4-Pyridone-3-carboxamide-1-β-d-ribonucleoside Triphosphate (4PyTP), a Novel NAD+ Metabolite Accumulating in Erythrocytes of Uremic Children: A Biomarker for a Toxic NAD+ Analogue in Other Tissues?

Abstract
:Abbreviations
| 2PY and 4PY | N1-methyl-2-pyridone-5-carboxamide and N1-methyl-4-pyridone-3/5-carboxamide |
| 4KNTP | 4-ketonicotinamide riboside triphosphate alias 4PyTP |
| 4Py-riboside | the N1-ribonucleoside of 4-pyridone-3/5-carboxamide |
| 4PyTP, 4PyDP, 4PyMN | the triphosphate, diphosphate and mononucleotide of 4-pyridone-3/5-carboxamide-1β-D-ribonucleoside |
| GFR | glomerular filtration rate for creatinine |
| HD | (mechanical) haemodialysis |
| IMPDH | inosine monophosphate dehydrogenase, (EC 1.1.1.205, IMP:NAD oxidoreductase) |
| N-Me-nic | N1-methylnicotinamide |
| NAD+ | nicotinamide adenine dinucleotide |
| NMN | nicotinamide mononucleotide |
| NR | nicotinamide riboside |
| PCNR | 4-pyridone carboxamide ribonucleoside (alias 4Py-riboside) |
| PD | patients undergoing continuous ambulatory peritoneal dialysis |
| r | Pearson correlation coefficient |
1. Introduction
2. Materials and Methods
2.1. Patients with Renal Failure
2.2. Processing of Blood Samples for Liquid Chromatography
