Nymphoides peltata Alleviates Patulin-Induced Glutamine Metabolic Stress and Epithelial Toxicity in Small Intestinal Epithelial Cells


Abstract

1. Introduction
2. Results
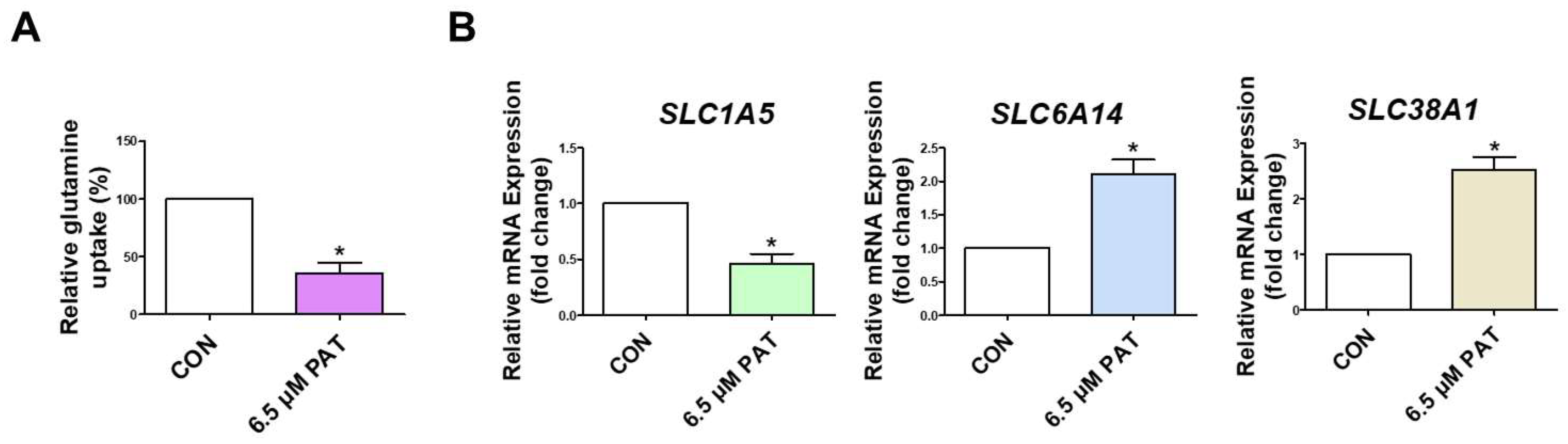
2.1. PAT Suppresses GLN Uptake and Alters GLN Transporter Expression
2.2. PAT Induces GLN Synthetase and Glutaminase Activity
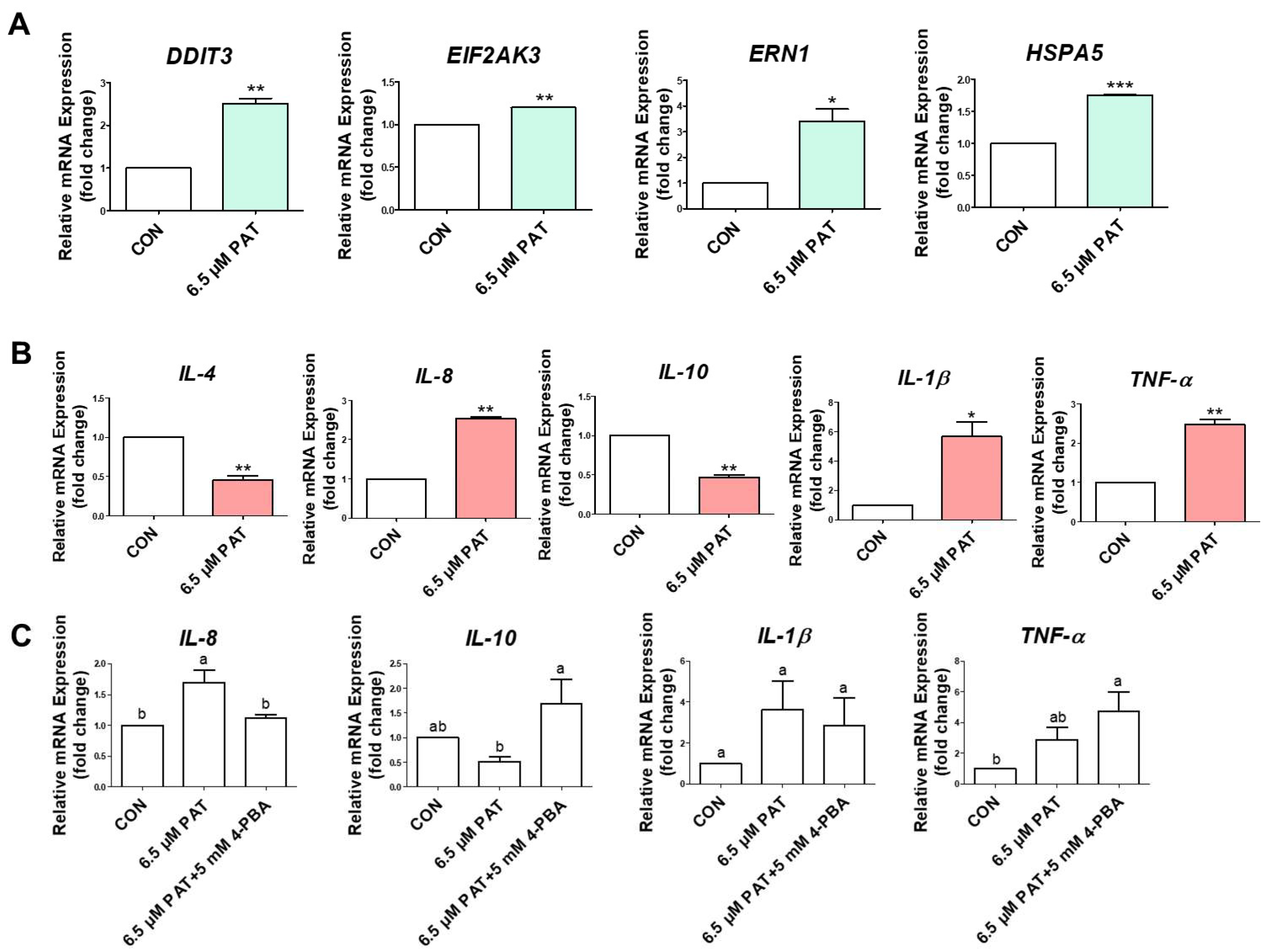
2.3. PAT Activates ER Stress and Inflammatory Cytokine Expression
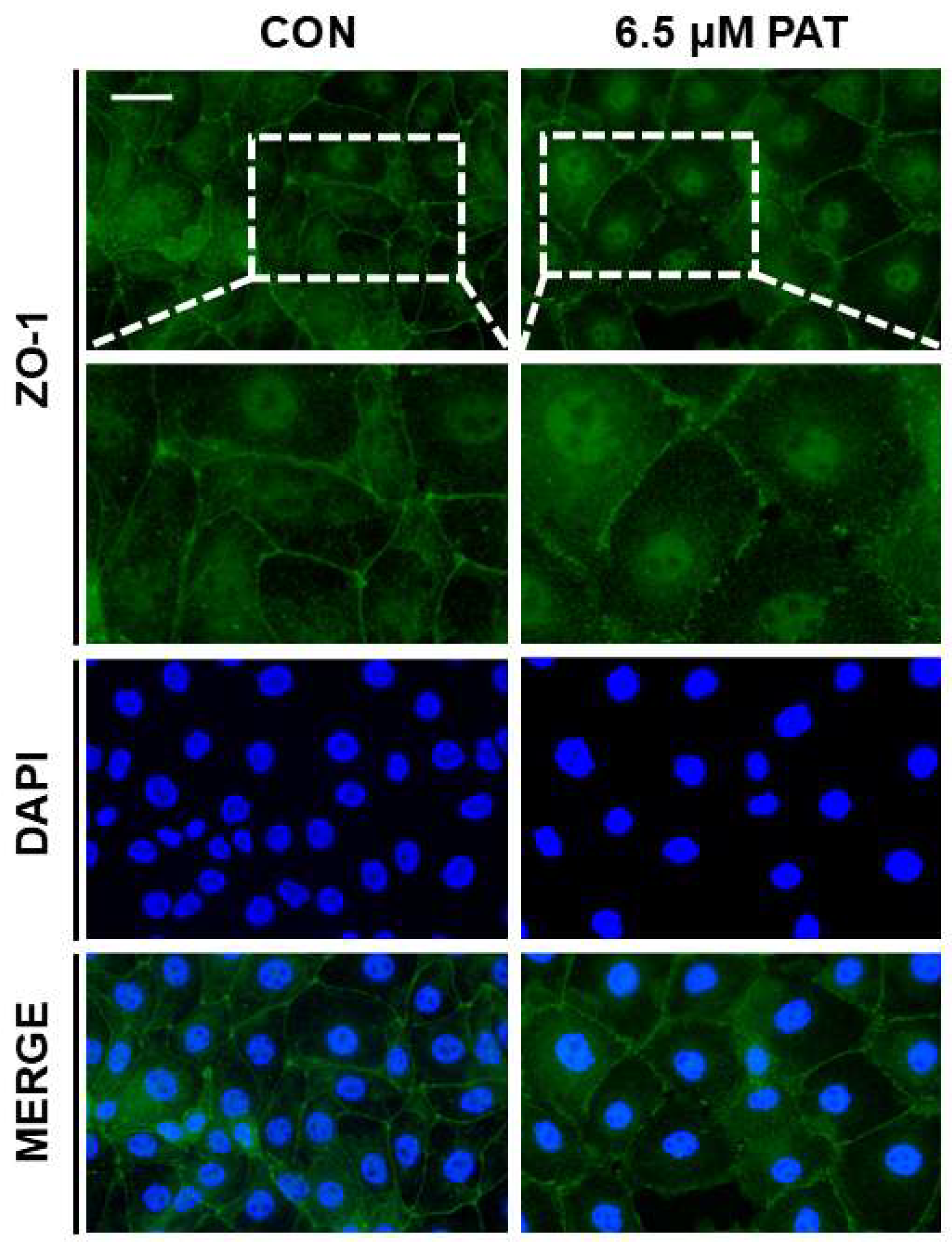
2.4. PAT Disrupts Epithelial Barrier Integrity
2.5. Effects of GLN on PAT-Induced ER Stress, Inflammatory Responses, and Barrier Dysfunction in IPEC-J2 Cells
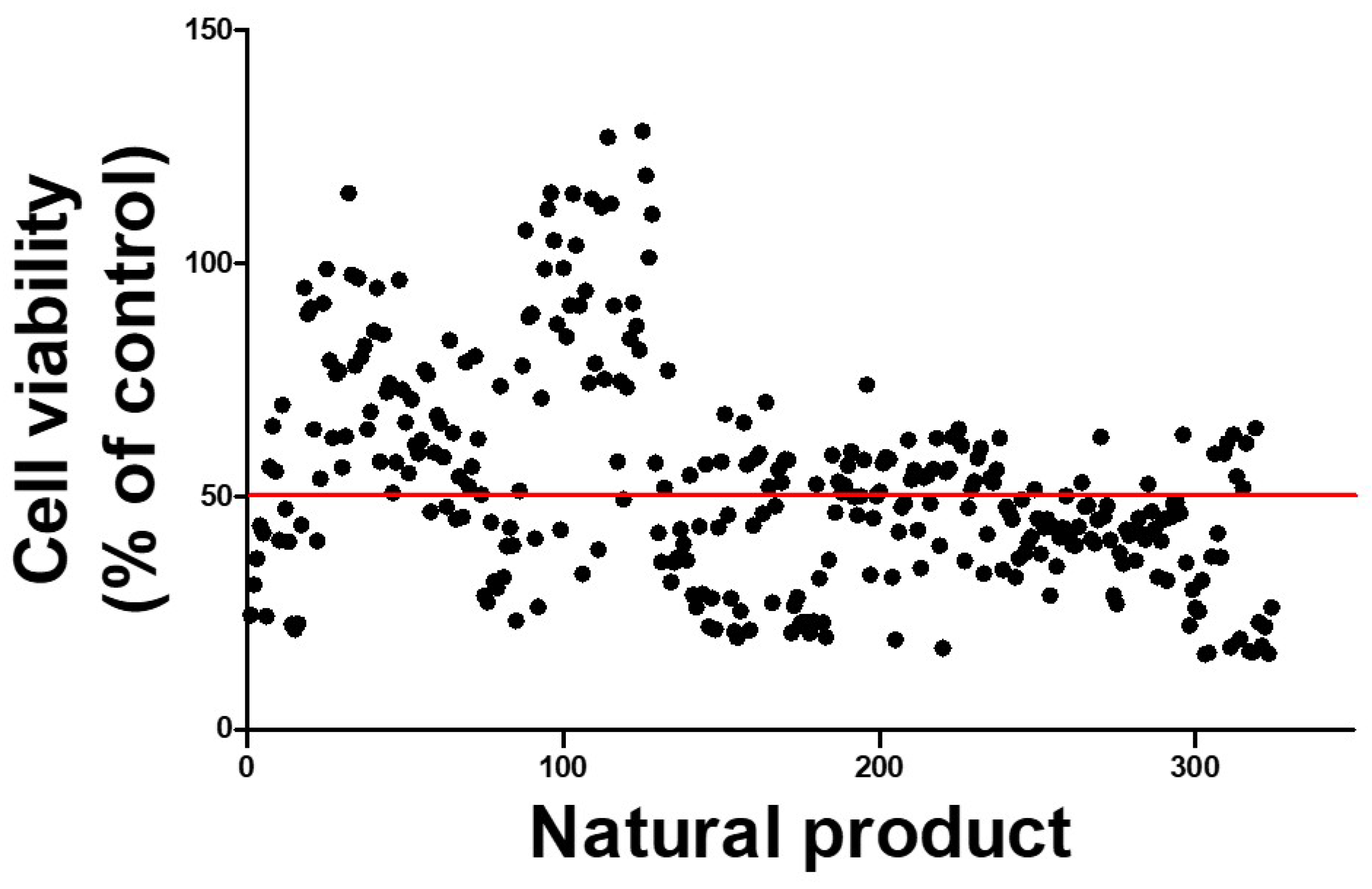
2.6. Protective Natural Products Showing Promise in Counteracting PAT-Induced Cytotoxicity
2.7. Nymphoides peltata Extract Modulates GLN Uptake, ER Stress, Inflammation, and Tight Junction Localization in PAT-Treated IPEC-J2 Cells
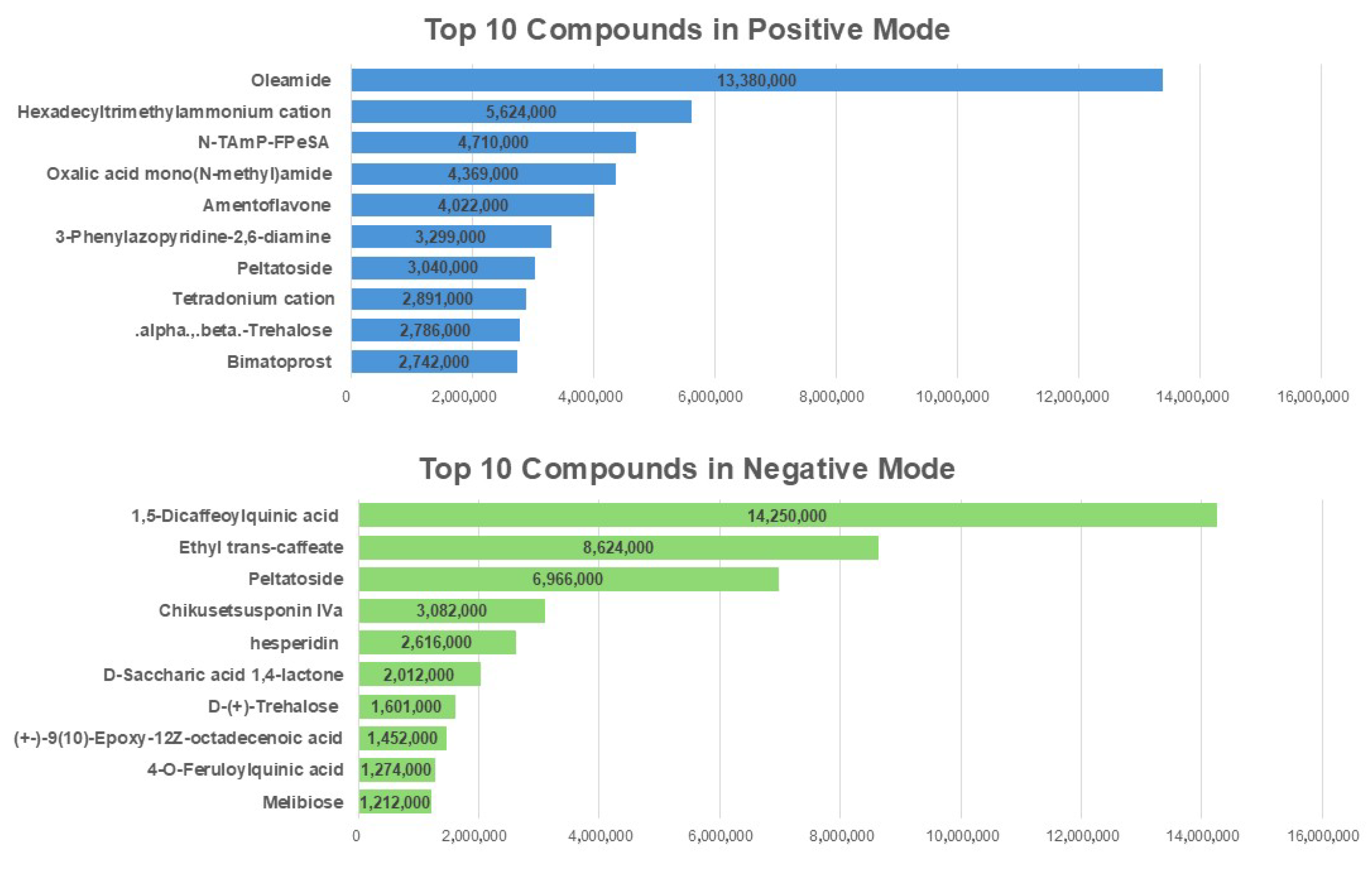
2.8. The Identification of the Abundant Bioactive Compounds in N. peltata Extract Using LC-MS/MS
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Reagents
5.2. Cell Culturing and Treatment
5.3. Glutamine Uptake Assay
5.4. Glutamine Synthetase Activity Assay
5.5. Quantitative Real-Time PCR
5.6. Immunofluorescence
5.7. High-Throughput Screening of Natural Products
5.8. LC-MS/MS Analysis of Nymphoides peltata Extract
5.9. Statistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| PAT | Patulin |
| GLN | Glutamine |
| ER | Endoplasmic reticulum |
| N. peltata | Nymphoides peltata |
| GS | Glutamine synthetase |
| ZO-1 | Zonula occludin 1 |
| PCR | Polymerase chain reaction |
References
- Speijers, G.J.; Franken, M.A.; van Leeuwen, F.X. Subacute toxicity study of patulin in the rat: Effects on the kidney and the gastro-intestinal tract. Food Chem. Toxicol. 1988, 26, 23–30. [Google Scholar] [CrossRef]
- McKinley, E.R.; Carlton, W.W.; Boon, G.D. Patulin mycotoxicosis in the rat: Toxicology, pathology and clinical pathology. Food Chem. Toxicol. 1982, 20, 289–300. [Google Scholar] [CrossRef]
- Escoula, L.; Thomsen, M.; Bourdiol, D.; Pipy, B.; Peuriere, S.; Roubinet, F. Patulin immunotoxicology: Effect on phagocyte activation and the cellular and humoral immune system of mice and rabbits. Int. J. Immunopharmacol. 1988, 10, 983–989. [Google Scholar] [CrossRef] [PubMed]
