scholar] [CrossRef] [PubMed]
- Bischoff, V.; Cookson, S.J.; Wu, S.; Scheible, W.-R. Thaxtomin A affects CESA-complex density, expression of cell wall genes, cell wall composition, and causes ectopic lignification in Arabidopsis thaliana seedlings. J. Exp. Bot. 2009, 60, 955–965. [Google Scholar] [CrossRef] [PubMed]
- Halloin, J.M.; De Zoeten, G.A.; Gaard, G.R.; Walker, J.C. Effects of tentoxin on chlorophyll synthesis and plastid structure in cucumber and cabbage. Plant Physiol. 1970, 45, 310–314. [Google Scholar] [CrossRef] [PubMed]
- Duke, S.O.; Wickliff, J.L.; Vaughn, K.C.; Paul, R.N. Tentoxin does not cause chlorosis in greening mung bean leaves by inhibiting photophosphorylation. Physiol. Plant. 1982, 56, 387–398. [Google Scholar]
- Selman, B.R.; Durbin, R.D. Evidence for a catalytic function of the coupling factor 1 protein reconstituted with chloroplast thylakoid membranes. Biochim. Biophys. Acta 1978, 502, 29–37. [Google Scholar]
- Reimer, S.; Selman, B.R. Tentoxin-induced energy-independent adenine nucleotide exchange and ATPase activity with chloroplast coupling factor 1. J. Biol. Chem. 1978, 253, 7249–7255. [Google Scholar]
- Vaughn, K.C.; Duke, S.O. Tentoxin stops the processing of polyphenol oxidase into an active enzyme. Physiol. Plant. 1984, 60, 257–261. [Google Scholar]
- Duke, S.O. Tentoxin effects on variable fluorescence and P515 electrochromic absorbance changes in tentoxin-sensitive and -resistant plant species. Plant Sci. 1993, 90, 119–126. [Google Scholar]
- Avni, A.; Anderson, J.D.; Hollan, N.; Rochaix, J.D.; Gromet-Elhanan, Z.; Edelman, M. Tentoxin sensitivity of chloroplasts determined by codon 83 of β subunit of proton ATPase. Science 1992, 257, 1245–1247. [Google Scholar]
- Holland, N.; Evron, Y.; Jansen, M.A.K.; Edelman, M.; Pick, U. Involvement of thylakoid overenergization in tentoxin-induced chlorosis in Nicotiana spp. Plant Physiol. 1997, 114, 887–892. [Google Scholar] [PubMed]
- Trebst, A.; Depka, B. Polyphenol oxidase and photosynthesis research. Photosynthesis Res. 1995, 46, 41–44. [Google Scholar]
- Vaughn, K.C.; Lax, A.R.; Duke, S.O. Polyphenol oxidase: The chloroplast oxidase with no established function. Physiol. Plant. 1988, 72, 659–665. [Google Scholar]
- Shavit, N.; San Pietro, A. K+-dependent uncoupling of photophosphorylation by nigericin. Biochim. Biophys. Acta 1967, 28, 277–283. [Google Scholar]
- Quick, P.; Scheibe, R.; Stitt, M. Use of tentoxin and nigericin to investigate the possible contribution of ΔpH to energy dissipation and the control of electron transport in spinach leaves. Biochim. Biophys. Acta 1989, 974, 282–288. [Google Scholar]
- Lee, E.-S.J.; Gleason, F.K. A second algicidal natural product from the cyanobacterium, Scytonema hofmanni. Plant Sci. 1994, 103, 155–160. [Google Scholar] [CrossRef]
- Srivastava, A.; Juttner, F.; Strasser, R.J. Action of the allelochemical, fischerellin A, on photosystem II. Biochim. Biophys. Acta Bioenerg. 1998, 1365, 326–336. [Google Scholar]
- Oettmeier, W.; Godde, D.; Kunze, B.; Hoefle, G. Stigmatellin. A dual type inhibitor of photosynthetic electron transport. Biochim. Biophys. Acta Bioenerg. 1985, 807, 216–219. [Google Scholar] [CrossRef]
- Oettmeier, W.; Dostatni, R.; Majewski, C.; Hoefle, G.; Fecker, T.; Kunze, B.; Reichenbach, H. The aurachins, naturally occurring inhibitors of photosynthetic electron flow through photosystem II and cytochrome b6/f-complex. Z. Naturforsch. 1990, 45C, 322–328. [Google Scholar]
- Gerwick, B.C.; Fields, S.S.; Graupner, P.R.; Gray, J.A.; Chapin, E.L.; Cleveland, J.A.; Heim, D.R. Pyridazocidin, a new microbial phytotoxin with activity in the Mehler reaction. Weed Sci. 1997, 45, 654–657. [Google Scholar]