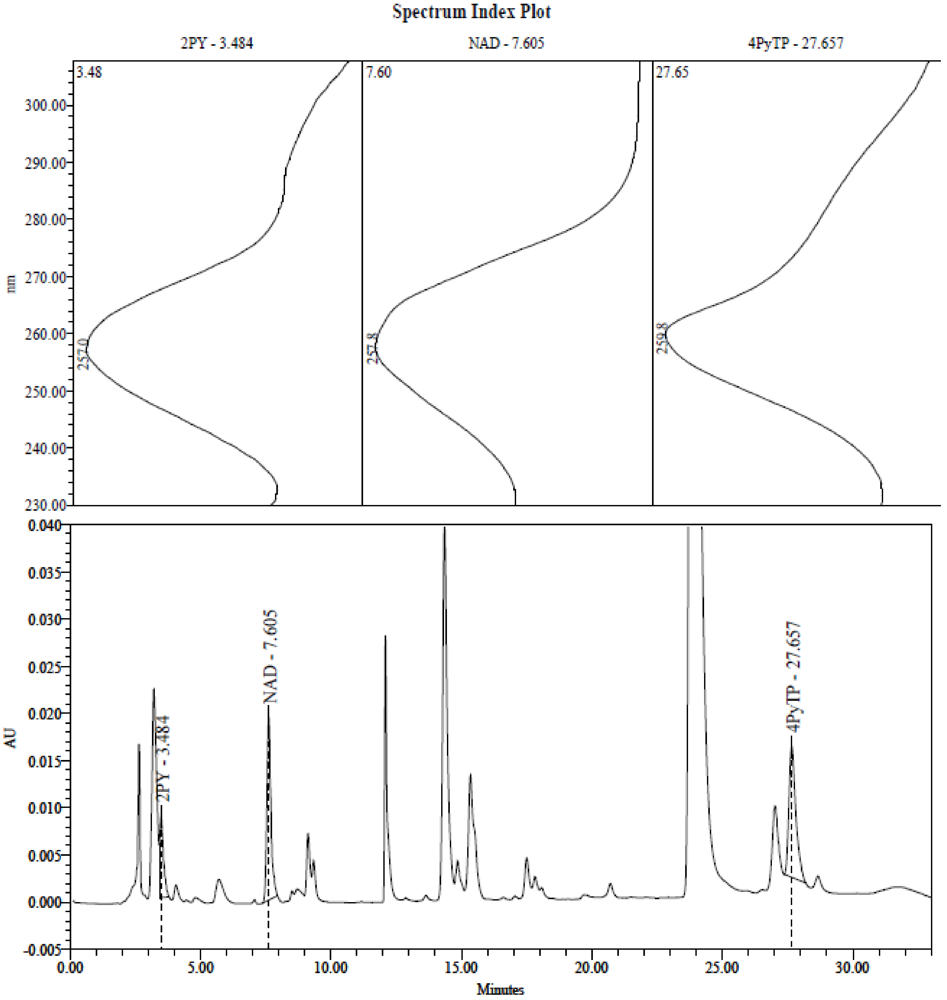
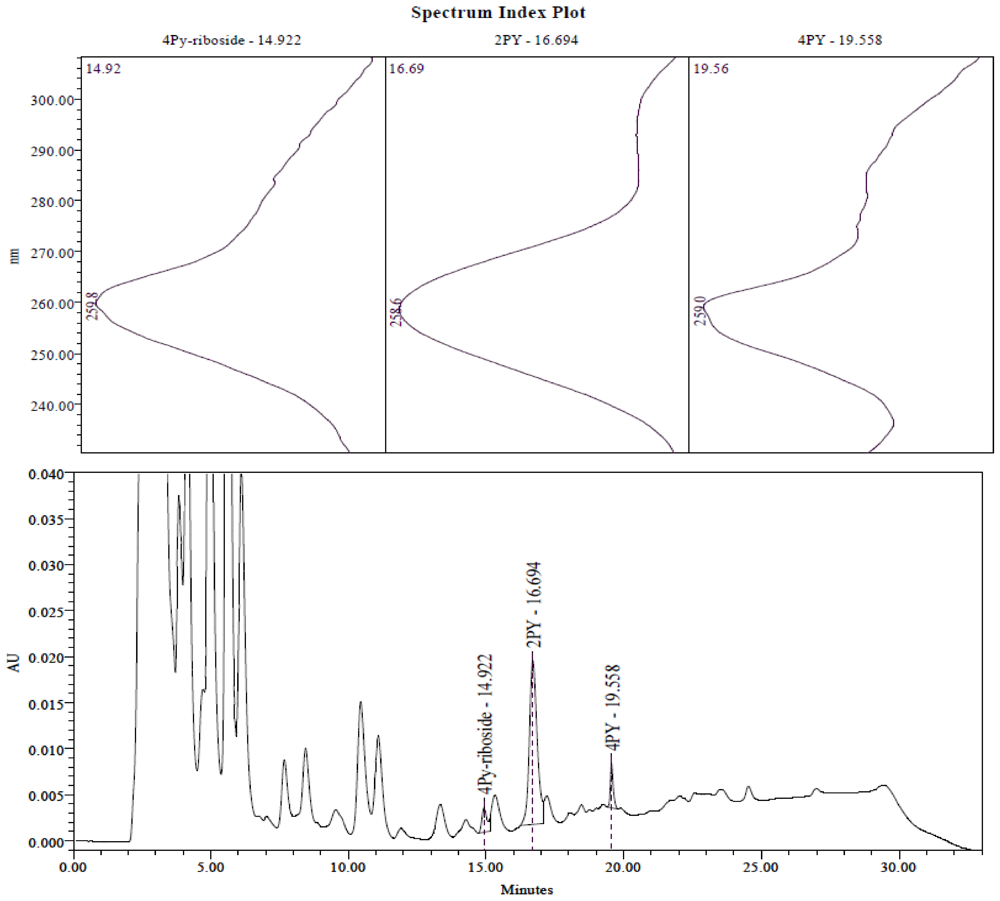
2.3. Identification of the Pyridine and Pyridone Bases and 4Py-Ribonucleoside in HPLC Profiles
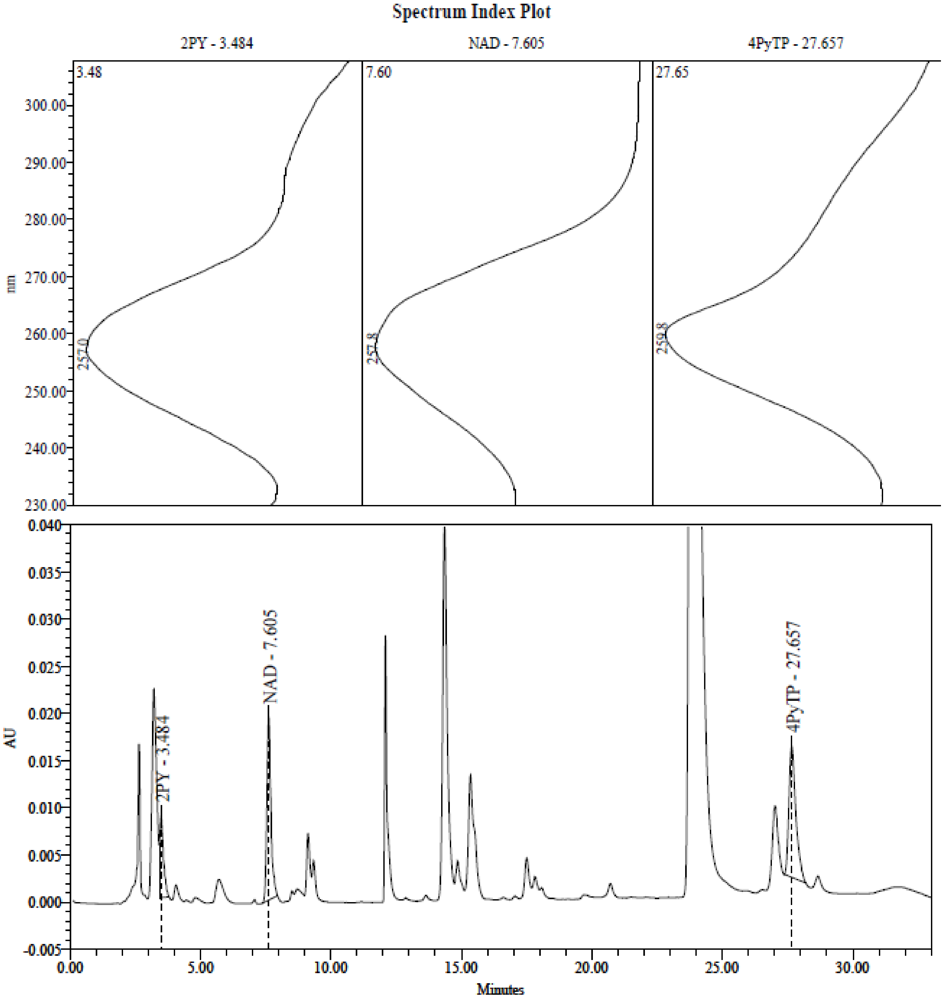
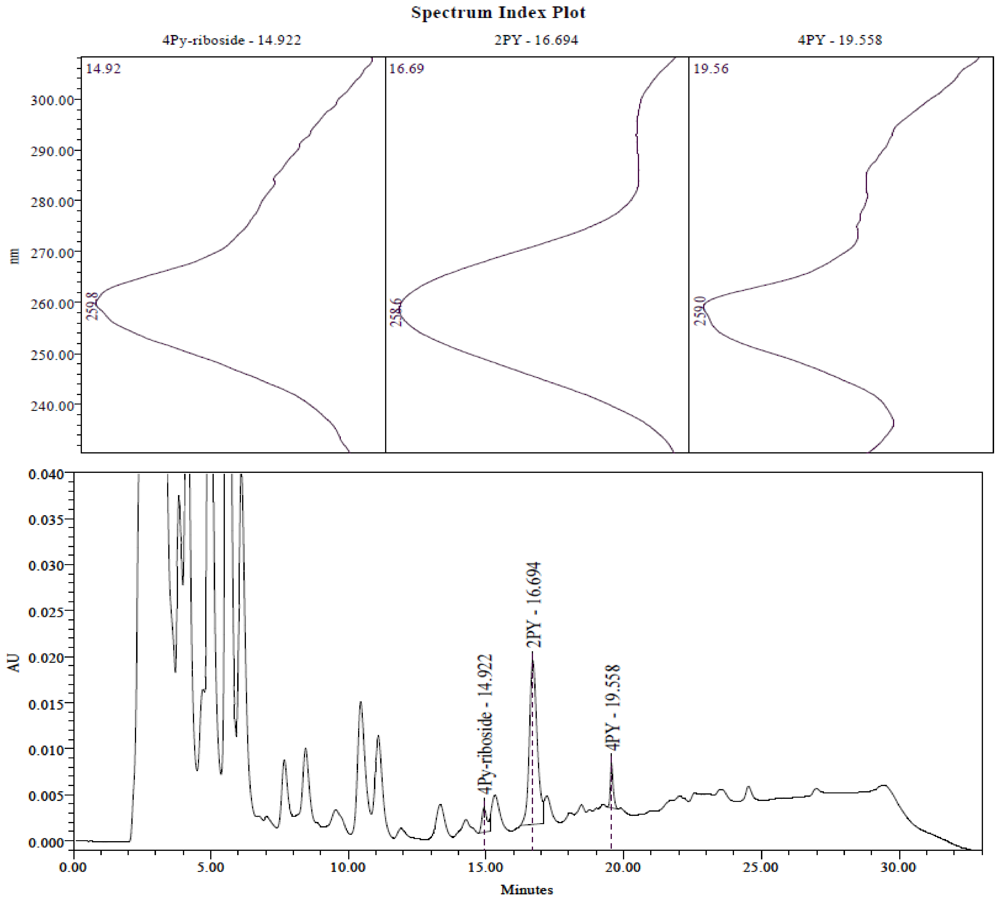
2.4. Analysis of Plasma Samples by LC-MS
3. Results and Discussion
3.1. Degree of Renal Failure and Uremia
3.2. Correlation of Methylated Pyridones and Tryptophan in Plasma with Severity of Renal Failure

{kind=link}
{kind=link}
{kind=link}
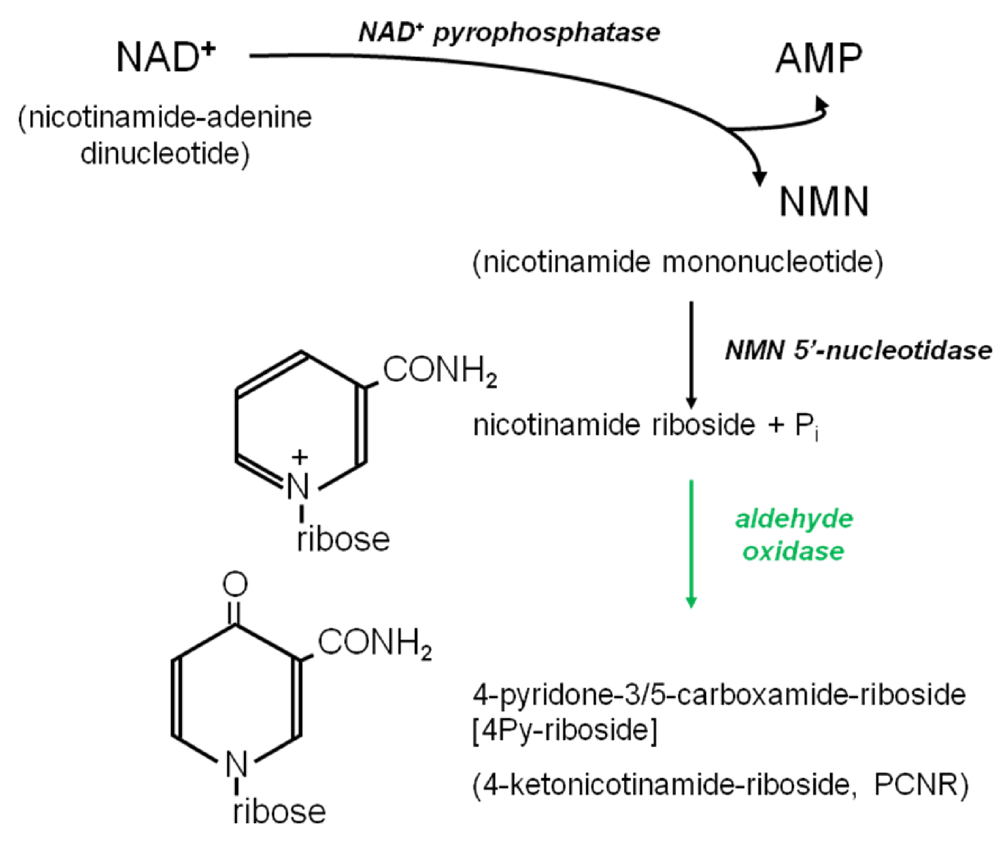
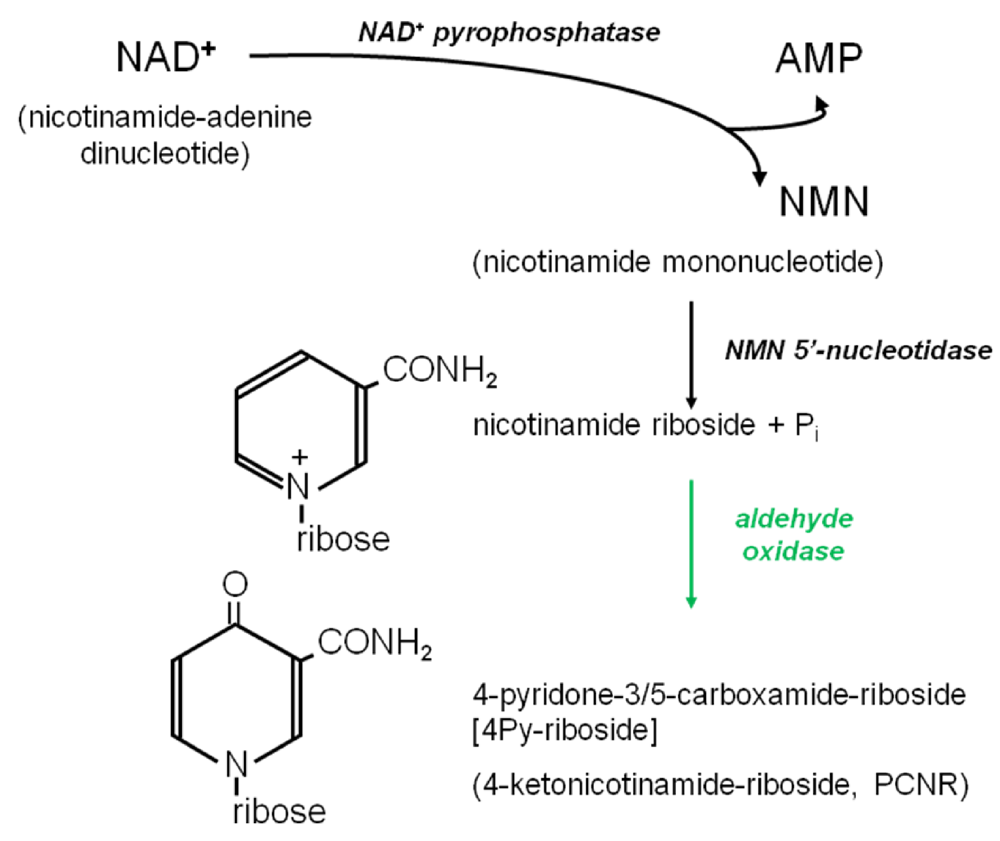{kind=link}
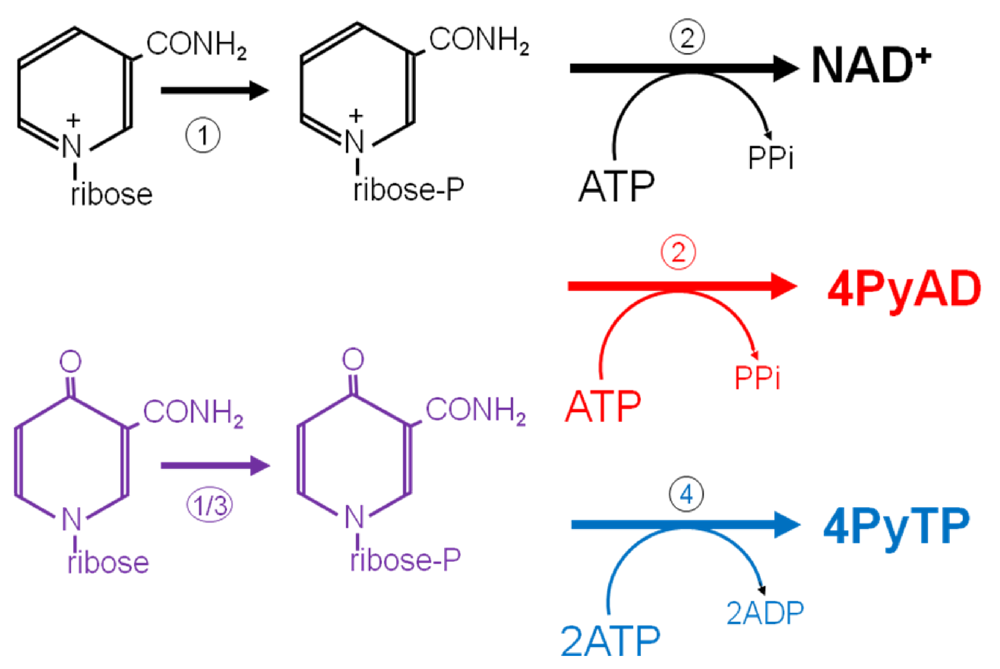{kind=link}
| Low-Normal n = 5 | Mild n = 9 | Mod-Severe n = 8 | Peritoneal n = 6 | Haemodialysis n = 6 | |
|---|---|---|---|---|---|
| GFR | 95.00 ± 20.6 | 59.22 ± 5.26 | 34.75 ± 12.83 | ≤10 | ≤10 |
| Creatinine | 95.8 ± 27.8 | 140.1 ± 27.4 | 248.5 ± 116.9 | 839.2 ± 223.9 | 1005.3 ± 258.6 |
| Urea | 4.66 ± 1.25 | 8.42 ± 3.33 | 14.85 ± 5.19 | 13.80 ± 2.83 | 23.22 ± 3.03 |
| Uric acid | 374.4 ± 56.4 | 454.3 ± 26.6 | 439.1 ± 19.5 | 426.2 ± 29.4 | 545.2 ± 51.1 |
| Tryptophan | 31.79 ± 6.58 | 29.41 ± 8.21 | 18.80 ± 8.89 | 10.53 ± 6.84 | 4.96 ± 7.75 |