- Fliege, R.; Metzler, M. Electrophilic properties of patulin. N-acetylcysteine and glutathione adducts. Chem. Res. Toxicol. 2000, 13, 373–381. [Google Scholar] [CrossRef]
- Fliege, R.; Metzler, M. The mycotoxin patulin induces intra- and intermolecular protein crosslinks in vitro involving cysteine, lysine, and histidine side chains, and alpha-amino groups. Chem. Biol. Interact. 1999, 123, 85–103. [Google Scholar] [CrossRef]
- Puel, O.; Galtier, P.; Oswald, I.P. Biosynthesis and toxicological effects of patulin. Toxins 2010, 2, 613–631. [Google Scholar] [CrossRef] [PubMed]
- Goossens, J.; Pasmans, F.; Verbrugghe, E.; Vandenbroucke, V.; De Baere, S.; Meyer, E.; Haesebrouck, F.; De Backer, P.; Croubels, S. Porcine intestinal epithelial barrier disruption by the Fusarium mycotoxins deoxynivalenol and T-2 toxin promotes transepithelial passage of doxycycline and paromomycin. BMC Vet. Res. 2012, 8, 245. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Bansal, M.; Pal, S.; Alam, S.; Jagdale, P.; Ayanur, A.; Ansari, K.M. COX-2/EP2-EP4/β-catenin signaling regulates patulin-induced intestinal cell proliferation and inflammation. Toxicol. Appl. Pharmacol. 2018, 356, 224–234. [Google Scholar] [CrossRef]
- Zhang, T.; Chang, M.; Hou, X.; Yan, M.; Zhang, S.; Song, W.; Sheng, Q.; Yuan, Y.; Yue, T. Apple polyphenols prevent patulin-induced intestinal damage by modulating the gut microbiota and metabolism of the gut-liver axis. Food Chem. 2025, 463, 141049. [Google Scholar] [CrossRef]
- Assunção, R.; Alvito, P.; Kleiveland, C.R.; Lea, T.E. Characterization of in vitro effects of patulin on intestinal epithelial and immune cells. Toxicol. Lett. 2016, 250–251, 47–56. [Google Scholar] [CrossRef]
- Li, P.; Kim, S.W.; Li, X.; Datta, S.; Pond, W.G.; Wu, G. Dietary supplementation with cholesterol and docosahexaenoic acid affects concentrations of amino acids in tissues of young pigs. Amino Acids 2009, 37, 709–716. [Google Scholar] [CrossRef]
- Wu, G.; Bazer, F.W.; Johnson, G.A.; Knabe, D.A.; Burghardt, R.C.; Spencer, T.E.; Li, X.L.; Wang, J.J. TRIENNIAL GROWTH SYMPOSIUM: Important roles for L-glutamine in swine nutrition and production1,2. J. Anim. Sci. 2011, 89, 2017–2030. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, D.C.; da Silva Lima, F.; Sartori, T.; Santos, A.C.A.; Rogero, M.M.; Fock, R.A. Glutamine metabolism and its effects on immune response: Molecular mechanism and gene expression. Nutrire 2016, 41, 14. [Google Scholar] [CrossRef]