- Ichihara, A.; Shiraishi, K.; Sato, H.; Sakamura, S.; Nishiyama, K.; Sakai, R.; Furusaki, A.; Matsumoto, T. The structure of coronatine. J. Am. Chem. Soc. 1977, 99, 636–637. [Google Scholar]
- Block, A.; Schmelz, E.; Jones, J.B.; Klee, H.J. Coronatine and salicylic acid: the battle between Arabidopsis and Pseudomonas for phytohorome control. Mol. Plant Pathol. 2005, 6, 79–83. [Google Scholar] [CrossRef] [PubMed]
- Irvine, N.M.; Yerkes, C.N.; Graupner, P.R.; Roberts, R.E.; Hahn, D.R.; Pearce, C.; Gerwick, B.C. Synthesis and characterization of synthetic analogs of cinnacidin, a novel phytotoxin from Nectria sp. Pest Manag. Sci. 2008, 64, 891–899. [Google Scholar] [CrossRef] [PubMed]
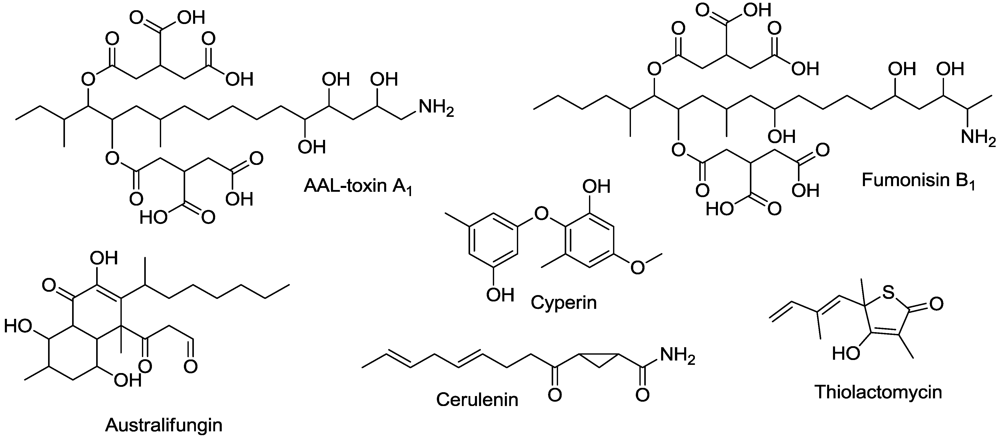
- Abbas, H.K.; Duke, S.O.; Tanaka, T. Phytotoxicity of fumonisins and related compounds. J. Toxicol.-Toxin Rev. 1993, 12, 225–251. [Google Scholar]
- Abbas, H.K.; Duke, S.O.; Shier, W.T.; Riley, R.T.; Kraus, G.A. The Chemistry and Biological Activities of the Natural Products AAL-Toxin and the Fumonisins. In Natural Toxins 2. Structure, Mechanism of Action, and Detection; Singh, B.R., Tu, A.T., Eds.; Plenum: New York, NY, USA, 1996; pp. 293–308. [Google Scholar]
- Abbas, H.K.; Duke, S.O.; Shier, W.T.; Duke, M.V. Inhibition of Ceramide Synthesis in Plants by Phytotoxins. In Advances in Microbial Toxin Research and Its Biotechnological Exploitation; Upadhyay, R.K., Ed.; Kluwer Academic/Plenum Pub.: London, UK, 2002; pp. 211–229. [Google Scholar]
- Abbas, H.K.; Duke, S.O.; Merrill, A.H.; Wang, E.; Shier, W.T. Phytotoxicity of australifungin, AAL-toxins and fumonisin B1 to Lemna pausicostata. Phytochemistry 1998, 47, 1509–1514. [Google Scholar]
- Abbas, H.K.; Tanaka, T.; Duke, S.O.; Porter, J.K.; Wray, E.M.; Hodges, L.; Sessions, A.E.; Wang, E.; Merrill, A.H.; Riley, R.T. Fumonisin and AAL-toxin-induced disruption of sphingolipid metabolism with accumulation of free sphingoid bases: Involvement in plant disease. Plant Physiol. 1994, 106, 1085–1093. [Google Scholar]
- Lincoln, J.E.; Richael, C.; Overduin, B.; Smith, K.; Bostock, R.; Gilchrist, D.G. Expression of the antiapoptotic baculovirus p35 gene in tomato blocks programmed cell death and provides broad-spectrum resistance to disease. Proc. Natl. Acad. Sci. USA 2002, 99, 15217–15221. [Google Scholar]
- Gechev, T.S.; Gadjev, I.Z.; Hille, J. An extensive microarray analysis of AAL-toxin-induced cell death in Arabidopsis thaliana brings new insights into the complexity of programmed cell death in plants. Cell. Mol. Life Sci. 2004, 61, 1185–1197. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Abbas, H.K.; Duke, S.O. Structure-dependent phytotoxicity of fumonisins and related compounds in a duckweed bioassay. Phytochemistry 1993, 33, 779–785. [Google Scholar]
- Shi, L.; Bielawski, J.; Mu, J.; Dong, H.; Teng, C.; Zhang, J.; Yang, X.; Tomishige, N.; Hanada, K.; Hannun, Y.A.; Zuo, J. Involvement of sphingoid bases in mediating reactive oxygen intermediate production and programmed cell death in Arabidopsis. Cell Res. 2007, 17, 1030–1040. [Google Scholar] [CrossRef] [PubMed]
- Brown, M.S.; Akopiants, K.; Resceck, D.M.; McArthur, H.A.I.; McCormick, E.; Reynolds, K.A. Biosynthetic origins of the natural product, thiolactomycin: A unique and selective inhibitor of type II dissociated fatty acid synthases. J. Am. Chem. Soc. 2003, 125, 10166–10167. [Google Scholar]
- Focke, M.; Feld, A.; Lichtenthaler, H.K. Effect of thiolactomycin on de novo fatty acid biosynthesis in plants. Z. Naturforsch. 1990, 45C, 518–520. [Google Scholar]
- Feld, A.; Kobek, K.; Lichtenthaler, H.K. Inhibition of de novo fatty-acid biosynthesis in isolated chloroplasts by different antibiotics and herbicides. Z. Naturforsch. 1989, 44C, 976–978. [Google Scholar]