| 2PY | 2.55 ± 1.05 | 6.84 ± 3.38 | 10.84 ± 3.32 | 34.90 ± 8.23 | 53.38 ± 23.60 |
| 4PY | 0.16 ± 0.23 | 1.06 ± 0.79 | 1.77 ± 0.94 | 6.45 ± 1.01 | 9.50 ± 3.06 |
| Nicotinamide | 0.38 ± 0.30 | 0.44 ± 0.19 | 0.57 ± 0.29 | 0.58 ± 0.40 | 0.94 ± 0.77 |
| N-Me-nic | 0.05 ± 0.03 | 0.08 ± 0.03 | 0.06 ± 0.03 | 0.06 ± 0.03 | 0.14 ± 0.07 |
| 4Py-riboside | 0.03 ± 0.04 | 0.08 ± 0.08 | 0.10 ± 0.12 | 0.42 ± 0.23 | 0.28 ± 0.26 |
3.2.1. Renal Failure and the Accumulation of Methylated Pyridone and Pyridine Bases
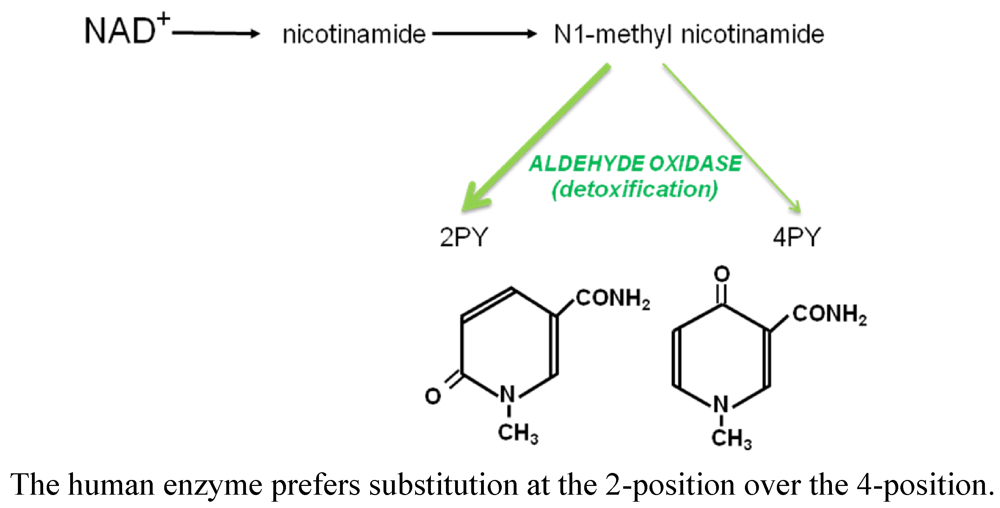
3.2.2. Presence of 2PY in Erythrocytes
3.3. Concentration of Nucleotides in Erythrocytes: Correlation with Severity of Renal Failure
| Low-Normal n = 5 | Mild n = 9 | Mod-Severe n = 8 | Peritoneal n = 6 | Haemodialysis n = 6 | |
|---|---|---|---|---|---|
| 4PyTP | 20.76 ± 5.85 | 28.96 ± 13.49 | 55.70 ± 31.28 | 200.57 ± 85.11 | 76.62 ± 25.00 |
| 4PyDP | 1.32 ± 1.44 | 2.34 ± 1.73 | 7.26 ± 6.59 | 10.38 ± 6.00 | 4.48 ± 5.42 |
| 2PY | n.d. | n.d. | n.d. | 10.5 ± 4.36 | 11.72 ± 8.46 |
| NAD | 56.82 ± 6.79 | 59.74 ± 7.05 | 67.81 ± 6.16 | 72.90 ± 9.81 | 76.48 ± 13.35 |
| NADP | 43.54 ± 2.61 | 48.8 ± 6.61 | 46.24 ± 3.53 | 47.55 ± 10.93 | 45.12 ± 4.40 |