- Santos, A.A.; Braga-Neto, M.B.; Oliveira, M.R.; Freire, R.S.; Barros, E.B.; Santiago, T.M.; Rebelo, L.M.; Mermelstein, C.; Warren, C.A.; Guerrant, R.L.; et al. Glutamine and alanyl-glutamine increase RhoA expression and reduce Clostridium difficile toxin-a-induced intestinal epithelial cell damage. Biomed Res. Int. 2013, 2013, 152052. [Google Scholar] [CrossRef] [PubMed]
- Xing, S.; Zhang, B.; Lin, M.; Zhou, P.; Li, J.; Zhang, L.; Gao, F.; Zhou, G. Effects of alanyl-glutamine supplementation on the small intestinal mucosa barrier in weaned piglets. Asian-Australas J. Anim. Sci. 2017, 30, 236–245. [Google Scholar] [CrossRef]
- Yeh, S.-L.; Shih, Y.-M.; Lin, M.-T. Chapter 25—Glutamine and Its Antioxidative Potentials in Diabetes. In Diabetes, 2nd ed.; Preedy, V.R., Ed.; Academic Press: Cambridge, MA, USA, 2020; pp. 255–264. [Google Scholar]
- Griffiths, R.D.; Esanu-Varnav, C. Glutamine-Supplemented Parenteral Nutrition. In Problems of the Gastrointestinal Tract in Anesthesia, the Perioperative Period, and Intensive Care; Springer: Berlin/Heidelberg, Germany, 1999; pp. 195–211. [Google Scholar]
- Amores-Sánchez, M.a.I.; Medina, M.Á. Glutamine, as a Precursor of Glutathione, and Oxidative Stress. Mol. Genet. Metab. 1999, 67, 100–105. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zhang, C.; Wu, G.; Sun, Y.; Wang, B.; He, B.; Dai, Z.; Wu, Z. Glutamine enhances tight junction protein expression and modulates corticotropin-releasing factor signaling in the jejunum of weanling piglets. J. Nutr. 2015, 145, 25–31. [Google Scholar] [CrossRef]
- Chen, S.; Xia, Y.; Zhu, G.; Yan, J.; Tan, C.; Deng, B.; Deng, J.; Yin, Y.; Ren, W. Glutamine supplementation improves intestinal cell proliferation and stem cell differentiation in weanling mice. Food Nutr. Res. 2018, 62. [Google Scholar] [CrossRef]
- Xi, P.; Jiang, Z.; Dai, Z.; Li, X.; Yao, K.; Zheng, C.; Lin, Y.; Wang, J.; Wu, G. Regulation of protein turnover by l-glutamine in porcine intestinal epithelial cells. J. Nutr. Biochem. 2012, 23, 1012–1017. [Google Scholar] [CrossRef]