- Feld, A.; Kobek, K.; Litchtenthaler, H.K. Inhibition of fatty-acid biosynthesis in isolated chloroplasts by the antibiotics cerulenin and thiolactomycin. Brighton Crop Prot. Conf. Weeds 1989, 2, 479–486. [Google Scholar]
- Weber, H.A.; Gloer, J.B. Interference competition among natural fungal competitors: An antifungal metabolite from the coprophilous fungus Preussia Fleischhakii. J. Nat. Prod. 1988, 51, 879–883. [Google Scholar] [CrossRef] [PubMed]
- Stierle, A.; Upadhyay, R.; Strobel, G. Cyperine, a phytotoxin produced by Ascochyta cypericola, a fungal pathogen of Cyperus rotundus. Phytochemistry 1992, 30, 2191–2192. [Google Scholar]
- Venkatasubbaiah, P.; van Dyke, C.G.; Chilton, W.S. Phytotoxic metabolites of Phoma sorghina, a new foliar pathogen of pokeweed. Mycologia 1992, 84, 715–723. [Google Scholar] [CrossRef]
- Dayan, F.E.; Ferreira, D.; Wang, Y.-H.; Khan, I.A.; McInroy, J.A.; Pan, Z. A pathogenic fungi diphenyl ether phytotoxin targets plant enoyl (acyl carrier protein) reductase. Plant Physiol. 2008, 147, 1062–1071. [Google Scholar]
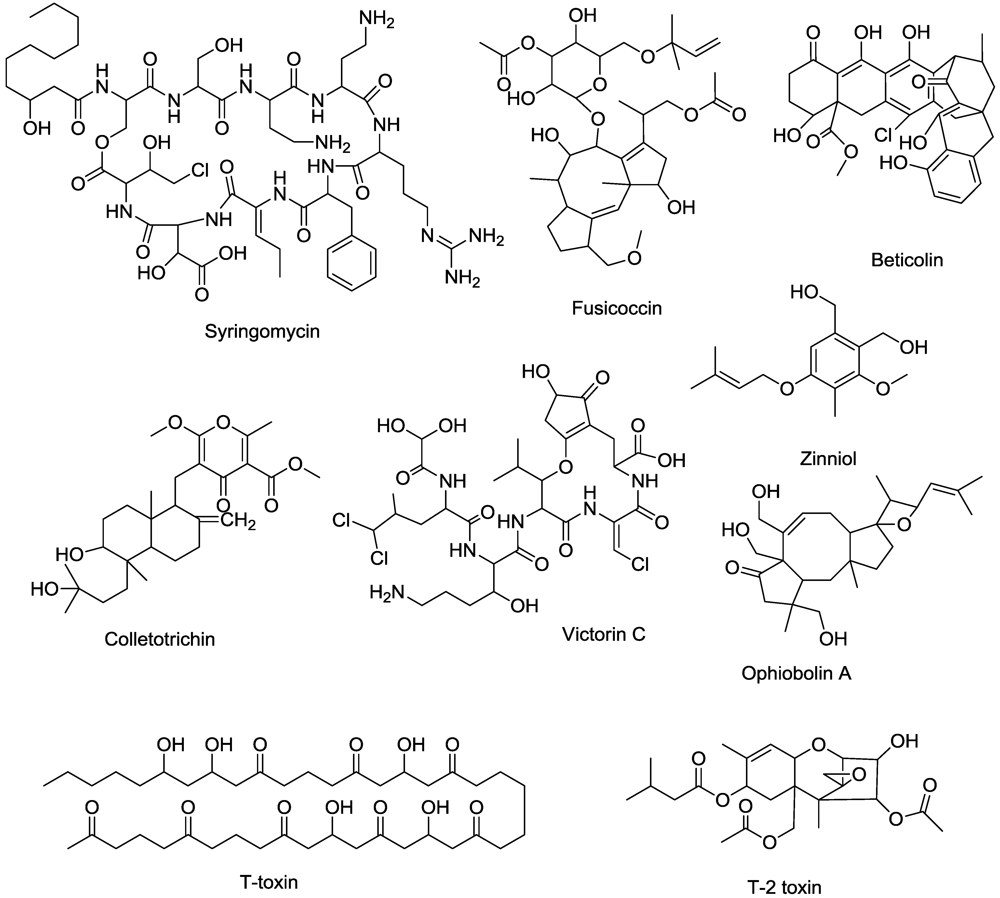
- Isogai, A.; Fukuchi, N.; Yamashita, S.; Suyama, K.; Suzuki, A. Structures of syringostatin A and syringostatin B, novel phytotoxins produced by Pseudomonas syringae pv. syringae isolated from lilac blights. Tetrahedron Lett. 1990, 31, 695–698. [Google Scholar]
- Ballio, A.; Bossa, F.; Collina, A.; Gallo, M.; Iacobellis, N.S.; Paci, M.; Pucci, P.; Scaloni, A.; Segre, A.; Simmaco, M. Structure of syringotoxin, a bioactive metabolite of Pseudomonas syringae pv. syringae. FEBS Lett. 1990, 269, 377–380. [Google Scholar] [CrossRef]
- Scaloni, A.; Bachmann, R.C.; Takemoto, J.Y.; Barra, D.; Simmaco, M.; Ballio, A. Stereochemical structure of syringomycin, a phytotoxic metabolite of Pseudomonas syringae pv. syringae. Nat. Prod. Lett. 1994, 4, 159–164. [Google Scholar] [CrossRef]
- Bender, C.L.; Alarcón-Chaidez, F.; Gross, D.C. Pseudomonas syringae phytotoxins: Mode of action, regulation, and biosynthesis by peptide and polyketide synthetases. Microbiol. Mol. Biol. Rev. 1999, 63, 266–292. [Google Scholar] [PubMed]
- Goudet, C.; Very, A.A.; Milat, M.L.; Ildefonse, M.; Thibaud, J.B.; Sentenac, H.; Blein, J.P. Magnesium ions promote assembly of channel-like structures from beticolin 0, a non-peptide fungal toxin purified from Cercospora beticola. Plant J. 1998, 14, 359–364. [Google Scholar] [CrossRef] [PubMed]
- Goudet, C.; Benitah, J.P.; Milat, M.L.; Sentenac, H.; Thibaud, J.B. Cluster organization and pore structure of ion channels formed by beticolin 3, a nonpeptidic fungal toxin. Biophys. J. 1999, 77, 3052–3059. [Google Scholar]
- Tugeon, B.; Baker, S.E. Genetic and genomic dissection of the Cochliobolus heterostrophus Tox 1 locus controlling biosynthesis of the polyketide virulence factor T-toxin. Mol. Plant Microbe Interact. 2006, 19, 219–261. [Google Scholar]
- Levings, S.S.; Rhoads, D.M.; Siedow, J.N. Molecular interactions of Bipolaris maydis T-toxin and maize. Can. J. Bot. 1995, 73, S484–S489. [Google Scholar]
- Aducci, P.; Marra, M.; Fogliano, V.; Fullone, M.R. Fusicoccin receptors: perception and transduction of the fusicoccin signal. J. Exp. Bot. 1995, 46, 1463–1478. [Google Scholar]
- Gomarasca, S.; Vannini, C.; Venegoni, A.; Talarico, A.; Marre, T.; Soave, C. A mutant of Arabidopsis thaliana with a reduced response to fusicoccin. Plant Physiol. 1993, 103, 165–170. [Google Scholar]
- Curtis, M.J.; Wolpert, T.J. The victorin-induced mitochondrial permeability transition precedes cell shrinkage and biochemical markers of cell death, and shrinkage occurs without loss of membrane integrity. Plant J. 2004, 38, 244–259. [Google Scholar]