| GTP | 44.78 ± 2.62 | 44.31 ± 3.23 | 45.26 ± 4.82 | 88.63 ± 12.74 | 81.78 ± 27.57 * |
| ATP | 1222 ± 172 | 1350 ± 261 | 1486 ± 197 | 1754 ± 306 | 1449 ± 310 * |
3.4. Measurement of 4Py-Riboside in Plasma
3.4.1. Correlation of 4Py-Riboside and 4PyTP Concentrations
3.4.2. Origins of 4Py-Riboside
3.5. The “Paradoxical” Elevation of GTP Concentrations
3.6. Further Metabolism of 4Py-Ribonucleoside
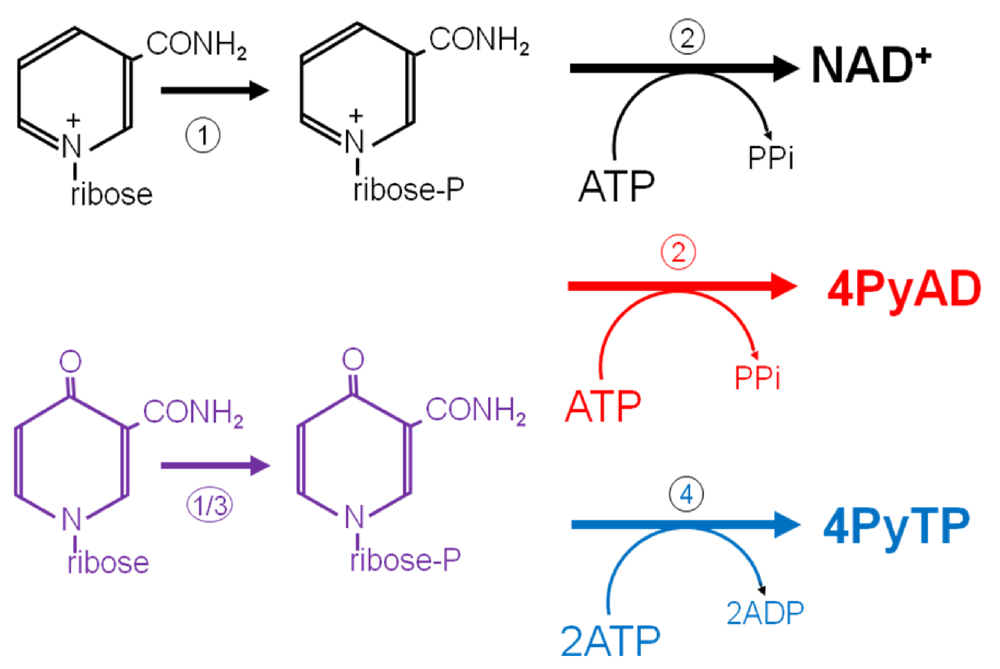
4. Conclusions
Drugs Implicated in Inhibition of IMPDH
Acknowledgments
Conflict of Interest
References
- Bürkle, A. Physiology and pathophysiology of poly(ADP-ribosyl)ation. BioEssays 2001, 23, 795–806. [Google Scholar]
- Slominska, E.M.; Smolenski, R.T.; Szolkiewicz, M.; Leaver, N.; Rutkowski, B.; Simmonds, H.A.; Swierczynski, J. Accumulation of plasma N-methyl-2-pyridone-5-carboxamide in patients with chronic renal failure. Mol. Cell. Biochem. 2002, 231, 83–88. [Google Scholar]
- Carrey, E.A.; Smolenski, R.T.; Edbury, S.M.; Laurence, A.; Marinaki, A.M.; Duley, J.A.; Zhu, L.M.; Goldsmith, D.J.A.; Simmonds, H.A. Origin and characteristics of an unusual pyridine nucleotide accumulating in erythrocytes: Positive correlation with degree of renal failure. Clin. Chim. Acta 2003, 335, 117–129. [Google Scholar]