- Yadav, V.; Mythri, C.; Kumarasamy, M. Natural products as potential modulators of pro-inflammatory cytokines signalling in Alzheimer′s disease. Brain Behav. Immun. Integr. 2024, 5, 100048. [Google Scholar] [CrossRef]
- Cragg, G.M.; Pezzuto, J.M. Natural Products as a Vital Source for the Discovery of Cancer Chemotherapeutic and Chemopreventive Agents. Med. Princ. Pract. 2016, 25 (Suppl. S2), 41–59. [Google Scholar] [CrossRef]
- Wang, C.; Fu, Y.; Wang, R.; Wang, Q.; Yu, H.; Zhang, J. Quercetin Attenuates the Combined Effects of Zearalenone and Lipopolysaccharide on IPEC-J2 Cell Injury through Activating the Nrf2 Signaling Pathway. Toxins 2023, 15, 679. [Google Scholar] [CrossRef] [PubMed]
- Najafzadeh, M.; Najafzadeh, M.; Adoul, Z.; Jafarinejad, S.; Baumgartner, A.; Reynolds, D.; Isreb, M.; Ghaderi, R.; Sefat, F.; Heidari, S.; et al. P161 The use of Novel Natural Treatments for Inflammatory Bowel Diseases. J. Crohn’s Colitis 2024, 18, i466. [Google Scholar] [CrossRef]
- Debnath, T.; Kim, D.H.; Lim, B.O. Natural Products as a Source of Anti-Inflammatory Agents Associated with Inflammatory Bowel Disease. Molecules 2013, 18, 7253–7270. [Google Scholar] [CrossRef]
- Chen, W.; Jia, Z.; Pan, M.-H.; Anandh Babu, P.V. Natural Products for the Prevention of Oxidative Stress-Related Diseases: Mechanisms and Strategies. Oxidative Med. Cell. Longev. 2016, 2016, 4628502. [Google Scholar] [CrossRef]
- Pal, S.; Singh, N.; Ansari, K.M. Toxicological effects of patulin mycotoxin on the mammalian system: An overview. Toxicol. Res. 2017, 6, 764–771. [Google Scholar] [CrossRef] [PubMed]
- Bacha, S.A.S.; Li, Y.; Nie, J.; Xu, G.; Han, L.; Farooq, S. Comprehensive review on patulin and Alternaria toxins in fruit and derived products. Front. Plant Sci. 2023, 14. [Google Scholar] [CrossRef]
- Zhang, T.; Guo, Z.; Che, J.; Yan, M.; Liang, J.; Wang, F.; Hu, J.; Song, W.; Yuan, Y.; Yue, T. Dysbiosis of intestinal microbiota and metabolism caused by acute patulin exposure in mice. Food Front. 2024, 5, 1819–1832. [Google Scholar] [CrossRef]
- Mahfoud, R.; Maresca, M.; Garmy, N.; Fantini, J. The mycotoxin patulin alters the barrier function of the intestinal epithelium: Mechanism of action of the toxin and protective effects of glutathione. Toxicol. Appl. Pharmacol. 2002, 181, 209–218. [Google Scholar] [CrossRef]