- Tada, Y.; Yasuomi, K.; Koh, B.; Betsuyaku, S.; Shinogi, T.; Sakamoto, M.; Ohura, Y.; Hata, S.; Mori, Y.; Tosa, Y.; Mayama, S. Victorin triggers programmed cell death and the defense response via interaction with a cell surface mediator. Plant Cell Physiol. 2005, 46, 1787–1798. [Google Scholar]
- Gohbara, M.; Kosuge, Y.; Yamaaki, S.; Kimura, Y.; Suzuki, A.; Tamura, S. Isolation, structures and biological activities of colletotrichins, phytototoxic substances from Colletotrichum nicotianae. Agric. Biol. Chem. 1978, 42, 1037–1043. [Google Scholar] [CrossRef]
- Duke, S.O.; Gohbara, M.; Paul, R.N.; Duke, M.V. Colletothricin causes rapid membrane damage to plant cells. J. Phytopathol. 1992, 134, 289–305. [Google Scholar]
- Sze, H. Nigericin-stimulated ATPase activity in microsomal vesicles of tobacco callus. Proc. Natl. Acad. Sci. USA 1989, 77, 5904–5908. [Google Scholar]
- Thuleau, P.; Graziana, A.; Rossignol, M.; Kauss, H.; Auriol, P.; Ranjeva, R. Binding of the phytotoxin zinniol stimulates the entry of calcium into plant protoplasts. Proc. Natl. Acad. Sci. USA 1988, 85, 5932–5935. [Google Scholar]
- Iacobellis, N.S.; Bottalico, A. Effects of some trichothecenes produced by Fusarium spp. on electrolyte leakage from tomato leak disk. Phytopathol. Medierranea 1981, 20, 129–132. [Google Scholar]
- Au, T.K.; Wallace, S.H.C.; Leung, P.C. The biology of ophioboins. Life Sci. 2000, 67, 733–742. [Google Scholar]
- Leung, P.C.; Taylor, W.A.; Wang, J.H.; Tipton, C.L. Role of calmodulin inhibition in the mode of action of ophiobolin A. Plant Physiol. 1985, 77, 303–308. [Google Scholar]
- Kuhn, S.; Bussemer, J.; Chigri, F.; Vothknecht, U.C. Calcium depletion and calmodulin inhibition affect the import of nuclear-encoded proteins into plant mitochrondria. Plant J. 2009, 58, 694–705. [Google Scholar]
- Chigri, F.; Sol, J.; Vothknecht, U.C. Calcium regulation of chloroplast protein import. Plant J. 2005, 42, 821–831. [Google Scholar]
- Vaughn, K.C.; Vaughan, M.A. Mitotic disrupters from higher plants. Am. Chem. Soc. Symp. Ser. 1988, 380, 273–293. [Google Scholar]
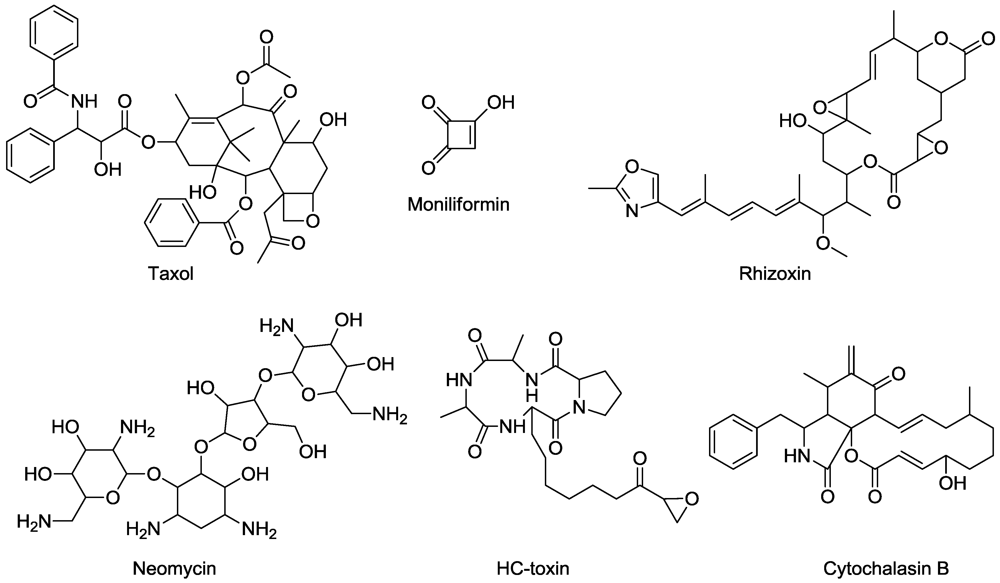
- Stierle, A.; Strobel, G.; Stierle, D. Taxol and taxane production by Taxomyces andreanae, an endophytic fungus of Pacific yew. Science 1993, 260, 214–216. [Google Scholar] [PubMed]
- Strobel, G.; Yang, X.; Sears, J.; Kramver, R.; Sidhu, R.S.; Hess, W.M. Taxol from Pestalotiopsis microspora, an endophytic fungus of Taxus wallachiana. Microbiology 1996, 142, 435–440. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, J.C.; Vaughn, K.C. Post-translational tubulin modifications in spermatogeneous cells of the pteridophyte Ceratopteris richardii. Protoplasma 1995, 186, 169–182. [Google Scholar] [CrossRef]
- Vaughn, K.C.; Vaughan, M.A. Dinitroaniline Resistance in Eleucine Indica may be due to Hyperstabilized Microtubules. In Herbicide Resistance in Weeds and Crops; Caseley, J.C., Cussans, G.W., Atkin, R.K., Eds.; Butterworth-Heinemann Ltd.: Oxford, UK, 1991; pp. 177–186. [Google Scholar]
- Scherlach, K.; Partida-Martinez, L.P.; Dahse, H.M.; Herweck, C. Antimitotic rhizotoxin derivatives from a cultured bacterial endosymbiont of the rice pathogenic fungus Rhizopus microsporus. J. Am. Chem. Soc. 2006, 128, 11529–11536. [Google Scholar] [PubMed]
- Partida-Martinez, L.P.; Hertweck, C. Pathogenic fungus harbours endosymbiont bacteria for toxin production. Nature 2005, 437, 884–888. [Google Scholar]
- Schmitt, I.; Partida-Martinez, L.P.; Winkler, R.; Voigt, K.; Einax, E.; Dolz, F.; Telle, S.; Woestemeyer, J.; Hertweck, C. Evolution of host resistance in a toxin-producing bacterial-fungal alliance. ISME. J. 2008, 2, 632–641. [Google Scholar]
- Cutler, H.G. A Fresh Look at the Wheat Coleoptile Bioassay. In Proceedings of the 11th Annual Meeting of the Plant Growth Regular Society of American, Boston, MA, USA, 29 July-1 August 1984; 11, pp. 1–9.