- Carrey, E.A.; Smolenski, R.T.; Edbury, S.M.; Laurence, A.; Marinaki, A.M.; Duley, J.A.; Zhu, L.M.; Goldsmith, D.J.A.; Simmonds, H.A. An unusual pyridine nucleotide accumulating in erythrocytes: Its identity, and positive correlation with degree of renal failure. Nucleosides, Nucleotides Nucleic Acids 2004, 23, 1135–1139. [Google Scholar] [CrossRef] [PubMed]
- Slominska, E.M.; Carrey, E.A.; Foks, H.; Orlewska, C.; Wieczerzak, E.; Sowinski, P.; Yacoub, M.H.; Marinaki, A.M.; Simmonds, H.A.; Smolenski, R.T. A novel nucleotide found in human erythrocytes, 4-pyridone-3-carboxamide-1-β-D-ribonucleoside triphosphate. J. Biol. Chem. 2006, 281, 32057–32064. [Google Scholar]
- Carrey, E.A.; Synesiou, E.; Simmonds, H.A.; Fairbanks, L.D. The novel nucleotide 4kntp, in high concentrations in erythrocytes of renal failure children: A comparison with accumulation of other putative precursors in the plasma. Nucleosides, Nucleotides Nucleic Acids 2006, 25, 1051–1054. [Google Scholar] [CrossRef] [PubMed]
- Fadem, S.Z. Pediatric calculators—CKiD Schwartz and bedside Schwartz (revised pediatric equations). Available online: http://nephron.com/cgi-bin/peds_nic.cgi (accessed on 1 June 2011).