- Lee, C.H.; Shin, S.; Lee, S.I. 10-Eicosanol Alleviates Patulin-Induced Cell Cycle Arrest and Apoptosis by Activating AKT (Protein Kinase B) in Porcine Intestinal Epithelial Cells. Int. J. Mol. Sci. 2024, 25, 8597. [Google Scholar] [CrossRef]
- Nicklin, P.; Bergman, P.; Zhang, B.; Triantafellow, E.; Wang, H.; Nyfeler, B.; Yang, H.; Hild, M.; Kung, C.; Wilson, C.; et al. Bidirectional Transport of Amino Acids Regulates mTOR and Autophagy. Cell 2009, 136, 521–534. [Google Scholar] [CrossRef]
- Pan, P.; Ying, Y.; Ma, F.; Zou, C.; Yu, Y.; Li, Y.; Li, Z.; Fang, Y.; Huang, T.; Ge, R.-S.; et al. Zearalenone disrupts the placental function of rats: A possible mechanism causing intrauterine growth restriction. Food Chem. Toxicol. 2020, 145, 111698. [Google Scholar] [CrossRef] [PubMed]
- Sikder, M.O.F.; Yang, S.; Ganapathy, V.; Bhutia, Y.D. The Na+/Cl−-Coupled, Broad-Specific, Amino Acid Transporter SLC6A14 (ATB0,+): Emerging Roles in Multiple Diseases and Therapeutic Potential for Treatment and Diagnosis. AAPS J. 2017, 20, 12. [Google Scholar] [CrossRef]
- Bröer, A.; Rahimi, F.; Bröer, S. Deletion of Amino Acid Transporter ASCT2 (SLC1A5) Reveals an Essential Role for Transporters SNAT1 (SLC38A1) and SNAT2 (SLC38A2) to Sustain Glutaminolysis in Cancer Cells. J. Biol. Chem. 2016, 291, 13194–13205. [Google Scholar] [CrossRef] [PubMed]
- Issaq, S.H.; Mendoza, A.; Fox, S.D.; Helman, L.J. Glutamine synthetase is necessary for sarcoma adaptation to glutamine deprivation and tumor growth. Oncogenesis 2019, 8, 20. [Google Scholar] [CrossRef]
- Deng, S.J.; Chen, H.Y.; Zeng, Z.; Deng, S.; Zhu, S.; Ye, Z.; He, C.; Liu, M.L.; Huang, K.; Zhong, J.X.; et al. Nutrient Stress-Dysregulated Antisense lncRNA GLS-AS Impairs GLS-Mediated Metabolism and Represses Pancreatic Cancer Progression. Cancer Res. 2019, 79, 1398–1412. [Google Scholar] [CrossRef]
- He, L.; Zhou, X.; Wu, Z.; Feng, Y.; Liu, D.; Li, T.; Yin, Y. Glutamine in suppression of lipopolysaccharide-induced piglet intestinal inflammation: The crosstalk between AMPK activation and mitochondrial function. Anim. Nutr. 2022, 10, 137–147. [Google Scholar] [CrossRef]
- Schemitt, E.; Rosa, C.; Fillmann, H.; Dias, A.; Marroni, C.; Marroni, N. Glutamine Modulates Heat Shock Proteins, Endoplasmic Reticulum Stress and Cell Death in Rats with Thioacetamide-Induced Severe Acute Liver Failure. J. Biomed. Res. Environ. Sci. 2024, 5, 021–032. [Google Scholar] [CrossRef]