- Whitton, B.A. Effect of light on toxicity of various substances to Anacystis nidulans. Plant Cell Physiol. 1968, 9, 23–26. [Google Scholar]
- Larson, P.M.; Chen, T.L.L.; Wolniak, S.M. Neomycin reversibly disrupts mitotic progression in stamen hair cells of Tradescantia. J. Cell Sci. 1991, 98, 159–168. [Google Scholar]
- Styer, C.H.; Cutler, H.G. Effects of moniliformin on mitosis in maize (Zea mays L.). Plant Cell Physiol. 1984, 25, 1077–1082. [Google Scholar]
- Wyss, R.; Tamm, C.; Vederas, J.C. Differential hydrogen exchange during biosynthesis of cytochalasins B and D. Helv. Chim. Acta 1980, 63, 1538–1541. [Google Scholar]
- Yang, C.-J.; Zhai, Z.-X.; Guo, Y.-H.; Gao, P. Effects of acetylcholine, cytochalasin B and amiprophos-methyl on phloem transport in radish (Raphanus sativa). J. Integr. Plant Biol. 2007, 49, 550–550. [Google Scholar] [CrossRef]
- Walton, J.D. HC-toxin. Phytochemistry 2006, 67, 1406–1413. [Google Scholar]
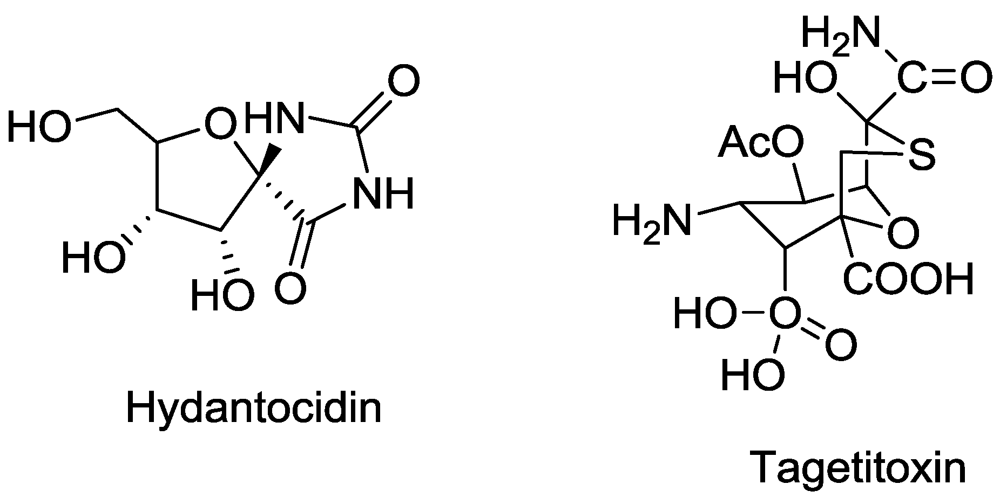
- Lukens, J.H.; Durbin, R.D. Tagetitoxin inhibits RNA synthesis directed by RNA polymerases from chloroplasts and Escherichia coli. J. Biol. Chem. 1990, 265, 493–498. [Google Scholar] [PubMed]
- Steinberg, T.H.; Matthews, D.E.; Durbin, R.D.; Burgess, R.R. Tagetitoxin: A new inhibitor of eukaryotic transcription by RNA polymerase III. J. Biol. Chem. 1990, 265, 499–505. [Google Scholar]
- Steinberg, T.H.; Burgess, R.R. Tagetitoxin inhibition of RNA polymerase III transcription results from enhanced pausing at discrete sites and is template-dependent. J. Biol. Chem. 1992, 267, 20204–20211. [Google Scholar]
- Nakajima, M.; Itoi, K.; Takamatsu, Y.; Kinoshita, T.; Okazaki, T.; Kawakubo, K.; Shindo, M.; Honma, T.; Tohjigamori, M.; Haneishi, T. Hydantocydin: A new compound with herbicidal activity from Streptomyces hygroscopicus. J. Antiobiot. 1991, 44, 293–300. [Google Scholar]
- Heim, D.R.; Cseke, C.; Gerwick, B.C.; Murdoch, M.G.; Green, S.B. Hydantocidin: A possible pro-herbicide inhibiting purine biosynthesis at the site of adenylosuccinate synthetase. Pestic. Biochem. Physiol. 1996, 55, 210–217. [Google Scholar]
- Siehl, D.L.; Subramanian, M.V.; Walters, E.W.; Lee, S.F.; Anderson, R.J.; Toschi, A.G. Adenylosuccinate synthetase: Site of action of hydantocidin, a microbial phytotoxin. Plant Physiol. 1996, 110, 753–758. [Google Scholar]
- Fonne-Pfister, R.; Chemla, P.; Ward, E.; Girardet, M.; Kreuz, E.; Honzatko, R.B.; Fromm, H.J.; Schar, H.P.; Grutter, M.G.; Cowan-Jacob, S.W. The mode of action and the structure of a herbicide in complex with its target: Binding of activated hydantocidin to the feedback regulation site of adenylsuccinate synthetase. Proc. Natl. Acad. Sci. USA 1996, 93, 9431–9436. [Google Scholar]
- Cseke, C.; Gerwick, B.C.; Crouse, G.D.; Murdoch, M.G.; Green, S.B.; Heim, D.R. 2α-phosphohydantocidin: The in vivo adenylosuccinate synthetase inhibitor responsible for hydantocidin phytotoxicity. Pestic. Biochem. Physiol. 1996, 55, 210–217. [Google Scholar] [CrossRef]
- Walters, E.W.; Lee, S.F.; Niderman, T.; Bernasconi, P.; Subramanian, M.V.; Siehl, D.L. Adenylosuccinate synthetase from maize. Purification, properties, and mechanism of inhibition by 5'-phosphohydantocidin. Plant Phyisol. 1997, 114, 549–555. [Google Scholar]
- Schmitzer, P.R.; Graupner, P.R.; Chapin, E.L.; Fields, S.C.; Gilbert, J.R.; Peacock, C.L.; Gerwick, B.C. Ribofuranosyl triazolone: A natural product herbicide with activity on adenylosuccinate synthetase following phosphorylation. J. Nat. Prod. 2000, 63, 777–781. [Google Scholar]