- Schwartz, G.J.; Haycock, G.B.; Edelmann, C.M., Jr.; Spitzer, A. A simple estimate of glomerular filtration rate in children derived from body length and plasma creatinine. Pediatrics 1976, 58, 259–263. [Google Scholar]
- Pawlak, D.; Pawlak, K.; Malyszko, J.; Mysliwiec, M.; Buczko, W. Accumulation of toxic products degradation of kynurenine in hemodialyzed patients. Int. Urol. Nephrol. 2001, 33, 399–404. [Google Scholar]
- Unge, G.; Lins, L.E.; Hultman, E. Tryptophan in patients on chronic haemodialysis. Lancet 1997, 2, 937. [Google Scholar]
- Qureshi, A.R.; Lindholm, B.; Garcia, E.; Groth, C.G.; Bergstrom, J. Tryptophan and its metabolites in patients on continuous ambulatory peritoneal dialysis and following renal transplantation. Nephrol. Dial. Transplant. 1994, 9, 791–796. [Google Scholar]
- Cernacek, P.; Becvarova, H.; Gerova, Z.; Valek, A.; Spustova, V. Plasma tryptophan level in chronic renal failure. Clin. Nephrol. 1980, 14, 246–249. [Google Scholar]
- Saito, A.; Niwa, T.; Maeda, K.; Kobayashi, K.; Yamamoto, Y.; Ohta, K. Tryptophan and indolic tryptophan metabolites in chronic renal failure. Am. J. Clin. Nutr. 1980, 33, 1402–1406. [Google Scholar]
- Qureshi, G.A.; Baig, S.M. The role of tryptophan, 5-hydroxy indole-3-acetic acid and their protein binding in uremic patients. Biochem. Mol. Biol. Int. 1993, 29, 411–419. [Google Scholar]
- de Torrente, A.; Glazer, G.B.; Gulyassy, P. Reduced in vivo binding of trptophan by plasma in uremia. Kidney Int. 1974, 6, 222–229. [Google Scholar]
- Mingrone, G.; DeSmet, R.; Greco, A.V.; Bertuzzi, A.; Gandolfi, A.; Ringoir, S.; Vanholder, R. Serum uremic toxins from patients with chronic renal failure displace the binding of l-tryptophan to human serum albumin. Clin. Chim. Acta 1997, 260, 27–34. [Google Scholar]
- Saito, K.; Fujigaki, S.; Heyes, M.P.; Shibata, K.; Takemura, M.; Fujii, H.; Wada, H.; Noma, A.; Seishima, M. Mechanism of increases in L-kynurenine and quinolinic acid in renal insufficiency. Am. J. Physiol. Ren. Physiol. 2000, 279, F565–F572. [Google Scholar]
- McCreanor, G.M.; Bender, D.A. The role of catabolism in controlling tissue concentrations of nicotinamide nucleotide coenzymes. Biochim. Biophys. Acta 1983, 759, 222–228. [Google Scholar]
- Fallarino, F.; Grohmann, U.; Vacca, C.; Orabana, C.; Spreca, A.; Fioretti, M.C.; Puccetti, P. T cell apoptosis by kynurenines. Adv. Exp. Med. Biol. 2003, 527, 183–190. [Google Scholar]
- Schoots, A.C.; de Vries, P.M.; Thiemann, R.; Hazejager, W.A.; Visser, S.L.; Oe, P.L. Biochemical and neurophysiological parameters in hemodialyzed patients with chronic renal failure. Clin. Chim. Acta 1989, 185, 91–107. [Google Scholar]
- Bogan, K.L.; Brenner, C. Nicotinic acid, nicotinamide, and nicotinamide riboside: A molecular evaluation of NAD+ precursor vitamins in human nutrition. Annu. Rev. Nutr. 2008, 28, 115–130. [Google Scholar] [CrossRef] [PubMed]
- Mrochek, J.E.; Jolley, R.L.; Young, D.S.; Turner, W.J. Metabolic response of humans to ingestion of nicotinic acid and nicotinamide. Clin. Chem. 1976, 22, 1821–1827. [Google Scholar]
- Fumagalli, R. Pharmacokinetics of nicotinic acid and some of its derivatives. In Metabolic Effects of Nicotinic Acid and Its Derivatives; Gey, K.F., Carlson, L.A., Eds.; Hans Huber: Bern, Switzerland, 1971; pp. 33–49. [Google Scholar]
- Stanulović, M.; Chaykin, S. Metabolic origins of the pyridones of N1-methylnicotinamide in man and rat. Arch. Biochem. Biophys. 1971, 145, 35–42. [Google Scholar]
- Beedham, C. Molybdenum hydroxylases: Biological distribution and substrate-inhibitor specificity. Prog. Med. Chem. 1987, 24, 85–127. [Google Scholar]
- Slominska, E.M.; Rutkowski, P.; Smolenski, R.T.; Szutowicz, A.; Rutkowski, B.; Swierczynski, J. The age-related increase in N-methyl-2-pyridone-5-carboxamide (NAD catabolite) in human plasma. Mol. Cell. Biochem. 2004, 267, 25–30. [Google Scholar]
- Knox, W.E.; Grossman, W.I. A new metabolite of nicotinamide. J. Biol. Chem. 1946, 166, 391–392. [Google Scholar]
- Jaeken, J.; van den Berghe, G. An infantile autistic syndrome characterized by the presence of succinyl purines in body fluids. Lancet 1984, 324, 1058–1061. [Google Scholar]