- Xu, H.; Liu, G.; Gu, H.; Wang, J.; Li, Y. Glutamine protects intestine against ischemia-reperfusion injury by alleviating endoplasmic reticulum stress induced apoptosis in rats. Acta Cirúrgica Bras. 2020, 35, e202000104. [Google Scholar] [CrossRef] [PubMed]
- Van Itallie, C.M.; Fanning, A.S.; Bridges, A.; Anderson, J.M. ZO-1 stabilizes the tight junction solute barrier through coupling to the perijunctional cytoskeleton. Mol. Biol. Cell 2009, 20, 3930–3940. [Google Scholar] [CrossRef]
- Rodgers, L.S.; Beam, M.T.; Anderson, J.M.; Fanning, A.S. Epithelial barrier assembly requires coordinated activity of multiple domains of the tight junction protein ZO-1. J. Cell Sci. 2013, 126 Pt 7, 1565–1575. [Google Scholar] [CrossRef]
- Wei, S.C.; Yang-Yen, H.F.; Tsao, P.N.; Weng, M.T.; Tung, C.C.; Yu, L.C.H.; Lai, L.C.; Hsiao, J.H.; Chuang, E.Y.; Shun, C.T.; et al. SHANK3 Regulates Intestinal Barrier Function Through Modulating ZO-1 Expression Through the PKCε-dependent Pathway. Inflamm. Bowel. Dis. 2017, 23, 1730–1740. [Google Scholar] [CrossRef] [PubMed]
- Su, X.; Wei, J.; Qi, H.; Jin, M.; Zhang, Q.; Zhang, Y.; Zhang, C.; Yang, R. LRRC19 Promotes Permeability of the Gut Epithelial Barrier Through Degrading PKC-ζ and PKCι/λ to Reduce Expression of ZO1, ZO3, and Occludin. Inflamm. Bowel Dis. 2021, 27, 1302–1315. [Google Scholar] [CrossRef]
- Gu, A.; Yang, L.; Wang, J.; Li, J.; Shan, A. Protective effect of glutamine and alanyl-glutamine against zearalenone-induced intestinal epithelial barrier dysfunction in IPEC-J2 cells. Res. Vet. Sci. 2021, 137, 48–55. [Google Scholar] [CrossRef] [PubMed]
- Amin, A.; Tuenter, E.; Exarchou, V.; Upadhyay, A.; Cos, P.; Maes, L.; Apers, S.; Pieters, L. Phytochemical and Pharmacological Investigations on Nymphoides indica Leaf Extracts. Phytother. Res. 2016, 30, 1624–1633. [Google Scholar] [CrossRef]
- Khan, Z.R.; Chowdhury, N.S.; Sharmin, S.; Sohrab, M.H. Medicinal values of aquatic plant genus Nymphoides grown in Asia: A review. Asian Pac. J. Trop. Biomed. 2018, 8, 113–119. [Google Scholar]
- Hou, T.; Yang, M.; Yan, K.; Fan, X.; Ci, X.; Peng, L. Amentoflavone Ameliorates Carrageenan-Induced Pleurisy and Lung Injury by Inhibiting the NF-κB/STAT3 Pathways via Nrf2 Activation. Front. Pharmacol. 2022, 13, 763608. [Google Scholar] [CrossRef] [PubMed]