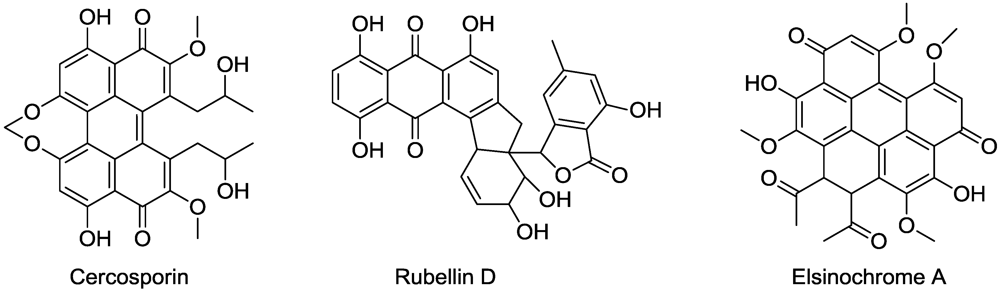
- Daub, M.E.; Hangarter, R.P. Light-induced production of singlet oxygen and superoxide by the fungal toxin, cercosporin. Plant Physiol. 1983, 73, 855–857. [Google Scholar]
- Tabuchi, H.; Tajimi, A.; Ichihara, A. (+)-Isocercosporin, a phytotoxic compound isolated from Scolecotrichum grammis Fuckel. Agric. Biol. Chem. 1991, 55, 2675–2676. [Google Scholar] [CrossRef]
- Liao, H.L.; Chung, K.R. Genetic dissection defines the roles of elsinochrome phytotoxin for fungal pathogenesis of the citrus pathogen Elsinoe fawcettii. Mol. Plant Microbe Interact. 2008, 21, 469–479. [Google Scholar] [CrossRef] [PubMed]
- Daub, M.E.; Herrero, S.; Chung, K.-R. Photoactivated perylenequinone toxins in fungal pathogenesis of plants. FEMS Microbiol. Lett. 2005, 252, 197–206. [Google Scholar]
- Daub, M.E. The fungal photosensitizer cercosporin and its role in plant disease. Amer. Chem. Soc. Symp. Ser. 1987, 339, 271–280. [Google Scholar]
- Hieser, I.; Hess, M.; Schmidtke, K.U.; Vogler, U.; Meithbauer, S.; Liebermann, B. Fatty acid peroxidation by rubellin B, C and D, phytotoxins produced by Ramularia collo-cygni (Sutton et Waller). Physiol. Mol. Plant Pathol. 2004, 64, 135–143. [Google Scholar] [CrossRef]
- Harrington, P.M.; Singh, B.K.; Szamosi, I.T.; Birk, J.H. Synthesis and herbicidal activity of cyperin. J. Agric. Food Chem. 1995, 43, 804–808. [Google Scholar]
- Kahn, A.; Kannangara, C.G. Gabaculine-resistant mutants of Chlamydomonas reinhardtii with elevated glutamate 1-semialdehyde aminotransferase activity. Carlsberg Res. Commun. 1987, 52, 73–81. [Google Scholar] [CrossRef]
- Allison, G.; Gough, K.; Rogers, L.; Smith, A. A suicide vector for allelic recombination involving the gene for glutamate 1-semialdehyde aminotransferase in the cyanobacterium Synechococcus PCC 7942. Mol. Gen. Gen. 1997, 255, 392–399. [Google Scholar]
- Werck-Reichhart, D.; Jones, O.T.; Durst, F. Haem synthesis during cytochrome P-450 induction in higher plants. 5-Aminolaevulinic acid synthesis through a five-carbon pathway in Helianthus tuberosis tuber tissues aged in the dark. Biochem. J. 1988, 249, 473–480. [Google Scholar] [PubMed]
- May, T.B.; Guikema, J.A.; Henry, R.L.; Schuler, M.K.; Wong, P.P. Gabaculine inhibition of chlorophyll biosynthesis and nodulation in Phaseolus lunatus L. Plant Physiol. 1987, 84, 1309–1313. [Google Scholar] [CrossRef] [PubMed]
- Gardner, G.; Gorton, H.L. Inhibition of phytochrome synthesis by gabaculine. Plant Physiol. 1985, 77, 540–543. [Google Scholar]
- Matson, R.S.; Kimura, T. Ferredoxin biosynthesis in Euglena gracilis. Biochim. Biophys. Acta 1976, 442, 76–87. [Google Scholar] [PubMed]
- Ohta, N.; Sager, R. Identification of a chloroplast ribosomal protein altered by a chloroplast mutation in Chlamydomonas. J. Biol. Chem. 1975, 250, 3655–3659. [Google Scholar] [PubMed]
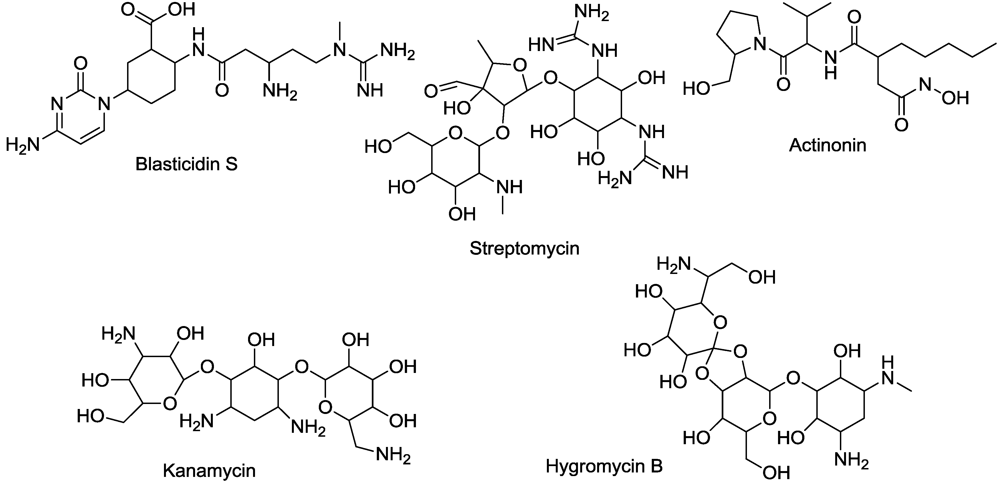
- Torregrosa, L.; Peros, J.-P.; Bouquet, L.A. Effect of hygromycin, kanamycin and phosphinothricin on the embryonic callus development and axillary micropropagation of Vitus vinifera L. Acta Hort. 1998, 528, 401–406. [Google Scholar]
- Joersbo, M.; Okkels, F.T. Calcium reduces toxicity of aminoglycoside antibiotics in sugar beet explants in vitro. Phyiol. Plant. 1996, 97, 245–250. [Google Scholar]
- Spahn, C.M.; Prescott, C.D. Throwing a spanner in the works: Antibiotics and the translation apparatus. J. Mol. Med. 1996, 74, 423–439. [Google Scholar]