- Grossman, L.; Kaplan, N.O. Nicotinamide riboside phosphorylase from human erythrocytes. J. Biol. Chem. 1958, 231, 717–726. [Google Scholar]
- Wielgus-Kutrowska, B.; Kulikowska, E.; Wierchowski, J.; Bzowska, A.; Shugar, D. Nicotinamide riboside, an unusual, non-typical substrate of purified purine-nucleoside phosphorylases. Eur. J. Biochem. 1997, 243, 408–414. [Google Scholar]
- Dutta, S.P.; Crain, P.F.; McCloskey, J.A.; Chheda, G.B. Isolation and characterization of 1-β-D-ribofuranosylpyridin-4-one-3-carboxamide from human urine. Life Sci. 1979, 24, 1381–1388. [Google Scholar]
- Pane, F.; Oriani, G.; Kuo, K.C.T.; Gehrke, C.W.; Salvatore, F.; Sacchetti, L. Reference intervals for eight modified nucleosides in serum in a healthy population from Italy and the United States. Clin. Chem. 1992, 38, 671–677. [Google Scholar]
- Millis, G.C.; Davis, N.J.; Lertratanangkoon, K. Isolation and identification of 1-ribosyl pyridine nucleosides from human urine. Nucleosides, Nucleotidesand Nucleic Acids 1989, 8, 415–430. [Google Scholar] [CrossRef]
- Speer, J.; Gehrke, C.W.; Kuo, K.C.; Waalkes, T.P.; Borek, E. tRNA breakdown product as markers for cancer. Cancer 1979, 44, 2120–2123. [Google Scholar]
- Intrieri, M.; Calcagno, G.; Oriani, G.; Pane, F.; Zarrilli, F.; Cataldo, P.T.; Foggia, M.; Piazza, M.; Salvatore, F.; Sacchetti, L. Pseudouridine and 1-ribosylpyridin-4-one-3-carboxamide (PCNR) serum concentrations in human immunodeficiency virus type 1-infected patients are independent predictors for AIDS progression. J. Infect. Dis. 1996, 174, 199–203. [Google Scholar]
- Reiter, S.; Simmonds, H.A.; Zöllner, N.; Braun, S.L.; Knedel, M. Demonstration of a combined deficiency of xanthine oxidase and aldehyde oxidase in xanthinuric patients not forming oxipurinol. Clin. Chim. Acta 1990, 187, 221–234. [Google Scholar]
- Magni, G.; Amici, A.; Emanueli, M.; Orsomando, G.; Raffaelli, N.; Ruggierri, S. Enzymology of NAD+ homeostasis in man. Cell. Mol. Life Sci. 2004, 61, 19–34. [Google Scholar]
- Schmidt-Brauns, J.; Herbert, M.; Kemmer, G.; Kraiss, A.; Schlor, S.; Reidl, J. Is a NAD pyrophosphatase activity necessary for Haemophilus influenzae type b multiplication in the blood stream? Int. J. Med. Microbiol. 2001, 291, 219–225. [Google Scholar] [CrossRef] [PubMed]
- MacGregor, J.T.; Burkhalter, A. Biliary excretion of nicotinamide riboside. A possible role in the regulation of hepatic pyridine nucleotide dynamics. Biochem. Pharmacol. 1973, 22, 2645–2658. [Google Scholar] [CrossRef] [PubMed]
- Klecker, R.W.; Cysyk, R.L.; Collins, J.L. Zebularine metabolism by aldehyde oxidase in hepatic cytosol from humans, monkeys, dogs, rats and mice: Influence of sex and inhibitors. Bioorg. Med. Chem. 2006, 14, 62–66. [Google Scholar]
- Goldsmith, D.; Carrey, E.A.; Ebury, S.; Smolenski, R.T.; Jagodzinski, P.; Simmonds, H.A. Mycophenolate mofetil, an inhibitor of inosine monophosphate dehydrogenase, causes a paradoxical elevation of GTP in erythrocytes of renal transplant patients. Clin. Sci. 2004, 107, 63–68. [Google Scholar]
- Jagodzinski, P.; Lizakowski, S.; Smolenski, R.T.; Smolenska, E.M.; Goldsmith, D.; Simmonds, H.A.; Rutkowski, B. Mycophenolate mofetil treatment following renal transplantation decreases GTP concentrations in mononuclear leucocytes. Clin. Sci. 2004, 107, 69–74. [Google Scholar]
- Montero, C.; Duley, J.A.; Fairbanks, L.D.; McBride, M.A.; Micheli, V.; Cant, A.; Morgan, G. Demonstration of induction of erythrocyte inosine monophosphate dehydrogenase activity in Ribavirin-treated patients using a high performance liquid chromatography linked method. Clin. Chim. Acta 1995, 238, 169–178. [Google Scholar]
- Nimmersgen, E.; Fox, T.; Fleming, M.A.; Thomson, J.A. Conformational changes and stabilization of inosine 5'-monophosphate dehydrogenase associated with ligand binding and inhibition by mycophenolic acid. J. Biol. Chem. 1996, 271, 19421–19427. [Google Scholar]
- Slominska, E.M.; Orlewska, C.; Yuen, A.; Romaszko, P.; Sokolowska, E.; Foks, H.; Simmonds, H.A.; Yacoub, M.H.; Smolenski, R.T. Metabolism of 4-pyridone-3-carboxamide-1-β-D-ribonucleoside triphosphate and its nucleoside precursor in the erythrocytes. Nucleosides, Nucleotides and Nucleic Acids 2008, 27, 830–834. [Google Scholar] [CrossRef]