- Moon, S.M.; Lee, S.A.; Hong, J.H.; Kim, J.S.; Kim, D.K.; Kim, C.S. Corrigendum to “Oleamide suppresses inflammatory responses in LPS-induced RAW264.7 murine macrophages and alleviates paw edema in a carrageenan-induced inflammatory rat model” [Int. Immunopharmacol. 56 (2018) 179–185]. Int. Immunopharmacol. 2018, 60, 246. [Google Scholar] [CrossRef]
- Ali, Y.A.; Soliman, H.A.; Abdel-Gabbar, M.; Ahmed, N.A.; Attia, K.A.A.; Shalaby, F.M.; El-Nahass, E.S.; Ahmed, O.M. Rutin and Hesperidin Revoke the Hepatotoxicity Induced by Paclitaxel in Male Wistar Rats via Their Antioxidant, Anti-Inflammatory, and Antiapoptotic Activities. Evid. Based Complement Altern. Med. 2023, 2023, 2738351. [Google Scholar] [CrossRef]
- Li, X.; Li, K.; Xie, H.; Xie, Y.; Li, Y.; Zhao, X.; Jiang, X.; Chen, D. Antioxidant and Cytoprotective Effects of the Di-O-Caffeoylquinic Acid Family: The Mechanism, Structure-Activity Relationship, and Conformational Effect. Molecules 2018, 23, 222. [Google Scholar] [CrossRef]
- Chen, F.; Wang, Y.; Wang, K.; Chen, J.; Jin, K.; Peng, K.; Chen, X.; Liu, Z.; Ouyang, J.; Wang, Y.; et al. Effects of Litsea cubeba essential oil on growth performance, blood antioxidation, immune function, apparent digestibility of nutrients, and fecal microflora of pigs. Front. Pharmacol. 2023, 14. [Google Scholar] [CrossRef]
- Bai, M.; Liu, H.; Yan, Y.; Duan, S.; Szeto, I.M.-Y.; He, J.; Hu, J.; Fu, Y.; Xu, K.; Xiong, X. Hydrolyzed protein formula improves the nutritional tolerance by increasing intestinal development and altering cecal microbiota in low-birth-weight piglets. Front. Nutr. 2024, 11. [Google Scholar] [CrossRef] [PubMed]
- Janczyk, M.; Pfister, R. Unterschiedshypothesen Bei Bis zu Zwei Gruppen: T-Tests. In Inferenzstatistik Verstehen: Von A Wie Signifikanztest Bis Z Wie Konfidenzintervall; Janczyk, M., Pfister, R., Eds.; Springer: Berlin/Heidelberg, Germany, 2020; pp. 45–68. [Google Scholar]
- Lynch, S.M. Comparing Means Across Multiple Groups: Analysis of Variance (ANOVA). In Using Statistics in Social Research: A Concise Approach; Lynch, S.M., Ed.; Springer: New York, NY, USA, 2013; pp. 117–126. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene a | Sequence (5′–3′) | Accession No. |
|---|---|---|
| GAPDH | Forward: ACACCGAGCATCTCCTGACT Reverse: GACGAGGCAGGTCTCCCTAA | NM_001206359 |