- Umezawa, H.; Aoyagi, T.; Tanaka, T.; Suda, H.; Okuyama, A.; Naganawa, H.; Hamada, M.; Takeuchi, T. Production of actinonin, an inhibitor of aminopeptidase M, by actinomycetes. J. Antibiot. 1985, 38, 1629–1630. [Google Scholar] [PubMed]
- Hou, C.-X.; William, M. Actinonin-induced inhibition of plant peptide deformylase: a paradigm for the design of novel broad-spectrum herbicides. Am. Chem. Soc. Symp. Ser. 2006, 927, 243–254. [Google Scholar]
- Hou, C.-X.; Dirk, L.M.A.; Pattanik, S.; Das, N.C.; Maiti, I.B.; Houtz, R.L.; Williams, M.A. Plant peptide deformylase: A novel selectable marker and herbicide target based on essential cotranslational chloroplast protein processing. Plant Biotechnol. J. 2007, 5, 275–281. [Google Scholar]
- Fernandez-San Milan, A.; Obregon, P.; Veramendi, J. Over-expression of peptide deformylase in chloroplasts confers actinonin resistance, but is not a suitable selective marker system for plastid transformation. Transgenic Res. 2011, 20, 613–624. [Google Scholar]
- McLaughlin, J.E.; Bin-Umer, M.A.; Tortora, A.; Mendez, N.; McCormic, S.; Turner, N.E. A genome-wide screen in Saccharomyces cerevisiae reveals a critical role for the mitochondria in the toxicity of a trichothecene mycotoxin. Proc. Natl. Acad. Sci. USA 2009, 106, 21883–21888. [Google Scholar]
- Di, R.; Blechl, A.; Dill-Macky, R.; Tortora, A.; Turner, N.E. Expression of a truncated form of yeast ribosomal protein L3 in transgenic wheat improves resistance to Fusarium head blight. Plant Sci. 2010, 178, 374–380. [Google Scholar]
- Larsen, S.H.; Berry, D.M.; Paschal, J.W.; Gilliam, J.M. 5-Hydroxymethylblasticidin S and blasticidin S from Streptomyces setonii culture A83094. J. Antibiot. 1989, 42, 470–471. [Google Scholar] [PubMed]
- Nishimura, M.; Matsuo, H.; Nakamura, A.; Sugiyama, M. Purification and characterization of a puromycin-hydrolyzing enzyme from blastocidin S-producing Streptomyces morookaensis. J. Biochem. 1998, 123, 247–252. [Google Scholar] [PubMed]
- Scacchi, A.; Bortolo, R.; Cassani, G.; Pirali, G.; Nielson, E. Detection, characterization, and phytotoxic activity of the nucleoside antibiotics, blasticidin S and 5-hydroxylmethyl-blasticidin S. J. Plant Growth Regul. 1992, 11, 39–46. [Google Scholar] [CrossRef]
- Petropoulos, A.D.; Xaplanteri, M.A.; Dinos, G.P.; Wilson, D.N.; Kalpaxis, D.L. Polyamines affect diversely the antibiotic potency: Insight gained from kinetic studies of the blasticidin S and spiramycin interactions with functional ribosomes. J. Biol. Chem. 2004, 279, 26518–26525. [Google Scholar]
- Kalpaxis, D.L.; Dimitrios, L.; Theocharis, D.A.; Coutsogeorgopoulos, C. Kinetic studies on ribosomal peptidyltransferase. The behavior of the inhibitor blasticidin S. Fed. Eur. J. Biochem. 1986, 154, 267–271. [Google Scholar] [CrossRef]
- Rodriguez-Foneca, C.; Amils, R.; Garrett, R.A. Fine structure of the peptidyl transferase center on 23 S-like rRNAs deduced from chemical probing of anitbiotic-ribosome complexes. J. Mol. Biol. 1995, 247, 224–235. [Google Scholar]
- Rouxel, T.; Chupeau, Y.; Fritz, R.; Kollmann, A.; Blouquet, J.F. Biological effects of sirodesmin PL, a phytotoxin produced by Leptosphaeria maculans. Plant Sci. 1988, 57, 45–53. [Google Scholar] [CrossRef]
- Chai, C.L.L.; Waring, P. Redox sensitive epidithiodioxopiperazines in biological mechanisms of toxicity. Redox Rep. 2000, 5, 257–264. [Google Scholar]
- Wright, J.M. Phytotoxic effects of some antibiotics. Ann. Bot. 1951, 15, 493–499. [Google Scholar]