- Romaszko, P.; Slominska, E.M.; Orlewska, C.; Lipinski, M.; Smolenski, R.T. Metabolism of 4-pyridone-3-carboxamide-1-β-D-ribonucleoside in rodent tissues and in vivo. Mol. Cell. Biochem. 2011, 351, 143–148. [Google Scholar]
- Saunders, P.P.; Spindler, C.D.; Tan, M.T.; Alvarez, E.; Robins, R.K. Tiazofurin is phosphorylated by three enzymes from Chinese hamster ovary cells. Cancer Res. 1990, 50, 5269–5274. [Google Scholar]
- Jäger, W.; Salamon, A.; Szekeres, T. Metabolism of the novel IMP dehydrogenase inhibitor benzamide riboside. Curr. Med. Chem. 2002, 9, 781–786. [Google Scholar]
- Wu, J.Z.; Larson, G.; Walker, H.; Shim, J.H.; Hong, Z. Phosphorylation of ribavirin and viramidine by adenosine kinase and cytosolic 5'-nucleotidase II, implications for ribavirin metabolism in erythrocytes. Antimicrob. Agents Chemother. 2005, 49, 2164–2171. [Google Scholar]
- Yamada, Y.; Natsumeda, Y.; Weber, G. Action of the active metabolites of tiazofurin and ribavirin on purified IMP dehydrogenase. Biochemistry 1988, 27, 2193–2196. [Google Scholar]
- Saunders, P.P.; Kuttan, R.; Lai, M.M.; Robins, R.K. Action of 2-β-D-ribofuranosylthiazole-4-carboxamide (Tiazofurin) in Chinese hamster ovary and variant cell lines. Mol. Pharmacol. 1983, 23, 534–539. [Google Scholar]
- Zhen, W.; Jayaram, H.N.; Weber, G. Antitumor activity of tiazofurin in human colon carcinoma HY-29. Cancer Invest. 1992, 10, 505–511. [Google Scholar]
- Monks, A.; Marquez, V.E.; Mao, D.T.; Cysyk, R.L. Uptake of 2-β-D-ribofuranosylthiazole-4-carboxamide (Tiazofurin) and analogues by the facilitated transport mechanism of erythrocytes. Cancer Lett. 1985, 28, 1–8. [Google Scholar]
- Damaraju, V.L.; Visser, F.; Zhang, J.; Mowles, D.; Ng, A.M.L.; Young, J.D.; Jayaram, H.N.; Cass, C.E. Role of human nucleoside transporters in the cellular uptake of two inhibitors of IMP dehydrogenase, tiazofurin and benzamide riboside. Mol. Pharmacol. 2005, 67, 273–279. [Google Scholar]
- Roberts, J.D.; Tong, W.P.; Hartshorn, J.N.; Hacker, M.P. Metabolism of tiazofurin by human erythrocytes and mononuclear blood cells in vitro. Cancer Lett. 1986, 32, 193–197. [Google Scholar]
- Zhen, W.; Jayaram, H.N.; Weber, G. Determination of thiazole-4-carboxamide adenine dinucleotide (TAD) levels in mononuclear cells of leukemic patients treated with tiazofurin. Biochem. Pharmacol. 1991, 41, 281–286. [Google Scholar]
- Vranić-Mandušić, V.; Subota, V.; Savovski, K.; Medić, L.; Dramićanin, T.; Jozanov-Stankov, O.; Popov-Čeleketić, D.; Jokanović, M.; Dimitrijević, B. Erythrocytotoxicity of tiazofurin in vivo and in vitro detected by scanning probe microscopy. Toxicol. Lett. 2004, 146, 275–284. [Google Scholar]
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Synesiou, E.; Fairbanks, L.D.; Simmonds, H.A.; Slominska, E.M.; Smolenski, R.T.; Carrey, E.A. 4-Pyridone-3-carboxamide-1-β-d-ribonucleoside Triphosphate (4PyTP), a Novel NAD+ Metabolite Accumulating in Erythrocytes of Uremic Children: A Biomarker for a Toxic NAD+ Analogue in Other Tissues? Toxins 2011, 3, 520-537. https://doi.org/10.3390/toxins3060520
Synesiou E, Fairbanks LD, Simmonds HA, Slominska EM, Smolenski RT, Carrey EA. 4-Pyridone-3-carboxamide-1-β-d-ribonucleoside Triphosphate (4PyTP), a Novel NAD+ Metabolite Accumulating in Erythrocytes of Uremic Children: A Biomarker for a Toxic NAD+ Analogue in Other Tissues? Toxins. 2011; 3(6):520-537. https://doi.org/10.3390/toxins3060520
Chicago/Turabian StyleSynesiou, Elena, Lynnette D. Fairbanks, H. Anne Simmonds, Ewa M. Slominska, Ryszard T. Smolenski, and Elizabeth A. Carrey. 2011. "4-Pyridone-3-carboxamide-1-β-d-ribonucleoside Triphosphate (4PyTP), a Novel NAD+ Metabolite Accumulating in Erythrocytes of Uremic Children: A Biomarker for a Toxic NAD+ Analogue in Other Tissues?" Toxins 3, no. 6: 520-537. https://doi.org/10.3390/toxins3060520
APA StyleSynesiou, E., Fairbanks, L. D., Simmonds, H. A., Slominska, E. M., Smolenski, R. T., & Carrey, E. A. (2011). 4-Pyridone-3-carboxamide-1-β-d-ribonucleoside Triphosphate (4PyTP), a Novel NAD+ Metabolite Accumulating in Erythrocytes of Uremic Children: A Biomarker for a Toxic NAD+ Analogue in Other Tissues? Toxins, 3(6), 520-537. https://doi.org/10.3390/toxins3060520