| SLC1A5 | Forward: CTTGACTCTTGTCCCCATCC Reverse: CAACCCATCAGAACCTCCTT | XM_003127238.5 |
| SLC6A14 | Forward: TAACTTTGGGTTGCTGCTTG Reverse: CAATGAATCTGTTCCCTCCAT | NM_001348402.1 |
| SLC38A1 | Forward: CTGGGGGTGTTGTTTTCTCT Reverse: AAGGAAGTGATGAGGGGTTG | XM_003355629.4 |
| GLUL | Forward: ATGCGAGAGGAGAATGGTCT Reverse: TCGTTGATGTTGGAGGTTTC | NM_213909.1 |
| GLS | Forward: CTCTTGACCAGGGGTGTTCT Reverse: GGTGTTTGGGGCTGAATAGT | XM_021076049.1 |
| DDIT3 | Forward: TGAAAGCAGAGCCTAATCCA Reverse: CCAGGGGGTGAGACATAGTT | NM_001144845.1 |
| EIF2AK3 | Forward: AAAGGTCTCGGTTGCTGATT Reverse: AAAAGGCTGATGGGAATGAC | XM_003124925.4 |
| ERN1 | Forward: GTCTCTGCCCATCAACCTCT Reverse: ATCTTGTAGTCCCCGTCGTC | XM_005668695.3 |
| HSPA5 | Forward: AGGGAAGGGGAGAAGAACAT Reverse: GGTAGAACGGAAAAGGTCCA | XM_001927795.7 |
| IL-4 | Forward: TCCACGGACACAAGTGCGAC Reverse: TGTTTGCCATGCTGCTCAGG | NM_214123.1 |
| IL-8 | Forward: GGCTGTTGCCTTCTTGGCAG Reverse: TTTGGGGTGGAAAGGTGTGG | NM_213867 |
| IL-10 | Forward: CATCCACTTCCCAACCAGCC Reverse: CTCCCCATCACTCTCTGCCTTC | NM_214041 |
| IL-1β | Forward: GAACAAGAGCATCAGGCAGA Reverse: TGGCATCACAGACAAAGTCA | NM_001305893 |
| TNF-α | Forward: TTTCTGTGAAAACGGAGCTG Reverse: CAGCGATGTAGCGACAAGTT | NM_214022 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, C.H.; Shin, S.; Kim, T.H.; Lee, S.I. Nymphoides peltata Alleviates Patulin-Induced Glutamine Metabolic Stress and Epithelial Toxicity in Small Intestinal Epithelial Cells. Toxins 2025, 17, 337. https://doi.org/10.3390/toxins17070337
Lee CH, Shin S, Kim TH, Lee SI. Nymphoides peltata Alleviates Patulin-Induced Glutamine Metabolic Stress and Epithelial Toxicity in Small Intestinal Epithelial Cells. Toxins. 2025; 17(7):337. https://doi.org/10.3390/toxins17070337
Chicago/Turabian StyleLee, Chae Hyun, Sangsu Shin, Tae Hyun Kim, and Sang In Lee. 2025. "Nymphoides peltata Alleviates Patulin-Induced Glutamine Metabolic Stress and Epithelial Toxicity in Small Intestinal Epithelial Cells" Toxins 17, no. 7: 337. https://doi.org/10.3390/toxins17070337
APA StyleLee, C. H., Shin, S., Kim, T. H., & Lee, S. I. (2025). Nymphoides peltata Alleviates Patulin-Induced Glutamine Metabolic Stress and Epithelial Toxicity in Small Intestinal Epithelial Cells. Toxins, 17(7), 337. https://doi.org/10.3390/toxins17070337