- Tanaka, T.; Hanato, K.; Watanabe, M.; Abbas, H.K. Isolation, purification and identification of 2,5-anhydro-D-glucitol as a phytotoxin from Fusarium solanii. J. Nat. Toxins 1996, 5, 317–329. [Google Scholar]
- Dayan, F.E.; Rimando, A.M.; Tellez, M.R.; Scheffler, B.E.; Roy, T.; Abbas, H.K.; Duke, S.O. Bioactivation of the fungal phytotoxin 2,4-anhydro-d-glucitol by glycolytic enzymes is an essential component of its mechanism of action. Z. Naturforsch. 2002, 57C, 645–653. [Google Scholar]
- Graupner, P.R.; Carr, A.; Clancy, E.; Gilbert, J.; Bailey, K.L.; Derby, J.A.; Gerwick, B.C. The macrocidins: Novel cyclic tetramic acids with herbicidal activity produced by Phoma macrostoma. J. Nat. Prod. 2003, 66, 1558–1561. [Google Scholar] [CrossRef] [PubMed]
- Graupner, P.R.; Gerwick, B.C.; Siddall, T.L.; Carr, A.W.; Clancy, E.; Gilbert, J.R.; Bailey, K.L.; Derby, J.A. Chlorosis inducing phytotoxic metabolites: New herbicides from Phoma macrostoma. Am. Chem. Soc. Symp. Ser. 2006, 927, 37–47. [Google Scholar]
- Okuhara, M.; Kuroda, Y.; Goto, T.; Okamoto, M.; Terano, H.; Kohsaka, M.; Aoki, H.; Imanaka, H. Studies on new phosphonic acid antibiotics. III. Isolation and characterization of FR-31564, FR-32863 and FR-33289. J. Antibiot. 1980, 33, 24–28. [Google Scholar] [PubMed]
- Lichtenthaler, H.K.; Zeidler, J.; Schwender, J.; Muller, C. The non-mevalonate isoprenoid biosynthesis of plants as a test system for new herbicides and drugs against bacteria and the malaria parasite. Z. Naturforsch. 2000, 55C, 305–313. [Google Scholar]
- Zhen, R.-G.; Singh, B.K. From inhibitors to target site genes and beyond-herbicidal inhibitors as powerful tools for functional genomics. Weed Sci. 2001, 49, 266–272. [Google Scholar]
- Jacyno, J.M.; Cutler, H.G.; Roberts, R.G.; Waters, R.M. Effects on plant growth of the HMG-CoA synthase inhibitor, 1233A/F-244/L-659,699, isolated from Scopulariopsis candidus. Agric. Biol. Chem. 1991, 55, 3129–3131. [Google Scholar] [CrossRef]
- Omura, S.; Tomoda, H.; Kumagai, H. Potent inhibitory effect of antibiotic 1233A, which specifically blocks 3-hydroxy-3-methylglutaryl coenzyme A synthase, on cholesterol biosynthesis. J. Antibiot. 1987, 40, 1356–1357. [Google Scholar] [PubMed]
- Greenspan, M.D.; Yudkovitz, J.B.; Lo, C.Y.; Chen, J.S.; Alberts, A.W.; Hunt, V.M.; Chang, M.N.; Yang, S.S.; Thompson, K.L.; Chiang, Y.C.; Chabala, J.C.; Monaghan, R.L.; Schwartz, R.L. Inhibition of hydroxymethylglutaryl-coenzyme A synthase by L-659,699. Proc. Natl. Acad. Sci. USA 1987, 84, 7488–7492. [Google Scholar]
- Duke, S.O.; Baerson, S.R.; Gressel, J. Genomics and Weeds: A synthesis. In Weedy and Invasive Plant Genomics; Stewart, C.N., Ed.; Blackwell Publishing: Singapore, 2009; pp. 221–247. [Google Scholar]
- Gerwick, B.C. Thirty years of herbicide discovery: Surveying the past and contemplating the future. Agrow 2010, VII–IX. [Google Scholar]
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Duke, S.O.; Dayan, F.E. Modes of Action of Microbially-Produced Phytotoxins. Toxins 2011, 3, 1038-1064. https://doi.org/10.3390/toxins3081038
Duke SO, Dayan FE. Modes of Action of Microbially-Produced Phytotoxins. Toxins. 2011; 3(8):1038-1064. https://doi.org/10.3390/toxins3081038
Chicago/Turabian StyleDuke, Stephen O., and Franck E. Dayan. 2011. "Modes of Action of Microbially-Produced Phytotoxins" Toxins 3, no. 8: 1038-1064. https://doi.org/10.3390/toxins3081038
APA StyleDuke, S. O., & Dayan, F. E. (2011). Modes of Action of Microbially-Produced Phytotoxins. Toxins, 3(8), 1038-1064. https://doi.org/10.3390/toxins3081038